Up-Regulation of Rhoa/Rho Kinase Pathway by Translationally Controlled Tumor Protein in Vascular Smooth Muscle Cells

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
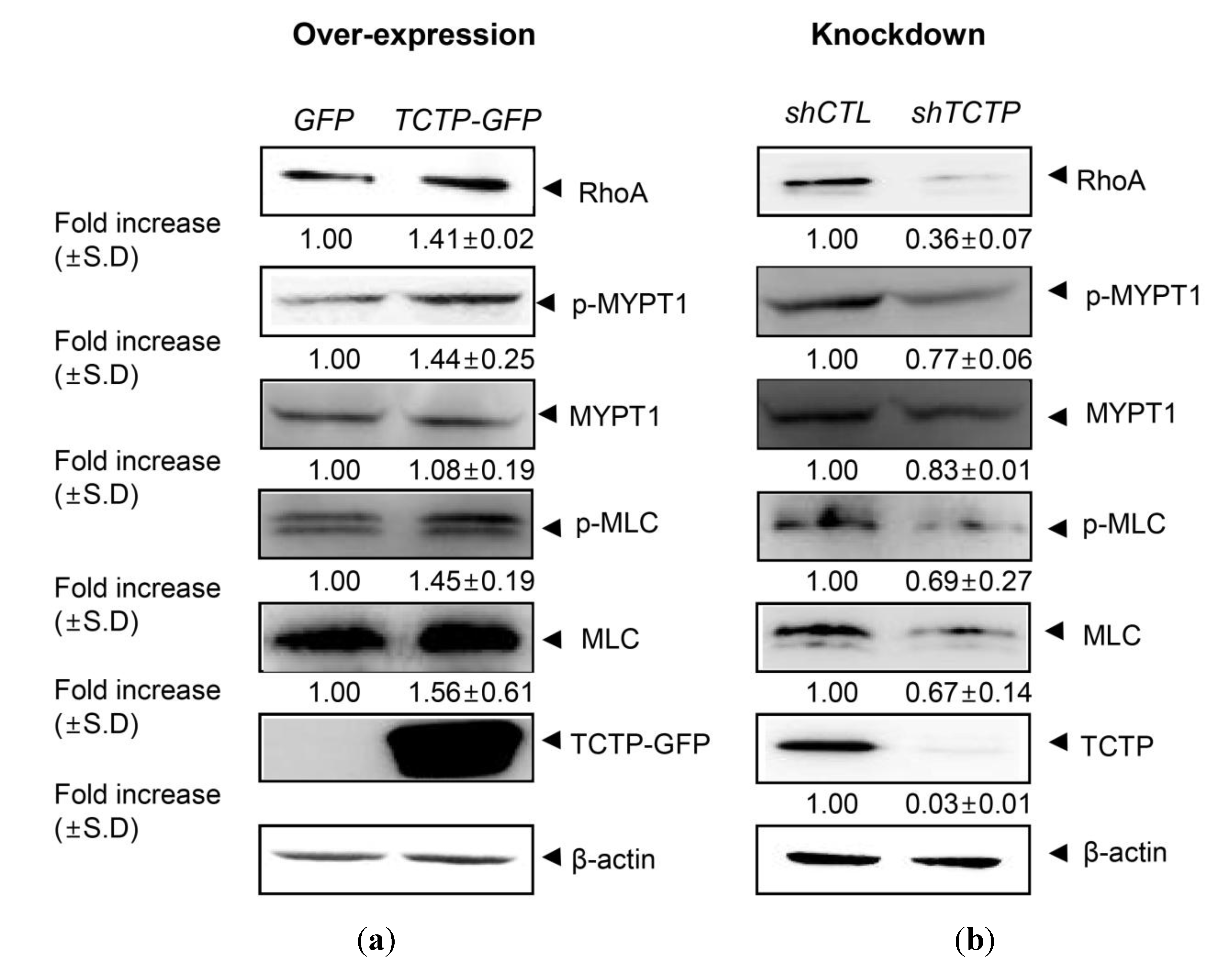
2.1. Translationally Controlled Tumor Protein (TCTP) Regulates the RhoA/Rho Kinase Pathway in Vascular Smooth Muscle Cells (VSMCs)
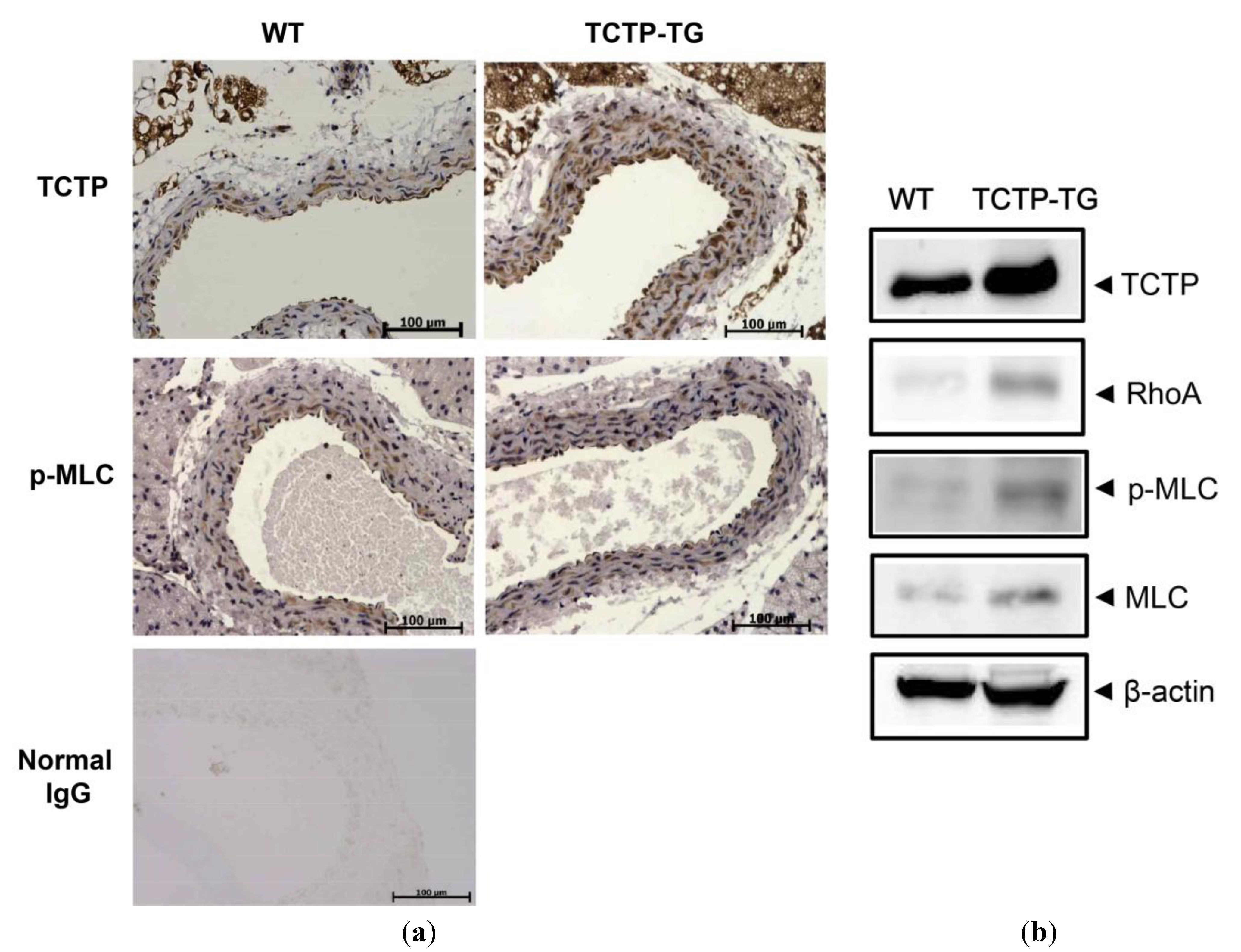
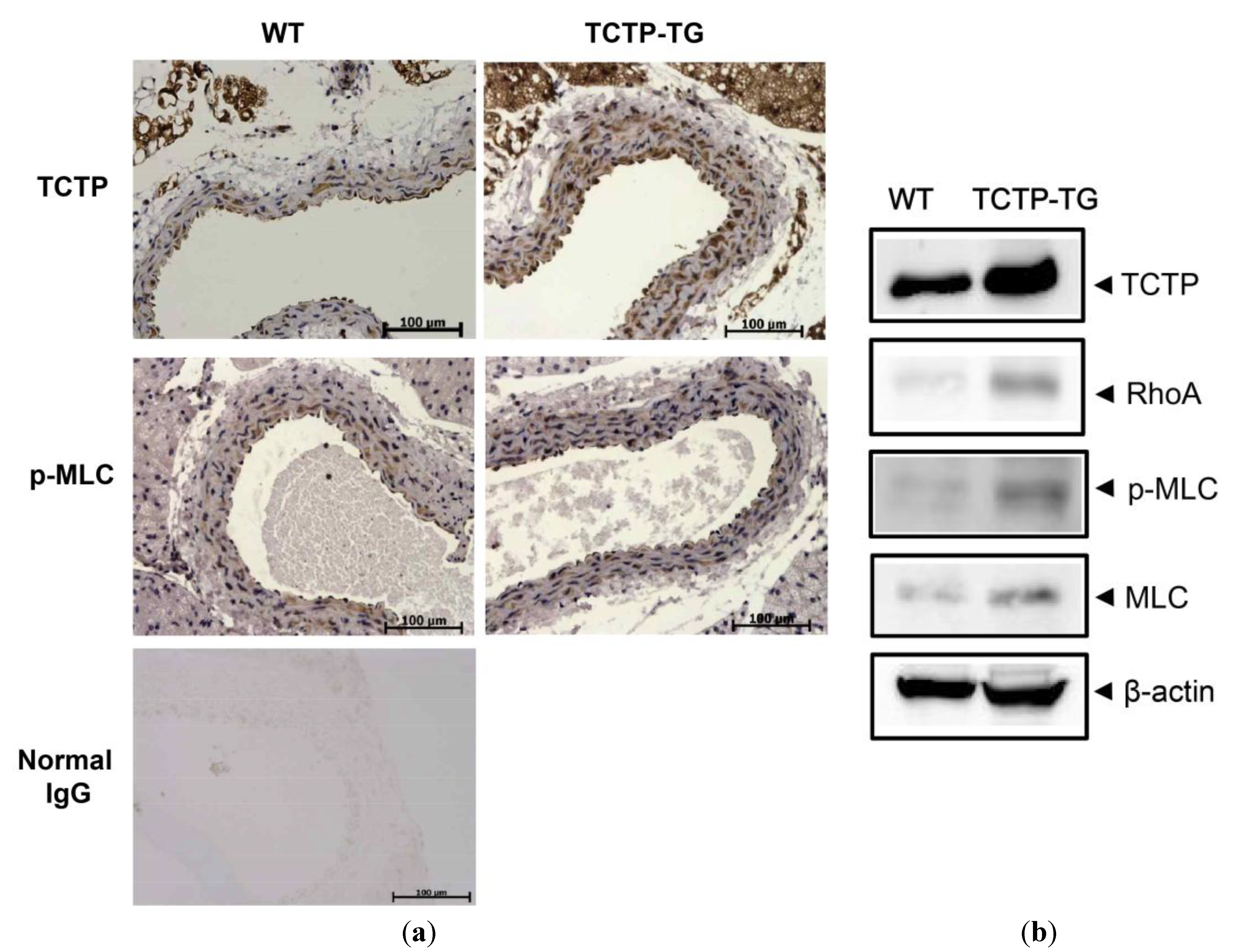
2.2. Up-Regulation of RhoA Expression and Phosphorylated Myosin Light Chain (p-MLC) in TCTP-Over-Expressing Transgenic (TCTP-TG) Mice

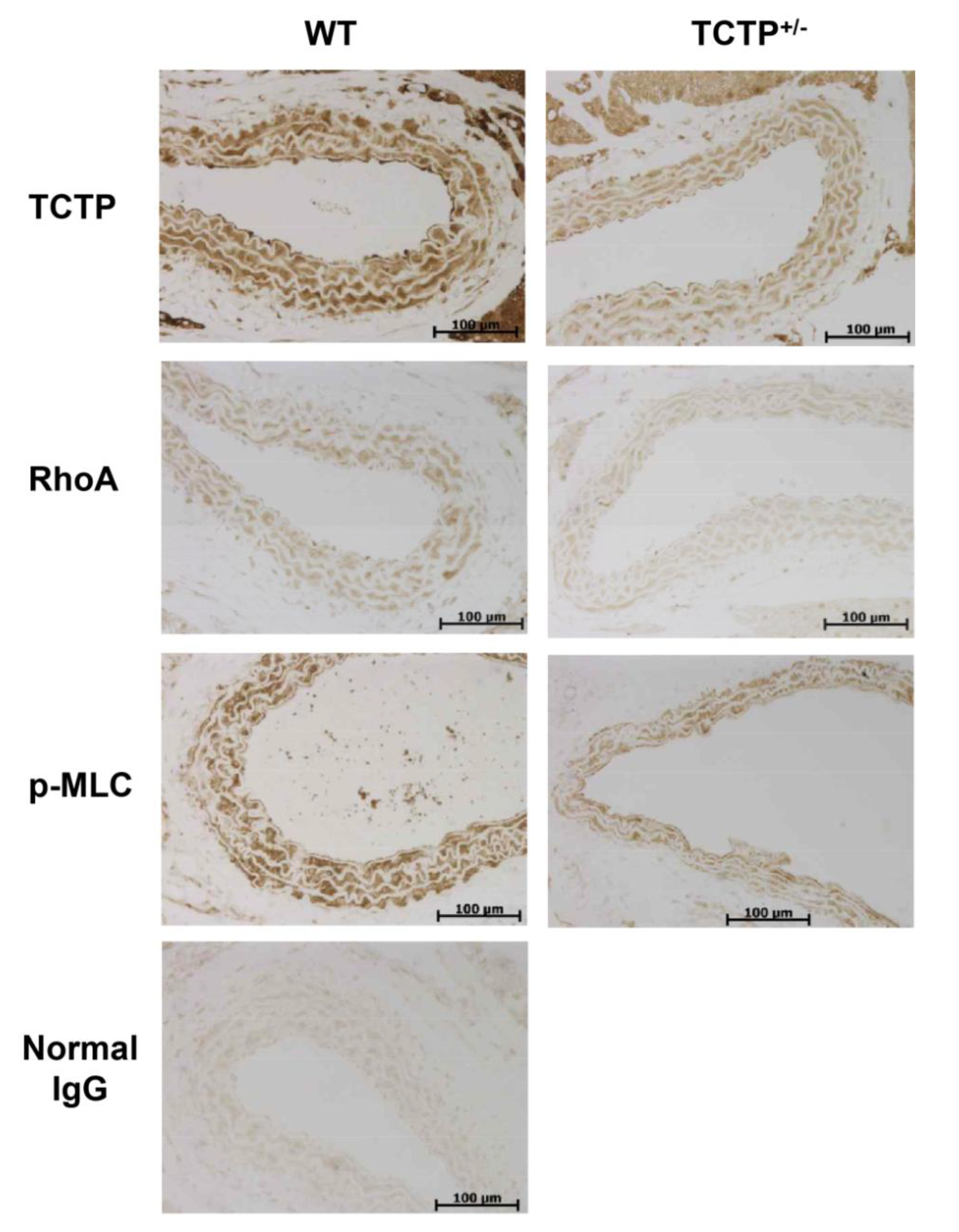
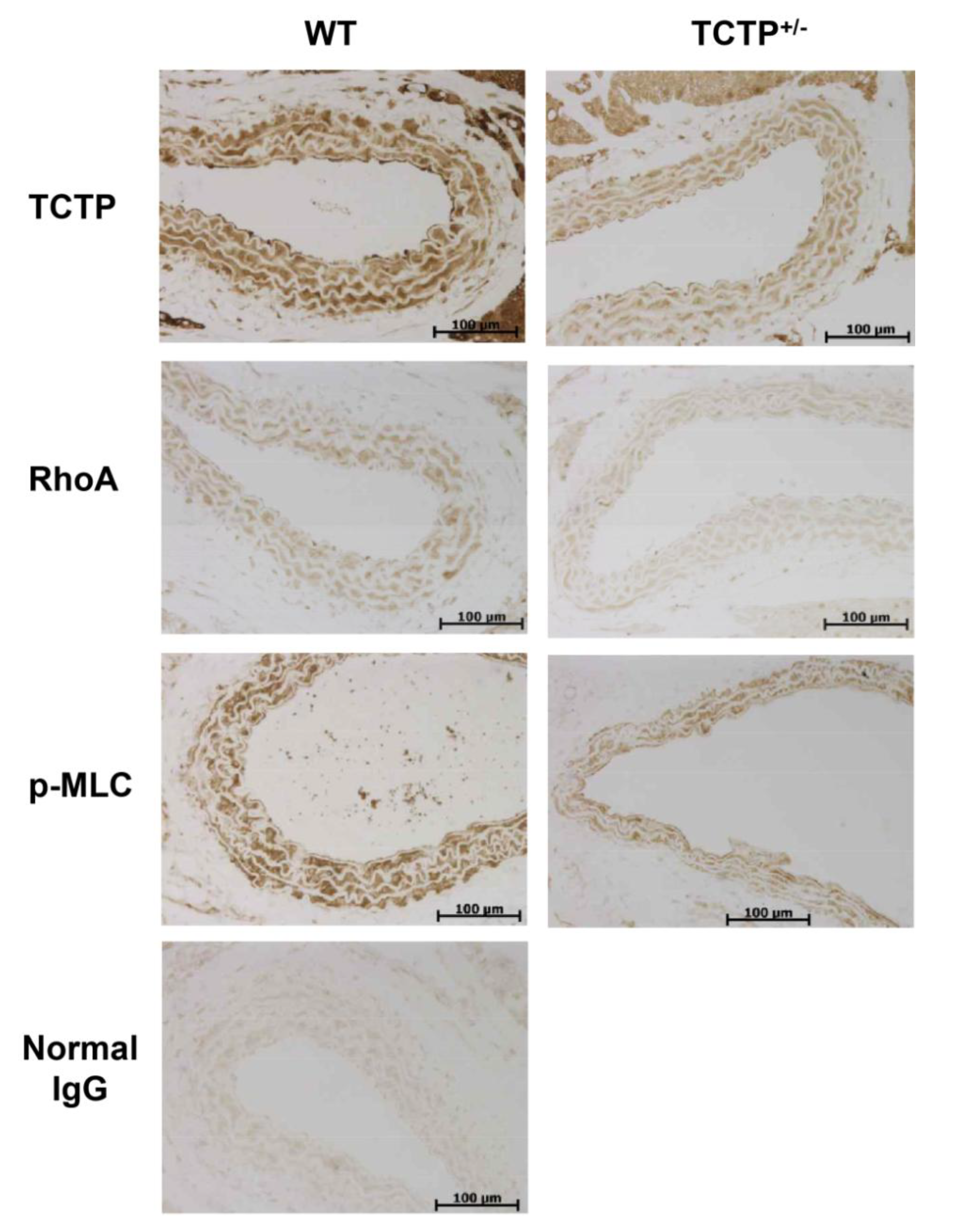
2.3. Down-Regulation of RhoA Expression and p-MLC in TCTP+/− Mice
3. Discussion
4. Experimental Section
4.1. Animal Studies
4.2. Cell Culture
4.3. Isolation of Vascular Smooth Muscle Cell (VSMC) from Rat Aorta
4.4. Virus Induction of TCTP Over-Expression
4.5. Western Blotting
4.6. Tissue Western Blotting
4.7. Immunohistochemistry
4.8. Assay of Rho Kinase Activity in VSMC
4.9. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bommer, U.A.; Thiele, B.J. The translationally controlled tumour protein (TCTP). Int. J. Biochem. Cell Biol. 2004, 36, 379–385. [Google Scholar] [CrossRef]
