Role of mtDNA Haplogroups in the Prevalence of Knee Osteoarthritis in a Southern Chinese Population


Abstract
:1. Introduction
2. Results and Discussion
2.1. Result
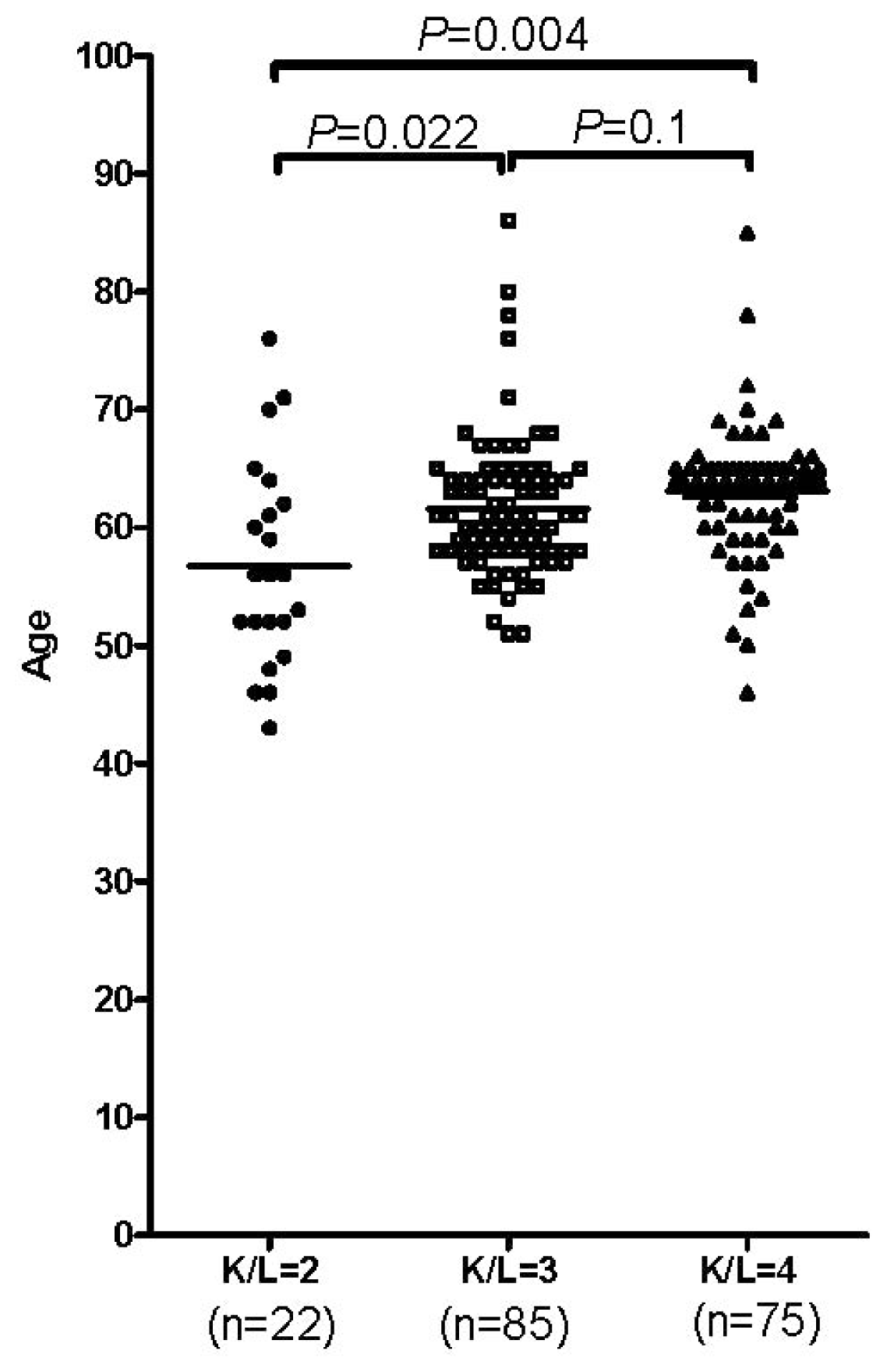
2.1.1. Age and Gender in the Occurrence of OA
2.1.2. mtDNA Haplogroup and SNPs Analysis in OA Patients
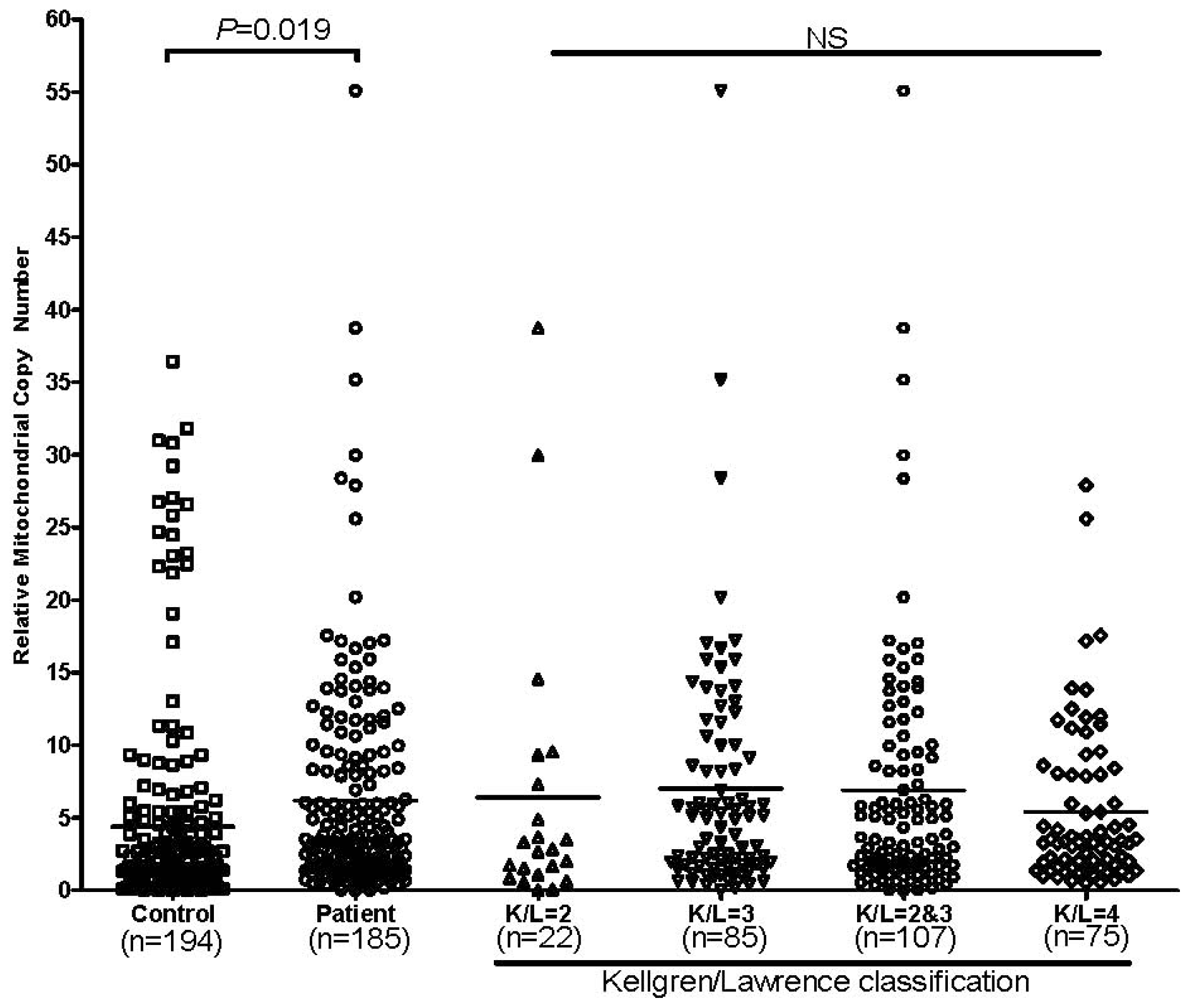
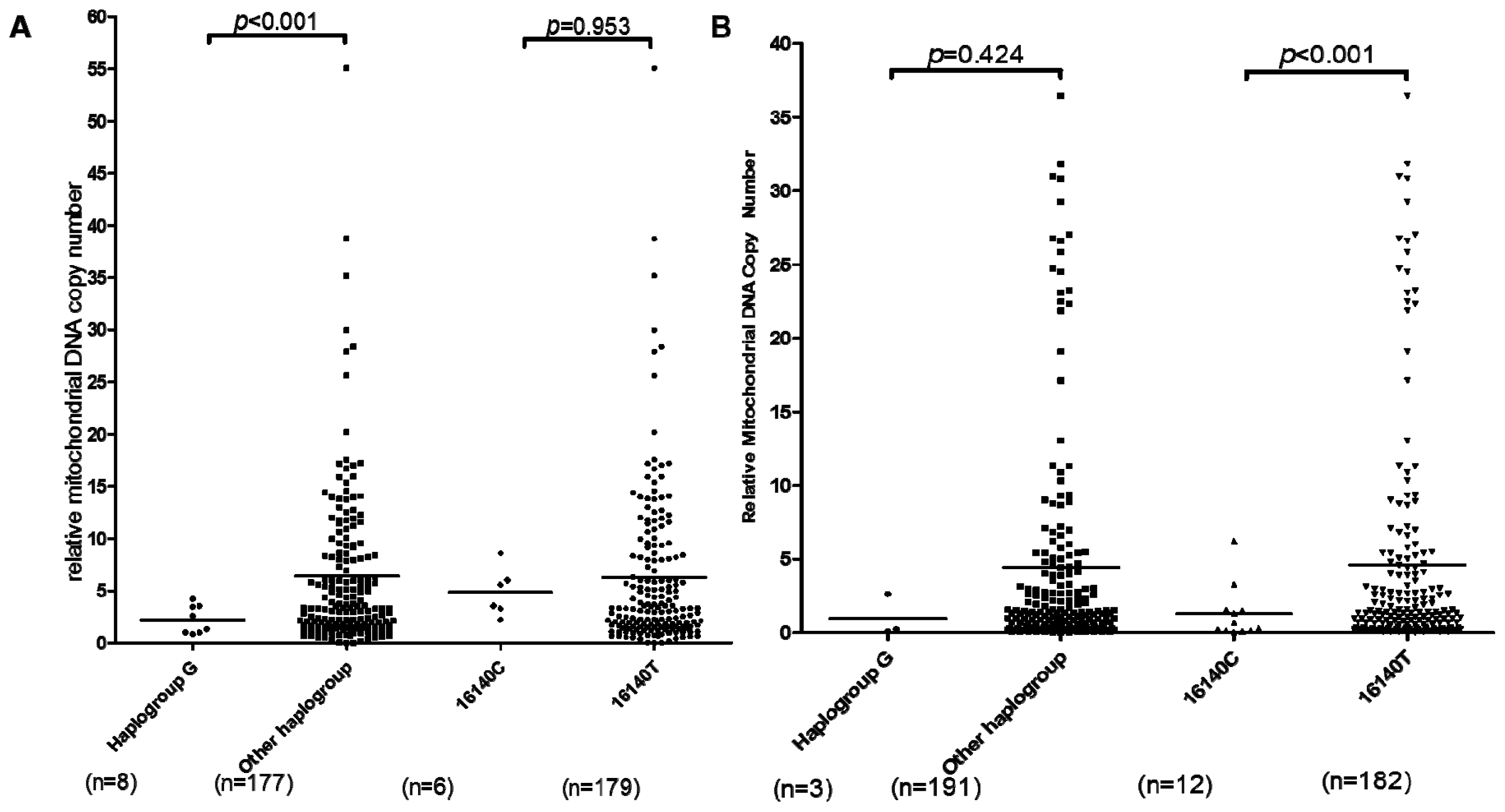
2.1.3. mtDNA Copy Number Alternation in OA Patients
2.2. Discussion
3. Experimental Section
3.1. Subjects
3.2. mtDNA Sequencing and Genotyping
3.3. mtDNA Copy Number Evaluation
3.4. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Glasson, S.S.; Askew, R.; Sheppard, B.; Carito, B.; Blanchet, T.; Ma, H.L.; Flannery, C.R.; Peluso, D.; Kanki, K.; Yang, Z.; et al. Deletion of active ADAMTS5 prevents cartilage degradation in a murine model of osteoarthritis. Nature 2005, 434, 644–648. [Google Scholar]
- Chen, S.; Zhou, Y.; Li, J.; Shan, L.Q.; Fan, Q.Y. The effect of bradykinin B2 receptor polymorphisms on the susceptibility and severity of osteoarthritis in a Chinese cohort. J. Biomed. Biotechnol 2012, 2012, 597637. [Google Scholar]
- Spector, T.D.; MacGregor, A.J. Risk factors for osteoarthritis: Genetics. Osteoarthr. Cartil 2004, 12, S39–S44. [Google Scholar]
- Blanco, F.J.; Rego, I.; Ruiz-Romero, C. The role of mitochondria in osteoarthritis. Nat. Rev. Rheumatol 2011, 7, 161–169. [Google Scholar]
- Blanco, F.J.; Lopez-Armada, M.J.; Maneiro, E. Mitochondrial dysfunction in osteoarthritis. Mitochondrion 2004, 4, 715–728. [Google Scholar]
- Lin, C.S.; Sharpley, M.S.; Fan, W.; Waymire, K.G.; Sadun, A.A.; Carelli, V.; Ross-Cisneros, F.N.; Baciu, P.; Sung, E.; McManus, M.J.; et al. Mouse mtDNA mutant model of Leber hereditary optic neuropathy. Proc. Natl. Acad. Sci. USA 2013, 109, 20065–20070. [Google Scholar]
- Sharpley, M.S.; Marciniak, C.; Eckel-Mahan, K.; McManus, M.; Crimi, M.; Waymire, K.; Lin, C.S.; Masubuchi, S.; Friend, N.; Koike, M.; et al. Heteroplasmy of mouse mtDNA is genetically unstable and results in altered behavior and cognition. Cell 2012, 151, 333–343. [Google Scholar]
