The Effect of Radiation on the Immune Response to Cancers

{kind=link}
Abstract
:1. Introduction
2. Effects of Radiation on the Endogenous Immune System
2.1. Systemic Antitumor Immune Response (Abscopal Effect)
2.2. Tumor Antigens and Antigen Presentation
2.2.1. Cytotoxic T Lymphocytes and Dendritic Cells
2.2.2. MHC Class I and II
2.3. Immune Modulators; HMGB-1 and TLRs
2.4. Regulatory T cells
2.5. Clinical Implications of Concomitant Immunity
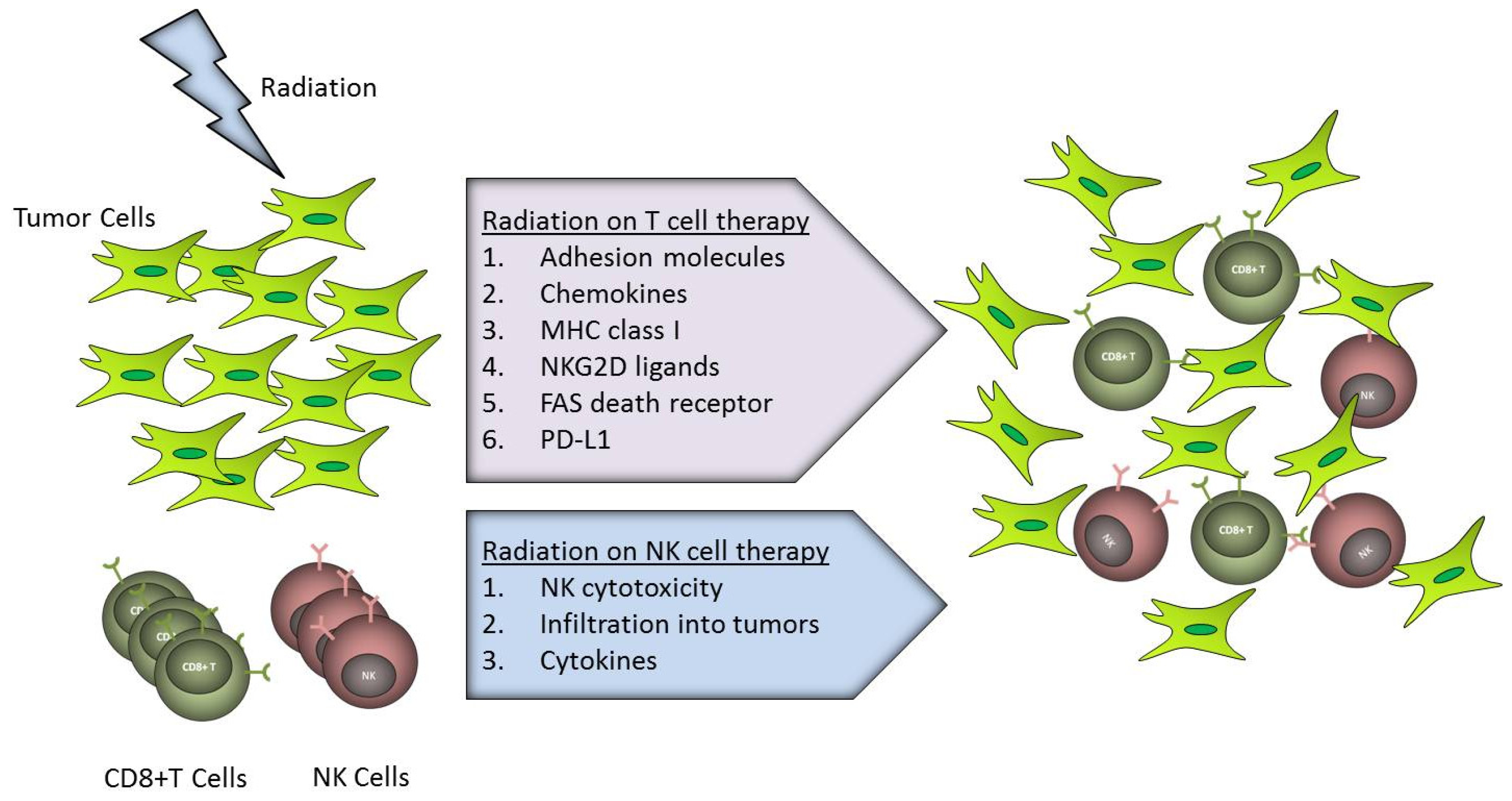
3. Effects of Radiation on Transferred Immune System
3.1. Adoptive T Cell Therapy
3.1.1. Lymphocyte Trafficking
3.1.2. Tumor Recognition and Killing
3.2. Adoptive Natural Killer (NK) Cell Therapy
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Aoyama, H.; Shirato, H.; Kakuto, Y.; Inakoshi, H.; Nishio, M.; Yoshida, H.; Hareyama, M.; Yanagisawa, T.; Watarai, J.; Miyasaka, K. Pathologically-proven intracranial germinoma treated with radiation therapy. Radiother. Oncol 1998, 47, 201–205. [Google Scholar]
- Ross, G.M. Induction of cell death by radiotherapy. Endocr.-Relat. Cancer 1999, 6, 41–44. [Google Scholar]
- Nam, H.Y.; Han, M.W.; Chang, H.W.; Kim, S.Y.; Kim, S.W. Prolonged autophagy by MTOR inhibitor leads radioresistant cancer cells into senescence. Autophagy 2013, 9, 1631–1632. [Google Scholar]
- Nam, H.Y.; Han, M.W.; Chang, H.W.; Lee, Y.S.; Lee, M.; Lee, H.J.; Lee, B.W.; Lee, K.E.; Jung, M.K.; Jeon, H.; et al. Radioresistant cancer cells can be conditioned to enter senescence by mTOR inhibition. Cancer Res 2013, 73, 4267–4277. [Google Scholar]
- Truman, J.P.; Gueven, N.; Lavin, M.; Leibel, S.; Kolesnick, R.; Fuks, Z.; Haimovitz-Friedman, A. Down-regulation of ATM protein sensitizes human prostate cancer cells to radiation-induced apoptosis. J. Biol. Chem 2005, 280, 23262–23272. [Google Scholar]
- Eriksson, D.; Stigbrand, T. Radiation-induced cell death mechanisms. Tumour Biol 2010, 31, 363–372. [Google Scholar]
- Verheij, M.; Bartelink, H. Radiation-induced apoptosis. Cell Tissue Res 2000, 301, 133–142. [Google Scholar]
- Ohba, K.; Omagari, K.; Nakamura, T.; Ikuno, N.; Saeki, S.; Matsuo, I.; Kinoshita, H.; Masuda, J.; Hazama, H.; Sakamoto, I.; et al. Abscopal regression of hepatocellular carcinoma after radiotherapy for bone metastasis. Gut 1998, 43, 575–577. [Google Scholar]
- Burnette, B.C.; Liang, H.; Lee, Y.; Chlewicki, L.; Khodarev, N.N.; Weichselbaum, R.R.; Fu, Y.X.; Auh, S.L. The efficacy of radiotherapy relies upon induction of type I interferon-dependent innate and adaptive immunity. Cancer Res 2011, 71, 2488–2496. [Google Scholar]
