Increased Preventive Effect on Colon Carcinogenesis by Use of Resistant Starch (RS3) as the Carrier for Polysaccharide of Larimichthys Crocea Swimming Bladder

Abstract
:1. Introduction
2. Results and Discussion
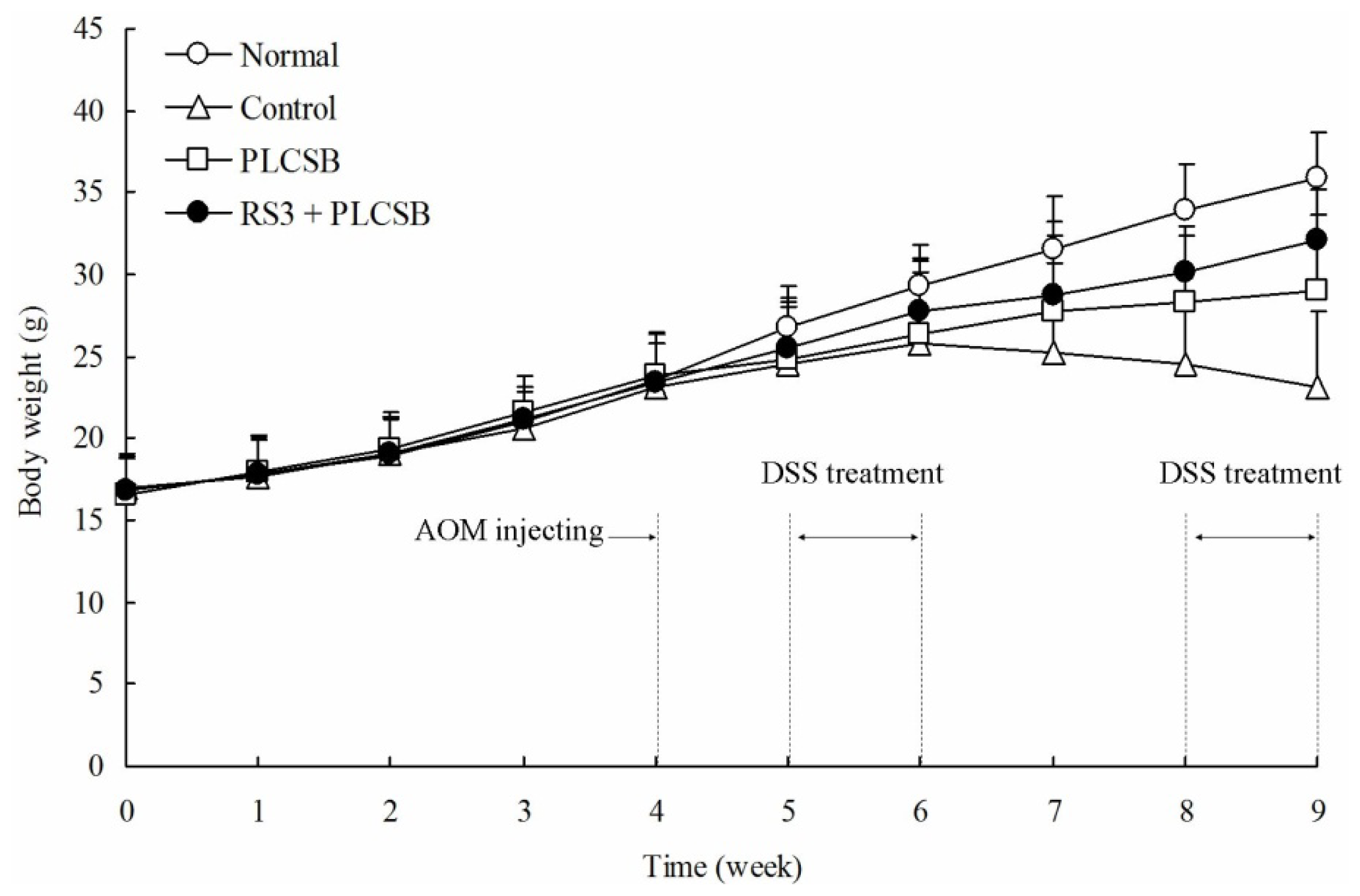
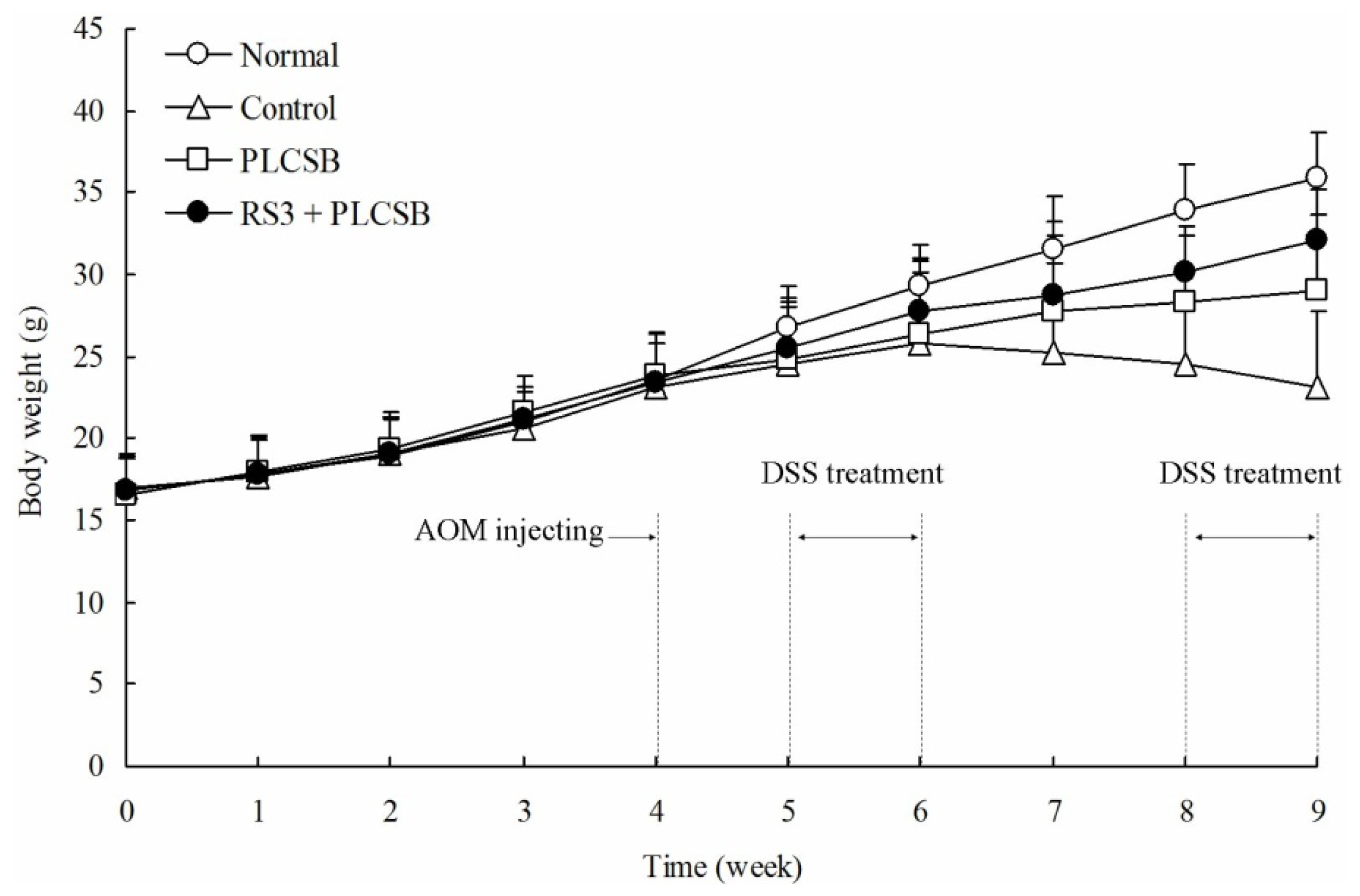
