Melatonin Signaling and Its Modulation of PfNF-YB Transcription Factor Expression in Plasmodium falciparum


{kind=link}
Abstract
:1. Introduction
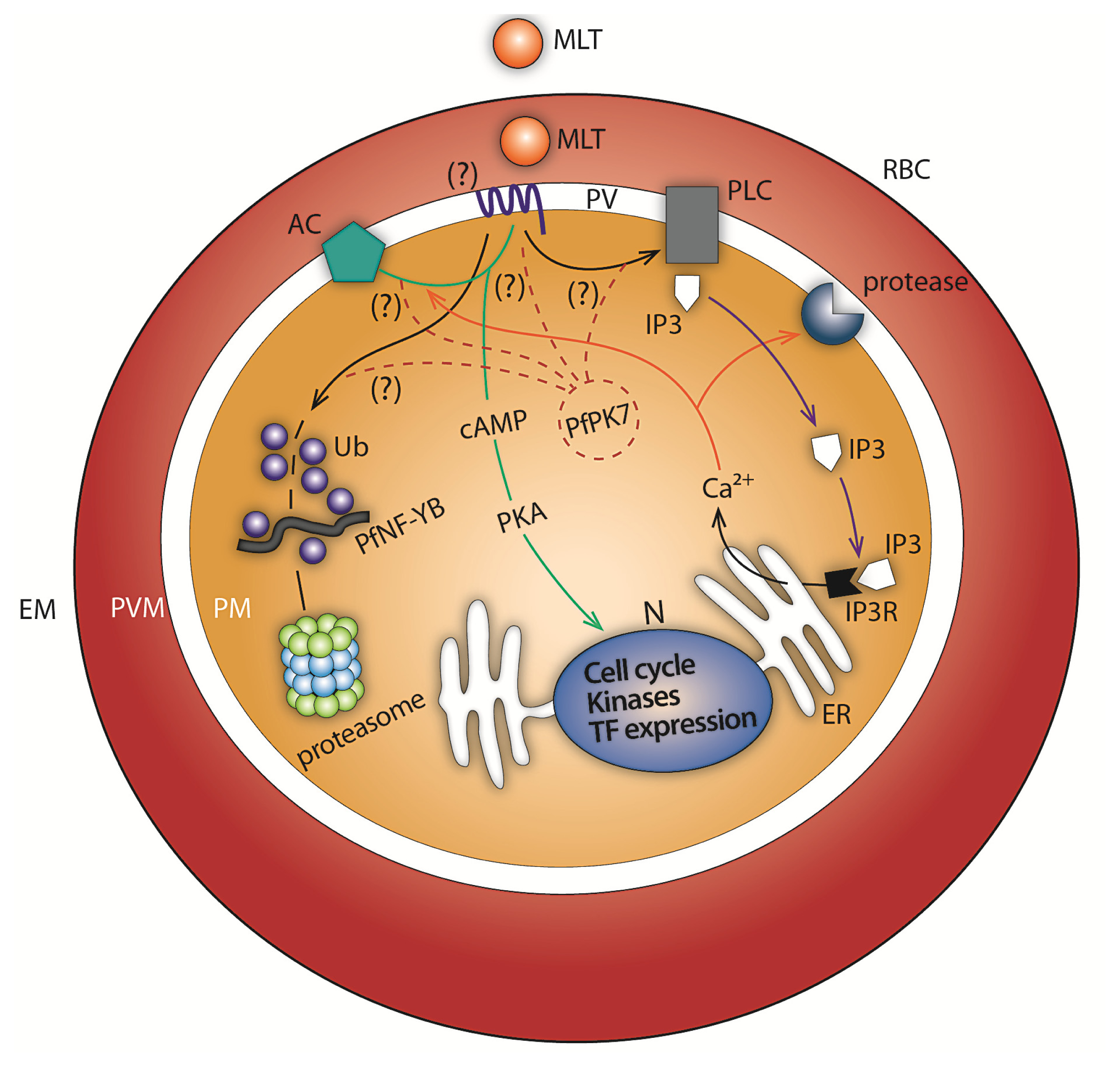
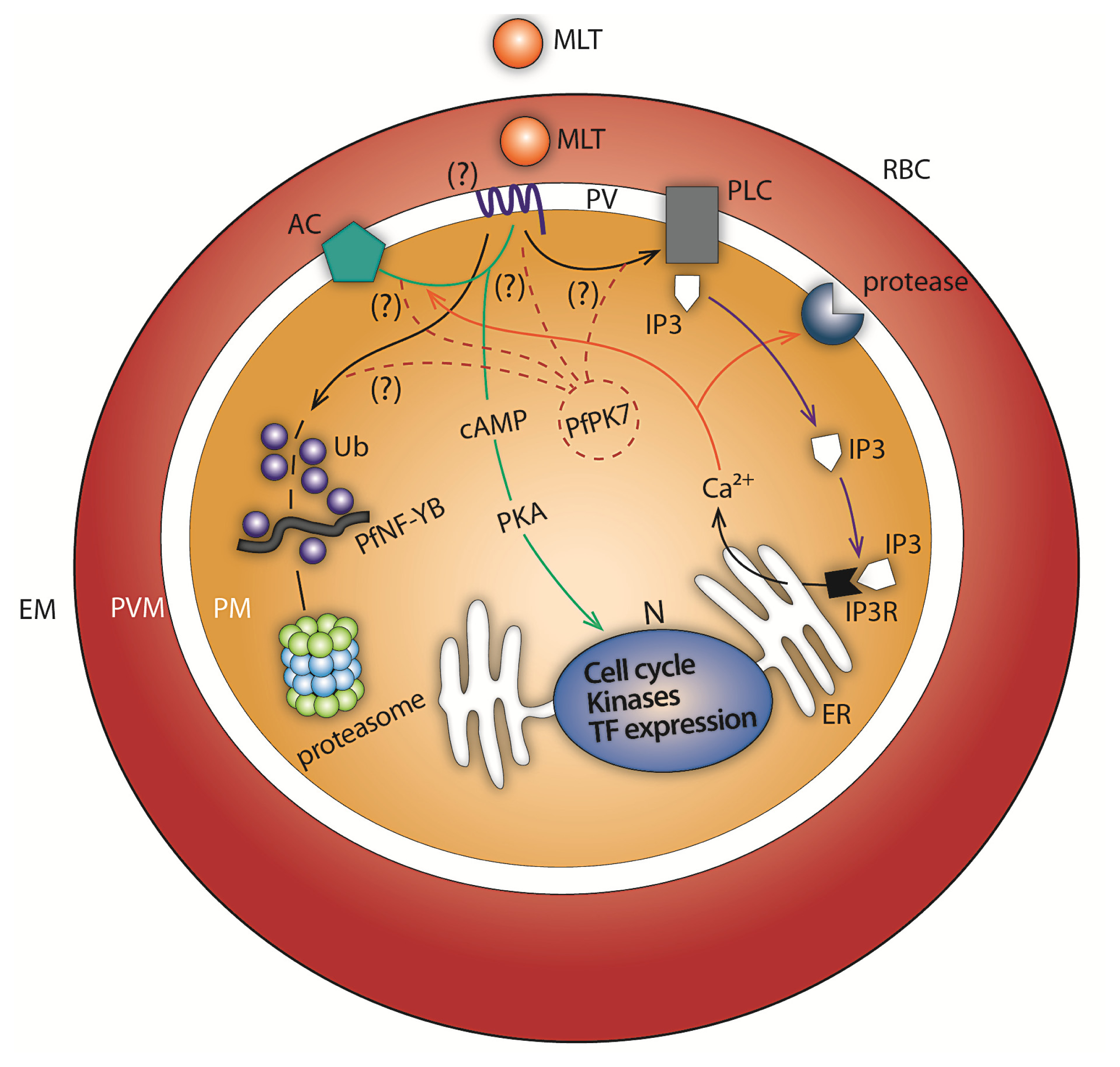
2. Melatonin Signaling: Calcium and cAMP Generation in P. falciparum and P. chabaudi
3. Melatonin Modulates the Expression of a Subset of Genes of the Ubiquitin Proteasome System (UPS) in P. falciparum
4. Melatonin Signaling Modulates PfNF-YB Transcription Factor Expression in P. falciparum
5. Melatonin Blockers as Potential Antimalarial Drugs
6. Conclusions
Acknowledgments
Conflict of Interest
References
- Hempelmann, E.; Tesarowicz, I.; Oleksyn, B.J. From onions to artemisinin. Brief history of malaria chemotherapy. Pharm. Unserer Zeit 2009, 38, 500–507. [Google Scholar]
- World Malaria Report 2012. Available online: http://www.who.int/malaria/publications/world_malaria_report_2012/en/index.html (on accessed 19 April 2013).