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: A dawn for evolutionary medicine. Annu. Rev. Genet 2005, 39, 359–407. [Google Scholar]
- Suissa, S.; Wang, Z.; Poole, J.; Wittkopp, S.; Feder, J.; Shutt, T.E.; Wallace, D.C.; Shadel, G.S.; Mishmar, D. Ancient mtDNA genetic variants modulate mtDNA transcription and replication. PLoS Genet 2009, 5, e1000474. [Google Scholar]
- Kazuno, A.A.; Munakata, K.; Nagai, T.; Shimozono, S.; Tanaka, M.; Yoneda, M.; Kato, N.; Miyawaki, A.; Kato, T. Identification of mitochondrial DNA polymorphisms that alter mitochondrial matrix pH and intracellular calcium dynamics. PLoS Genet 2006, 2, e128. [Google Scholar]
- Pello, R.; Martin, M.A.; Carelli, V.; Nijtmans, L.G.; Achilli, A.; Pala, M.; Torroni, A.; Gomez-Duran, A.; Ruiz-Pesini, E.; Martinuzzi, A.; et al. Mitochondrial DNA background modulates the assembly kinetics of OXPHOS complexes in a cellular model of mitochondrial disease. Hum. Mol. Genet 2008, 17, 4001–4011. [Google Scholar]
- Fuku, N.; Park, K.S.; Yamada, Y.; Nishigaki, Y.; Cho, Y.M.; Matsuo, H.; Segawa, T.; Watanabe, S.; Kato, K.; Yokoi, K.; et al. Mitochondrial haplogroup N9a confers resistance against type 2 diabetes in Asians. Am. J. Hum. Genet 2007, 80, 407–415. [Google Scholar]
- Tanaka, M.; Fuku, N.; Nishigaki, Y.; Matsuo, H.; Segawa, T.; Watanabe, S.; Kato, K.; Yokoi, K.; Ito, M.; Nozawa, Y.; et al. Women with mitochondrial haplogroup N9a are protected against metabolic syndrome. Diabetes 2007, 56, 518–521. [Google Scholar]
- Ji, Y.; Zhang, A.M.; Jia, X.; Zhang, Y.P.; Xiao, X.; Li, S.; Guo, X.; Bandelt, H.J.; Zhang, Q.; Yao, Y.G. Mitochondrial DNA haplogroups M7b1’2 and M8a affect clinical expression of leber hereditary optic neuropathy in Chinese families with the m.11778G-->a mutation. Am. J. Hum. Genet 2008, 83, 760–768. [Google Scholar]
- Tanaka, N.; Goto, Y.; Akanuma, J.; Kato, M.; Kinoshita, T.; Yamashita, F.; Tanaka, M.; Asada, T. Mitochondrial DNA variants in a Japanese population of patients with Alzheimer’s disease. Mitochondrion 2010, 10, 32–37. [Google Scholar]
- Hendrickson, S.L.; Hutcheson, H.B.; Ruiz-Pesini, E.; Poole, J.C.; Lautenberger, J.; Sezgin, E.; Kingsley, L.; Goedert, J.J.; Vlahov, D.; Donfield, S.; et al. Mitochondrial DNA haplogroups influence AIDS progression. AIDS 2008, 22, 2429–2439. [Google Scholar]
