Role of Candida albicans-Secreted Aspartyl Proteinases (Saps) in Severe Early Childhood Caries

Abstract
:1. Introduction
2. Results and Discussion
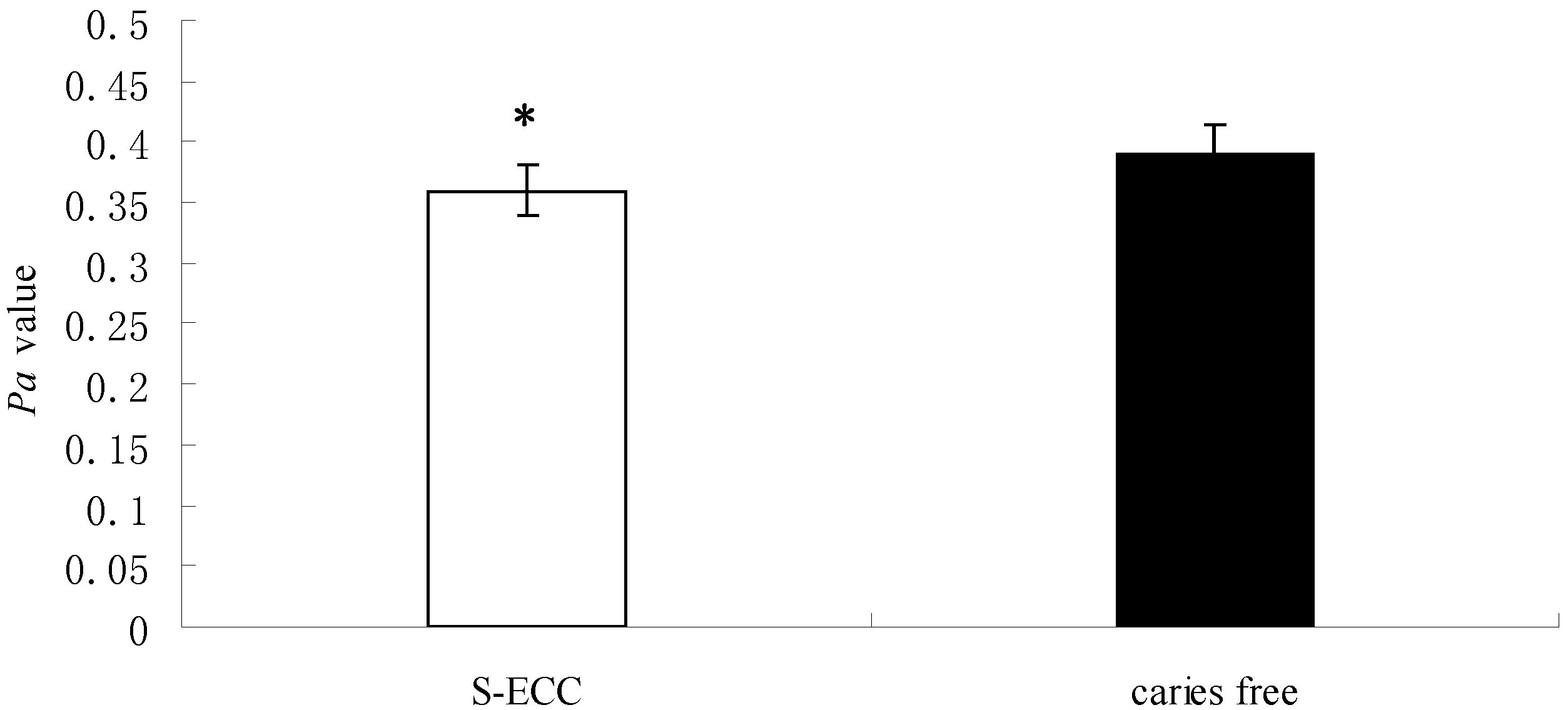
2.1. The Enzyme Activity of Saps Determined by the Yeast Nitrogen Base–Bovine Serum Albumi (YNB–BSA) Agar Plate Method

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | n | Diameter of Colony (mm) | Diameter of Colony plus Precipitation Zone (mm) | Pa | p |
|---|---|---|---|---|---|
| S-ECC | 26 | 7.13 ± 0.49 | 19.76 ± 2.63 | 0.36 ± 0.03 | 0.031 |
| Caries-free | 14 | 6.78 ± 0.32 | 17.52 ± 1.65 | 0.39 ± 0.05 |

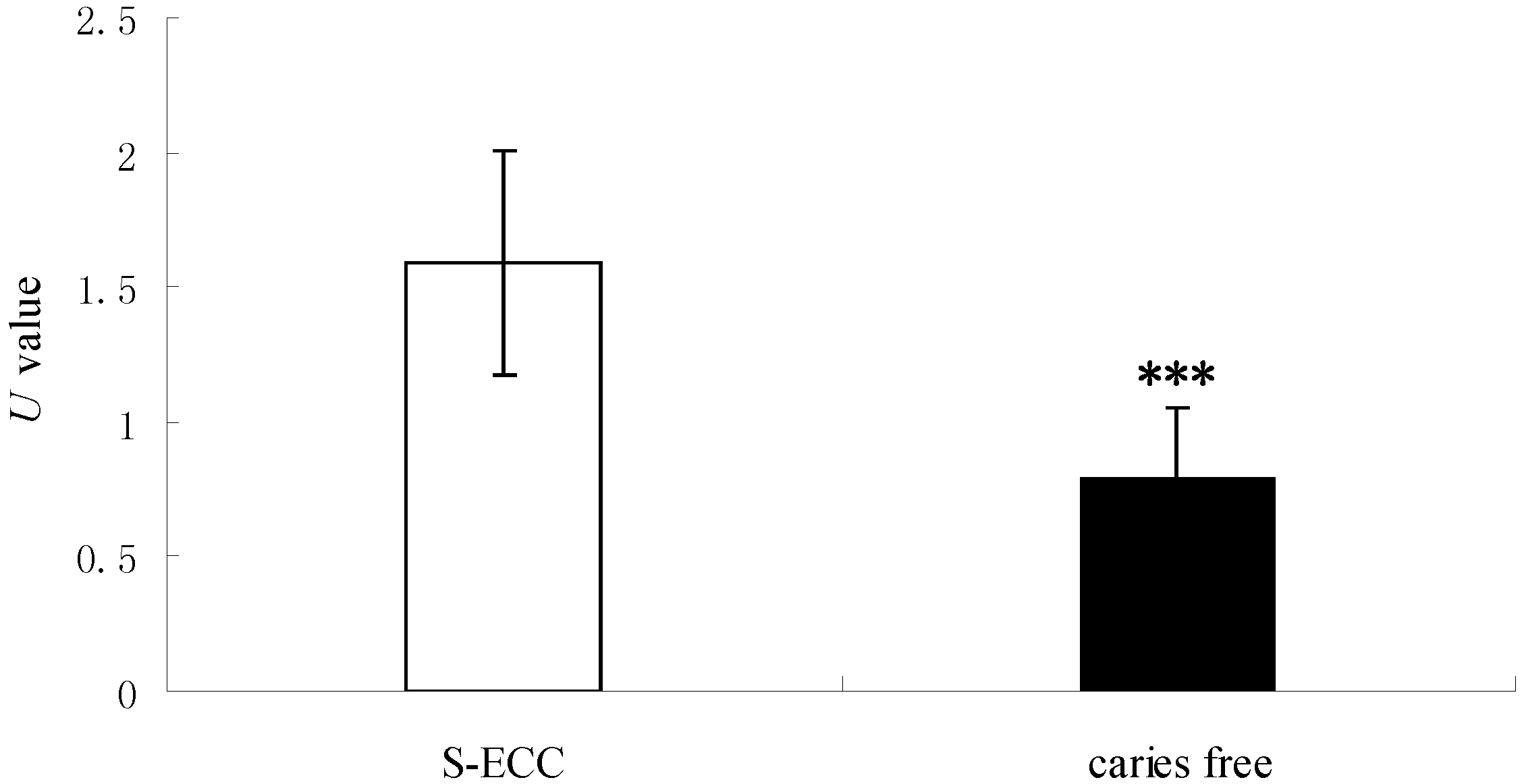
2.2. The Enzyme Activity of Saps Determined by the Bovine Serum Albumin–MTT (BSA–MTT) Method
| Groups | n | OD280 | MTT Value | U | p |
|---|---|---|---|---|---|
| S-ECC | 26 | 0.53 ± 0.25 | 0.40 ± 0.19 | 1.59 ± 0.92 | 0.001 |
| Caries-free | 14 | 0.42 ± 0.13 | 0.54 ± 0.12 | 0.79 ± 0.26 |
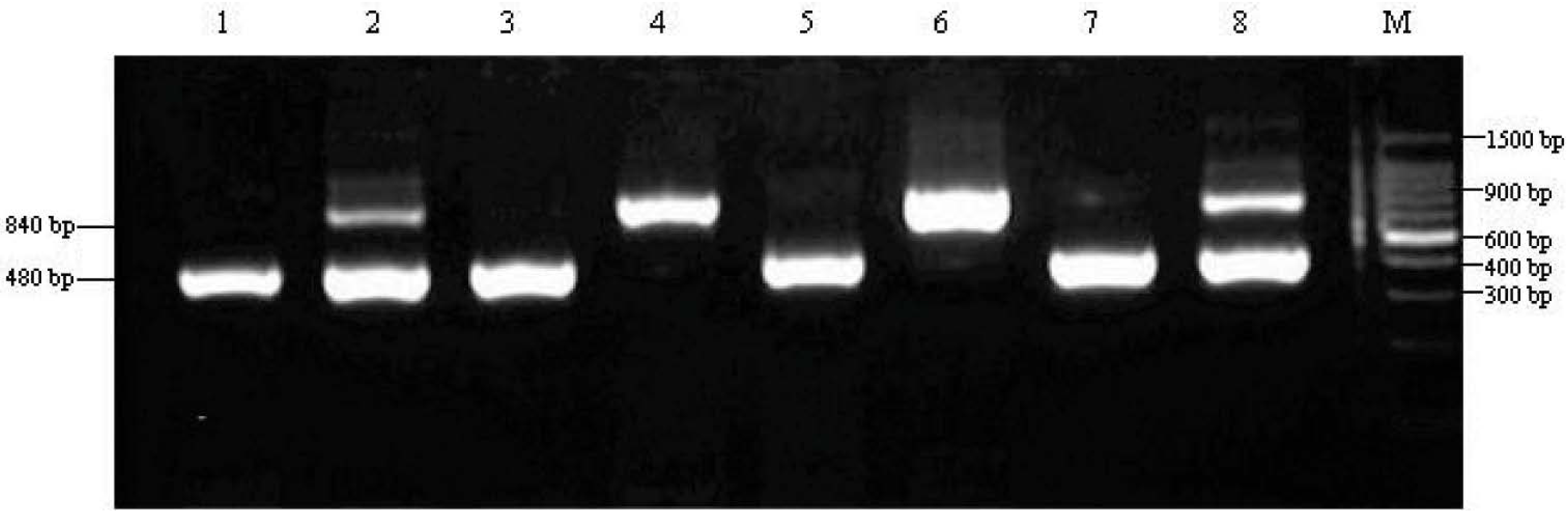
2.3. Genotypes of Candida albicans from Severe Early Childhood Caries (S-ECC) Children
| Groups | C. albicans Isolate | Genotypes | ||
|---|---|---|---|---|
| A | B | C | ||
| S-ECC group | 23 | 13 (56.5%) | 7 (30.4%) | 3 (13.1%) |
| CF group | 17 | 8 (47.1%) | ND | 9 (52.9%) |

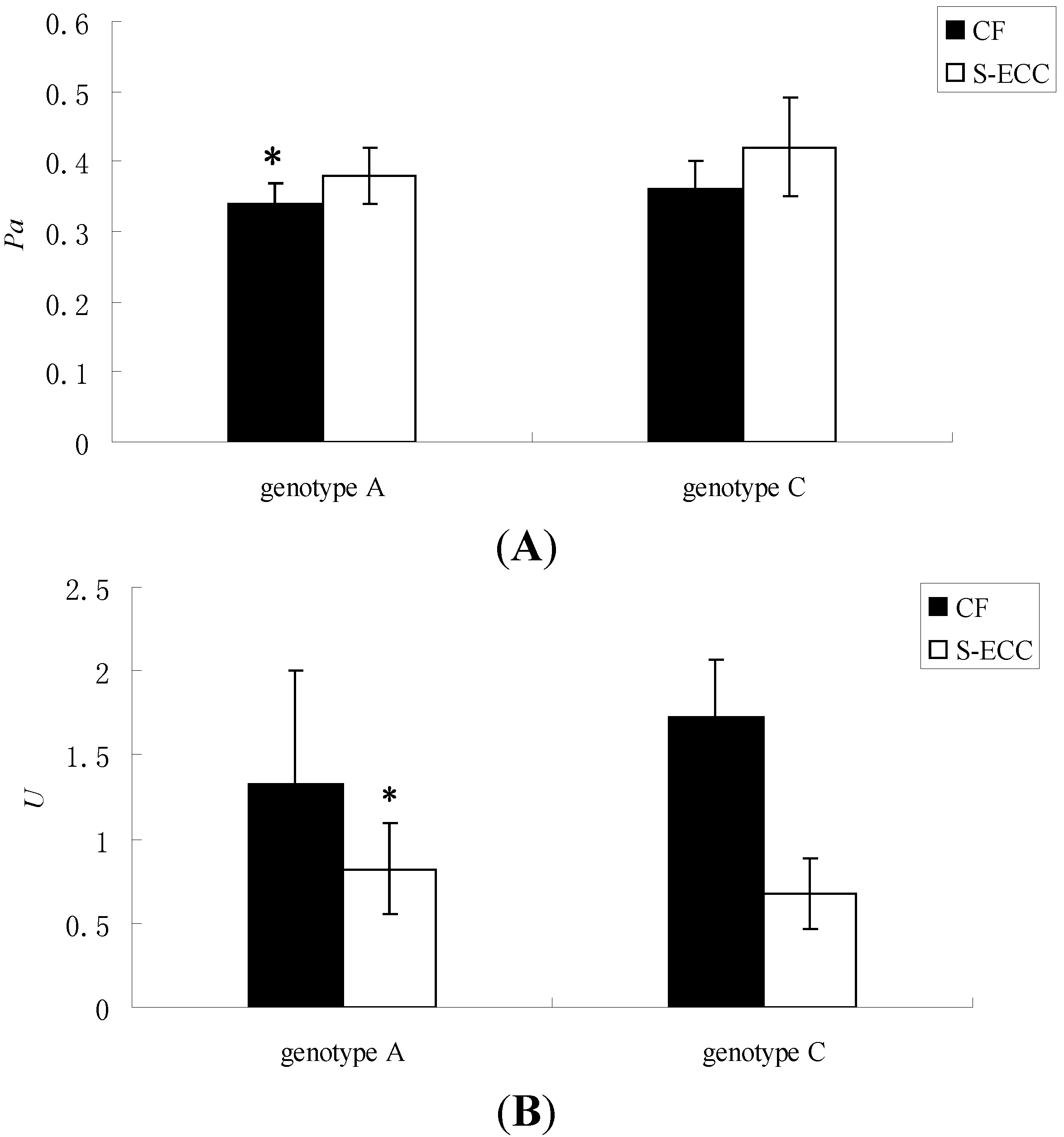
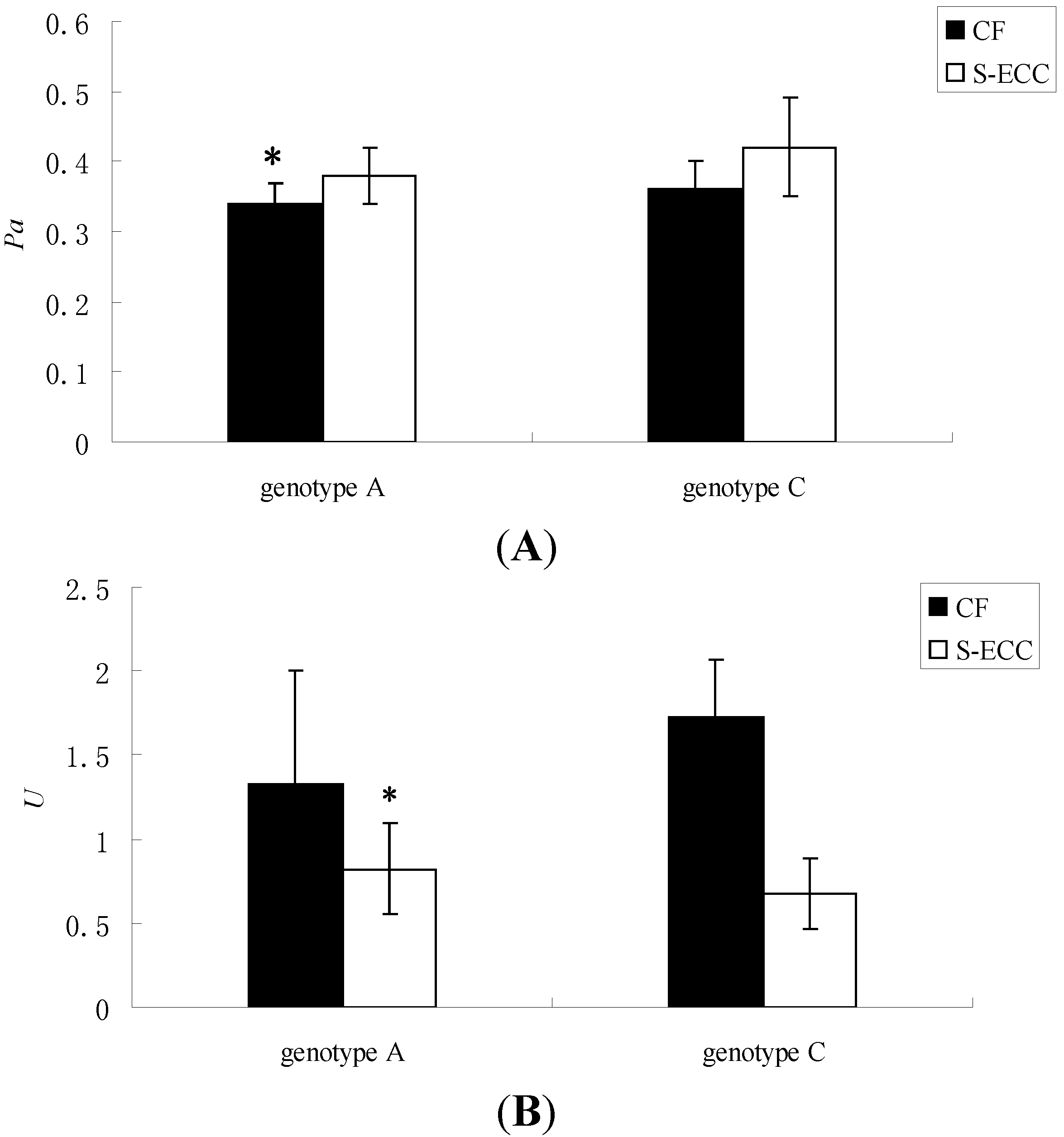
| Genotype | Enzyme Activity | Group | N | x ± s | F | t | Sig. |
|---|---|---|---|---|---|---|---|
