The Glutathione Peroxidase Gene Family in Thellungiella salsuginea: Genome-Wide Identification, Classification, and Gene and Protein Expression Analysis under Stress Conditions

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
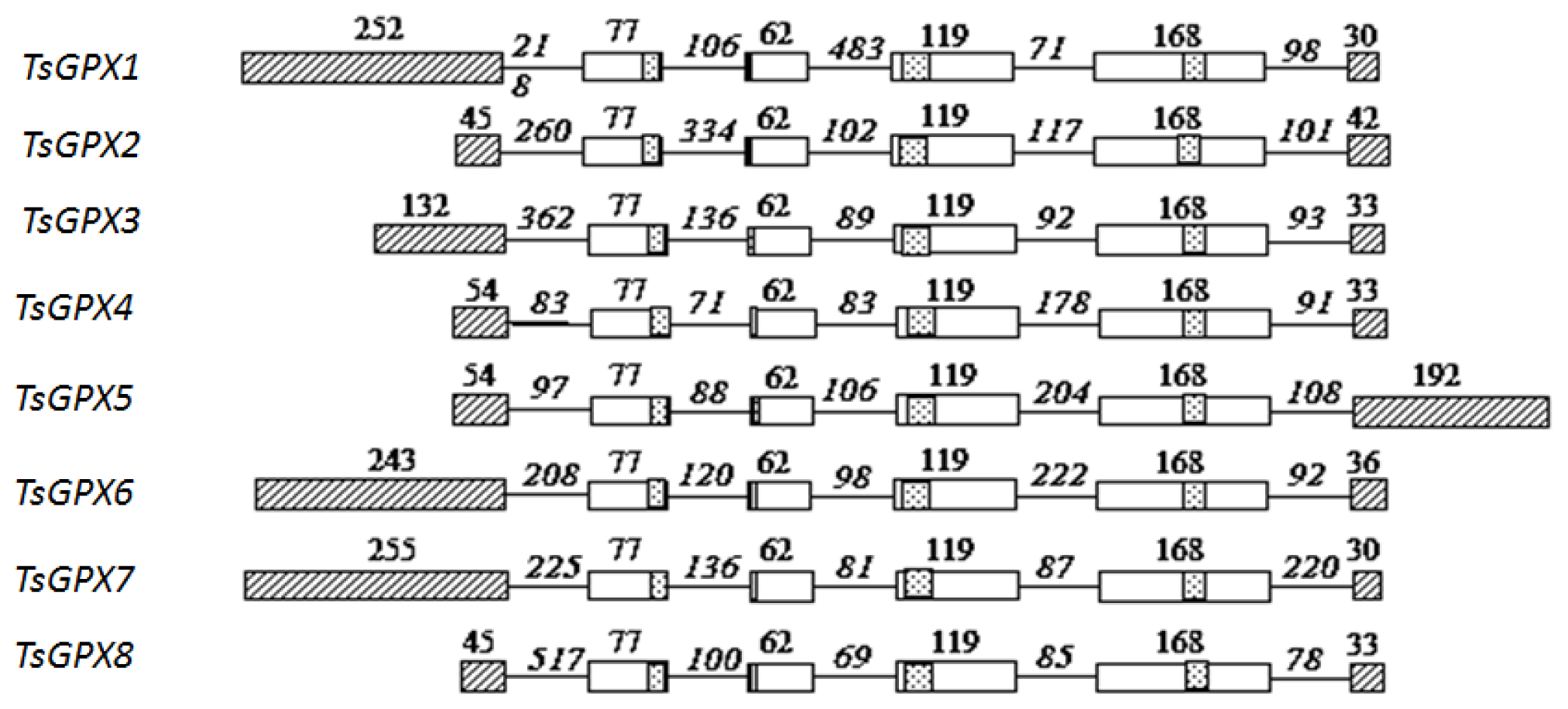
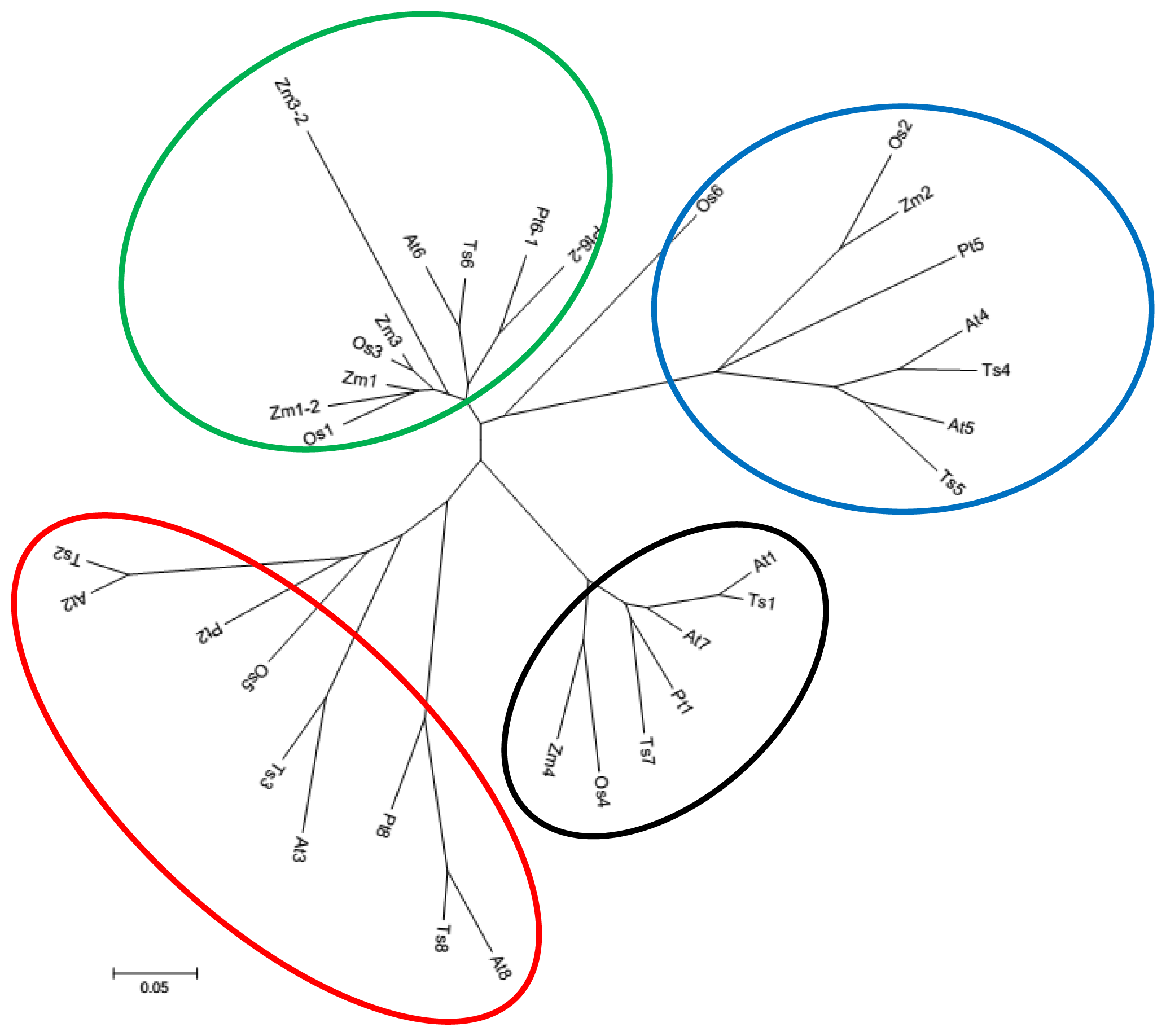