- MacDonald, S.M.; Rafnar, T.; Langdon, J.; Lichtenstein, L.M. Molecular identification of an IgE-dependent histamine-releasing factor. Science 1995, 269, 688–690. [Google Scholar]
- Gachet, Y.; Tournier, S.; Lee, M.; Lazaris-Karatzas, A.; Poulton, T.; Bommer, U.A. The growth-related, translationally controlled protein P23 has properties of a tubulin binding protein and associates transiently with microtubules during the cell cycle. J. Cell Sci. 1999, 112, 1257–1271. [Google Scholar]
- Li, F.; Zhang, D.; Fujise, K. Characterization of fortilin, a novel antiapoptotic protein. J. Biol. Chem. 2001, 276, 47542–47549. [Google Scholar] [CrossRef]
- Cans, C.; Passer, B.J.; Shalak, V.; Nancy-Portebois, V.; Crible, V.; Amzallag, N.; Allanic, D.; Tufino, R.; Argentini, M.; Moras, D.; et al. Translationally controlled tumor protein acts as a guanine nucleotide dissociation inhibitor on the translation elongation factor eEF1A. Proc. Natl. Acad. Sci. USA 2003, 100, 13892–13897. [Google Scholar] [CrossRef]
- Tuynder, M.; Fiucci, G.; Prieur, S.; Lespagnol, A.; Geant, A.; Beaucourt, S.; Duflaut, D.; Besse, S.; Susini, L.; Cavarelli, J.; et al. Translationally controlled tumor protein is a target of tumor reversion. Proc. Natl. Acad. Sci. USA 2004, 101, 15364–15369. [Google Scholar] [CrossRef]