- Shiraishi, K.; Ishiwata, Y.; Nakagawa, K.; Yokochi, S.; Taruki, C.; Akuta, T.; Ohtomo, K.; Matsushima, K.; Tamatani, T.; Kanegasaki, S. Enhancement of antitumor radiation efficacy and consistent induction of the abscopal effect in mice by ECI301, an active variant of macrophage inflammatory protein-1alpha. Clin. Cancer Res 2008, 14, 1159–1166. [Google Scholar]
- Demaria, S.; Kawashima, N.; Yang, A.M.; Devitt, M.L.; Babb, J.S.; Allison, J.P.; Formenti, S.C. Immune-mediated inhibition of metastases after treatment with local radiation and CTLA-4 blockade in a mouse model of breast cancer. Clin. Cancer Res 2005, 11, 728–734. [Google Scholar]
- Demaria, S.; Pilones, K.A.; Formenti, S.C.; Dustin, M.L. Exploiting the stress response to radiation to sensitize poorly immunogenic tumors to anti-CTLA-4 treatment. Oncoimmunology 2013, 2, e23127. [Google Scholar]
- Postow, M.A.; Callahan, M.K.; Barker, C.A.; Yamada, Y.; Yuan, J.; Kitano, S.; Mu, Z.; Rasalan, T.; Adamow, M.; Ritter, E.; et al. Immunologic correlates of the abscopal effect in a patient with melanoma. N. Engl. J. Med 2012, 366, 925–931. [Google Scholar]
- Meng, Y.; Efimova, E.V.; Hamzeh, K.W.; Darga, T.E.; Mauceri, H.J.; Fu, Y.X.; Kron, S.J.; Weichselbaum, R.R. Radiation-inducible immunotherapy for cancer: Senescent tumor cells as a cancer vaccine. Mol. Ther 2012, 20, 1046–1055. [Google Scholar]
- Lee, Y.; Auh, S.L.; Wang, Y.; Burnette, B.; Meng, Y.; Beckett, M.; Sharma, R.; Chin, R.; Tu, T.; Weichselbaum, R.R.; et al. Therapeutic effects of ablative radiation on local tumor require CD8+ T cells: Changing strategies for cancer treatment. Blood 2009, 114, 589–595. [Google Scholar]
- Liao, Y.P.; Wang, C.C.; Butterfield, L.H.; Economou, J.S.; Ribas, A.; Meng, W.S.; Iwamoto, K.S.; McBride, W.H. Ionizing radiation affects human MART-1 melanoma antigen processing and presentation by dendritic cells. J. Immunol 2004, 173, 2462–2469. [Google Scholar]
- Reits, E.A.; Hodge, J.W.; Herberts, C.A.; Groothuis, T.A.; Chakraborty, M.; Wansley, E.K.; Camphausen, K.; Luiten, R.M.; de Ru, A.H.; Neijssen, J.; et al. Radiation modulates the peptide repertoire, enhances MHC class I expression, and induces successful antitumor immunotherapy. J. Exp. Med 2006, 203, 1259–1271. [Google Scholar]
- Sharma, A.; Bode, B.; Wenger, R.H.; Lehmann, K.; Sartori, A.A.; Moch, H.; Knuth, A.; Boehmer, L.; Broek, M. γ-Radiation promotes immunological recognition of cancer cells through increased expression of cancer-testis antigens in vitro and in vivo. PLoS One 2011, 6, e28217. [Google Scholar]
- Newcomb, E.W.; Demaria, S.; Lukyanov, Y.; Shao, Y.; Schnee, T.; Kawashima, N.; Lan, L.; Dewyngaert, J.K.; Zagzag, D.; McBride, W.H.; et al. The combination of ionizing radiation and peripheral vaccination produces long-term survival of mice bearing established invasive GL261 gliomas. Clin. Cancer Res 2006, 12, 4730–4737. [Google Scholar]