| Genotype A | Pa | CF | 8 | 0.34 ± 0.03 | 0.426 | −2.385 | 0.028 |
| S-ECC | 13 | 0.38 ± 0.04 | |||||
| U | CF | 8 | 1.33 ± 0.67 | 8.687 | 2.296 | 0.034 | |
| S-ECC | 13 | 0.82 ± 0.27 | |||||
| Genotype C | Pa | CF | 9 | 0.36 ± 0.04 | 2.029 | −1.898 | 0.087 |
| S-ECC | 3 | 0.42 ± 0.07 | |||||
| U | CF | 9 | 1.73 ± 0.34 | 4.88 | 1.307 | 0.22 | |
| S-ECC | 3 | 0.68 ± 0.21 |
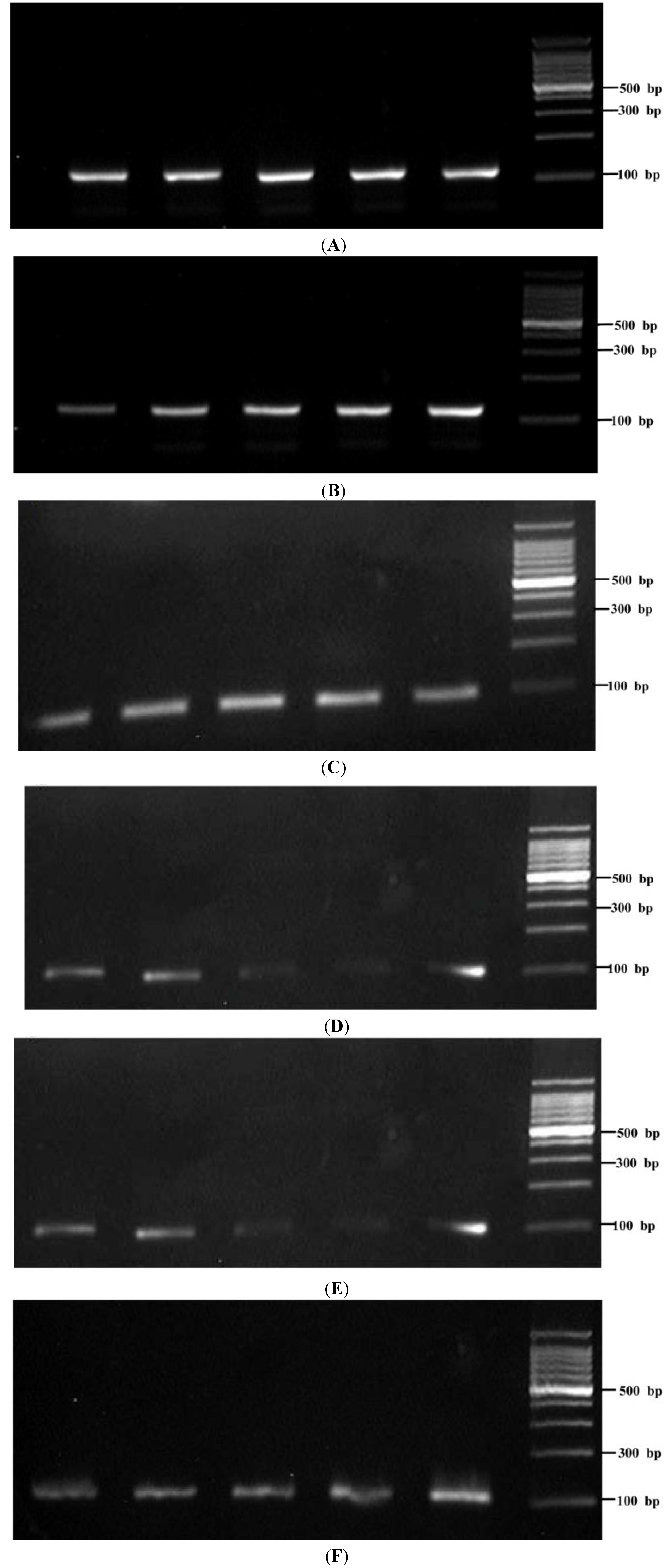
2.4. Gene Expression of Sap1–5 in Dental Biofilm from S-ECC and CF Children


3. Materials and Methods
3.1. Study Population
3.2. Sampling Procedures
3.3. Candida albicans Identification
3.4. Saps Enzyme Activity Determination by the Agar Plate Method
3.5. Saps Enzyme Activity Determination by MTT Methods
3.6. DNA Extraction of C. albicans
3.7. Genotype Determination of C. albicans by Polymerase Chain Reaction (PCR)
3.8. RNA Extraction and Reverse Transcription
3.9. Real-Time RT-PCR for Gene Expression of Sap1–5
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- American Academy on Pediatric Dentistry; American Academy of Pediatrics. Policy on early childhood caries (ECC): Classifications, consequences, and preventive strategies. Pediatr. Dent. 2008, 30, 40–43. [Google Scholar]
- Berkowitz, R.J. Causes, treatment and prevention of early childhood caries: A microbiologic perspective. J. Can. Dent. Assoc. 2003, 69, 304–307. [Google Scholar]