- Kim, M.; Min, H.J.; Won, H.Y.; Park, H.; Lee, J.C.; Park, H.W.; Chung, J.; Hwang, E.S.; Lee, K. Dimerization of translationally controlled tumor protein is essential for its cytokine-like activity. PLoS One 2009, 4, e6464. [Google Scholar]
- Kim, M.J.; Kwon, J.S.; Suh, S.H.; Suh, J.K.; Jung, J.; Lee, S.N.; Kim, Y.H.; Cho, M.C.; Oh, G.T.; Lee, K. Transgenic over-expression of translationally controlled tumor protein induces systemic hypertension via repression of Na+,K+-ATPase. J. Mol. Cell. Cardiol. 2008, 44, 151–159. [Google Scholar] [CrossRef]
- Jung, J.; Kim, M.; Kim, M.J.; Kim, J.; Moon, J.; Lim, J.S.; Lee, K. Translationally controlled tumor protein interacts with the third cytoplasmic domain of Na,K-ATPase alpha subunit and inhibits the pump activity in HeLa cells. J. Biol. Chem. 2004, 279, 49868–49875. [Google Scholar]
- Sweeney, G.; Klip, A. Regulation of the Na+/K+-ATPase by insulin: Why and how? Mol. Cell Biochem. 1998, 182, 121–133. [Google Scholar] [CrossRef]
- Xie, Z.; Askari, A. Na+/K+-ATPase as a signal transducer. Eur. J. Biochem. 2002, 269, 2434–2439. [Google Scholar] [CrossRef]