- Hendrickson, S.L.; Kingsley, L.A.; Ruiz-Pesini, E.; Poole, J.C.; Jacobson, L.P.; Palella, F.J.; Bream, J.H.; Wallace, D.C.; O’Brien, S.J. Mitochondrial DNA haplogroups influence lipoatrophy after highly active antiretroviral therapy. J. Acquir. Immune Defic. Syndr 2009, 51, 111–116. [Google Scholar]
- Fang, H.; Shen, L.; Chen, T.; He, J.; Ding, Z.; Wei, J.; Qu, J.; Chen, G.; Lu, J.; Bai, Y. Cancer type-specific modulation of mitochondrial haplogroups in breast, colorectal and thyroid cancer. BMC Cancer 2010, 10, 421. [Google Scholar]
- Chatterjee, A.; Mambo, E.; Sidransky, D. Mitochondrial DNA mutations in human cancer. Oncogene 2006, 25, 4663–4674. [Google Scholar]
- Yao, Y.G.; Kong, Q.P.; Zhang, Y.P. Mitochondrial DNA 5178A polymorphism and longevity. Hum. Genet 2002, 111, 462–463. [Google Scholar]
- Zeng, Q.Y.; Chen, R.; Darmawan, J.; Xiao, Z.Y.; Chen, S.B.; Wigley, R.; Le Chen, S.; Zhang, N.Z. Rheumatic diseases in China. Arthritis Res. Ther 2008, 10, R17. [Google Scholar]
- Du, H.; Chen, S.L.; Bao, C.D.; Wang, X.D.; Lu, Y.; Gu, Y.Y.; Xu, J.R.; Chai, W.M.; Chen, J.; Nakamura, H.; et al. Prevalence and risk factors of knee osteoarthritis in Huang-Pu District, Shanghai, China. Rheumatol. Int 2005, 25, 585–590. [Google Scholar]
- Kang, X.; Fransen, M.; Zhang, Y.; Li, H.; Ke, Y.; Lu, M.; Su, S.; Song, X.; Guo, Y.; Chen, J.; et al. The high prevalence of knee osteoarthritis in a rural Chinese population: The Wuchuan osteoarthritis study. Arthritis Rheum 2009, 61, 641–647. [Google Scholar]
- Yao, Y.G.; Kong, Q.P.; Bandelt, H.J.; Kivisild, T.; Zhang, Y.P. Phylogeographic differentiation of mitochondrial DNA in Han Chinese. Am. J. Hum. Genet 2002, 70, 635–651. [Google Scholar]
- Neogi, T. The epidemiology and impact of pain in osteoarthritis. Osteoarthr. Cartil 2013, 21, 1145–1153. [Google Scholar]
- Valdes, A.M.; Spector, T.D. Genetic epidemiology of hip and knee osteoarthritis. Nat. Rev 2011, 7, 23–32. [Google Scholar]
- Cushnaghan, J.; Dieppe, P. Study of 500 patients with limb joint osteoarthritis. I. Analysis by age, sex, and distribution of symptomatic joint sites. Ann. Rheum. Dis 1991, 50, 8–13. [Google Scholar]