- Ge, Y.; Caufield, P.W.; Fisch, G.S.; Li, Y. Streptococcus mutans and Streptococcus sanguinis colonization correlated with caries experience in children. Caries Res. 2008, 42, 444–448. [Google Scholar] [CrossRef]
- Marchant, S.; Brailsford, S.R.; Twomey, A.C.; Roberts, G.J.; Beighton, D. The predominant microflora of nursing caries lesions. Caries Res. 2001, 35, 397–406. [Google Scholar] [CrossRef]
- Gross, E.L.; Leys, E.J.; Gasparovich, S.R.; Firestone, N.D.; Schwartzbaum, J.A.; Janies, D.A.; Asnani, K.; Griffen, A.L. Bacterial 16S sequence analysis of severe caries in young permanent teeth. J. Clin. Microbiol. 2010, 48, 4121–4128. [Google Scholar]
- Tanner, A.C.; Mathney, J.M.; Kent, R.L.; Chalmers, N.I.; Hughes, C.V.; Loo, C.Y.; Pradhan, N.; Kanasi, E.; Hwang, J.; Dahlan, M.A. Cultivable anaerobic microbiota of severe early childhood caries. J. Clin. Microbiol. 2011, 49, 1464–1474. [Google Scholar] [CrossRef]
- Akdeniz, B.G.; Koparal, E.; Sen, B.H.; Ates, M.; Denizci, A.A. Prevalence of Candida albicans in oral cavities and root canals of children. ASDC J. Dent. Child. 2002, 69, 289–292. [Google Scholar]
- De Carvalho, F.G.; Silva, D.S.; Hebling, J.; Spolidorio, L.C.; Spolidorio, D.M. Presence of mutans Streptococci and Candida spp. in dental plaque/dentine of carious teeth and early childhood caries. Arch. Oral Biol. 2006, 51, 1024–1028. [Google Scholar] [CrossRef]
