Over-Expression of Platelet-Derived Growth Factor-D Promotes Tumor Growth and Invasion in Endometrial Cancer

Abstract
:1. Introduction
2. Results
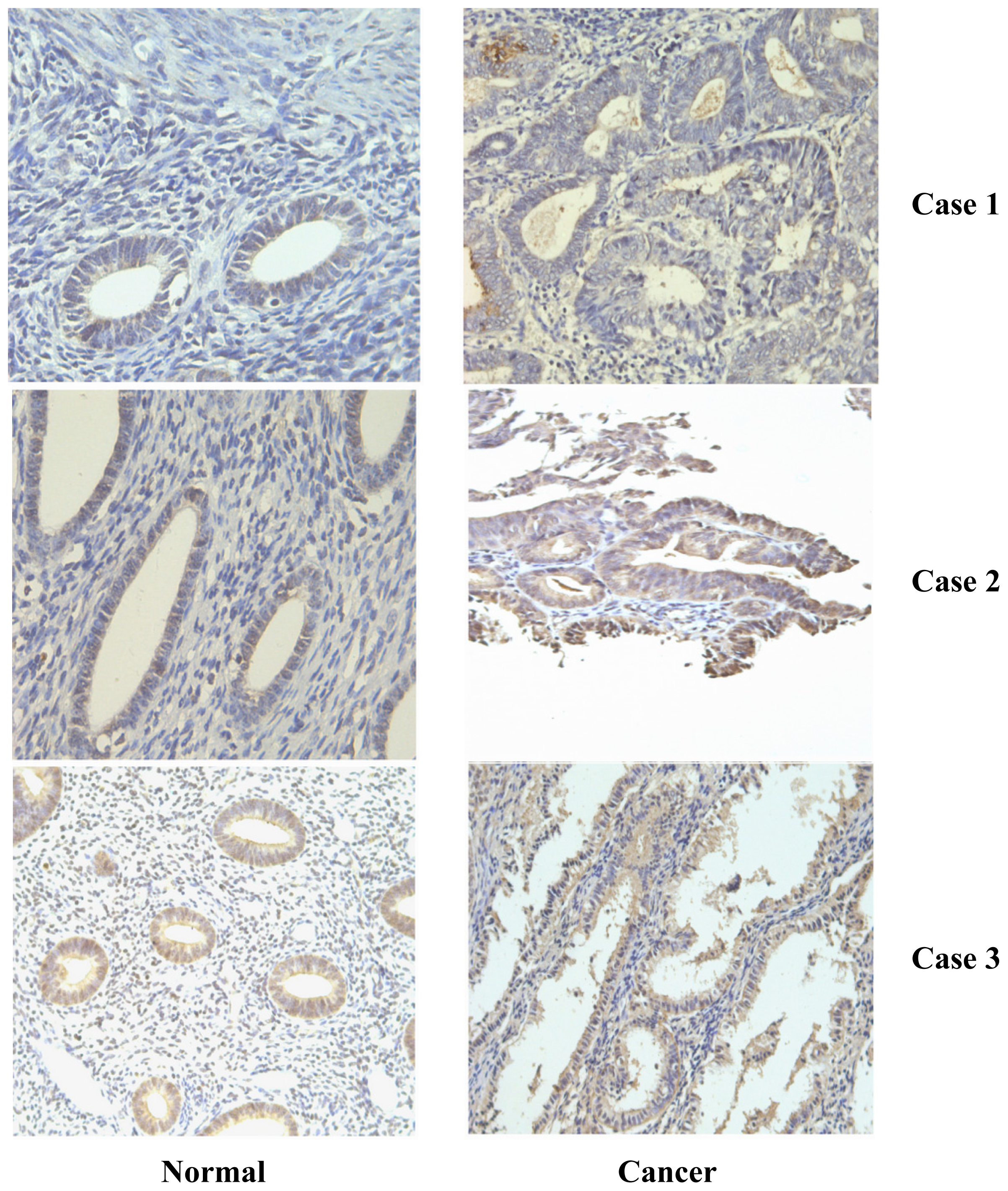
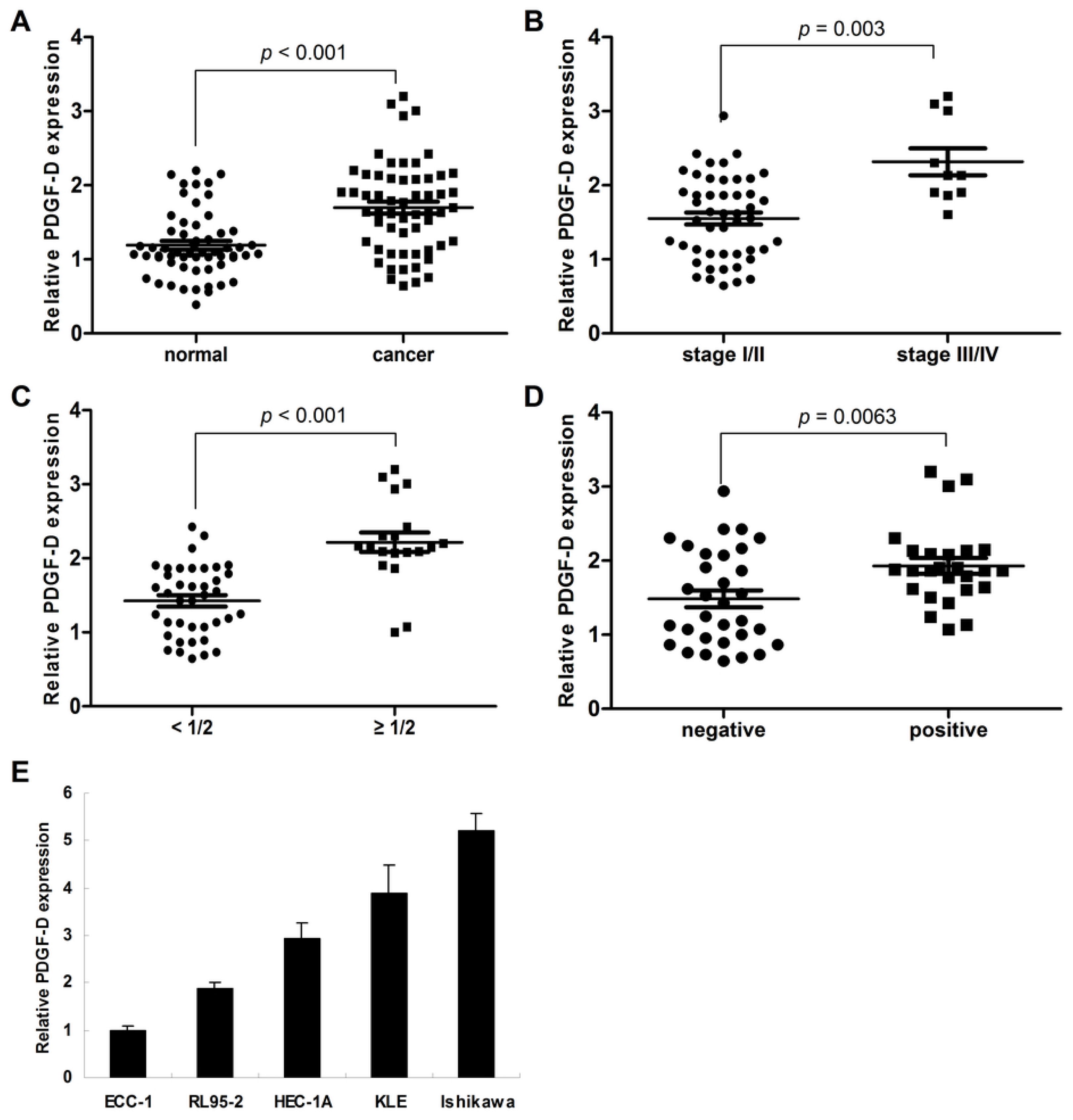
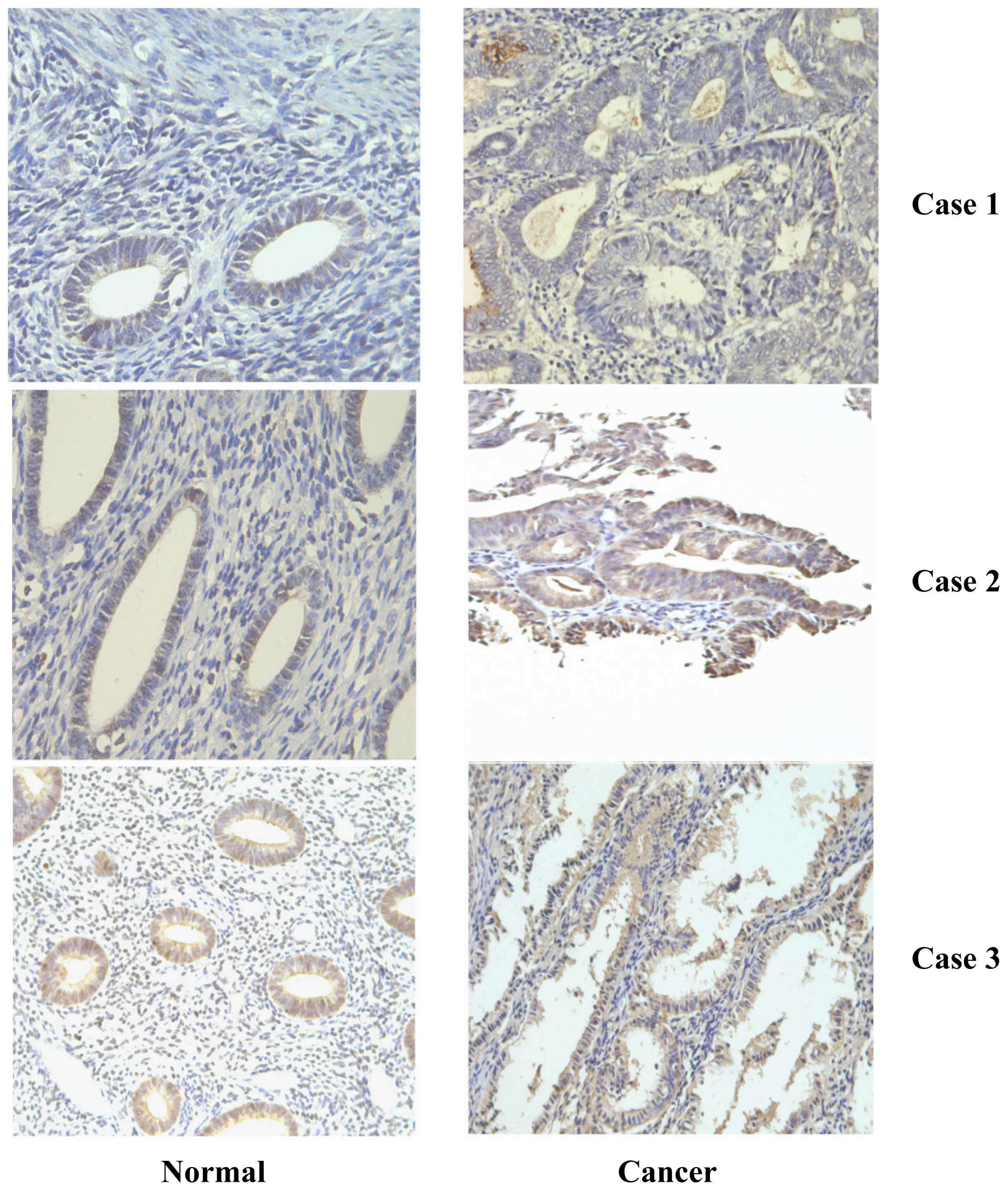
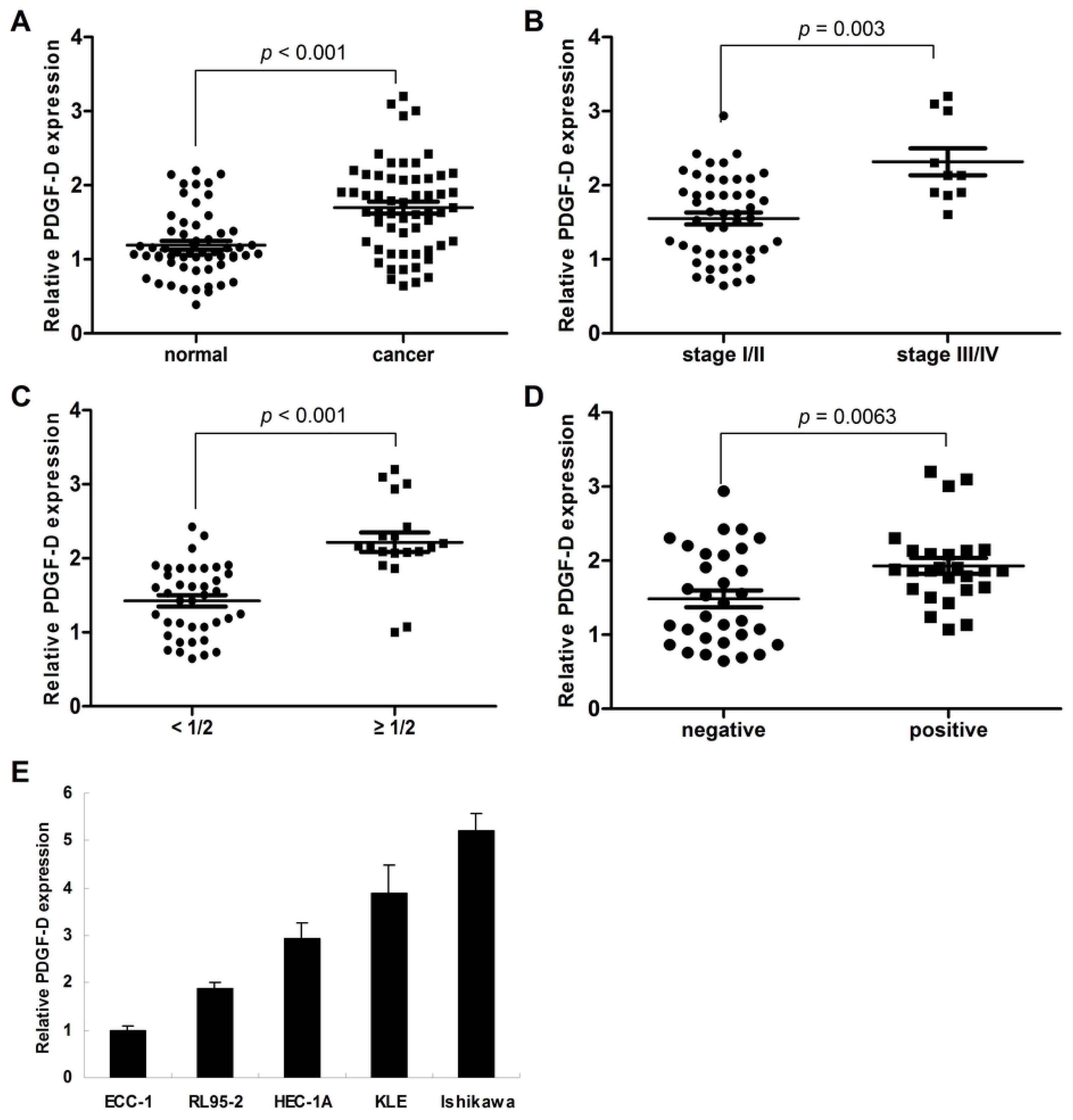
2.1. Frequent Over-Expression of PDGF-D in Human Endometrial Cancer
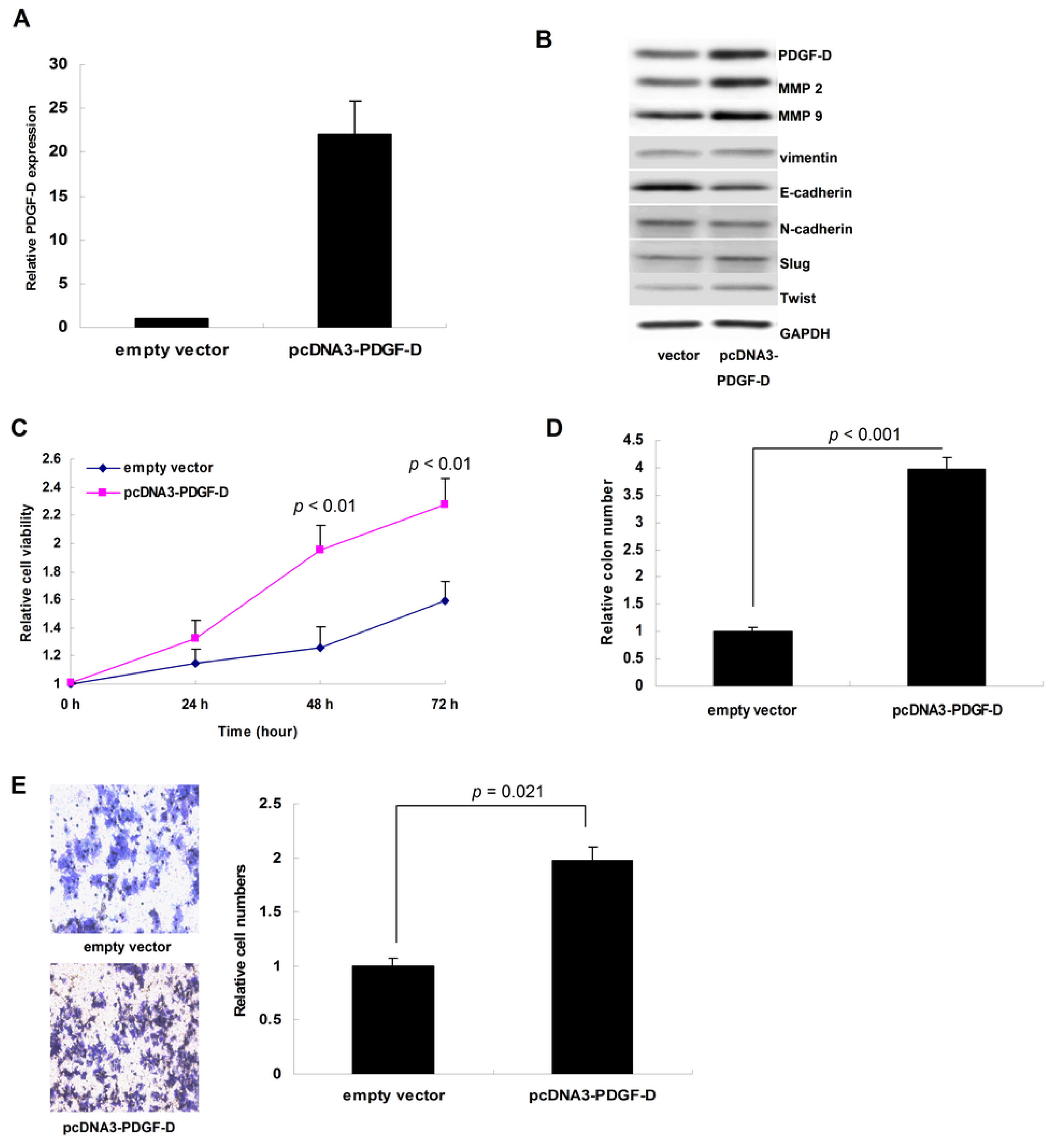
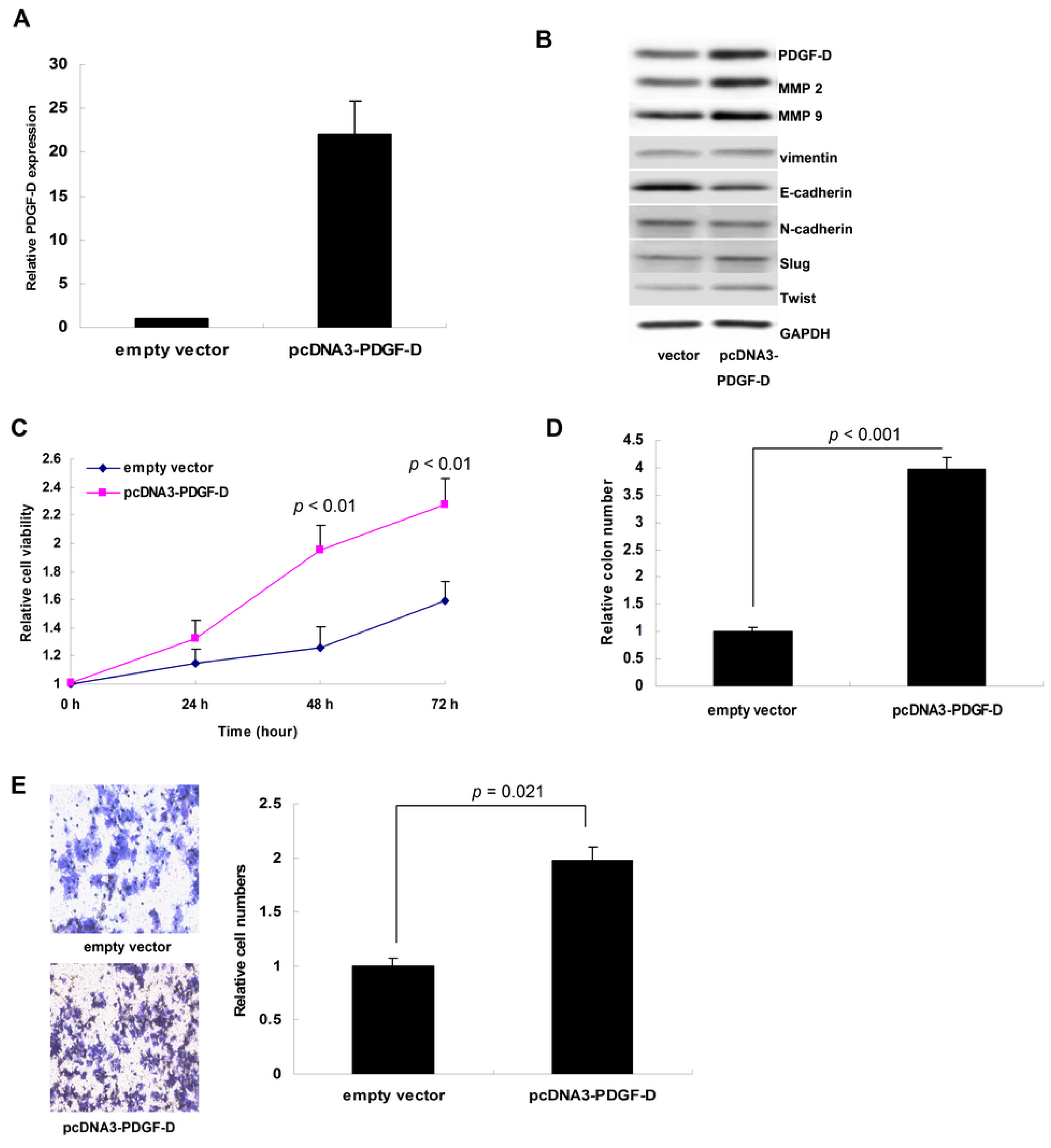
2.2. Up-Regulation of PDGF-D Accelerated Cell Proliferation and Invasion in ECC-1 Cells
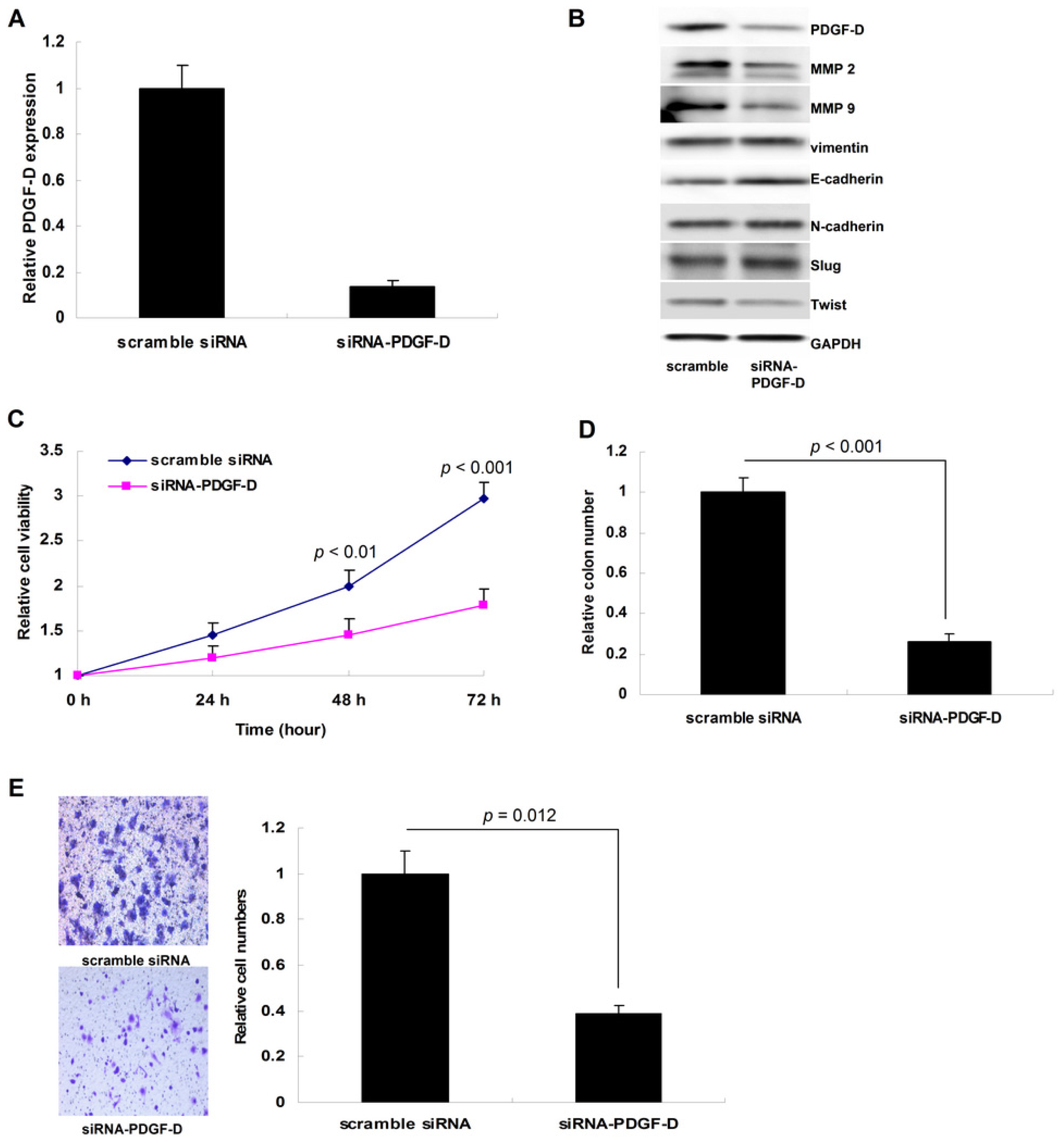
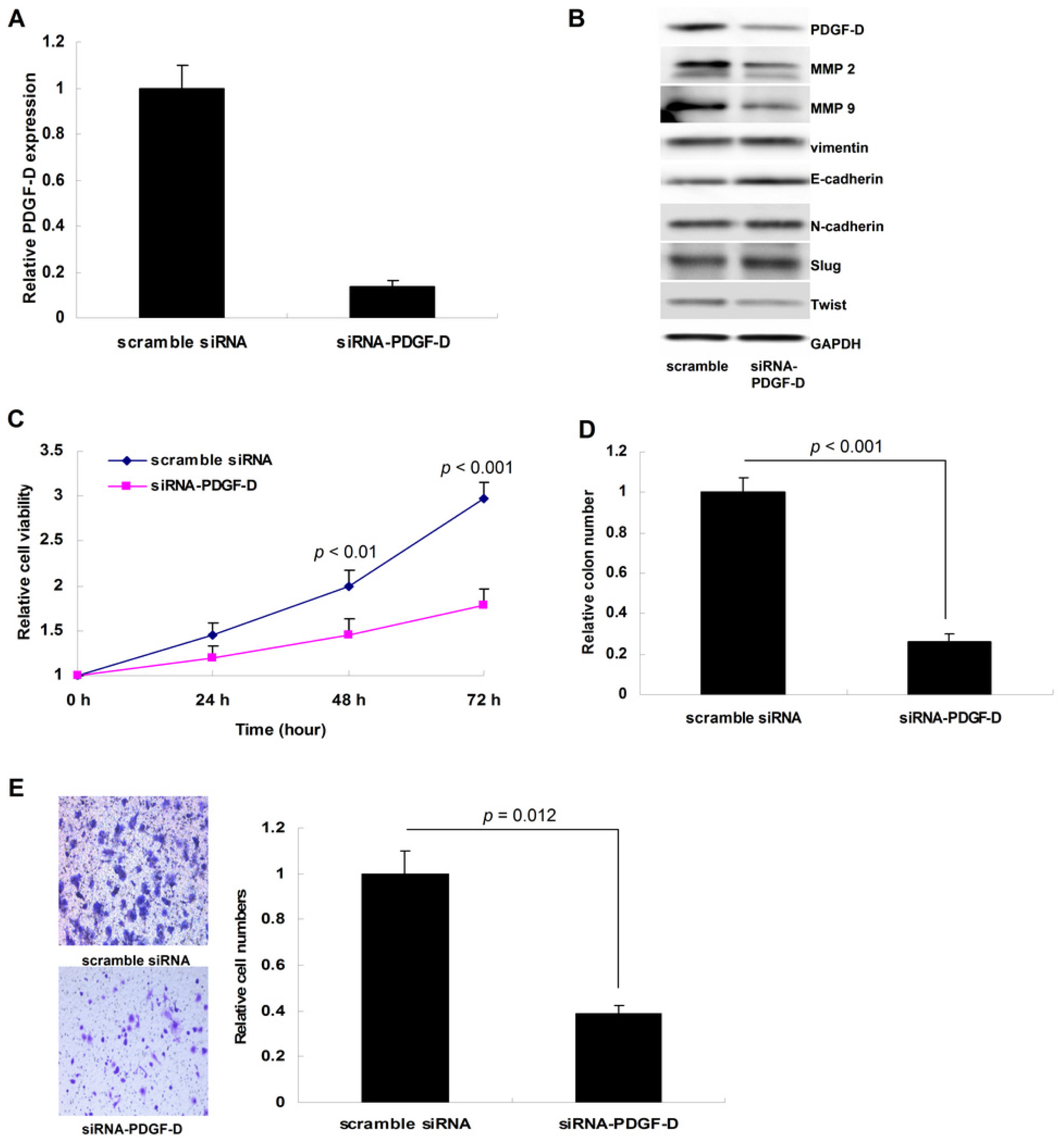
2.3. Silencing PDGF-D Suppressed Tumor Growth and Invasion in Ishikawa Cells
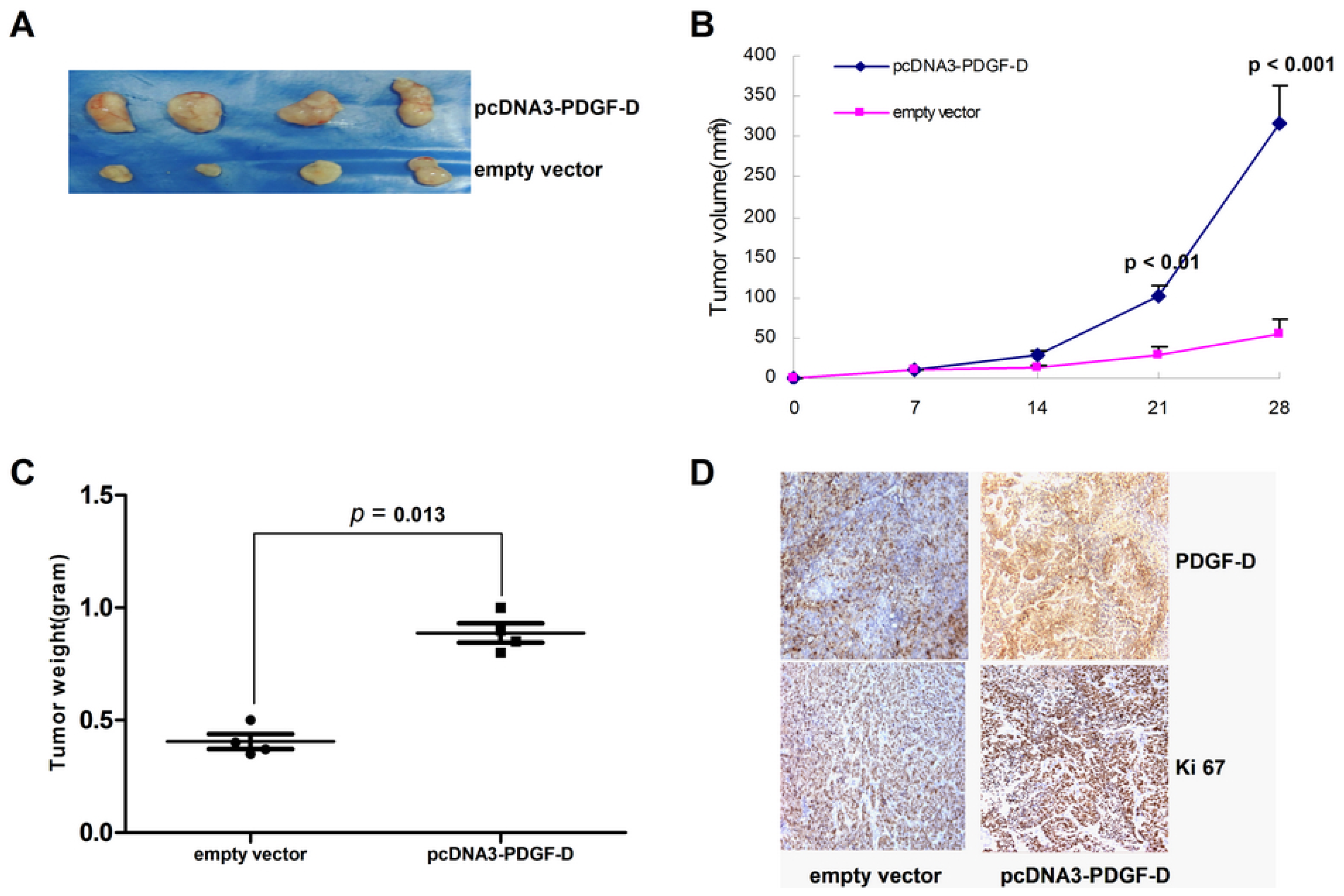
2.4. PDGF-D Promoted Tumor Xenografts Growth
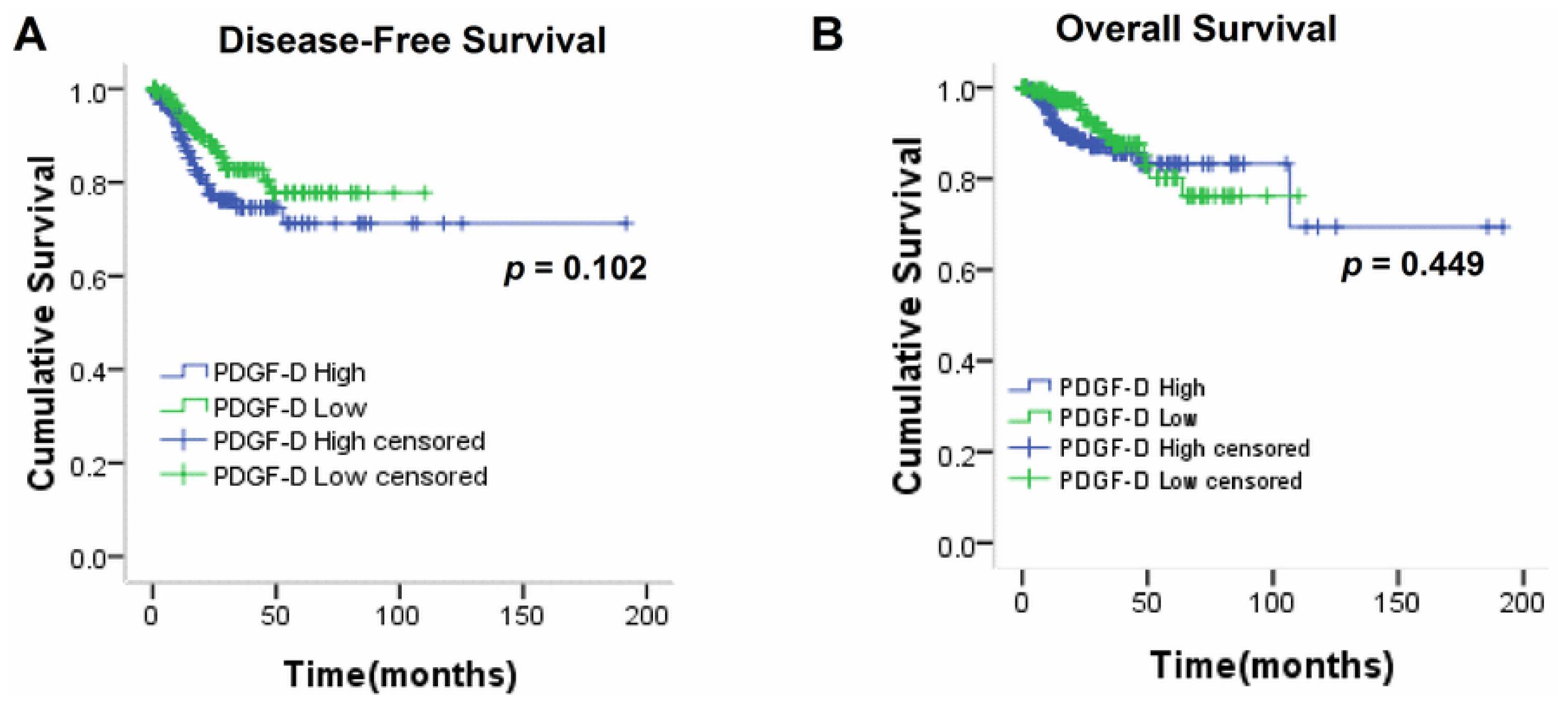
2.5. High PDGF-D Copy Number could Predict Worse Disease-Free Survival
3. Discussion
4. Experimental Section
4.1. Tissue Samples Collection
4.2. Immunohistochemistry
4.3. Cancer Cell Lines and Culture
4.4. Over-Expressing PDGF-D in ECC-1 Cells