2.1. Body Weight and Colon Observations
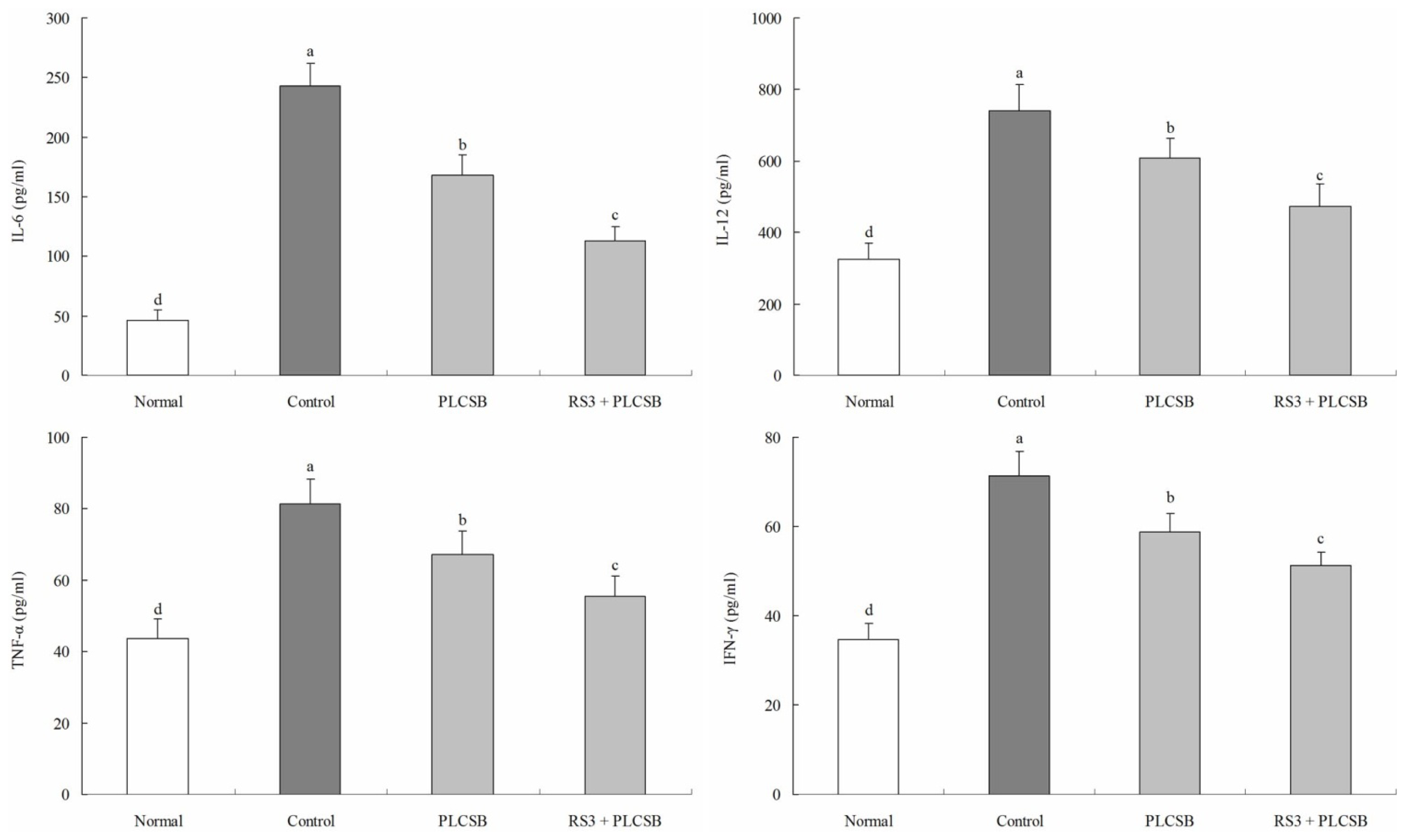
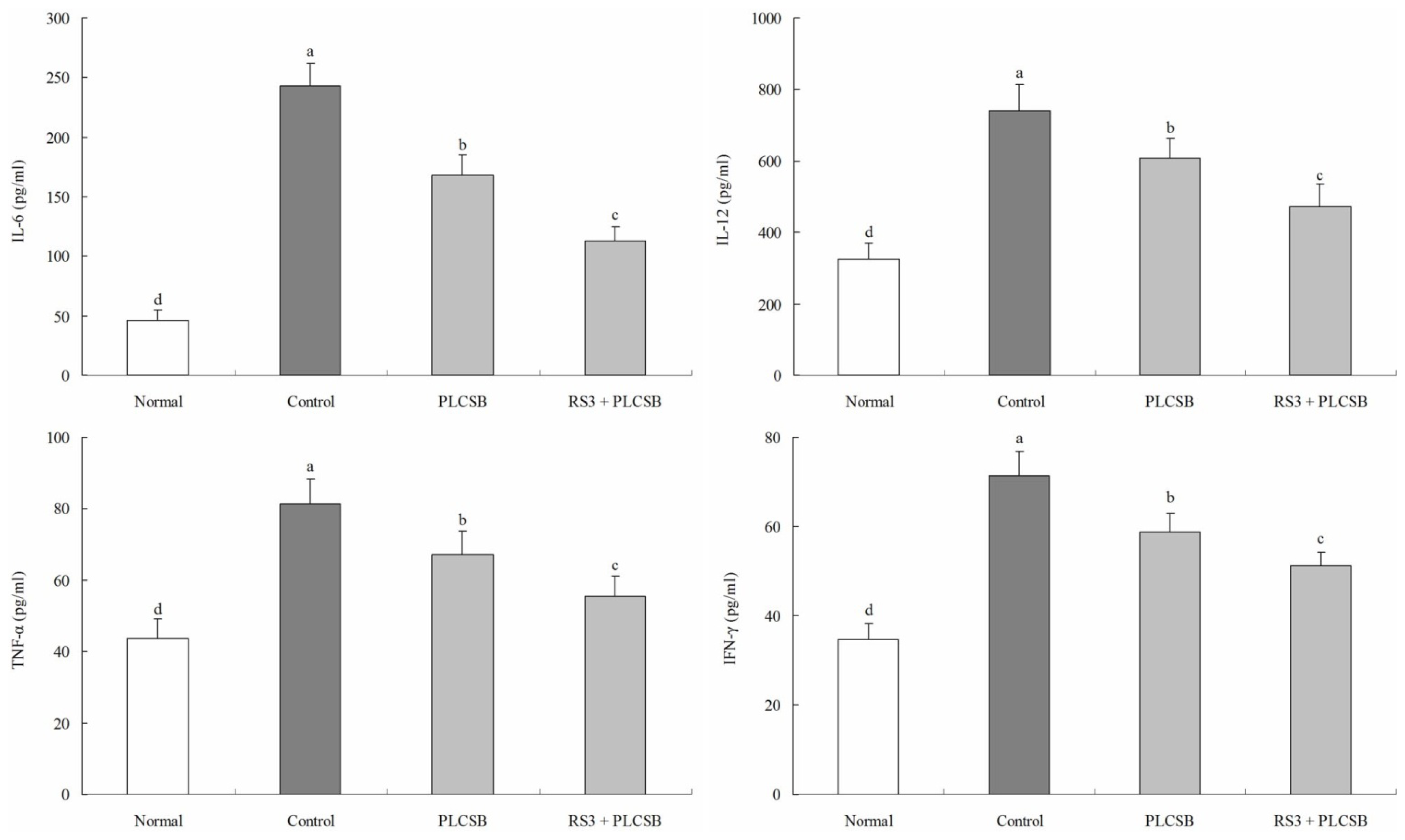
2.2. Effect of PLCSB and RS3 + PLCSB on Serum Levels of IL-6, IL-12, TNF-α and IFN-γ
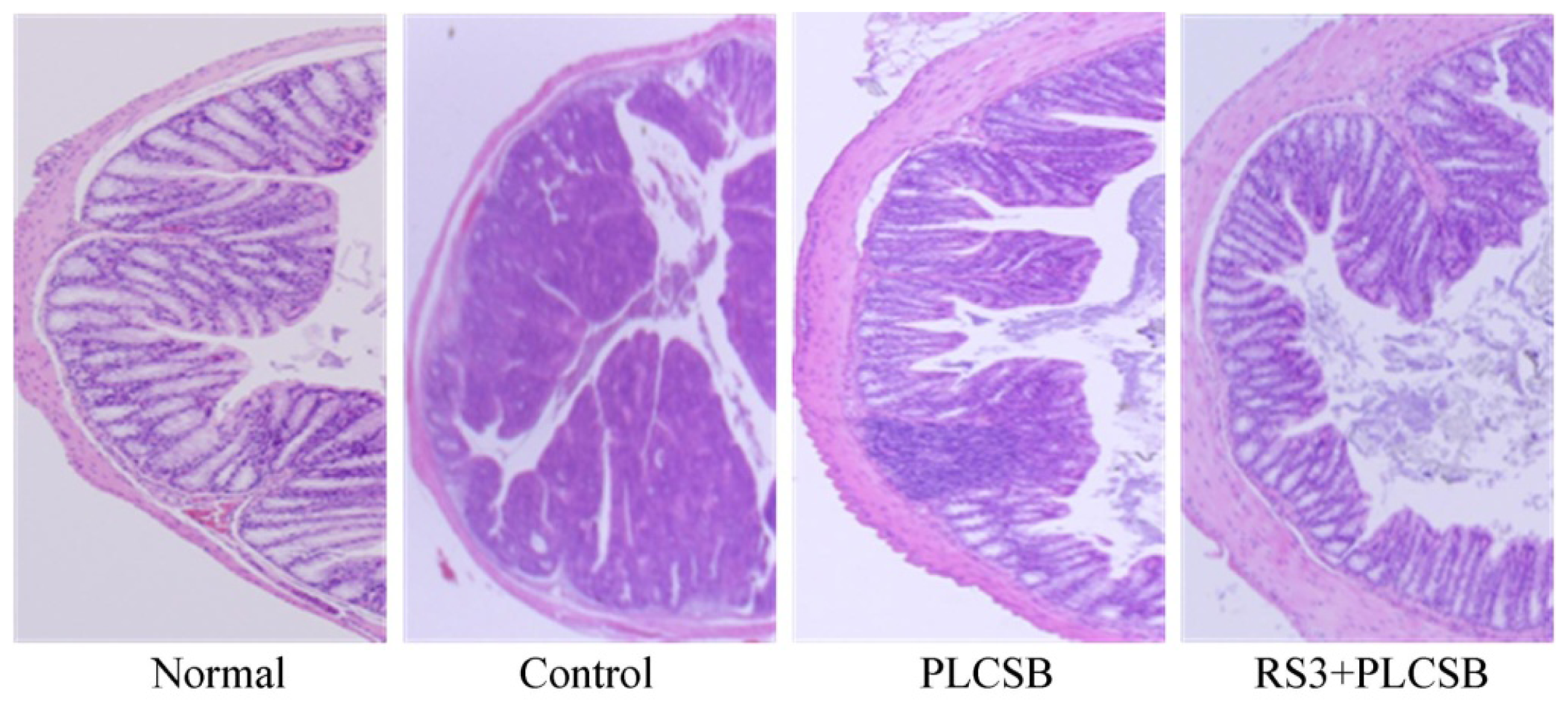
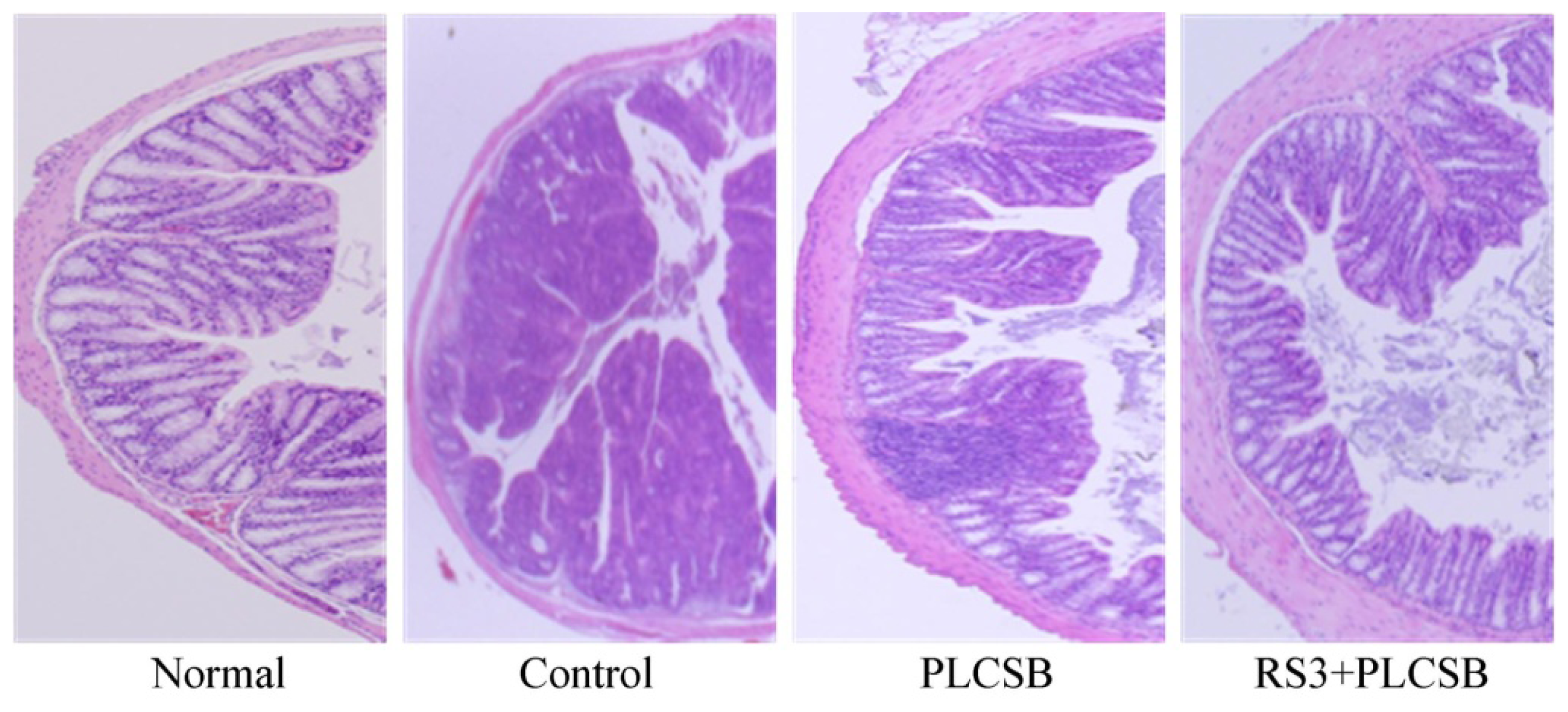