- Rozkiewicz, D.; Daniluk, T.; Zaremba, M.L.; Cylwik-Rokicka, D.; Stokowska, W.; Pawinska, M.; Dabrowska, E.; Marczuk-Kolada, G.; Waszkiel, D. Oral Candida albicans carriage in healthy preschool and school children. Adv. Med. Sci. 2006, 51, 187–190. [Google Scholar]
- Monod, M.; Hube, B.; Hess, D.; Sanglard, D. Differential regulation of Sap8 and Sap9, which encode two new members of the secreted aspartic proteinase family in Candida albicans. Microbiology 1998, 144, 2731–2737. [Google Scholar] [CrossRef]
- Monod, M.; Togni, G.; Hube, B.; Sanglard, D. Multiplicity of genes encoding secreted aspartic proteinases in Candida species. Mol. Microbiol. 1994, 13, 357–368. [Google Scholar] [CrossRef]
- Naglik, J.R.; Newport, G.; White, T.C.; Fernandes-Naglik, L.L.; Greenspan, J.S.; Greenspan, D.; Sweet, S.P.; Challacombe, S.J.; Agabian, N. In vivo analysis of secreted aspartyl proteinase expression in human oral candidiasis. Infect. Immun. 1999, 67, 2482–2490. [Google Scholar]
- Schaller, M.; Schafer, W.; Korting, H.C.; Hube, B. Differential expression of secreted aspartyl proteinases in a model of human oral candidosis and in patient samples from the oral cavity. Mol. Microbiol. 1998, 29, 605–615. [Google Scholar] [CrossRef]
- Naglik, J.R.; Rodgers, C.A.; Shirlaw, P.J.; Dobbie, J.L.; Fernandes-Naglik, L.L.; Greenspan, D.; Agaban, N.; Challacomber, S.J. Differential expression of Candida albicans secreted aspartyl proteinase and phospholipase B genes in humans correlates with active oral and vaginal infections. J. Infect. Dis. 2003, 188, 469–479. [Google Scholar] [CrossRef]