- Manunta, P.; Rogowski, A.C.; Hamilton, B.P.; Hamlyn, J.M. Ouabain-induced hypertension in the rat: Relationships among plasma and tissue ouabain and blood pressure. J. Hypertens 1994, 12, 549–560. [Google Scholar]
- Cho, Y.; Maeng, J.; Ryu, J.; Shin, H.; Kim, M.; Oh, G.T.; Lee, M.Y.; Lee, K. Hypertension resulting from over-expression of translationally controlled tumor protein increases the severity of atherosclerosis in apolipoprotein E knock-out mice. Transgenic Res. 2012, 21, 1245–1254. [Google Scholar] [CrossRef]
- Hollander, W. Role of hypertension in atherosclerosis and cardiovascular disease. Am. J. Cardiol. 1976, 38, 786–800. [Google Scholar] [CrossRef]
- Pauletto, P.; Rattazzi, M. Inflammation and hypertension: The search for a link. Nephrol. Dial. Transplant. 2006, 21, 850–853. [Google Scholar] [CrossRef]
- Uehata, M.; Ishizaki, T.; Satoh, H.; Ono, T.; Kawahara, T.; Morishita, T.; Tamakawa, H.; Yamagami, K.; Inui, J.; Maekawa, M.; et al. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension. Nature 1997, 389, 990–994. [Google Scholar] [CrossRef]
- Hartshorne, D.J.; Ito, M.; Erdodi, F. Myosin light chain phosphatase: Subunit composition, interactions and regulation. J. Muscle Res. Cell Motil. 1998, 19, 325–341. [Google Scholar] [CrossRef]