- Chiriva-Internati, M.; Grizzi, F.; Pinkston, J.; Morrow, K.J.; D’Cunha, N.; Frezza, E.E.; Muzzio, P.C.; Kast, W.M.; Cobos, E. Gamma-radiation upregulates MHC class I/II and ICAM-I molecules in multiple myeloma cell lines and primary tumors. In vitro Cell. Dev. Biol. Anim 2006, 42, 89–95. [Google Scholar]
- Apetoh, L.; Ghiringhelli, F.; Tesniere, A.; Obeid, M.; Ortiz, C.; Criollo, A.; Mignot, G.; Maiuri, M.C.; Ullrich, E.; Saulnier, P.; et al. Toll-like receptor 4-dependent contribution of the immune system to anticancer chemotherapy and radiotherapy. Nat. Med 2007, 13, 1050–1059. [Google Scholar]
- Suzuki, Y.; Mimura, K.; Yoshimoto, Y.; Watanabe, M.; Ohkubo, Y.; Izawa, S.; Murata, K.; Fujii, H.; Nakano, T.; Kono, K. Immunogenic tumor cell death induced by chemoradiotherapy in patients with esophageal squamous cell carcinoma. Cancer Res 2012, 72, 3967–3976. [Google Scholar]
- Turk, M.J.; Guevara-Patino, J.A.; Rizzuto, G.A.; Engelhorn, M.E.; Sakaguchi, S.; Houghton, A.N. Concomitant tumor immunity to a poorly immunogenic melanoma is prevented by regulatory T cells. J. Exp. Med 2004, 200, 771–782. [Google Scholar]
- Qu, Y.; Jin, S.; Zhang, A.; Zhang, B.; Shi, X.; Wang, J.; Zhao, Y. Gamma-ray resistance of regulatory CD4+CD25+Foxp3+ T cells in mice. Radiat. Res 2010, 173, 148–157. [Google Scholar]
- Kachikwu, E.L.; Iwamoto, K.S.; Liao, Y.P.; DeMarco, J.J.; Agazaryan, N.; Economou, J.S.; McBride, W.H.; Schaue, D. Radiation enhances regulatory T cell representation. Int. J. Radiat. Oncol. Biol. Phys 2011, 81, 1128–1135. [Google Scholar]
- Schaue, D.; Ratikan, J.A.; Iwamoto, K.S.; McBride, W.H. Maximizing tumor immunity with fractionated radiation. Int. J. Radiat. Oncol. Biol. Phys 2012, 83, 1306–1310. [Google Scholar]
- Mandapathil, M.; Szczepanski, M.J.; Szajnik, M.; Ren, J.; Lenzner, D.E.; Jackson, E.K.; Gorelik, E.; Lang, S.; Johnson, J.T.; Whiteside, T.L. Increased ectonucleotidase expression and activity in regulatory T cells of patients with head and neck cancer. Clin. Cancer Res 2009, 15, 6348–6357. [Google Scholar]
- Chen, W.; Jin, W.; Hardegen, N.; Lei, K.J.; Li, L.; Marinos, N.; McGrady, G.; Wahl, S.M. Conversion of peripheral CD4+CD25− naive T cells to CD4+CD25+ regulatory T cells by TGF-β induction of transcription factor Foxp3. J. Exp. Med 2003, 198, 1875–1886. [Google Scholar]
- Antoniades, J.; Brady, L.W.; Lightfoot, D.A. Lymphangiographic demonstration of the abscopal effect in patients with malignant lymphomas. Int. J. Radiat. Oncol. Biol. Phys 1977, 2, 141–147. [Google Scholar]