- Schaller, M.; Bein, M.; Korting, H.C.; Baur, S.; Hamm, G.; Monod, M.; Beinhauer, S.; Hube, B. The secreted aspartyl proteinases Sap1 and Sap2 cause tissue damage in an in vitro model of vaginal candidiasis based on reconstituted human vaginal epithelium. Infect. Immun. 2003, 71, 3227–3234. [Google Scholar] [CrossRef]
- Staib, P.; Kretschmar, M.; Nichterlein, T.; Hof, H.; Morschhauser, J. Differential activation of a Candida albicans virulence gene family during infection. Proc. Natl. Acad. Sci. USA 2000, 97, 6102–6107. [Google Scholar]
- Kretschmar, M.; Felk, A.; Staib, P.; Schaller, M.; Hess, D.; Callapina, M.; Morschhauser, J.; Schafer, W.; Korting, H.C.; Hof, H.; et al. Individual acid aspartic proteinases (Saps) 1–6 of Candida albicans are not essential for invasion and colonization of the gastrointestinal tract in mice. Microb. Pathog. 2002, 32, 61–70. [Google Scholar] [CrossRef]
- Price, M.F.; Wilkinson, I.D.; Gentry, L.O. Plate method for detection of phospholipase activity in Candida albicans. Sabouraudia 1982, 20, 7–14. [Google Scholar] [CrossRef]
- Yang, X.Q.; Zhang, Q.; Lu, L.Y.; Yang, R.; Liu, Y.; Zou, J. Genotypic distribution of Candida albicans in dental biofilm of chinese children associated with severe early childhood caries. Arch. Oral Biol. 2012, 57, 1048–1053. [Google Scholar] [CrossRef]