2.3. Effects of PLCSB and RS3 + PLCSB on Histological Changes
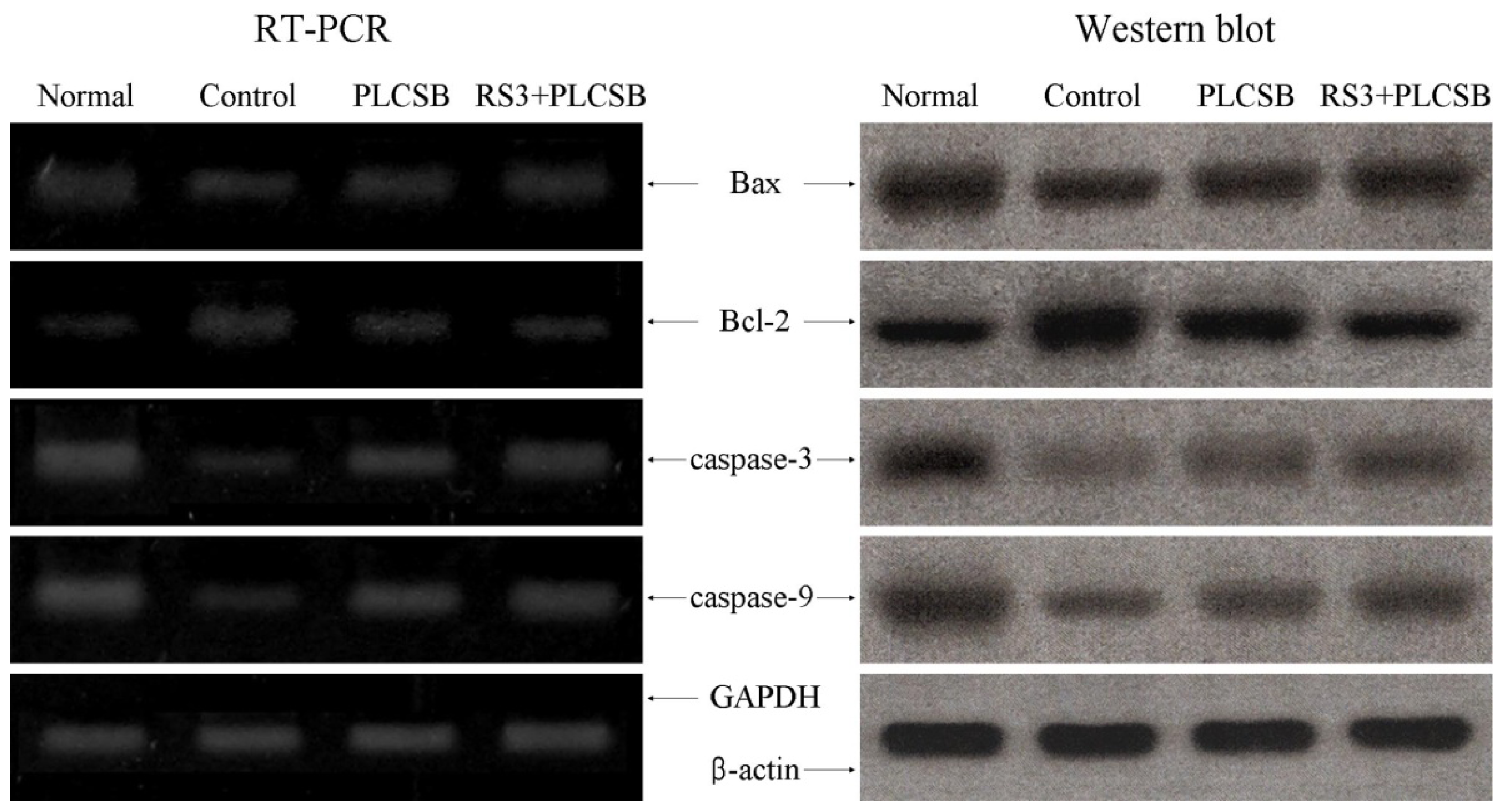
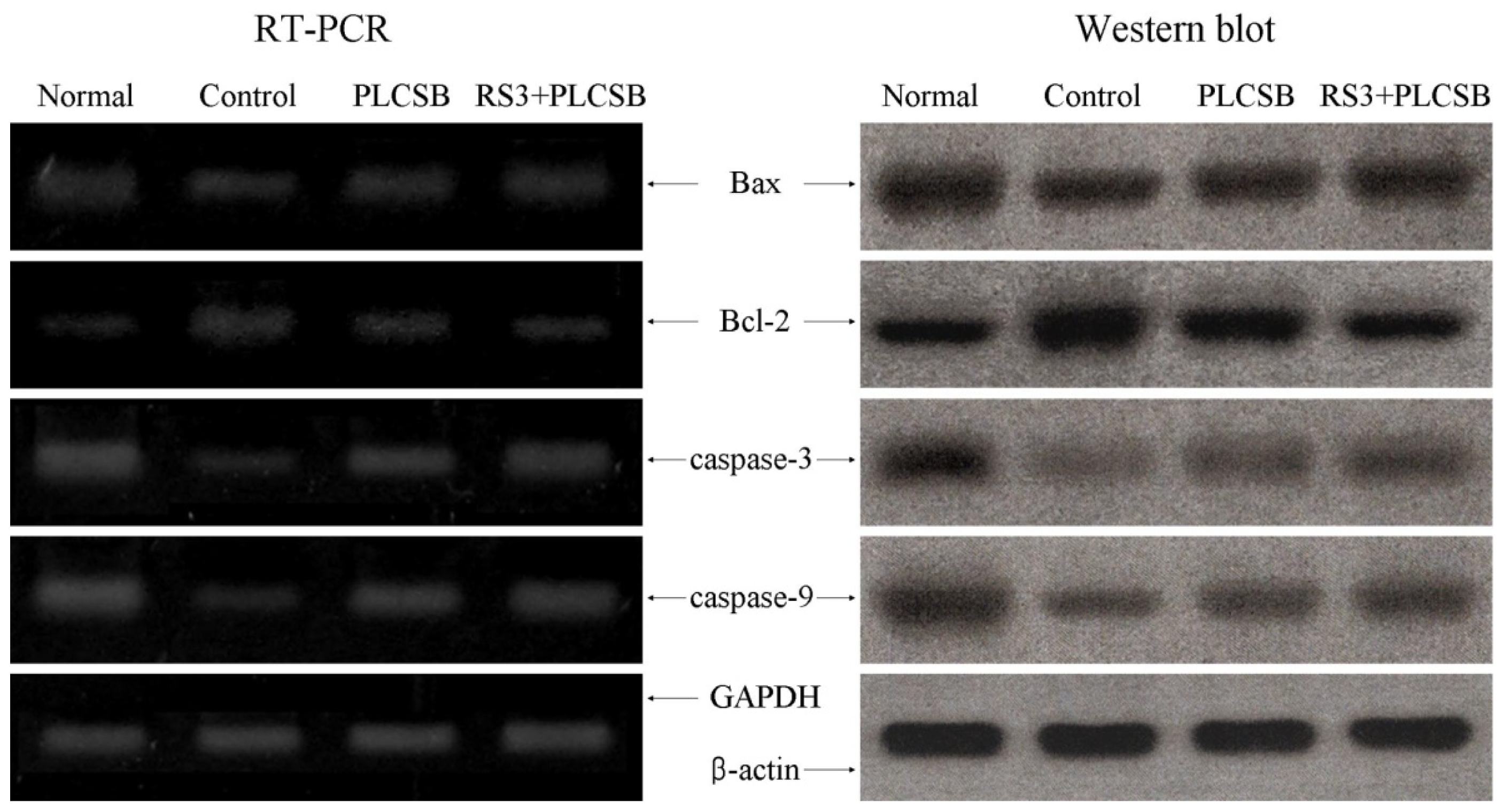
2.4. Effects of PLCSB and RS3 + PLCSB on Apoptosis-Related Gene Expression of Bax, Bcl-2 and Caspases
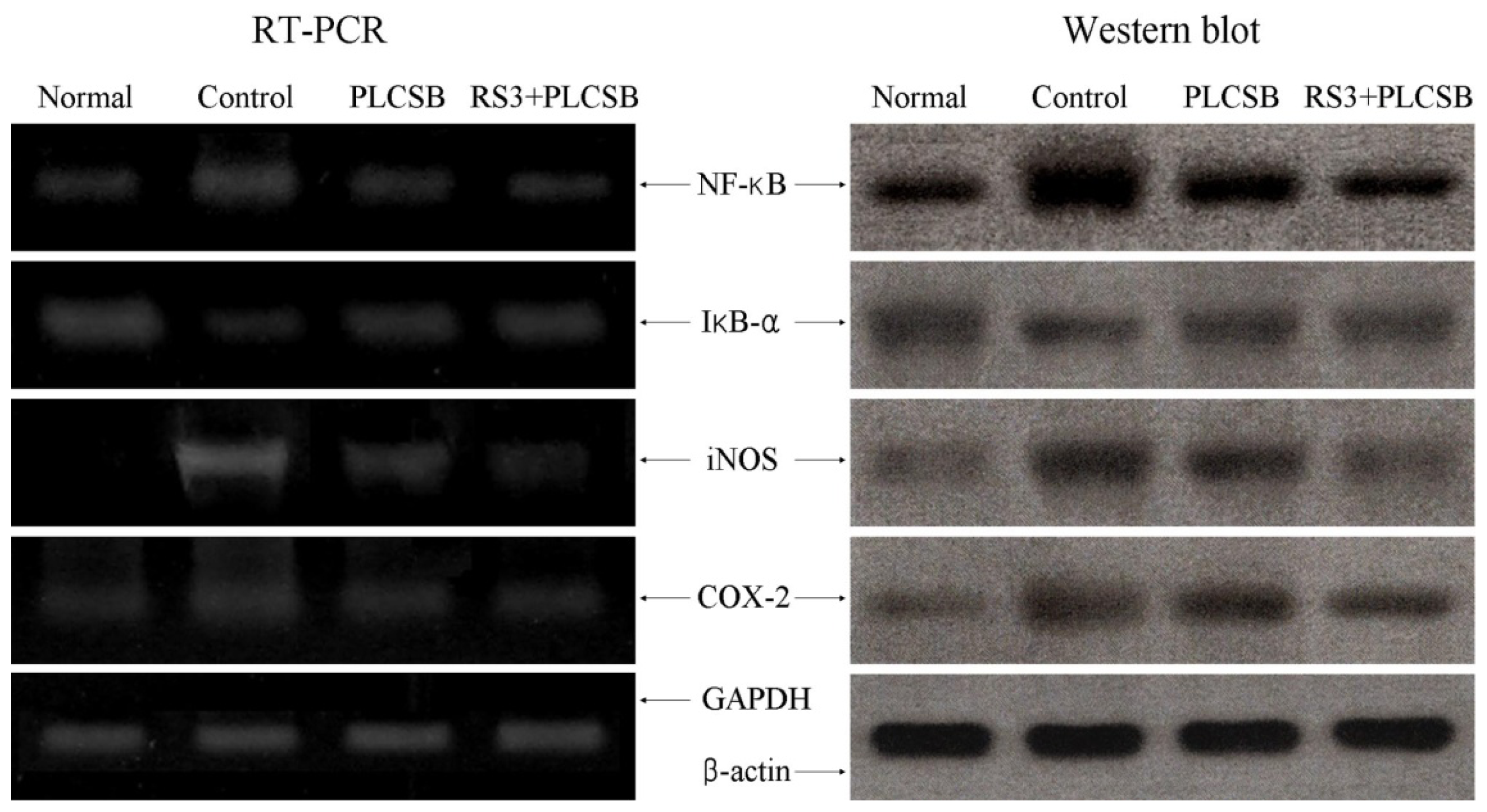
2.5. Effects of PLCSB and RS3 + PLCSB on Inflammation-Related Gene Expression of NF-κB, IκB-α, iNOS and COX-2
3. Experimental Section
3.1. Polysaccharide Larimichthys crocea Swimming Bladder (PLCSB) Preparation
3.2. Animals
3.3. Azoxymethane (AOM) and Dextran Sulfate Sodium (DSS)-Induced Colorectal Carcinogenesis Model
3.4. Analysis of Inflammation-Related Cytokines in Serum by Enzyme-Linked Immunosorbent Assay (ELISA)
3.5. Histological Analysis
3.6. RT-PCR Assay
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsXin Zhao made contribution to conception, design, and give final approval of the version to be submitted; Lian-Hong Chen, Xin Zhao, Jia-Le Song did the experiments and Lian-Hong Chen, Jia-Le Song, Yu Qian made contribution to acquisition of data, analysis and interpretation of data; Hua-Yi Suo and Jian Li made contribution to revise the manuscript.