- Rego, I.; Fernandez-Moreno, M.; Fernandez-Lopez, C.; Gomez-Reino, J.J.; Gonzalez, A.; Arenas, J.; Blanco, F.J. Role of European mitochondrial DNA haplogroups in the prevalence of hip osteoarthritis in Galicia, Northern Spain. Ann. Rheum. Dis 2010, 69, 210–213. [Google Scholar]
- Rego-Perez, I.; Fernandez-Moreno, M.; Fernandez-Lopez, C.; Arenas, J.; Blanco, F.J. Mitochondrial DNA haplogroups: Role in the prevalence and severity of knee osteoarthritis. Arthritis Rheum 2008, 58, 2387–2396. [Google Scholar]
- Hudson, G.; Panoutsopoulou, K.; Wilson, I.; Southam, L.; Rayner, N.W.; Arden, N.; Birrell, F.; Carluke, I.; Carr, A.; Chapman, K.; et al. No evidence of an association between mitochondrial DNA variants and osteoarthritis in 7393 cases and 5122 controls. Ann. Rheum. Dis 2013, 72, 136–139. [Google Scholar]
- Zheng, S.; Qian, P.; Li, F.; Qian, G.; Wang, C.; Wu, G.; Li, Q.; Chen, Y.; Li, J.; Li, H.; et al. Association of mitochondrial DNA variations with lung cancer risk in a Han Chinese population from southwestern China. PLoS One 2012, 7, e31322. [Google Scholar]
- Wu, D.; Cheng, S.; Chen, X.; Sun, M.; Wang, G.; Fu, S.; Dong, G.; Wang, L.; Wu, Y. Mitochondrial haplogroup B4 may be a protective factor to oral lichen planus susceptibility in Chinese. Oral Dis 2013, 20, 62–68. [Google Scholar]
- Tiao, M.M.; Liou, C.W.; Huang, L.T.; Wang, P.W.; Lin, T.K.; Chen, J.B.; Chou, Y.M.; Huang, Y.H.; Lin, H.Y.; Chen, C.L.; et al. Associations of mitochondrial haplogroups b4 and e with biliary atresia and differential susceptibility to hydrophobic bile Acid. PLoS Genet 2013, 9, e1003696. [Google Scholar]
- He, W.; Newman, J.C.; Wang, M.Z.; Ho, L.; Verdin, E. Mitochondrial sirtuins: Regulators of protein acylation and metabolism. Trends Endocrinol. Metab 2012, 23, 467–476. [Google Scholar]
- Czarnecka, A.M.; Krawczyk, T.; Plak, K.; Klemba, A.; Zdrozny, M.; Arnold, R.S.; Kofler, B.; Golik, P.; Szybinska, A.; Lubinski, J.; et al. Mitochondrial genotype and breast cancer predisposition. Oncol. Rep 2010, 24, 1521–1534. [Google Scholar]