- Khan, S.M.; Waters, A.P. Malaria parasite transmission stages: An update. Trends Parasitol 2004, 20, 575–580. [Google Scholar]
- Bannister, L.H.; Hopkins, J.M.; Fowler, R.E.; Krishna, S.; Mitchell, G.H. A brief illustrated guide to the ultrastructure of Plasmodium falciparum asexual blood stages. Parasitol. Today 2000, 16, 427–433. [Google Scholar]
- Garcia, C.R.; de Azevedo, M.F.; Wunderlich, G.; Budu, A.; Young, J.A.; Bannister, L. Plasmodium in the postgenomic era: New insights into the molecular cell biology of malaria parasites. Int. Rev. Cell Mol. Biol 2008, 266, 85–156. [Google Scholar]
- Mita-Mendoza, N.K.; van de Hoef, D.L.; Lopera-Mesa, T.M.; Doumbia, S.; Konate, D.; Doumbouya, M.; Gu, W.; Anderson, J.M.; Santos-Argumedo, L.; Rodriguez, A.; et al. A potential role for plasma uric acid in the endothelial pathology of Plasmodium falciparum malaria. PLoS One 2013, 8, e54481. [Google Scholar]
- White, N.J.; Warrell, D.A.; Looareesuwan, S.; Chanthavanich, P.; Phillips, R.E.; Pongpaew, P. Pathophysiological and prognostic significance of cerebrospinal-fluid lactate in cerebral malaria. Lancet 1985, 1, 776–778. [Google Scholar]
- Reis, P.A.; Estato, V.; da Silva, T.I.; d’Avila, J.C.; Siqueira, L.D.; Assis, E.F.; Bozza, P.T.; Bozza, F.A.; Tibirica, E.V.; Zimmerman, G.A.; et al. Statins decrease neuroinflammation and prevent cognitive impairment after cerebral malaria. PLoS Pathog 2012, 8, e1003099. [Google Scholar]
- Hawking, F.; Worms, M.J.; Gammage, K. 24- and 48-hour cycles of malaria parasites in the blood; their purpose, production and control. Trans. R. Soc. Trop. Med. Hyg 1968, 62, 731–765. [Google Scholar]
- Newbold, C.I.; Boyle, D.B.; Smith, C.C.; Brown, K.N. Stage specific protein and nucleic acid synthesis during the asexual cycle of the rodent malaria Plasmodium chabaudi. Mol. Biochem. Parasitol 1982, 5, 33–44. [Google Scholar]
- Gardner, M.J.; Hall, N.; Fung, E.; White, O.; Berriman, M.; Hyman, R.W.; Carlton, J.M.; Pain, A.; Nelson, K.E.; Bowman, S.; et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002, 419, 498–511. [Google Scholar]
- Grover, M.; Chaubey, S.; Ranade, S.; Tatu, U. Identification of an exported heat shock protein 70 in Plasmodium falciparum. Parasite 2013, 20, 1–9. [Google Scholar]
- Cai, H.; Hong, C.; Gu, J.; Lilburn, T.G.; Kuang, R.; Wang, Y. Module-based subnetwork alignments reveal novel transcriptional regulators in malaria parasite Plasmodium falciparum. BMC Syst. Biol 2012, 6, S5, , doi:10.1186/1752-0509-6-S3-S5.. [Google Scholar]
- Doerig, C.; Baker, D.; Billker, O.; Blackman, M.J.; Chitnis, C.; Dhar Kumar, S.; Heussler, V.; Holder, A.A.; Kocken, C.; Krishna, S.; et al. Signalling in malaria parasites. The MALSIG consortium. Parasite 2009, 16, 169–182. [Google Scholar]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J 2006, 273, 2813–2838. [Google Scholar]
- Jan, J.E.; Reiter, R.J.; Wasdell, M.B.; Bax, M. The role of the thalamus in sleep, pineal melatonin production, and circadian rhythm sleep disorders. J. Pineal. Res 2009, 46, 1–7. [Google Scholar]
- Reiter, R.J.; Paredes, S.D.; Manchester, L.C.; Tan, D.X. Reducing oxidative/nitrosative stress: A newly-discovered genre for melatonin. Crit. Rev. Biochem. Mol. Biol 2009, 44, 175–200. [Google Scholar]