References
- Jian, J.C.; Wu, Z.H. Effects of traditional Chinese medicine on nonspecific immunity and disease resistance of large yellow croaker, Pseudosciaena crocea (Richardson). Aquaculture 2003, 218, 1–9. [Google Scholar]
- Li, C.; Yao, C.L. Molecular and expression characterizations of interleukin-8 gene in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol 2013, 34, 799–809. [Google Scholar]
- Liu, S.; Yu, B. Peptides from variegated carp (Aristichthys nobilis) swim bladder: Fermentation production and assessment of antioxidant properties. Food Sci 2009, 30, 332–334. [Google Scholar]
- Bisen, P.S.; Baghel, R.K.; Sanodiya, B.S.; Thakur, G.S.; Prasad, G.B. Lentinus edodes: A macrofungus with pharmacological activities. Curr. Med. Chem 2010, 17, 2419–2430. [Google Scholar]
- Chen, H.L.; Li, B.T.; Zhang, J.X.; Li, D.F.; Chang, B.Y.; Xu, L.T. Research development on the immumodulatory effect of polysaccharide and its mechanism. Chin. Pharm. Bull 2002, 3, 249–252. [Google Scholar]
- Qian, Y.; Zhao, X.; Song, J.L.; Zhu, K.; Sun, P.; Li, G.J.; Wang, R.; Kan, J.Q. Inhibitory effects of resistant starch (RS3) as a carrier for stachyose on dextran sulfate sodium-induced ulcerative colitis in C57BL/6 mice. Exp. Ther. Med 2013, 6, 1312–1316. [Google Scholar]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr 1992, 46, S33–S50. [Google Scholar]
- Baghurst, P.A.; Baghurst, K.I.; Record, S.J. Dietary fiber, non-starch polysaccharides and resistant starch—A review. Food Aust 1996, 48, S1–S36. [Google Scholar]
- Fan, Z.D.; Deng, D.W.; Gu, Z.Y.; Huang, G.H. Animal experimental study on physiological function of P. Lobata (Willd.) ohawi resistant starch. Food Sci 2006, 27, 492–494. [Google Scholar]
- Jeong, J.K.; Chang, H.K.; Park, K.Y. Inhibitory effects of meju prepared with mixed starter cultures on azoxymethane and dextran sulfate sodium-induced colon carcinogenesis in mice. J. Carcin 2012, 11, 30–38. [Google Scholar]
- Li, H.; Wu, W.K.; Li, Z.J.; Chan, K.M.; Wong, C.C.; Ye, C.G.; Yu, L.; Sung, J.J.; Cho, C.H.; Wang, M. 2,3′,4,4′,5′-Pentamethoxy-trans-stilbene, a resveratrol derivative, inhibits colitis-associated colorectal carcinogenesis in mice. Br. J. Pharmacol 2010, 160, 1352–1361. [Google Scholar]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar]
- Bai, A.; Peng, Z. Biological therapies of inflammatory bowel disease. Immunotherapy 2010, 2, 727–742. [Google Scholar]
- Perrier, C.; Rutgeerts, P. Cytokine blockade in inflammatory bowel diseases. Immunotherapy 2011, 3, 1341–1352. [Google Scholar]
- Lin, W.W.; Karin, M. A cytokine-mediated link between innate immunity, inflammation, and cancer. J. Clin. Invest 2007, 117, 1175–1183. [Google Scholar]
- Cahlin, C.; Körner, A.; Axelsson, H.; Wang, W.H.; Lundholm, K.; Svanberg, E. Experimental cancer cachexia: The role of host-derived cytokines interleukin (IL)-6, IL-12, interferon-γ, and tumor necrosis factor α evaluated in gene knockout, tumor-bearing mice on C57 Bl background and eicosanoid-dependent cachexia. Cancer Res 2000, 60, 5488–5493. [Google Scholar]
- Kishimoto, T.; Akira, S.; Taga, T. Interleukin-6 and its receptor: A paradigm for cytokines. Science 1992, 258, 593–597. [Google Scholar]
- Waldner, M.J.; Foersch, S.; Neurath, M.F. Interleukin-6—A key regulator of colorectal cancer development. Int. J. Biol. Sci 2012, 8, 1248–1253. [Google Scholar]
- Engel, M.A.; Neurath, M.F. Anticancer properties of the IL-12 family—Focus on colorectal cancer. Curr. Med. Chem 2010, 17, 3303–3308. [Google Scholar]
- Popivanova, B.K.; Kitamura, K.; Wu, Y.; Kondo, T.; Kagaya, T.; Kaneko, S.; Oshima, M.; Fujii, C.; Mukaida, N. Blocking TNF-α in mice reduces colorectal carcinogenesis associated with chronic colitis. J. Clin. Invest 2008, 118, 560–570. [Google Scholar]