- Isashiki, Y.; Sonoda, S.; Izumo, S.; Sakamoto, T.; Tachikui, H.; Inoue, I. Phylogenetic assessment of the mitochondrial DNA displacement loop haplotype in Japanese patients with Leber’s hereditary optic neuropathy harboring the mitochondrial DNA G11778A mutation. Ophthalmic Res 2003, 35, 224–231. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Kong, Q.P.; Bandelt, H.J.; Sun, C.; Yao, Y.G.; Salas, A.; Achilli, A.; Wang, C.Y.; Zhong, L.; Zhu, C.L.; Wu, S.F.; et al. Updating the East Asian mtDNA phylogeny: A prerequisite for the identification of pathogenic mutations. Hum. Mol. Genet 2006, 15, 2076–2086. [Google Scholar]
- Eaton, J.S.; Lin, Z.P.; Sartorelli, A.C.; Bonawitz, N.D.; Shadel, G.S. Ataxia-telangiectasia mutated kinase regulates ribonucleotide reductase and mitochondrial homeostasis. J. Clin. Investig 2007, 117, 2723–2734. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Gender | Patient (n) | Control (n) | OR (95% CI) | p value |
|---|---|---|---|---|
| Male | 52 | 183 | 0.497(0.342, 0.722) | p < 0.001 * |
| Female | 135 | 237 |
| Haplogroup | Patient (n = 187) | Control (n = 420) | OR (95% CI) | p value |
|---|---|---|---|---|
| M | 103 (55.1) | 211 (50.2) | 1.215 (0.859, 1.716) | 0.270 |
| D | 40 (21.2) | 90 (21.4) | 0.998 (0.655, 1.519) | 0.992 |
| D4 | 17 (9.1) | 57 (13.6) | 0.637 (0.360, 1.128) | 0.119 |
| D4a | 4 (2.1) | 18 (4.3) | 0.488 (0.163. 1.463) | 0.191 |
| D5 | 21 (11.2) | 31 (7.4) | 1.587 (0.886, 2.844) | 0.118 |
| D5a | 5 (2.7) | 8 (1.9) | 1.415 (0.457, 4.383) | 0.546 |
| M8 | 21 (11.2) | 39 (9.3) | 1.415 (0.457, 4.383) | 0.546 |
| M8a | 7 (3.7) | 10 (2.38) | 1.594 (0.597, 4.2550) | 0.348 |
| CZ | 14 (7.5) | 28 (6.67) | 1.136 (0.584, 2.211) | 0.708 |
| M7 | 8 (4.3) | 32 (7.6) | 0.542 (0.245, 1.2) | 0.126 |
| M7b | 3 (1.6) | 20 (4.8) | 0.326 (0.096, 1.111) | 0.06 |
| M7c | 4 (2.1) | 8 (1.9) | 1.114 (0.331, 3.744) | 0.862 |
| G | 8 (4.3) | 6 (1.4) | 3.084 (1.055, 9.017) | 0.031 * |
| M9 | 4 (2.1) | 4 (1.0) | 1.180 (0.291, 4.7860) | 0.819 |
| M10 | 5 (2.6) | 8 (1.9) | 1.415 (0.457, 4.384) | 0.546 |
| N | 84 (44.9) | 209 (49.8) | 1.215 (0.859, 1.716) | 0.270 |
| A | 18 (9.6) | 35 (8.33) | 1.171 (0.645, 2.127) | 0.603 |
| N9 | 12 (6.4) | 20 (4.8) | 1.548 (0.739, 3.243) | 0.243 |