- Ben-Nathan, D.; Maestroni, G.J.; Lustig, S.; Conti, A. Protective effects of melatonin in mice infected with encephalitis viruses. Arch. Virol 1995, 140, 223–230. [Google Scholar]
- Bonilla, E.; Valero, N.; Chacin-Bonilla, L.; Medina-Leendertz, S. Melatonin and viral infections. J. Pineal. Res 2004, 36, 73–79. [Google Scholar]
- McNulty, S.; Ross, A.W.; Barrett, P.; Hastings, M.H.; Morgan, P.J. Melatonin regulates the phosphorylation of CREB in ovine pars tuberalis. J. Neuroendocrinol 1994, 6, 523–532. [Google Scholar]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal. Res 1995, 18, 28–31. [Google Scholar]
- Paredes, S.D.; Korkmaz, A.; Manchester, L.C.; Tan, D.X.; Reiter, R.J. Phytomelatonin: A review. J. Exp. Bot 2009, 60, 57–69. [Google Scholar]
- Balzer, I.; Hardeland, R. Photoperiodism and effects of indoleamines in a unicellular alga, Gonyaulax polyedra. Science 1991, 253, 795–797. [Google Scholar]
- Hardeland, R.; Balzer, I.; Poeggeler, B.; Fuhrberg, B.; Uria, H.; Behrmann, G.; Wolf, R.; Meyer, T.J.; Reiter, R.J. On the primary functions of melatonin in evolution: Mediation of photoperiodic signals in a unicell, photooxidation, and scavenging of free radicals. J. Pineal. Res 1995, 18, 104–111. [Google Scholar]
- Yamada, Y.; Forger, D. Multiscale complexity in the mammalian circadian clock. Curr. Opin. Genet. Dev 2010, 20, 626–633. [Google Scholar]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar]
- Golombek, D.A.; Rosenstein, R.E. Physiology of circadian entrainment. Physiol. Rev 2010, 90, 1063–1102. [Google Scholar]
- Agez, L.; Laurent, V.; Guerrero, H.Y.; Pevet, P.; Masson-Pevet, M.; Gauer, F. Endogenous melatonin provides an effective circadian message to both the suprachiasmatic nuclei and the pars tuberalis of the rat. J. Pineal. Res 2009, 46, 95–105. [Google Scholar]
- Hotta, C.T.; Gazarini, M.L.; Beraldo, F.H.; Varotti, F.P.; Lopes, C.; Markus, R.P.; Pozzan, T.; Garcia, C.R. Calcium-dependent modulation by melatonin of the circadian rhythm in malarial parasites. Nat. Cell Biol 2000, 2, 466–468. [Google Scholar]
- Hotta, C.T.; Markus, R.P.; Garcia, C.R. Melatonin and N-acetyl-serotonin cross the red blood cell membrane and evoke calcium mobilization in malarial parasites. Braz. J. Med. Biol. Res 2003, 36, 1583–1587. [Google Scholar]
- Beraldo, F.H.; Garcia, C.R. Products of tryptophan catabolism induce Ca2+ release and modulate the cell cycle of Plasmodium falciparum malaria parasites. J. Pineal. Res 2005, 39, 224–230. [Google Scholar]
- Budu, A.; Peres, R.; Bueno, V.B.; Catalani, L.H.; Garcia, C.R. N1-acetyl-N2-formyl-5-methoxykynuramine modulates the cell cycle of malaria parasites. J. Pineal Res 2007, 42, 261–266. [Google Scholar]
- Alves, E.; Bartlett, P.J.; Garcia, C.R.; Thomas, A.P. Melatonin and IP3-induced Ca2+ release from intracellular stores in the malaria parasite Plasmodium falciparum within infected red blood cells. J. Biol. Chem 2011, 286, 5905–5912. [Google Scholar]
- Beraldo, F.H.; Almeida, F.M.; da Silva, A.M.; Garcia, C.R. Cyclic AMP and calcium interplay as second messengers in melatonin-dependent regulation of Plasmodium falciparum cell cycle. J. Cell Biol 2005, 170, 551–557. [Google Scholar]