4.5. Knockdown of PDGF-D Using siRNA in Ishikawa Cells
4.6. MTT Assay
4.7. Colony Formation Assay
4.8. Cell Invasion Assay
4.9. RNA Extraction and qRT-PCR Amplification
4.10. Western Blot
4.11. Tumor Xenograft Assay
4.12. TCGA (The Cancer Genome Atlas) Data Extraction and Analysis
4.13. Statistical Analysis
5. Conclusions
Supplementary Information
ijms-15-04780-s001.pdfAcknowledgments
Conflicts of Interest
- Author ContributionsJinjin Yu and Yuan Wang conceived and designed the experiments; Yuan Wang and Haifeng Qiu performed these experiments, Weixu Hu and Shaoru Li analyzed the data, Jinjin Yu and Haifeng Qiu wrote the paper.
References
- Amant, F.; Moerman, P.; Neven, P.; Timmerman, D.; van Limbergen, E.; Vergote, I. Endometrial cancer. Lancet 2005, 366, 491–505. [Google Scholar]
- American Cancer Society , Cancer Facts & Figures 2011; American Cancer Society: Atlanta, GA, USA, 2011.
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics 2013. CA Cancer J. Clin. 2013, 63, 11–30. [Google Scholar]
- Dedes, K.J.; Wetterskog, D.; Ashworth, A.; Kaye, S.B.; Reis-Filho, J.S. Emerging therapeutic targets in endometrial cancer. Nat. Rev. Clin. Oncol. 2011, 8, 261–271. [Google Scholar]