| N9a | 9 (4.8) | 14 (3.3) | 1.475 (0.627, 3.470) | 0.371 |
| Y | 3 (1.6) | 6 (1.4) | 1.125 (0.278, 4.5470) | 0.869 |
| R | 54 (28.9) | 156 (37.1) | 0.687 (0.473, 0.998) | 0.048 * |
| R11 | 4 (2.1) | 5 (1.2) | 1.814 (0.482, 6.834) | 0.372 |
| B | 19 (10.1) | 74 (17.6) | 0.529 (0.309, 0.904) | 0.018 * |
| B4 | 13 (7.0) | 52 (12.4) | 0.529 (0.280, 0.997) | 0.046 * |
| B5 | 6 (3.2) | 19 (4.5) | 0.717 (0.282, 1.8250) | 0.484 |
| R9 | 27 (14.4) | 71 (16.9) | 0.829 (0.513, 1.342) | 0.446 |
| F | 21 (11.2) | 62 (14.8) | 0.730 (0.431, 1.239) | 0.242 |
| F1 | 13 (7.0) | 38 (9.0) | 0.751 (0.390, 1.446) | 0.390 |
| F1a | 4 (2.1) | 25 (6.0) | 0.350 (0.12, 1.0) | 0.045 * |
| F1b | 6 (3.2) | 8 (1.9) | 1.707 (0.584, 4.991) | 0.323 |
| F2 | 8 (4.2) | 17 (4.0) | 1.059 (0.449, 2.500) | 0.895 |
| F2a | 4 (2.1) | 7 (1.67) | 1.290 (0.373, 4.460) | 0.687 |
| mtSNP | Patient (n = 187) | Control (n = 420) | OR (95% CI) | p value |
|---|---|---|---|---|
| A10397G | 21 (11.2) | 32 (7.6) | 1.534 (0.859, 2.739) | 0.146 |
| A10398G | 118 (63.1) | 244 (58.1) | 1.234 (0.865, 1.759) | 0.246 |
| C10400T | 103 (55.1) | 211 (50.2) | 1.215 (0.859, 1.716) | 0.270 |
| A16182C | 23 (12.3) | 52 (12.4) | 0.992 (0.588, 1.676) | 0.987 |
| T16362C | 91 (48.7) | 149 (35.5) | 1.724 (1.215, 2.445) | 0.002 * |
| G16129A | 28 (15.0) | 84 (20.0) | 0.704 (0.441, 1.124) | 0.140 |
| T16140C | 6 (3.2) | 32 (7.6) | 0.402 (0.165, 0.978) | 0.038 * |
| A16183C | 54 (28.9) | 104 (24.8) | 1.263 (0.859, 1.857) | 0.234 |
| T16189C | 64 (34.2) | 158 (37.6) | 0.863 (0.601, 1.238) | 0.423 |
| T16217C | 13 (7.0) | 52 (12.4) | 0.529 (0.280, 0.997) | 0.046 * |
| C16223T | 126 (67.4) | 250 (59.5) | 1.405 (0.977, 2.018) | 0.066 |
| T16298C | 25 (13.4) | 45 (10.7) | 1.286 (0.763, 2.168) | 0.344 |
| T16304C | 25 (13.4) | 74 (17.6) | 0.722 (0.442, 1.178) | 0.191 |
| T16311C | 24 (12.8) | 74 (17.6) | 0.688 (0.419, 1.131) | 0.139 |
| G16319A | 30 (16.0) | 54 (12.9) | 1.338 (0.828, 2.162) | 0.233 |
| T146C | 26 (13.9) | 64 (15.2) | 0.898 (0.549, 1.470) | 0.669 |
| C150T | 40 (21.3) | 88 (21.0) | 1.027 (0.674, 1.564) | 0.903 |
| T152C | 59 (31.6) | 103 (24.5) | 1.419 (0.970, 2.075) | 0.071 |