- Syin, C.; Parzy, D.; Traincard, F.; Boccaccio, I.; Joshi, M.B.; Lin, D.T.; Yang, X.M.; Assemat, K.; Doerig, C.; Langsley, G. The H89 cAMP-dependent protein kinase inhibitor blocks Plasmodium falciparum development in infected erythrocytes. Eur. J. Biochem 2001, 268, 4842–4849. [Google Scholar]
- Rached, F.B.; Ndjembo-Ezougou, C.; Chandran, S.; Talabani, H.; Yera, H.; Dandavate, V.; Bourdoncle, P.; Meissner, M.; Tatu, U.; Langsley, G. Construction of a Plasmodium falciparum Rab-interactome identifies CK1 and PKA as Rab-effector kinases in malaria parasites. Biol. Cell 2012, 104, 34–47. [Google Scholar]
- Merckx, A.; Bouyer, G.; Thomas, S.L.; Langsley, G.; Egee, S. Anion channels in Plasmodium-falciparum-infected erythrocytes and protein kinase A. Trends Parasitol 2009, 25, 139–144. [Google Scholar]
- Lasonder, E.; Green, J.L.; Camarda, G.; Talabani, H.; Holder, A.A.; Langsley, G.; Alano, P. The Plasmodium falciparum schizont phosphoproteome reveals extensive phosphatidylinositol and cAMP-protein kinase A signaling. J. Proteome Res 2012, 11, 5323–5337. [Google Scholar]
- Holder, A.A.; Mohd Ridzuan, M.A.; Green, J.L. Calcium dependent protein kinase 1 and calcium fluxes in the malaria parasite. Microb. Infect 2012, 14, 825–830. [Google Scholar]
- Berridge, M.J. Inositol trisphosphate and calcium signalling mechanisms. Biochim. Biophys. Acta 2009, 1793, 933–940. [Google Scholar]
- Ishino, T.; Orito, Y.; Chinzei, Y.; Yuda, M. A calcium-dependent protein kinase regulates Plasmodium ookinete access to the midgut epithelial cell. Mol. Microbiol 2006, 59, 1175–1184. [Google Scholar]
- Ono, T.; Cabrita-Santos, L.; Leitao, R.; Bettiol, E.; Purcell, L.A.; Diaz-Pulido, O.; Andrews, L.B.; Tadakuma, T.; Bhanot, P.; Mota, M.M.; et al. Adenylyl cyclase alpha and cAMP signaling mediate Plasmodium sporozoite apical regulated exocytosis and hepatocyte infection. PLoS Pathog 2008, 4, e1000008. [Google Scholar]
- Alleva, L.M.; Kirk, K. Calcium regulation in the intraerythrocytic malaria parasite Plasmodium falciparum. Mol. Biochem. Parasitol 2001, 117, 121–128. [Google Scholar]
- Biagini, G.A.; Bray, P.G.; Spiller, D.G.; White, M.R.; Ward, S.A. The digestive food vacuole of the malaria parasite is a dynamic intracellular Ca2+ store. J. Biol. Chem 2003, 278, 27910–27915. [Google Scholar]
- Billker, O.; Lourido, S.; Sibley, L.D. Calcium-dependent signaling and kinases in apicomplexan parasites. Cell Host Microb 2009, 5, 612–622. [Google Scholar]
- Valderramos, S.G.; Scanfeld, D.; Uhlemann, A.C.; Fidock, D.A.; Krishna, S. Investigations into the role of the Plasmodium falciparum SERCA (PfATP6) L263E mutation in artemisinin action and resistance. Antimicrob. Agents Chemotherap 2010, 54, 3842–3852. [Google Scholar]
- Agarwal, S.; Singh, M.K.; Garg, S.; Chitnis, C.E.; Singh, S. Ca(2+) -mediated exocytosis of subtilisin-like protease 1: A key step in egress of Plasmodium falciparum merozoites. Cell. Microbiol 2012. [Google Scholar] [CrossRef]
- Beraldo, F.H.; Mikoshiba, K.; Garcia, C.R. Human malarial parasite, Plasmodium falciparum, displays capacitative calcium entry: 2-aminoethyl diphenylborinate blocks the signal transduction pathway of melatonin action on the P. falciparum cell cycle. J. Pineal. Res 2007, 43, 360–364. [Google Scholar]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem 1998, 67, 425–479. [Google Scholar]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev 2002, 82, 373–428. [Google Scholar]