- Zhao, L.; Zhang, C.; Liao, G.; Long, J. Rnai-mediated inhibition of PDGF-D leads to decreased cell growth invasion and angiogenesis in the SGC-7901 gastric cancer xenograft model. Cancer Biol. Ther. 2010, 9, 42–48. [Google Scholar]
- Wang, Z.; Ahmad, A.; Li, Y.; Kong, D.; Azmi, A.S.; Banerjee, S.; Sarkar, F.H. Emerging roles of PDGF-D signaling pathway in tumor development and progression. Biochim. Biophys. Acta 2010, 1806, 122–130. [Google Scholar]
- Bergsten, E.; Uutela, M.; Li, X.; Pietras, K.; Ostman, A.; Heldin, C.H.; Alitalo, K.; Eriksson, U. PDGF-D is a specific protease-activated ligand for the PDGF β-receptor. Nat. Cell Biol. 2001, 3, 512–516. [Google Scholar]
- Li, X.; Eriksson, U. Novel PDGF family members: PDGF-C and PDGF-D. Cytokine Growth Factor Rev. 2003, 14, 91–98. [Google Scholar]
- Reigstad, L.J.; Varhaug, J.E.; Lillehaug, J.R. Structural and functional specificities of PDGF-C and PDGF-D the novel members of the platelet-derived growth factors family. FEBS J. 2005, 272, 5723–5741. [Google Scholar]