- Alexander, J.; Watanabe, T.; Wu, T.T.; Rashid, A.; Li, S.; Hamilton, S.R. Histopathological identification of colon cancer with microsatellite instability. Am. J. Pathol 2001, 158, 527–535. [Google Scholar]
- Milanezi, F.; Leitão, D.; Ricardo, S.; Augusto, I.; Schmitt, F. Evaluation of HER2 in breast cancer: Reality and expectations. Expert Opin. Med. Diagn 2009, 3, 607–620. [Google Scholar]
- Chao, D.T.; Korsmeyer, S.J. Bcl-2 family: Regulators of cell death. Annu. Rev. Immunol 1998, 16, 395–419. [Google Scholar]
- Kidd, V.J. Proteolytic activities that mediate apoptosis. Annu. Rev. Physiol 1998, 60, 533–573. [Google Scholar]
- Blanc, C.; Deveraux, Q.L.; Krajewski, S.; Jänicke, R.U.; Porter, A.G.; Reed, J.C.; Jaggi, R.; Marti, A. Caspase-3 is essential for procaspase-9 processing and cisplatin-induced apoptosis of MCF-7 breast cancer cells. Cancer Res 2000, 60, 4386–4390. [Google Scholar]
- Kirsch, D.G.; Doseff, A.; Chau, B.N.; Lim, D.S.; de Souza-Pinto, N.C.; Hansford, R.; Kastan, M.B.; Lazebnik, Y.A.; Hardwick, J.M. Caspase-3-dependent cleavage of Bcl-2 promotes release of cytochrome c. J. Biol. Chem 1999, 274, 21155–21161. [Google Scholar]
- Delić, R.; Stefanović, M. Optimal laboratory panel for predicting preeclampsia. J. Matern. Fetal. Neonatal. Med 2010, 23, 96–102. [Google Scholar]
- Baeuerle, P.A. IkappaB-NF-kappaB structures: At the interface of inflammation control. Cell 1998, 95, 729–731. [Google Scholar]
- Sánchez-Pérez, I.; Benitah, S.A.; Martínez-Gomariz, M.; Lacal, J.C.; Perona, R. Cell stress and MEKK1-mediated c-Jun activation modulate NFkappaB activity and cell viability. Mol. Biol. Cell 2002, 13, 2933–2945. [Google Scholar]
- Yan, C.; Yan, X. Study on extraction of Lycium barbarum polysaccharides by different methods and their antioxidant effects in vitro. Food Sci. 2008, 29, 183–187. [Google Scholar]
- Song, J.L.; Qian, Y.; Li, G.J.; Zhao, X. Anti-inflammatory effects of kudingcha methanol extract (Ilex kudingcha C.J. Tseng) in dextran sulfate sodium-induced ulcerative colitis. Mol. Med. Rep 2013, 8, 1256–1262. [Google Scholar]
- Zhao, X.; Kim, S.Y.; Park, K.Y. Bamboo salt has in vitro anticancer activity in HCT-116 cells and exerts anti-metastatic effects in vivo. J. Med. Food 2013, 16, 9–19. [Google Scholar]
- Zhao, X. Hawk tea (Litsea coreana Levl. var. lanuginose) attenuates CCl4-induced hepatic damage in Sprague-Dawley rats. Exp. Ther. Med 2013, 5, 555–560. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Colon length (mm) | Colon weight (g) | Colon weight/length (mg/cm) |
|---|---|---|---|
| Normal | 80.33 ± 3.86 a | 0.23 ± 0.03 d | 28.63 ± 7.77 d |
| Control | 72.14 ± 5.21 d | 0.47 ± 0.08 a | 65.15 ± 15.36 a |
| PLCSB | 75.47 ± 3.21 c | 0.35 ± 0.02 b | 46.38 ± 6.23 b |
| RS3 + PLCSB | 77.31 ± 2.58 b | 0.30 ± 0.03 c | 38.80 ± 11.63 c |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, L.-H.; Song, J.-L.; Qian, Y.; Zhao, X.; Suo, H.-Y.; Li, J. Increased Preventive Effect on Colon Carcinogenesis by Use of Resistant Starch (RS3) as the Carrier for Polysaccharide of Larimichthys Crocea Swimming Bladder. Int. J. Mol. Sci. 2014, 15, 817-829. https://doi.org/10.3390/ijms15010817
Chen L-H, Song J-L, Qian Y, Zhao X, Suo H-Y, Li J. Increased Preventive Effect on Colon Carcinogenesis by Use of Resistant Starch (RS3) as the Carrier for Polysaccharide of Larimichthys Crocea Swimming Bladder. International Journal of Molecular Sciences. 2014; 15(1):817-829. https://doi.org/10.3390/ijms15010817
Chicago/Turabian StyleChen, Lian-Hong, Jia-Le Song, Yu Qian, Xin Zhao, Hua-Yi Suo, and Jian Li. 2014. "Increased Preventive Effect on Colon Carcinogenesis by Use of Resistant Starch (RS3) as the Carrier for Polysaccharide of Larimichthys Crocea Swimming Bladder" International Journal of Molecular Sciences 15, no. 1: 817-829. https://doi.org/10.3390/ijms15010817