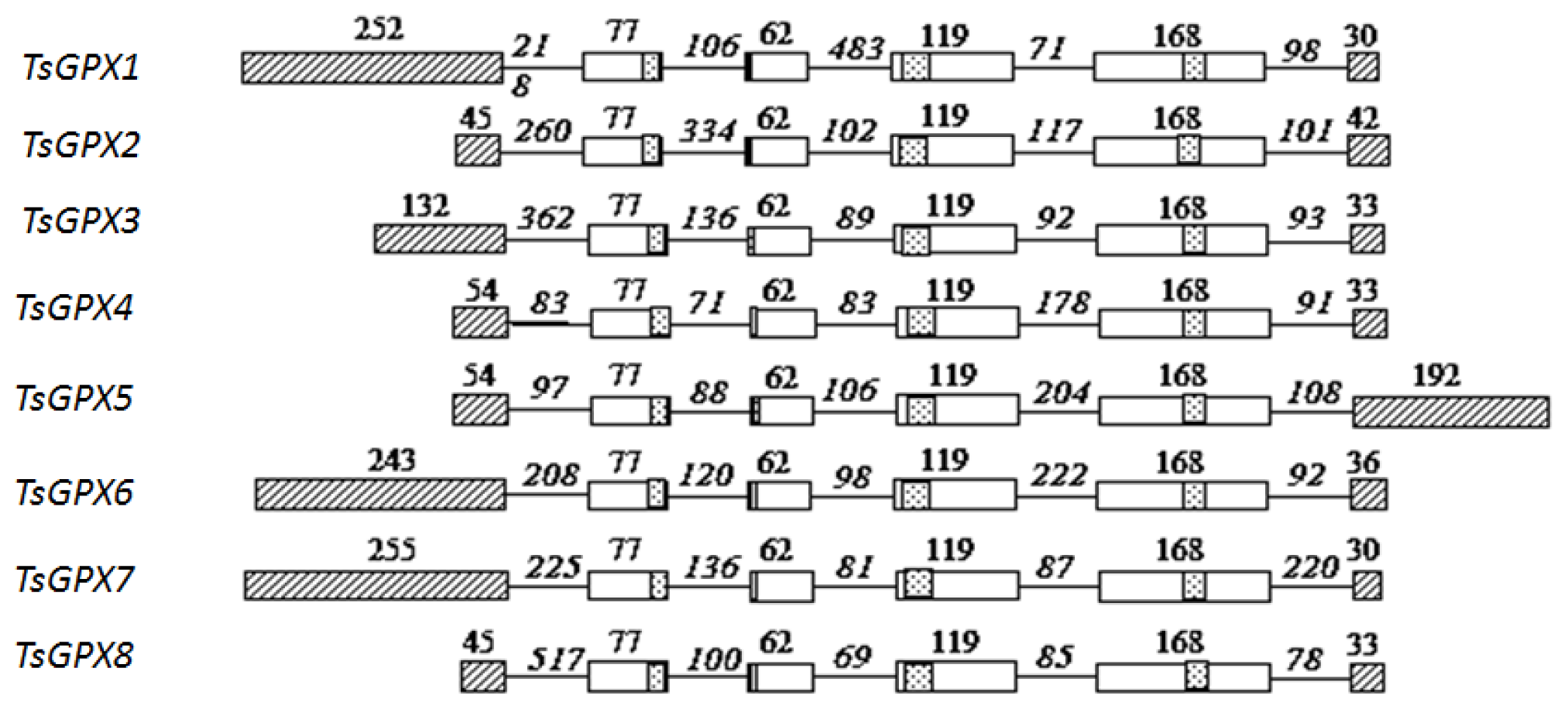
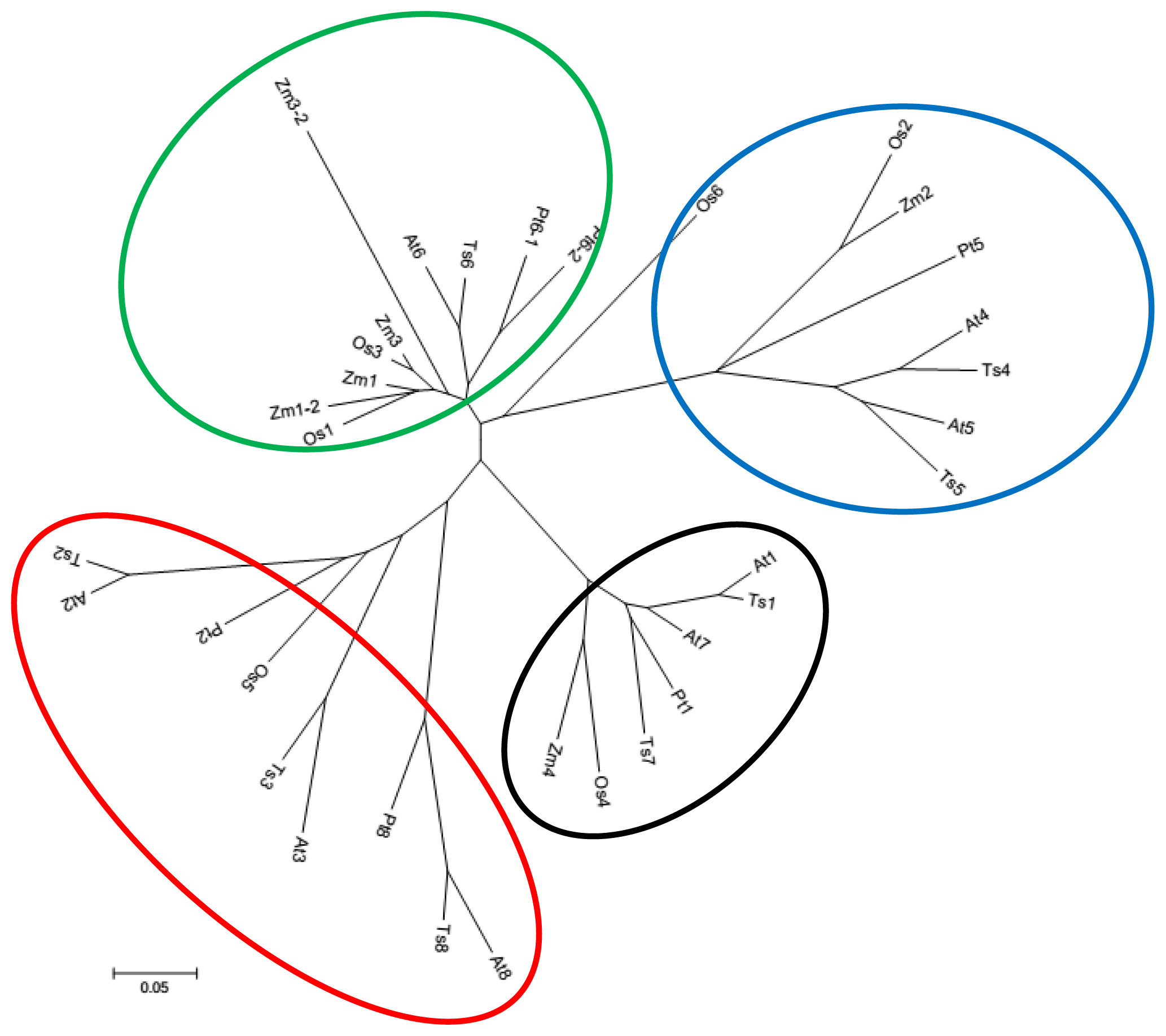
2.1. Identification and Characterization of TsGPX Genes
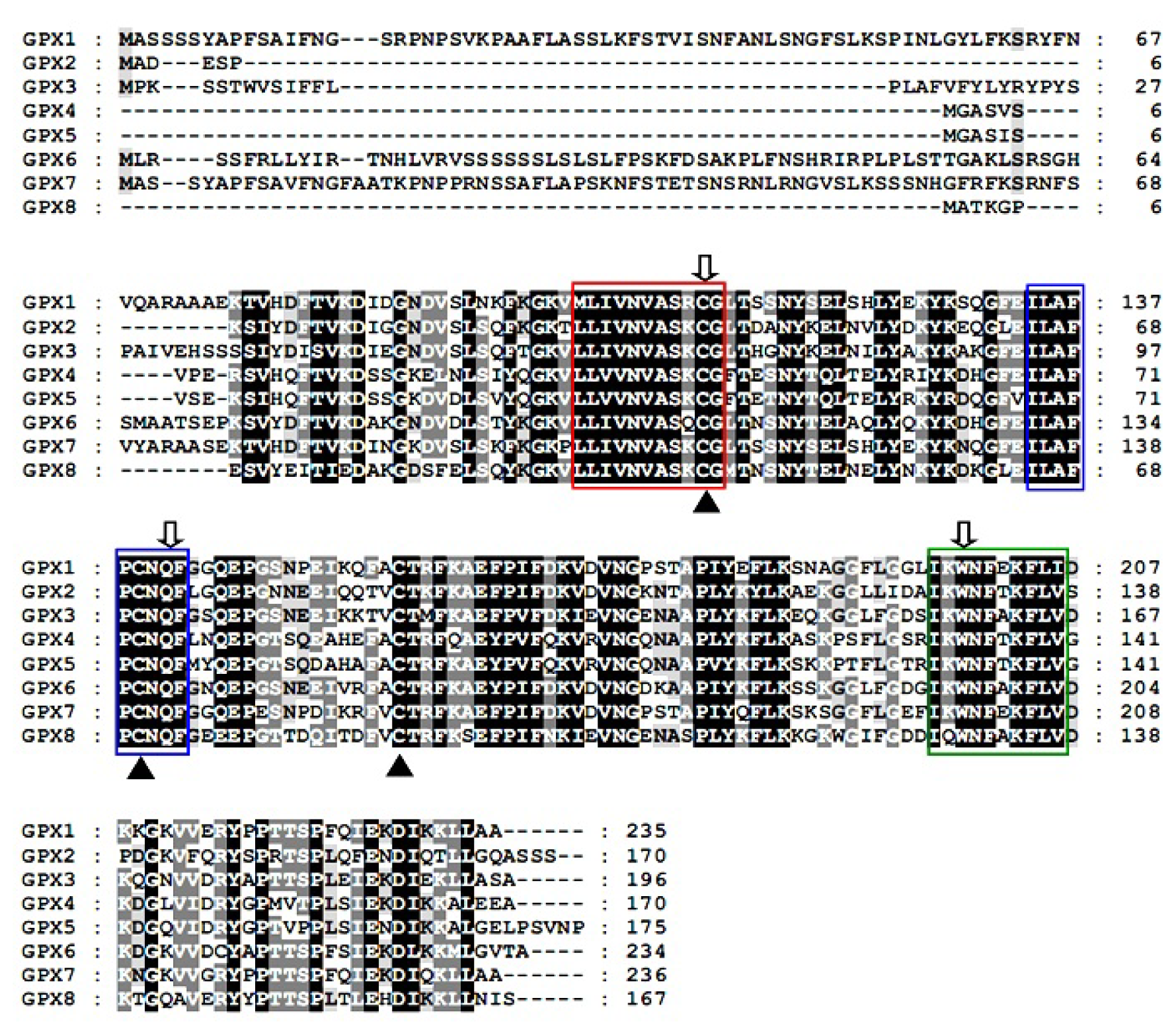
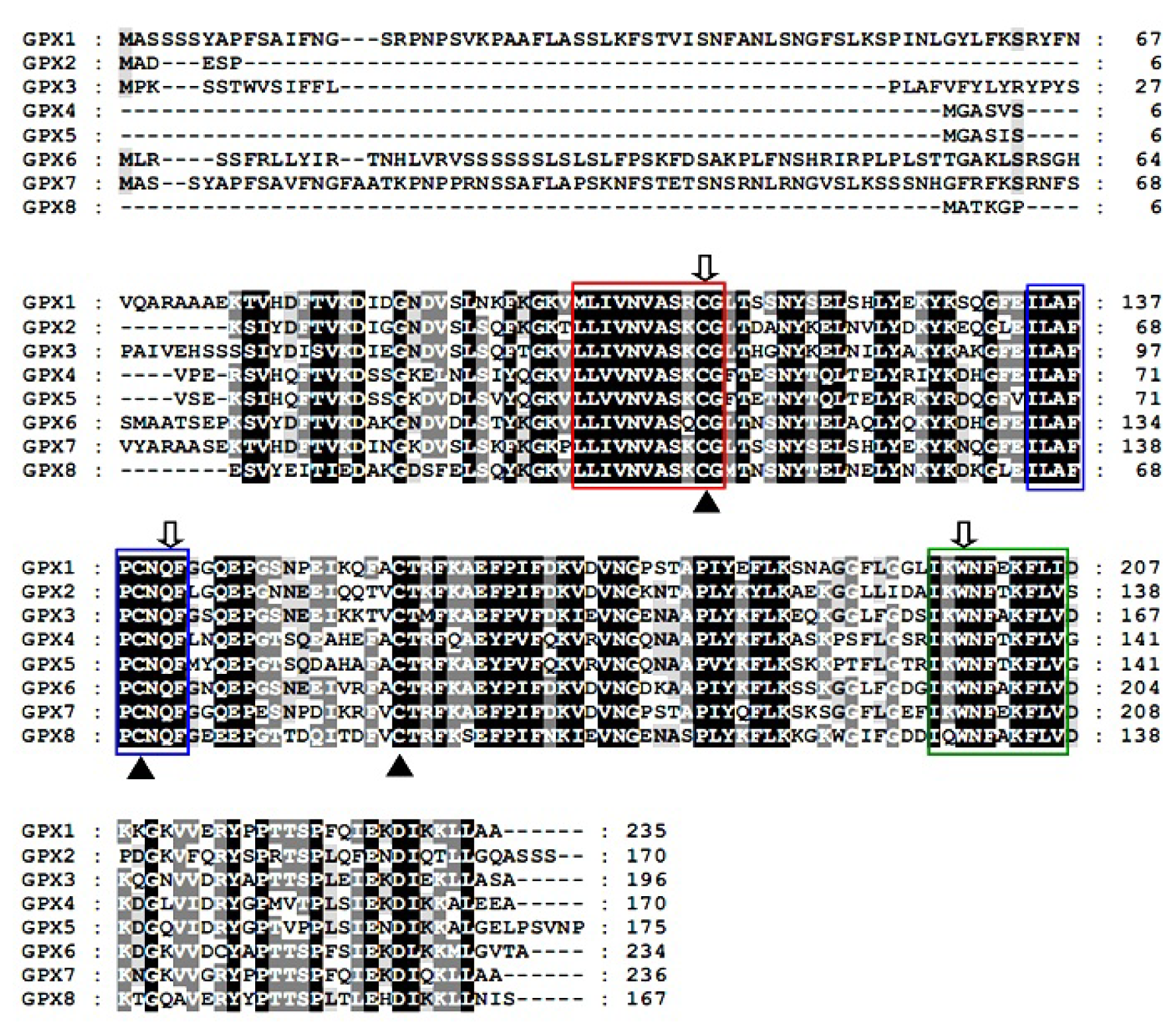
2.2. Predicted Properties of TsGPX Proteins