- Isobe, Y.; Aritaka, N.; Sasaki, M.; Oshimi, K.; Sugimoto, K. Spontaneous regression of natural killer cell lymphoma. J. Clin. Pathol 2009, 62, 647–650. [Google Scholar]
- Lakshmanagowda, P.B.; Viswanath, L.; Thimmaiah, N.; Dasappa, L.; Supe, S.S.; Kallur, P. Abscopal effect in a patient with chronic lymphocytic leukemia during radiation therapy: A case report. Cases J 2009, 2, 204. [Google Scholar]
- Cotter, S.E.; Dunn, G.P.; Collins, K.M.; Sahni, D.; Zukotynski, K.A.; Hansen, J.L.; O’Farrell, D.A.; Ng, A.K.; Devlin, P.M.; Wang, L.C. Abscopal effect in a patient with metastatic Merkel cell carcinoma following radiation therapy: Potential role of induced antitumor immunity. Arch. Dermatol 2011, 147, 870–872. [Google Scholar]
- Takaya, M.; Niibe, Y.; Tsunoda, S.; Jobo, T.; Imai, M.; Kotani, S.; Unno, N.; Hayakawa, K. Abscopal effect of radiation on toruliform para-aortic lymph node metastases of advanced uterine cervical carcinoma—A case report. Anticancer Res 2007, 27, 499–503. [Google Scholar]
- Okuma, K.; Yamashita, H.; Niibe, Y.; Hayakawa, K.; Nakagawa, K. Abscopal effect of radiation on lung metastases of hepatocellular carcinoma: A case report. J. Med. Case Rep 2011, 5, 111. [Google Scholar]
- Niibe, Y.; Chang, J.Y. Novel insights of oligometastases and oligo-recurrence and review of the literature. Pulm. Med 2012, 2012, 261096. [Google Scholar]
- Niibe, Y.; Hayakawa, K. Oligometastases and oligo-recurrence: The new era of cancer therapy. Jpn. J. Clin. Oncol 2010, 40, 107–111. [Google Scholar]
- Niibe, Y.; Nishimura, T.; Inoue, T.; Karasawa, K.; Shioyama, Y.; Ogawa, Y.; Takemoto, M.; Shirato, H. Oligometastases of brain only in patients with Non-Small Cell Lung Cancer (NSCLC) treated with Stereotactic Irradiation (STI): A multi-institutional study in Japan. Int. J. Radiat. Oncol 2010, 78, S497–S497. [Google Scholar]
- Corbin, K.S.; Hellman, S.; Weichselbaum, R.R. Extracranial oligometastases: A subset of metastases curable with stereotactic radiotherapy. J. Clin. Oncol 2013, 31, 1384–1390. [Google Scholar]
- Masucci, G.V.; Wersall, P.; Kiessling, R.; Lundqvist, A.; Lewensohn, R. Stereotactic Ablative Radio Therapy (SABR) followed by immunotherapy a challenge for individualized treatment of metastatic solid tumours. J. Transl. Med 2012, 10, 104. [Google Scholar]
- Seung, S.K.; Curti, B.D.; Crittenden, M.; Walker, E.; Coffey, T.; Siebert, J.C.; Miller, W.; Payne, R.; Glenn, L.; Bageac, A.; Urba, W.J. Phase 1 study of stereotactic body radiotherapy and interleukin-2—Tumor and immunological responses. Sci. Transl. Med 2012, 4. [Google Scholar] [CrossRef]
- Lugade, A.A.; Moran, J.P.; Gerber, S.A.; Rose, R.C.; Frelinger, J.G.; Lord, E.M. Local radiation therapy of B16 melanoma tumors increases the generation of tumor antigen-specific effector cells that traffic to the tumor. J. Immunol 2005, 174, 7516–7523. [Google Scholar]