- Al-Karaawi, Z.M.; Manfredi, M.; Waugh, A.C.; McCullough, M.J.; Jorge, J.; Scully, C.; Porter, S.R. Molecular characterization of Candida spp. isolated from the oral cavities of patients from diverse clinical settings. Oral Microbiol. Immunol. 2002, 17, 44–49. [Google Scholar] [CrossRef]
- Hube, B.; Monod, M.; Schofield, D.A.; Brown, A.J.; Gow, N.A. Expression of seven members of the gene family encoding secretory aspartyl proteinases in Candida albicans. Mol. Microbiol. 1994, 14, 87–99. [Google Scholar] [CrossRef]
- White, T.C.; Agabian, N. Candida albicans secreted aspartyl proteinases: Isoenzyme pattern is determined by cell type, and levels are determined by environmental factors. J. Bacteriol. 1995, 177, 5215–5221. [Google Scholar]
- Ripeau, J.S.; Fiorillo, M.; Aumont, F.; Belhumeur, P.; de Repentigny, L. Evidence for differential expression of Candida albicans virulence genes during oral infection in intact and human immunodeficiency virus Type 1-Transgenic mice. J. Infect. Dis. 2002, 185, 1094–1102. [Google Scholar] [CrossRef]
- Schaller, M.; Schackert, C.; Korting, H.C.; Januschke, E.; Hube, B. Invasion of Candida albicans correlates with expression of secreted aspartic proteinases during experimental infection of human epidermis. J. Investig. Dermatol. 2000, 114, 712–717. [Google Scholar] [CrossRef]
- Klinke, H.T.; Pönisch, R.; Kriegel, T.M.; Klimm, H.W. Immunohistochemical detection of the collagenolytic Candia albicans Sap2 proteinase in caries lesions. Caries Res. 2007, 41, 287. [Google Scholar]