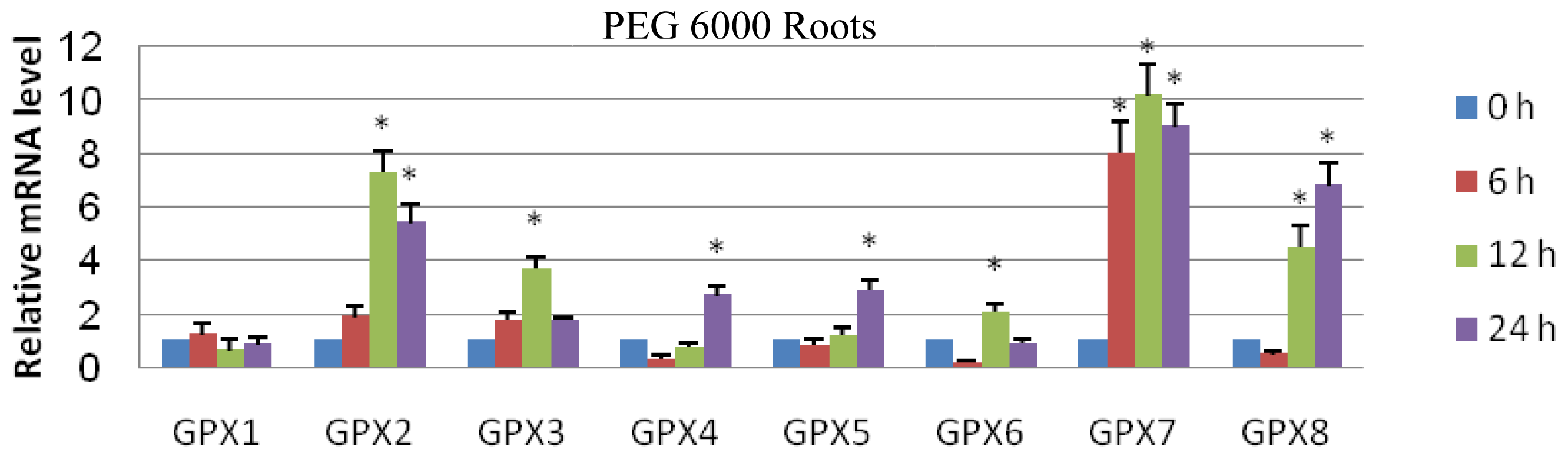
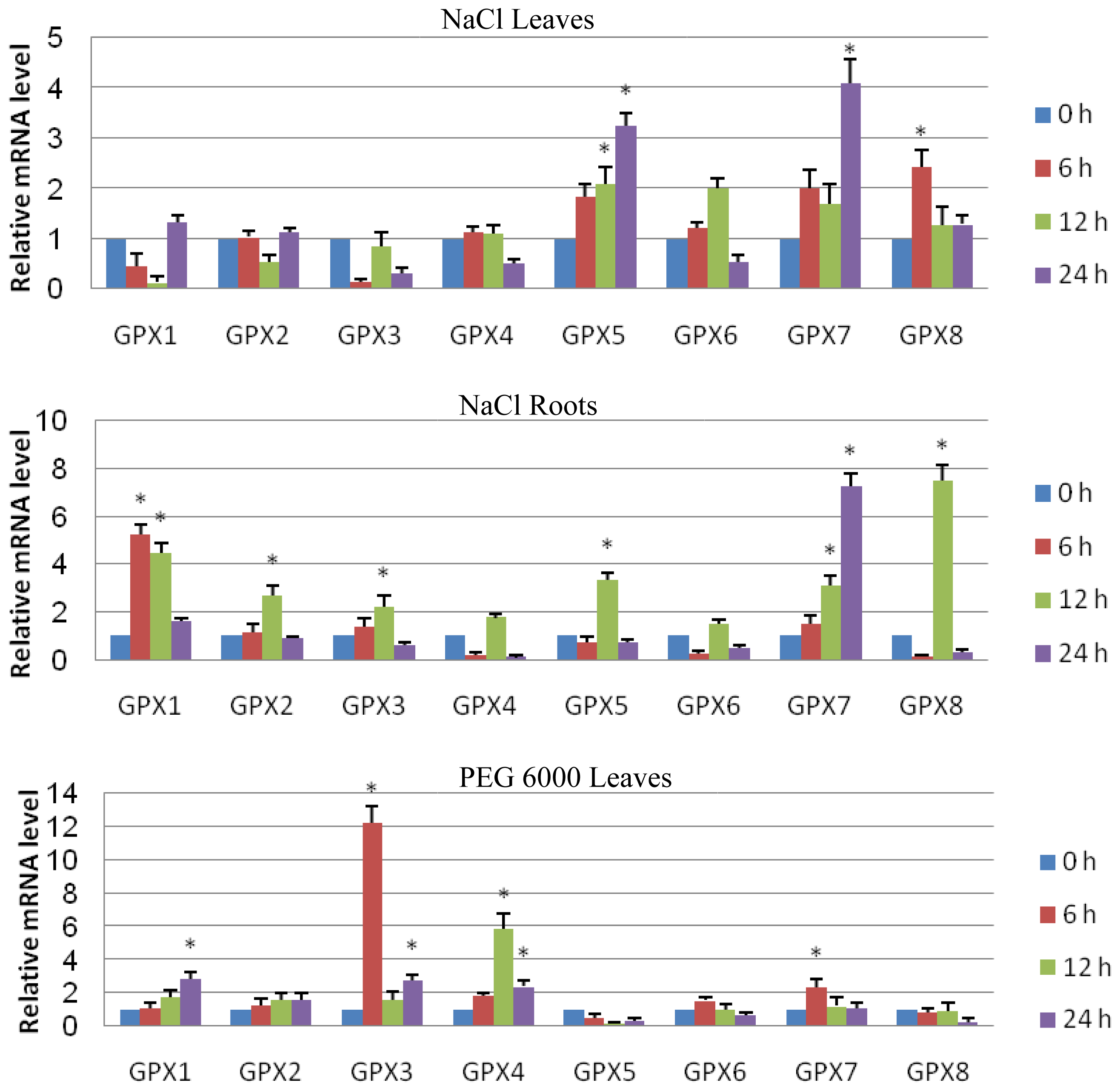
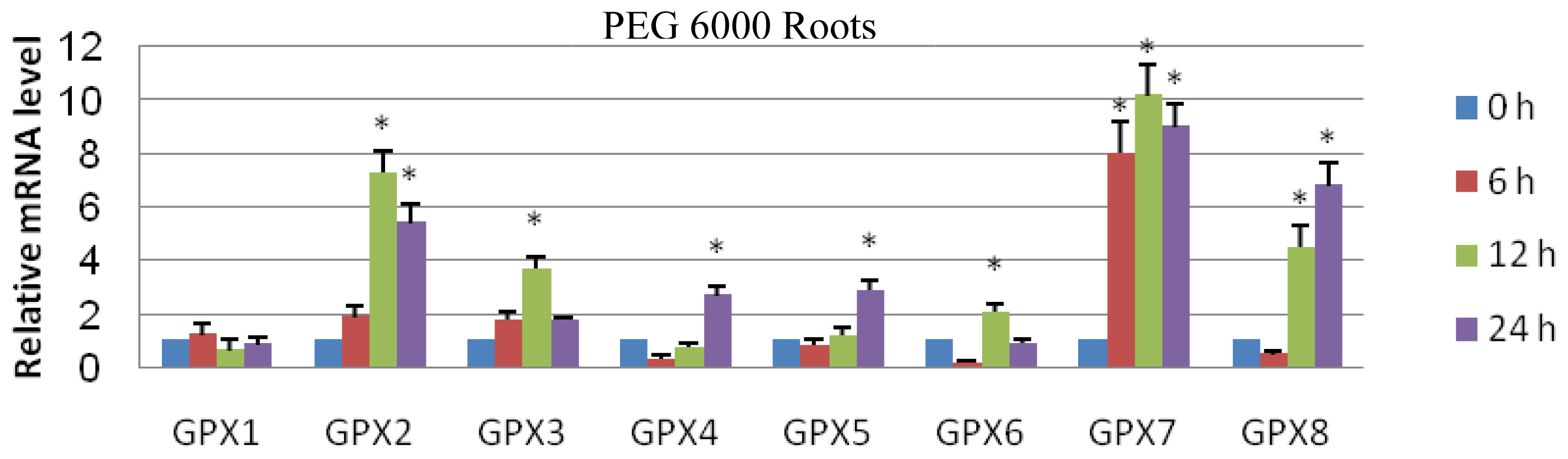
2.3. Expression Analyses of GPX Genes in T. salsuginea Plants Exposed to High Salinity and Osmotic Treatment
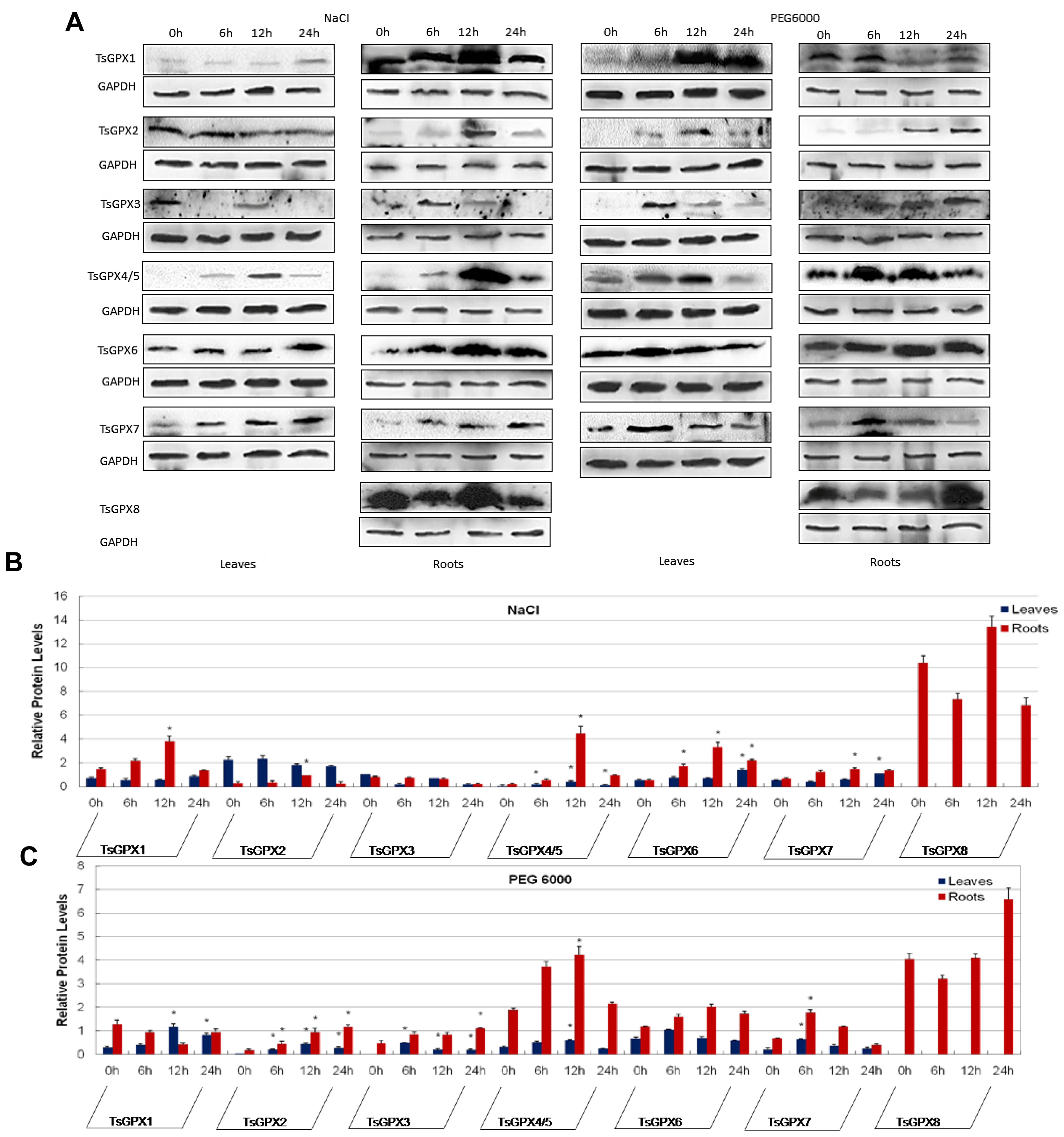
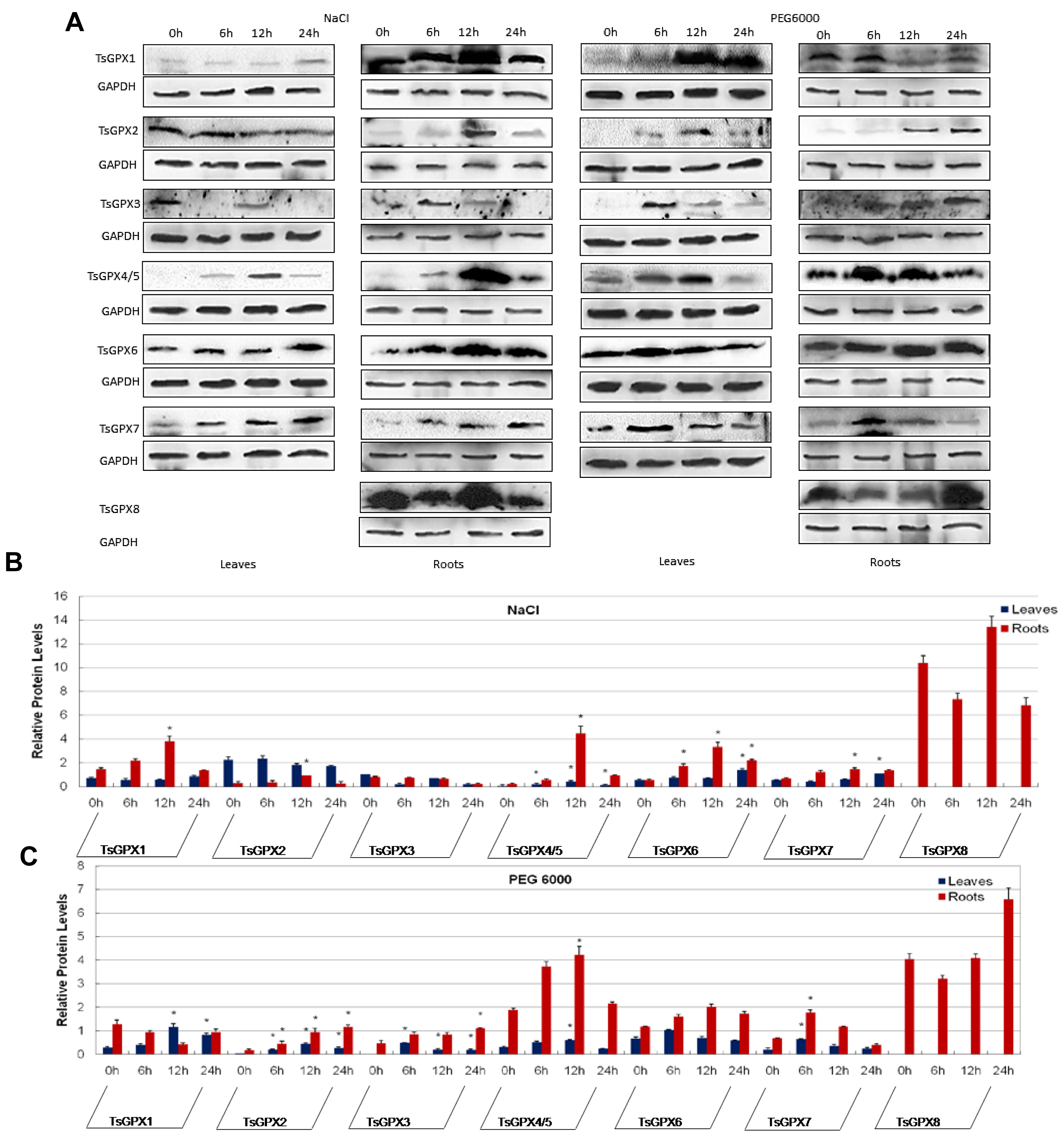
2.4. Protein Expression Analyses of GPX Genes in T. salsuginea Plants Exposed to High Salinity and Osmotic Treatments
3. Experimental Section
3.1. Plant Materials and Stress Treatments
3.2. Isolation of the Eight TsGPX Genes
3.3. Sequence Alignments, Phylogenetic Analysis, and Other Bioinformatics Analysis
3.4. Quantitative Real-Time RT-PCR
3.5. Antibody Preparation and Western Blotting Analysis
4. Conclusions
Supplementary Information
ijms-15-03319-s001.pdfAcknowledgments
Conflicts of Interest
- Author ContributionsYinjun Zhou and Fei Gao conceived and designed the experiments, supervised the project, interpreted the data, and wrote the manuscript. Tingting Ma and Jing Chen conducted the quantitative real-time RT-PCR. Jing Chen, Huayun Li, Zhanglei Li, Ning Wang and Zichen Zhang performed the Western blotting analysis. Tingting Ma and Huayun Li conducted the bioinformatics analysis.