- Fredriksson, L.; Li, H.; Eriksson, U. The PDGF family: Four gene products form five dimeric isoforms. Cytokine Growth Factor Rev. 2004, 15, 197–204. [Google Scholar]
- Yu, J.; Ustach, C.; Kim, H.R. Platelet-derived growth factor signaling and human cancer. J. Biochem. Mol. Biol. 2003, 36, 49–59. [Google Scholar]
- Wang, Z.; Kong, D.; Li, Y.; Sarkar, F.H. PDGF-D signaling: A novel target in cancer therapy. Curr. Drug Targets 2009, 10, 38–41. [Google Scholar]
- Xu, L.; Tong, R.; Cochran, D.M.; Jain, R.K. Blocking platelet-derived growth factor-D/platelet-derived growth factor receptor β signaling inhibits human renal cell carcinoma progression in an orthotopic mouse model. Cancer Res. 2005, 65, 5711–5719. [Google Scholar]
- Ustach, C.V.; Kim, H.R. Platelet-derived growth factor D is activated by urokinase plasminogen activator in prostate carcinoma cells. Mol. Cell. Biol. 2005, 25, 6279–6288. [Google Scholar]
- Wang, Z.; Kong, D.; Banerjee, S.; Li, Y.; Adsay, N.V.; Abbruzzese, J.; Sarkar, F.H. Down-regulation of platelet-derived growth factor-D inhibits cell growth and angiogenesis through inactivation of notch-1 and nuclear factor-kappab signaling. Cancer Res. 2007, 67, 11377–11385. [Google Scholar]