- Cao, Z.A.; Daniel, D.; Hanahan, D. Sub-lethal radiation enhances anti-tumor immunotherapy in a transgenic mouse model of pancreatic cancer. BMC Cancer 2002, 2, 11. [Google Scholar] [Green Version]
- Lugade, A.A.; Sorensen, E.W.; Gerber, S.A.; Moran, J.P.; Frelinger, J.G.; Lord, E.M. Radiation-induced IFN-gamma production within the tumor microenvironment influences antitumor immunity. J. Immunol 2008, 180, 3132–3139. [Google Scholar]
- Hareyama, M.; Imai, K.; Oouchi, A.; Takahashi, H.; Hinoda, Y.; Tsujisaki, M.; Adachi, M.; Shonai, T.; Sakata, K.; Morita, K. The effect of radiation on the expression of intercellular adhesion molecule-1 of human adenocarcinoma cells. Int. J. Radiat. Oncol. Biol. Phys 1998, 40, 691–696. [Google Scholar]
- Matsumura, S.; Wang, B.; Kawashima, N.; Braunstein, S.; Badura, M.; Cameron, T.O.; Babb, J.S.; Schneider, R.J.; Formenti, S.C.; Dustin, M.L.; et al. Radiation-induced CXCL16 release by breast cancer cells attracts effector T cells. J. Immunol 2008, 181, 3099–3107. [Google Scholar]
- Draghiciu, O.; Walczak, M.; Hoogeboom, B.N.; Franken, K.L.; Melief, K.J.; Nijman, H.W.; Daemen, T. Therapeutic immunization and local low-dose tumor irradiation, a reinforcing combination. Int. J. Cancer 2013, 134, 859–872. [Google Scholar]
- Gasser, S.; Orsulic, S.; Brown, E.J.; Raulet, D.H. The DNA damage pathway regulates innate immune system ligands of the NKG2D receptor. Nature 2005, 436, 1186–1190. [Google Scholar]
- Chakraborty, M.; Abrams, S.I.; Camphausen, K.; Liu, K.; Scott, T.; Coleman, C.N.; Hodge, J.W. Irradiation of tumor cells up-regulates Fas and enhances CTL lytic activity and CTL adoptive immunotherapy. J. Immunol 2003, 170, 6338–6347. [Google Scholar]
- Garnett, C.T.; Palena, C.; Chakraborty, M.; Tsang, K.Y.; Schlom, J.; Hodge, J.W. Sublethal irradiation of human tumor cells modulates phenotype resulting in enhanced killing by cytotoxic T lymphocytes. Cancer Res 2004, 64, 7985–7994. [Google Scholar]
- Chakraborty, M.; Abrams, S.I.; Coleman, C.N.; Camphausen, K.; Schlom, J.; Hodge, J.W. External beam radiation of tumors alters phenotype of tumor cells to render them susceptible to vaccine-mediated T-cell killing. Cancer Res 2004, 64, 4328–4337. [Google Scholar]
- Liang, H.; Deng, L.; Chmura, S.; Burnette, B.; Liadis, N.; Darga, T.; Beckett, M.A.; Lingen, M.W.; Witt, M.; Weichselbaum, R.R.; et al. Radiation-induced equilibrium is a balance between tumor cell proliferation and T cell-mediated killing. J. Immunol 2013, 190, 5874–5881. [Google Scholar]
- Caligiuri, M.A. Human natural killer cells. Blood 2008, 112, 461–469. [Google Scholar]