References
- Grene, R. Oxidative stress and acclimation mechanisms in plants. The Arabidopsis Book 2002, 1, e0036. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar]
- Dat, J.; Vandenabeele, S.; Vranová, E.; van Montagu, M.; Inzé, D.; van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci 2000, 57, 779–795. [Google Scholar]
- Ursini, F.; Maiorino, M.; Brigelius-Flohé, R.; Aumann, K.D.; Roveri, A.; Schomburg, D.; Flohé, L. Diversity of glutathione peroxidases. Methods Enzymol 1995, 252, 38–53. [Google Scholar]
- Herbette, S.; Roeckel-Drevet, P.; Drevet, J.R. Seleno-independent glutathione peroxidases: More than simple antioxidant scavengers. FEBS J 2007, 274, 2163–2180. [Google Scholar]
- Espinola-Klein, C.; Rupprecht, H.J.; Bickel, C.; Schnabel, R.; Genth-Zotz, S.; Torzewski, M.; Lackner, K.; Munzel, T.; Blankenberg, S. AtheroGene Investigators. Glutathione peroxidase-1 activity, atherosclerotic burden, and cardiovascular prognosis. Am. J. Cardiol 2007, 99, 808–812. [Google Scholar]
- Criqui, M.C.; Jamet, E.; Parmentier, Y.; Marbach, J.; Durr, A.; Fleck, J. Isolation and characterization of a plant cDNA showing homology to animal glutathione peroxidases. Plant Mol. Biol 1992, 18, 623–627. [Google Scholar]
- Holland, D.; Ben-Hayyim, G.; Faltin, Z.; Camoin, L.; Strosberg, A.D.; Eshdat, Y. Molecular characterization of salt-stress associated protein in citrus: protein and cDNA sequence homology to mammalian glutathione peroxidases. Plant Mol. Biol 1993, 21, 923–927. [Google Scholar]
- Johnson, R.R.; Cranston, H.J.; Chaverra, M.E.; Dyer, W.E. Characterization of cDNA clones for differentially expressed genes in embryos of dormant and non-dormant Avenua fatua (L.) caryopses. Plant Mol. Biol 1995, 28, 113–122. [Google Scholar]
- Gaber, A.; Ogata, T.; Maruta, T.; Yoshimura, K.; Tamoi, M.; Shigeoka, S. The involvement of Arabidopsis glutathione peroxidase 8 in the suppression of oxidative damage in the nucleus and cytosol. Plant Cell Physiol 2012, 53, 1596–1606. [Google Scholar]
- Eshdat, Y.; Holland, D.; Faltin, Z.; Ben-Hayyim, G. Plant glutathione peroxidases. Plant Physiol 1997, 100, 234–240. [Google Scholar]
- Sugimoto, M.; Furui, S.; Suzuki, Y. Molecular cloning and characterization of a cDNA encoding putative phospholipid hydroperoxide glutathione from spinach. Biosci. Biotechnol. Biochem 1997, 61, 1379–1381. [Google Scholar]
- Roeckel-Drevet, P.; Gagne, G.; de Labrouhe, T.D.; Dufaure, J.P.; Nicolas, P.; Drevet, J.R. Molecular characterization, organ distribution and stress-mediated induction of two glutathione peroxidase-encoding mRNAs in sunflower (Helianthus annuus). Physiol. Plant 1998, 103, 385–394. [Google Scholar]
- Mullineaux, P.M.; Karpinski, S.; Jimenez, A.; Clearly, S.P.; Robinson, C.; Creissen, G.P. Identification of cDNAs encoding plastid-targeted glutathione peroxidase. Plant J 1998, 13, 375–379. [Google Scholar]
- Depege, D.; Drevet, J.; Boyer, N. Molecular cloning and characterization of tomato cDNA encoding glutathione peroxidase-like proteins. Eur. J. Biochem 1998, 253, 445–451. [Google Scholar]
- Passaia, G.; Fonini, L.S.; Caverzan, A.; Jardim-Messeder, D.; Christoff, A.P.; Gaeta, M.L.; de Araujo Mariath, J.E.; Margis, R.; Margis-Pinheiro, M. The mitochondrial glutathione peroxidase GPX3 is essential for H2O2 homeostasis and root and shoot development in rice. Plant Sci 2013, 208, 93–101. [Google Scholar]
- Zhai, C.Z.; Zhao, L.; Yin, L.J.; Chen, M.; Wang, Q.Y.; Li, L.C.; Xu, Z.S.; Ma, Y.Z. Two wheat glutathione peroxidase genes whose products are located in chloroplasts improve salt and H2O2 tolerances in Arabidopsis. PLoS One 2013, 8, e73989. [Google Scholar]
- Kim, Y.J.; Jang, M.G.; Noh, H.Y.; Lee, H.J.; Sukweenadhi, J.; Kim, J.H.; Kim, S.Y.; Kwon, W.S.; Yang, D.C. Molecular characterization of two glutathione peroxidase genes of Panax ginseng and their expression analysis against environmental stresses. Gene 2014, 535, 33–41. [Google Scholar]
- Rodriguez Milla, M.A.; Maure, A.; Rodriguez Huete, A.; Gustafson, J.P. Glutathione peroxidase genes in Arabidopsis are ubiquitous and regulated by abiotic stresses through diverse signaling pathways. Plant J 2003, 36, 602–615. [Google Scholar]
- Romas, J.; Matamoros, M.A.; Naya, L.; James, E.K.; Rouhier, N.; Sato, S.; Tabata, S.; Becana, M. The glutathione peroxidase gene family of Lotus japonicus: Characterization of genomic clones, expression analyses and immunolocalization in legumes. New Phytol 2009, 181, 103–114. [Google Scholar]
- Navrot, N.; Collin, V.; Gualberto, J.; Gelhaye, E.; Hirasawa, M.; Rey, P.; Knaff, D.B.; Issakidis, E.; Jacquot, J.P.; Rouhier, N. Plant glutathione peroxidases are functional peroxiredoxins distributed in several subcellular compartments and regulated during biotic and abiotic stresses. Plant Physiol 2006, 142, 1364–1379. [Google Scholar]
- Koch, M.A.; German, D.A. Taxonomy and systematics are key to biological information: Arabidopsis, Eutrema (Thellungiella), Noccaea and Schrenkiella (Brassicaceae) as examples. Front. Plant Sci 2013, 4, 267. [Google Scholar]
- Batelli, G.; Oh, D.H.; D’Urzo, M.P.; Orsini, F.; Dassanayake, M.; Zhu, J.K.; Bohnert, H.J.; Bressan, R.A.; Maggio, A. Using Arabidopsis-related model species (ARMS): Growth, genetic transformation, and comparative genomics. Methods Mol. Biol 2014, 1062, 27–51. [Google Scholar]
- Lee, Y.P.; Giorgi, F.M.; Lohse, M.; Kvederaviciute, K.; Klages, S.; Usadel, B.; Meskiene, I.; Reinhardt, R.; Hincha, D.K. Transcriptome sequencing and microarray design for functional genomics in the extremophile Arabidopsis relative Thellungiella salsuginea (Eutrema salsugineum). BMC Genomics 2013, 14, 793. [Google Scholar]
- Taji, T.; Seki, M.; Satou, M.; Sakurai, T.; Kobayashi, M.; Ishiyama, K.; Narusaka, Y.; Narusaka, M.; Zhu, J.K.; Shinozaki, K. Comparative genomics in salt tolerance between Arabidopsis and Arabidopsis-related halophyte salt cress using Arabidopsis microarray. Plant Physiol 2004, 135, 1697–1709. [Google Scholar]
- M’rah, S.; Ouerghi, Z.; Eymery, F.; Rey, P.; Hajji, M.; Grignon, C.; Lachaâl, M. Efficiency of biochemical protection against toxic effects of accumulated salt differentiates Thellungiella halophila from Arabidopsis thaliana. J. Plant Physiol 2007, 164, 375–384. [Google Scholar]