- Borg-von, Z.M.; Beggah, S.; Boggian, K.; Sanglard, D.; Monod, M. The expression of the secreted aspartic proteinases Sap4 to Sap6 from Candida albicans in murine macrophages. Mol. Microb. 1998, 28, 543–554. [Google Scholar] [CrossRef]
- Caufield, P.W.; Saxena, D.; Fitch, D.; Li, Y. Population structure of lasmid-containing strains of Streptococcus mutans, a member of the human indigenous biota. J. Bacteriol. 2007, 189, 1238–1243. [Google Scholar] [CrossRef]
- Li, Y.; Ge, Y.; Saxena, D.; Caufield, P.W. Genetic profiling of the oral microbiota associated with severe early childhood caries. J. Clin. Microbiol. 2007, 45, 81–87. [Google Scholar] [CrossRef]
- Odds, F.C.; Bernaerts, R. CHROMagar Candida, a new differential isolation medium for presumptive identification of clinically important candida species. J. Clin. Microbiol. 1994, 32, 1923–1929. [Google Scholar]
- Barros, L.M.; Boriollo, M.F.; Alves, A.C.; Klein, M.I.; Gonçalves, R.B.; Hofling, J.F. Genetic diversity and exoenzyme activities of Candida albicans and Candida dubliniensis isolated from the oral cavity of brazilian periodontal patients. Arch. Oral Biol. 2008, 53, 1172–1178. [Google Scholar] [CrossRef]
- McCullough, M.J.; Clemons, K.V.; Stevens, D.A. Molecular and phenotypic characterization of genotypic Candida albicans subgroups and comparison with Candida dubliniensis and Candida stellatoidea. J. Clin. Microbiol. 1999, 37, 417–421. [Google Scholar]