- Kamm, K.E.; Stull, J.T. The function of myosin and myosin light chain kinase phosphorylation in smooth muscle. Annu. Rev. Pharmacol. Toxicol. 1985, 25, 593–620. [Google Scholar] [CrossRef]
- Sellers, J.R.; Spudich, J.A.; Sheetz, M.P. Light chain phosphorylation regulates the movement of smooth muscle myosin on actin filaments. J. Cell Biol. 1985, 101, 1897–1902. [Google Scholar] [CrossRef]
- Wirth, A. Rho kinase and hypertension. Biochim. Biophys. Acta 2010, 1802, 1276–1284. [Google Scholar] [CrossRef]
- Lee, D.L.; Webb, R.C.; Jin, L. Hypertension and RhoA/Rho-kinase signaling in the vasculature: Highlights from the recent literature. Hypertension 2004, 44, 796–799. [Google Scholar] [CrossRef]
- Masumoto, A.; Hirooka, Y.; Shimokawa, H.; Hironaga, K.; Setoguchi, S.; Takeshita, A. Possible involvement of Rho-kinase in the pathogenesis of hypertension in humans. Hypertension 2001, 38, 1307–1310. [Google Scholar] [CrossRef]
- Mita, M.; Yanagihara, H.; Hishinuma, S.; Saito, M.; Walsh, M.P. Membrane depolarization-induced contraction of rat caudal arterial smooth muscle involves Rho-associated kinase. Biochem. J. 2002, 364, 431–440. [Google Scholar] [CrossRef]