| 249del | 34 (18.2) | 95 (22.6) | 0.760 (0.491, 1.176) | 0.760 |
| T16172C | 16 (8.6) | 49 (11.7) | 0.708 (0.392, 1.281) | 0.253 |
| Haplogroup/SNPs | K/L = 2, 3 (n = 108) | K/L = 4 (n = 76) | OR (95% CI) | p value |
|---|---|---|---|---|
| M | 57 (52.8) | 45 (59.2) | 1.299 (0.717, 2.353) | 0.387 |
| D | 21 (19.4) | 19 (25.0) | 1.381 (0.683, 2.793) | 0.368 |
| D4 | 6 (5.6) | 11 (14.5) | 2.874 (1.014, 8.130) | 0.04 * |
| D4a | 0 (0) | 4 (5.3) | 2.500 (2.092, 2.994) | 0.016 * |
| D5 | 14 (13.0) | 7 (9.2) | 0.681 (0.261, 1.776) | 0.431 |
| D5a | 1 (0.9) | 4 (5.3) | 5.952 (0.651, 55.556) | 0.075 |
| M8 | 14 (13.0) | 7 (9.2) | 0.681 (0.261, 1.776) | 0.431 |
| CZ | 8 (7.4) | 6 (7.9) | 1.072 (0.356, 3.226) | 0.902 |
| M8a | 6 (5.6) | 1 (1.3) | 0.227 (0.027, 1.923) | 0.139 |
| M7 | 4 (3.7) | 4 (5.3) | 1.445 (0.350, 5.952) | 0.610 |
| G | 1 (0.9) | 7 (9.2) | 10.870 (1.307, 90.909) | 0.007 * |
| M9 | 3 (2.8) | 1 (1.3) | 0.467 (0.048, 4.566) | 0.503 |
| M10 | 4 (3.7) | 1 (1.3) | 0.347 (0.038, 3.165) | 0.327 |
| N | 51 (47.2) | 31 (40.8) | 1.299 (0.717, 2.353) | 0.387 |
| A | 12 (11.1) | 6 (7.9) | 0.686 (0.245, 1.916) | 0.470 |
| N9 | 11 (10.2) | 1 (1.3) | 0.118 (0.015, 0.931) | 0.016 * |
| N9a | 9 (8.3) | 0 (0) | 0.566 (0.497, 0.644) | 0.01 * |
| Y | 2 (1.9) | 1 (1.3) | 0.707 (0.063, 7.937) | 0.777 |
| R | 32 (29.7) | 22 (28.9) | 0.968 (0.508, 1.845) | 0.920 |
| R11 | 2 (1.9) | 2 (2.6) | 1.433 (0.197, 10.417) | 0.721 |
| B | 12 (11.1) | 7 (9.2) | 0.812 (0.304, 2.169) | 0.677 |
| B4 | 8 (7.4) | 5 (6.6) | 0.880 (0.277, 2.801) | 0.829 |
| B5 | 4 (3.7) | 2 (2.6) | 0.703 (0.125, 3.937) | 0.687 |
| R9 | 16 (14.8) | 11 (14.5) | 0.973 (0.424, 2.232) | 0.949 |
| F | 12 (11.1) | 9 (11.9) | 1.074 (0.429, 2.695) | 0.878 |
| F1 | 6 (5.6) | 7 (9.2) | 1.724 (0.556, 5.348) | 0.341 |
| F1a | 3 (2.8) | 1 (1.3) | 0.227 (0.023, 2.227) | 0.166 |
| F1b | 4 (3.7) | 2 (2.6) | 0.703 (0.125, 3.937) | 0.687 |
| F2 | 6 (5.6) | 2 (2.6) | 0.460 (0.090, 2.342) | 0.338 |
| F2a | 1 (0.9) | 3 (3.9) | 0.467 (0.048, 4.566) | 0.503 |
| mtSNP | K/L = 2, 3 (n = 108) | K/L = 4 (n = 76) | OR (95% CI) | p value |
|---|---|---|---|---|
| A10397G | 14 (13.0) | 7 (9.2) | 0.681 (0.261, 1.776) | 0.431 |