- Ljunggren, H.G.; Malmberg, K.J. Prospects for the use of NK cells in immunotherapy of human cancer. Nat. Rev. Immunol 2007, 7, 329–339. [Google Scholar]
- Watzl, C.; Long, E.O. Exposing tumor cells to killer cell attack. Nat. Med 2000, 6, 867–868. [Google Scholar]
- Pardoll, D.M. Immunology. Stress NK receptors, and immune surveillance. Science 2001, 294, 534–536. [Google Scholar]
- Zarybnicka, L.; Vavrova, J.; Havelek, R.; Tichy, A.; Pejchal, J.; Sinkorova, Z. Lymphocyte subsets and their H2AX phosphorylation in response to in vivo irradiation in rats. Int. J. Radiat. Biol 2013, 89, 110–117. [Google Scholar]
- Zarcone, D.; Tilden, A.B.; Lane, V.G.; Grossi, C.E. Radiation sensitivity of resting and activated nonspecific cytotoxic cells of T lineage and NK lineage. Blood 1989, 73, 1615–1621. [Google Scholar]
- Mizutani, Y.; Uchida, A.; Fujimoto, T.; Ikenaga, M.; Yoshida, O. Enhancement by X-ray irradiation of target cell susceptibility to natural killer cells. Immunol. Lett 1989, 22, 247–251. [Google Scholar]
- Uchida, A.; Mizutani, Y.; Nagamuta, M.; Ikenaga, M. Effects of X-ray irradiation on natural killer (NK) cell system. I. Elevation of sensitivity of tumor cells and lytic function of NK cells. Immunopharmacol. Immunotoxicol 1989, 11, 507–519. [Google Scholar]
- Halpern, J.N.; Cohen, E.; Mekori, T.; Beny, A.N.; Kuten, A.; Rosenblatt, E.A.; Robinson, E. The effect of in vitro irradiation on human NK cell activity. A preliminary report. J. Med 1991, 22, 243–254. [Google Scholar]
- Shin, S.C.; Lee, K.M.; Kang, Y.M.; Kim, K.; Kim, C.S.; Yang, K.H.; Jin, Y.W.; Kim, H.S. Alteration of cytokine profiles in mice exposed to chronic low-dose ionizing radiation. Biochem. Biophys. Res. Commun 2010, 397, 644–649. [Google Scholar]
- Sonn, C.H.; Choi, J.R.; Kim, T.J.; Yu, Y.B.; Kim, K.; Shin, S.C.; Park, G.H.; Shirakawa, T.; Kim, H.S.; Lee, K.M. Augmentation of natural cytotoxicity by chronic low-dose ionizing radiation in murine natural killer cells primed by IL-2. J. Radiat. Res 2012, 53, 823–829. [Google Scholar]
- Nakagawa, K.; Yoshida, F.; Omori, N.; Tsunoda, T.; Nose, T. The effect of radiation therapy combined with natural killer cells against spontaneous murine fibrosarcoma. Biotherapy 1990, 2, 69–75. [Google Scholar]
- Nowosielska, E.M.; Cheda, A.; Wrembel-Wargocka, J.; Janiak, M.K. Effect of low doses of low-let radiation on the innate anti-tumor reactions in radioresistant and radiosensitive mice. Dose Response 2012, 10, 500–515. [Google Scholar]
- Yang, K.L.; Wang, Y.S.; Chang, C.C.; Huang, S.C.; Huang, Y.C.; Chi, M.S.; Chi, K.H. Reciprocal complementation of the tumoricidal effects of radiation and natural killer cells. PLoS One 2013, 8, e61797. [Google Scholar]