- Zhao, R.W.; Yang, S.H.; Cai, L.Q.; Zhang, J.; Wang, J.; Wang, Z.H. Roles of vascular endothelial growth factor and platelet-derived growth factor in lymphangiogenesis in epithelial ovarian carcinoma. Zhonghua Fu Chan Ke Za Zhi 2009, 44, 760–764. (in Chinese). [Google Scholar]
- Wong, Y.F.; Cheung, T.H.; Lo, K.W.; Yim, S.F.; Siu, N.S.; Chan, S.C.; Ho, T.W.; Wong, K.W.; Yu, M.Y.; Wang, V.W.; et al. Identification of molecular markers and signaling pathway in endometrial cancer in hong kong chinese women by genome-wide gene expression profiling. Oncogene 2007, 26, 1971–1982. [Google Scholar]
- Zhang, Y.; Liu, Z.; Yu, X.; Zhang, X.; Lu, S.; Chen, X.; Lu, B. The association between metabolic abnormality and endometrial cancer: A large case-control study in China. Gynecol. Oncol. 2010, 117, 41–46. [Google Scholar]
- Wang, Y.; Hu, C.; Dong, R.; Huang, X.; Qiu, H. Platelet-derived growth factor-D promotes ovarian cancer invasion by regulating matrix metalloproteinases 2 and 9. Asian Pac. J. Cancer Prev. 2011, 12, 3367–3370. [Google Scholar]