- She, X.D.; Wang, X.J.; Fu, M.H.; Shen, Y.N.; Liu, W.D. Genotype comparisons of strains of Candida albicans from patients with cutaneous candidiasis and vaginal candidiasis. Chin. Med. J. 2008, 121, 1450–1455. [Google Scholar]
- Tamura, M.; Watanabe, K.; Mikami, Y.; Yazawa, K.; Nishimura, K. Molecular characterization of new clinical isolates of Candida albicans and C. dubliniensis in Japan: Analysis reveals a new genotype of C. albicans with Group I Intron. J. Clin. Microbiol. 2001, 39, 4309–4315. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, W.; Yu, D.; Gao, S.; Lin, J.; Chen, Z.; Zhao, W. Role of Candida albicans-Secreted Aspartyl Proteinases (Saps) in Severe Early Childhood Caries. Int. J. Mol. Sci. 2014, 15, 10766-10779. https://doi.org/10.3390/ijms150610766
Li W, Yu D, Gao S, Lin J, Chen Z, Zhao W. Role of Candida albicans-Secreted Aspartyl Proteinases (Saps) in Severe Early Childhood Caries. International Journal of Molecular Sciences. 2014; 15(6):10766-10779. https://doi.org/10.3390/ijms150610766
Chicago/Turabian StyleLi, Wenqing, Dongsheng Yu, Shuo Gao, Jiacheng Lin, Zhuoyu Chen, and Wei Zhao. 2014. "Role of Candida albicans-Secreted Aspartyl Proteinases (Saps) in Severe Early Childhood Caries" International Journal of Molecular Sciences 15, no. 6: 10766-10779. https://doi.org/10.3390/ijms150610766