- Begovic, M.; Herberman, R.; Gorelik, E. Increase in immunogenicity and sensitivity to natural cell-mediated cytotoxicity following in vitro exposure of MCA105 tumor cells to ultraviolet radiation. Cancer Res 1991, 51, 5153–5159. [Google Scholar]
- Begovic, M.; Herberman, R.B.; Gorelik, E. Ultraviolet light-induced increase in tumor cell susceptibility to TNF-dependent and TNF-independent natural cell-mediated cytotoxicity. Cell. Immunol 1991, 138, 349–359. [Google Scholar]
- Begovic, M.; Herberman, R.B.; Gorelik, E. Effect of UV light on tumor cell sensitivity to NK and NC cell-mediated lysis. Nat. Immun 1993, 12, 250–266. [Google Scholar]
- Liu, L.L.; Smith, M.J.; Sun, B.S.; Wang, G.J.; Redmond, H.P.; Wang, J.H. Combined IFN-gamma-endostatin gene therapy and radiotherapy attenuates primary breast tumor growth and lung metastases via enhanced CTL and NK cell activation and attenuated tumor angiogenesis in a murine model. Ann. Surg. Oncol 2009, 16, 1403–1411. [Google Scholar]
- Ni, J.; Miller, M.; Stojanovic, A.; Garbi, N.; Cerwenka, A. Sustained effector function of IL-12/15/18-preactivated NK cells against established tumors. J. Exp. Med 2012, 209, 2351–2365. [Google Scholar]
- Lim, S.A.; Kim, T.J.; Lee, J.E.; Sonn, C.H.; Kim, K.; Kim, J.; Choi, J.G.; Choi, I.K.; Yun, C.O.; Kim, J.H.; et al. Ex vivo expansion of highly cytotoxic human NK cells by cocultivation with irradiated tumor cells for adoptive immunotherapy. Cancer Res 2013, 73, 2598–2607. [Google Scholar]
- Fujisaki, H.; Kakuda, H.; Shimasaki, N.; Imai, C.; Ma, J.; Lockey, T.; Eldridge, P.; Leung, W.H.; Campana, D. Expansion of highly cytotoxic human natural killer cells for cancer cell therapy. Cancer Res 2009, 69, 4010–4017. [Google Scholar]
- Denman, C.J.; Senyukov, V.V.; Somanchi, S.S.; Phatarpekar, P.V.; Kopp, L.M.; Johnson, J.L.; Singh, H.; Hurton, L.; Maiti, S.N.; Huls, M.H.; et al. Membrane-bound IL-21 promotes sustained ex vivo proliferation of human natural killer cells. PLoS One 2012, 7, e30264. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Park, B.; Yee, C.; Lee, K.-M. The Effect of Radiation on the Immune Response to Cancers. Int. J. Mol. Sci. 2014, 15, 927-943. https://doi.org/10.3390/ijms15010927
Park B, Yee C, Lee K-M. The Effect of Radiation on the Immune Response to Cancers. International Journal of Molecular Sciences. 2014; 15(1):927-943. https://doi.org/10.3390/ijms15010927
Chicago/Turabian StylePark, Bonggoo, Cassian Yee, and Kyung-Mi Lee. 2014. "The Effect of Radiation on the Immune Response to Cancers" International Journal of Molecular Sciences 15, no. 1: 927-943. https://doi.org/10.3390/ijms15010927