- Ahmad, A.; Wang, Z.; Kong, D.; Ali, R.; Ali, S.; Banerjee, S.; Sarkar, F.H. Platelet-derived growth factor-D contributes to aggressiveness of breast cancer cells by up-regulating notch and nf-kappab signaling pathways. Breast Cancer Res. Treat. 2010, 126, 15–25. [Google Scholar]
- Chen, X.; Aravindakshan, J.; Yang, Y.; Tiwari-Pandey, R.; Sairam, M.R. Aberrant expression of PDGF ligands and receptors in the tumor prone ovary of follitropin receptor knockout (FORKO) mouse. Carcinogenesis 2006, 27, 903–915. [Google Scholar]
- Kong, D.; Banerjee, S.; Huang, W.; Li, Y.; Wang, Z.; Kim, H.R.; Sarkar, F.H. Mammalian target of rapamycin repression by 33′-diindolylmethane inhibits invasion and angiogenesis in platelet-derived growth factor-D-over-expressing pc3 cells. Cancer Res. 2008, 68, 1927–1934. [Google Scholar]
- Liu, J.; Liao, S.; Huang, Y.; Samuel, R.; Shi, T.; Naxerova, K.; Huang, P.; Kamoun, W.; Jain, R.K.; Fukumura, D.; et al. PDGF-D improves drug delivery and efficacy via vascular