- Chen, S.H.; Wu, P.S.; Chou, C.H.; Yan, Y.T.; Liu, H.; Weng, S.Y.; Yang-Yen, H.F. A knockout mouse approach reveals that TCTP functions as an essential factor for cell proliferation and survival in a tissue- or cell type-specific manner. Mol. Biol. Cell 2007, 18, 2525–2532. [Google Scholar] [CrossRef]
- Miao, L.; Calvert, J.W.; Tang, J.; Parent, A.D.; Zhang, J.H. Age-related RhoA expression in blood vessels of rats. Mech. Ageing Dev. 2001, 122, 1757–1770. [Google Scholar] [CrossRef]
- Seko, T.; Ito, M.; Kureishi, Y.; Okamoto, R.; Moriki, N.; Onishi, K.; Isaka, N.; Hartshorne, D.J.; Nakano, T. Activation of RhoA and inhibition of myosin phosphatase as important components in hypertension in vascular smooth muscle. Circ. Res. 2003, 92, 411–418. [Google Scholar] [CrossRef]
- Raja, S.G. Evaluation of clinical efficacy of fasudil for the treatment of pulmonary arterial hypertension. Recent Pat. Cardiovasc. Drug Discov. 2012, 7, 100–104. [Google Scholar] [CrossRef]
- Satoh, K.; Fukumoto, Y.; Shimokawa, H. Rho-kinase: Important new therapeutic target in cardiovascular diseases. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H287–H296. [Google Scholar] [CrossRef]
- Ratz, P.H.; Berg, K.M.; Urban, N.H.; Miner, A.S. Regulation of smooth muscle calcium sensitivity: KCl as a calcium-sensitizing stimulus. Am. J. Physiol. Cell Physiol. 2005, 288, C769–C783. [Google Scholar]