- Avsian-Kretchmer, O.; Gueta-Dahan, Y.; Lev-Yadun, S.; Ben-Hayyim, G. The salt-stress signal transduction pathway that activates the gpx1 promoter is mediated by intracellular H2O2, different from the pathway induced by extracellular H2O2. Plant Physiol 2004, 135, 1685–1696. [Google Scholar]
- Churin, Y.; Schilling, S.; Börner, T. A gene family encoding glutathione peroxidase homologues in Hordeum vulgare (barley). FEBS Lett 1999, 459, 33–38. [Google Scholar]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res 2002, 30, 325–327. [Google Scholar]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione peroxidase family—An evolutionary overview. FEBS J 2008, 275, 3959–3970. [Google Scholar]
- Kilian, J.; Whitehead, D.; Horak, J.; Wanke, D.; Weinl, S.; Batistic, O.; D’Angelo, C.; Bornberg-Bauer, E.; Kudla, J.; Harter, K. The AtGenExpress global stress expression data set: protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J 2007, 50, 347–363. [Google Scholar]
- Kant, S.; Kant, P.; Raveh, E.; Barak, S. Evidence that differential gene expression between the halophyte, Thellungiella halophila, and Arabidopsis thaliana is responsible for higher levels of the compatible osmolyte proline and tight control of Na+ uptake in T. halophila. Plant Cell Environ 2006, 29, 1220–1234. [Google Scholar]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci 2001, 6, 66–71. [Google Scholar]
- Lan, P.; Li, W.; Schmidt, W. Complementary proteome and transcriptome profiling in phosphate-deficient Arabidopsis roots reveals multiple levels of gene regulation. Mol. Cell. Proteomics 2012, 11, 1156–1166. [Google Scholar]
- Boyer, J.S.; James, R.A.; Munns, R.; Condon, A.G.; Passioura, J.B. Osmotic adjustment may lead to anomalously low estimates of relative water content in wheat and barley. Funct. Plant Biol 2008, 35, 1172–1182. [Google Scholar]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res 2003, 31, 3497–3500. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol 2013, 30, 2725–2729. [Google Scholar]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol 2000, 300, 1005–1016. [Google Scholar]
- Nakai, K.; Kanehisa, M. A knowledge base for predicting protein localization sites in eukaryotic cells. Genomics 1992, 14, 897–911. [Google Scholar]
- Reese, M.G. Application of a time-delay neural network to promoter annotation in the Drosophila melanogaster genome. Comput. Chem 2001, 26, 51–56. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar]
- Gao, F.; Zhou, Y.; Zhu, W.; Li, X.; Fan, L.; Zhang, G. Proteomic analysis of cold stress-responsive proteins in Thellungiella rosette leaves. Planta 2009, 230, 1033–1046. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Sequence ID | Name | Location in genome |
|---|---|---|---|
| 1 | Thhalv10000311m | GPX1 | scaffold_15: 4940063–4941983 |
| 2 | Thhalv10000351m | GPX1 isoform | scaffold_15: 4940063–4941470 |
| 3 | Thhalv10017319m | GPX2 | scaffold_10: 6460958–6462801 |
| 4 | Thhalv10001645m | GPX3 | scaffold_22: 63411–66356 |
| 5 | Thhalv10001644m | GPX3 isoform | scaffold_22: 63411–66356 |
| 6 | Thhalv10001660m | GPX4 | scaffold_22: 1860223–1861484 |
| 7 | Thhalv10006271m | GPX5 | scaffold_19: 155838–157677 |
| 8 | Thhalv10006247m | GPX5 isoform | scaffold_19: 155838–157955 |
| 9 | Thhalv10006248m | GPX5 isoform | scaffold_19: 155838–157677 |
| 10 | Thhalv10028932m | GPX6 | scaffold_3: 3990214–3992118 |
| 11 | Thhalv10029228m | Not determined | scaffold_3: 3955295–3956113 |
| 12 | Thhalv10026852m | GPX7 | scaffold_1: 3456273–3457732 |
| 13 | Thhalv10023725m | GPX8 | scaffold_8: 965149–966667 |
| Environmental stress or hormone | Cis-acting regulatory elements | Sequence | TsGPX | AtGPX |
|---|---|---|---|---|
| Abscisic acid (ABA) | ABRE | TACGTG | TsGPX2, 4, 5, and 7 | AtGPX1, 4,5, and 8 |
| Auxin | TGA-element | AACGAC | TsGPX1 and 8 | AtGPX2 and 8 |
| AuxRR-core | GGTCCAT | TsGPX3, 4, and 5 | - | |
| Salicylic acid (SA) | TCA-element | CCATCTTTTT | TsGPX1 and 4 | AtGPX4 |
| GAGAAGAATA | TsGPX7 | AtGPX3 and 7 | ||
| Gibberellin (GA) | TATC-box | TATCCCA | TsGPX1 | - |
| GARE-motif | TCTGTTG | TsGPX2 | - | |
| P-box | GCCTTTTGAGT | TsGPX4 and 5 | AtGPX5, and 7 | |
| GARE-motif | TCTGTTG | TsGPX5 | - | |
| AAACAGA | TsGPX7 | AtGPX3, 6, and 7 | ||
| Methyl Jasmonate (MeJA) | CGTCA-motif | CGTCA | TsGPX2, 3, 4, 5, 6, 7, and 8 | AtGPX3, 4, 5, 7, and 8 |
| TGACG-motif | TGACG | TsGPX2, 3, 4, 5, 6, 7, and 8 | AtGPX3, 4, 5, 7, and 8 | |
| Ethylene | ERE | ATTTCAAA | TsGPX6 | AtGPX1, 3 |
| Drought inducibility | MBS | TAACTG | TsGPX1, 2, 3, 4, and 7 | AtGPX2, 5, 7 |
| Low-temperature | LTR | CCGAAA | TsGPX1, 2, and 5 | AtGPX5 and 8 |
| Anaerobic induction | ARE | TGGTTT | TsGPX2, 3, 6,and 8 | AtGPX2, 3, 7, and 8 |
| Name | Length (aa) | Molecular mass (kDa) | pI | TP * | Subcellular localization * | Transmembrane domain * |
|---|---|---|---|---|---|---|
| TsGPX1 | 235 | 25.902 | 9.94 | 73 | chloro | no |
| TsGPX2 | 170 | 19.020 | 5.48 | – | cyt | no |
| TsGPX3 | 196 | 23.258 | 7.33 | – | SP, CM, ER | yes |
| TsGPX4 | 170 | 19.147 | 9.08 | – | cyt | no |
| TsGPX5 | 175 | 19.612 | 9.79 | – | cyt | no |
| TsGPX6 | 234 | 25.937 | 9.35 | 58 | mito | no |
| TsGPX7 | 236 | 26.227 | 10.30 | 75 | chloro | no |
| TsGPX8 | 167 | 18.990 | 4.75 | – | cyt | no |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gao, F.; Chen, J.; Ma, T.; Li, H.; Wang, N.; Li, Z.; Zhang, Z.; Zhou, Y. The Glutathione Peroxidase Gene Family in Thellungiella salsuginea: Genome-Wide Identification, Classification, and Gene and Protein Expression Analysis under Stress Conditions. Int. J. Mol. Sci. 2014, 15, 3319-3335. https://doi.org/10.3390/ijms15023319
Gao F, Chen J, Ma T, Li H, Wang N, Li Z, Zhang Z, Zhou Y. The Glutathione Peroxidase Gene Family in Thellungiella salsuginea: Genome-Wide Identification, Classification, and Gene and Protein Expression Analysis under Stress Conditions. International Journal of Molecular Sciences. 2014; 15(2):3319-3335. https://doi.org/10.3390/ijms15023319
Chicago/Turabian StyleGao, Fei, Jing Chen, Tingting Ma, Huayun Li, Ning Wang, Zhanglei Li, Zichen Zhang, and Yijun Zhou. 2014. "The Glutathione Peroxidase Gene Family in Thellungiella salsuginea: Genome-Wide Identification, Classification, and Gene and Protein Expression Analysis under Stress Conditions" International Journal of Molecular Sciences 15, no. 2: 3319-3335. https://doi.org/10.3390/ijms15023319