normalization but promotes lymphatic metastasis by activating CXCR4 in breast cancer. Clin. Cancer Res. 2011, 17, 3638–3648. [Google Scholar]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar]
- Kang, Y.; Massague, J. Epithelial-mesenchymal transitions: Twist in development and metastasis. Cell 2004, 118, 277–279. [Google Scholar]
- Kong, D.; Wang, Z.; Sarkar, S.H.; Li, Y.; Banerjee, S.; Saliganan, A.; Kim, H.R.; Cher, M.L.; Sarkar, F.H. Platelet-derived growth factor-d over-expression contributes to epithelial-mesenchymal transition of pc3 prostate cancer cells. Stem Cells 2008, 26, 1425–1435. [Google Scholar]
- Devarajan, E.; Song, Y.H.; Krishnappa, S.; Alt, E. Epithelial-mesenchymal transition in breast cancer lines is mediated through PDGF-D released by tissue-resident stem cells. Int. J. Cancer 2012, 131, 1023–1031. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Cases(n) | % |
|---|---|---|
| Age | ||
| <55 | 25 | 43.1% |
| ≥55 | 33 | 56.9% |
| Stage | ||
| I | 38 | 65.5% |
| II | 10 | 20.7% |
| III | 9 | 12.1% |
| IV | 1 | 1.7% |
| Grade | ||
| I | 36 | 62.1% |