- Kloc, M.; Tejpal, N.; Sidhu, J.; Ganachari, M.; Flores-Villanueva, P.; Jennings, N.B.; Sood, A.K.; Kubiak, J.Z.; Ghobrial, R.M. Inverse relationship between TCTP/RhoA and p53/cyclin A/actin expression in ovarian cancer cells. Folia Histochem. Cytobiol. 2012, 50, 358–367. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maeng, J.; Sheverdin, V.; Shin, H.; Ha, I.; Bae, S.S.; Yang-Yen, H.-F.; Lee, K. Up-Regulation of Rhoa/Rho Kinase Pathway by Translationally Controlled Tumor Protein in Vascular Smooth Muscle Cells. Int. J. Mol. Sci. 2014, 15, 10365-10376. https://doi.org/10.3390/ijms150610365
Maeng J, Sheverdin V, Shin H, Ha I, Bae SS, Yang-Yen H-F, Lee K. Up-Regulation of Rhoa/Rho Kinase Pathway by Translationally Controlled Tumor Protein in Vascular Smooth Muscle Cells. International Journal of Molecular Sciences. 2014; 15(6):10365-10376. https://doi.org/10.3390/ijms150610365
Chicago/Turabian StyleMaeng, Jeehye, Vadim Sheverdin, Hyekyoung Shin, Insu Ha, Sun Sik Bae, Hsin-Fang Yang-Yen, and Kyunglim Lee. 2014. "Up-Regulation of Rhoa/Rho Kinase Pathway by Translationally Controlled Tumor Protein in Vascular Smooth Muscle Cells" International Journal of Molecular Sciences 15, no. 6: 10365-10376. https://doi.org/10.3390/ijms150610365