| A10398G | 68 (63.0) | 49 (64.5) | 1.067 (0.579, 1.965) | 0.843 |
| C10400T | 57 (52.8) | 45 (59.2) | 1.299 (0.717, 2.353) | 0.387 |
| A16182C | 12 (11.1) | 11 (14.5) | 1.353 (0.563, 0.563) | 0.497 |
| T16362C | 46 (42.6) | 42 (55.3) | 1.664 (0.922, 3.012) | 0.09 |
| G16129A | 15 (13.8) | 13 (17.1) | 1.279 (0.570, 2.874) | 0.550 |
| T16140C | 5 (4.6) | 1 (1.3) | 0.275 (0.031, 2.398) | 0.213 |
| A16183C | 33 (30.6) | 21 (27.7) | 0.868 (0.454, 1.658) | 0.668 |
| T16189C | 38 (35.2) | 26 (34.2) | 0.958 (0.517, 1.776) | 0.891 |
| T16217C | 8 (7.4) | 5 (6.6) | 0.880 (0.276, 2.801) | 0.829 |
| C16223T | 72 (66.7) | 51 (67.1) | 1.020 (0.547, 1.905) | 0.950 |
| T16298C | 18 (16.7) | 7 (9.2) | 0.507 (0.201, 1.282) | 0.146 |
| T16304C | 12 (11.1) | 13 (17.1) | 1.650 (0.708, 3.846) | 0.243 |
| T16311C | 13 (12.0) | 12 (15.8) | 1.370 (0.588, 3.195) | 0.465 |
| G16319A | 18 (16.7) | 10 (13.2) | 0.758 (0.328, 1.748) | 0.514 |
| T146C | 19 (17.6) | 7 (9.2) | 0.475 (0.189, 1.195) | 0.108 |
| C150T | 29 (26.9) | 12 (15.8) | 0.511 (0.241, 1.08) | 0.076 |
| T152C | 31 (28.8) | 25 (32.9) | 1.218 (0.646, 2.299) | 0.543 |
| 249del | 18 (16.7) | 13 (17.1) | 1.032 (0.472, 2.257) | 0.938 |
| T16172C | 6 (5.6) | 10 (13.2) | 2.577 (0.894, 7.407) | 0.072 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fang, H.; Liu, X.; Shen, L.; Li, F.; Liu, Y.; Chi, H.; Miao, H.; Lu, J.; Bai, Y. Role of mtDNA Haplogroups in the Prevalence of Knee Osteoarthritis in a Southern Chinese Population. Int. J. Mol. Sci. 2014, 15, 2646-2659. https://doi.org/10.3390/ijms15022646
Fang H, Liu X, Shen L, Li F, Liu Y, Chi H, Miao H, Lu J, Bai Y. Role of mtDNA Haplogroups in the Prevalence of Knee Osteoarthritis in a Southern Chinese Population. International Journal of Molecular Sciences. 2014; 15(2):2646-2659. https://doi.org/10.3390/ijms15022646
Chicago/Turabian StyleFang, Hezhi, Xinwei Liu, Lijun Shen, Fengjie Li, Yihong Liu, Hongbo Chi, Huikai Miao, Jianxin Lu, and Yidong Bai. 2014. "Role of mtDNA Haplogroups in the Prevalence of Knee Osteoarthritis in a Southern Chinese Population" International Journal of Molecular Sciences 15, no. 2: 2646-2659. https://doi.org/10.3390/ijms15022646