| II | 12 | 20.7% |
| III | 10 | 17.2% |
| Myometrial invasion | ||
| <1/2 | 39 | 67.2% |
| ≥1/2 | 19 | 32.8% |
| Lympha node metastasis | ||
| Negative | 18 | 64.3% |
| Positive | 10 | 35.7% |
| Lympha vascular space invasion | ||
| Negative | 32 | 55.2% |
| Positive | 26 | 44.8% |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Y.; Qiu, H.; Hu, W.; Li, S.; Yu, J. Over-Expression of Platelet-Derived Growth Factor-D Promotes Tumor Growth and Invasion in Endometrial Cancer. Int. J. Mol. Sci. 2014, 15, 4780-4794. https://doi.org/10.3390/ijms15034780
Wang Y, Qiu H, Hu W, Li S, Yu J. Over-Expression of Platelet-Derived Growth Factor-D Promotes Tumor Growth and Invasion in Endometrial Cancer. International Journal of Molecular Sciences. 2014; 15(3):4780-4794. https://doi.org/10.3390/ijms15034780
Chicago/Turabian StyleWang, Yuan, Haifeng Qiu, Weixu Hu, Shaoru Li, and Jinjin Yu. 2014. "Over-Expression of Platelet-Derived Growth Factor-D Promotes Tumor Growth and Invasion in Endometrial Cancer" International Journal of Molecular Sciences 15, no. 3: 4780-4794. https://doi.org/10.3390/ijms15034780