- Ciechanover, A.; Schwartz, A.L. The ubiquitin-proteasome pathway: The complexity and myriad functions of proteins death. Proc. Natl. Acad. Sci. USA 1998, 95, 2727–2730. [Google Scholar]
- Baumeister, W.; Walz, J.; Zuhl, F.; Seemuller, E. The proteasome: Paradigm of a self-compartmentalizing protease. Cell 1998, 92, 367–380. [Google Scholar]
- Voges, D.; Zwickl, P.; Baumeister, W. The 26S proteasome: A molecular machine designed for controlled proteolysis. Annu. Rev. Biochem 1999, 68, 1015–1068. [Google Scholar]
- Naujokat, C.; Hoffmann, S. Role and function of the 26S proteasome in proliferation and apoptosis. Lab. Investig. J. Tech. Methods Pathol 2002, 82, 965–980. [Google Scholar]
- Rock, K.L.; York, I.A.; Saric, T.; Goldberg, A.L. Protein degradation and the generation of MHC class I-presented peptides. Adv. Immunol 2002, 80, 1–70. [Google Scholar]
- Kruger, E.; Kuckelkorn, U.; Sijts, A.; Kloetzel, P.M. The components of the proteasome system and their role in MHC class I antigen processing. Rev. Physiol. Biochem. Pharmacol 2003, 148, 81–104. [Google Scholar]
- Wolf, D.H.; Hilt, W. The proteasome: A proteolytic nanomachine of cell regulation and waste disposal. Biochim. Biophys. Acta 2004, 1695, 19–31. [Google Scholar]
- Goldberg, A.L. Protein degradation and protection against misfolded or damaged proteins. Nature 2003, 426, 895–899. [Google Scholar]
- Cardozo, T.; Pagano, M. The SCF ubiquitin ligase: Insights into a molecular machine. Nature Rev. Mol. Cell Biol 2004, 5, 739–751. [Google Scholar]
- Chung, D.W.; Ponts, N.; Prudhomme, J.; Rodrigues, E.M.; Le Roch, K.G. Characterization of the ubiquitylating components of the human malaria parasite’s protein degradation pathway. PLoS One 2012, 7, e43477. [Google Scholar]
- Ponder, E.L.; Bogyo, M. Ubiquitin-like modifiers and their deconjugating enzymes in medically important parasitic protozoa. Eukaryot. Cell 2007, 6, 1943–1952. [Google Scholar]
- Ponts, N.; Yang, J.; Chung, D.W.; Prudhomme, J.; Girke, T.; Horrocks, P.; Le Roch, K.G. Deciphering the ubiquitin-mediated pathway in apicomplexan parasites: A potential strategy to interfere with parasite virulence. PLoS One 2008, 3, e2386. [Google Scholar]
- Schneider, A.; Haas, S.L.; Hildenbrand, R.; Siegmund, S.; Reinhard, I.; Nakovics, H.; Singer, M.V.; Feick, P. Enhanced expression of interleukin-18 in serum and pancreas of patients with chronic pancreatitis. World J. Gastroenterol 2006, 12, 6507–6514. [Google Scholar]
- Dharmasiri, N.; Dharmasiri, S.; Estelle, M. The F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 441–445. [Google Scholar]
- Kepinski, S.; Leyser, O. The Arabidopsis F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 446–451. [Google Scholar]
- Murch, S.; Saxena, P.K. Melatonin: A potential regulator of plant growth and development? In Vitro Cell Dev. Biol. Plant 2002, 38, 531–536. [Google Scholar]
- Schroder, R.; Abele, C.; Gohr, P.; Stuhlfauth-Roisch, U.; Grosse, W. Latest on enzymology of serotonin biosynthesis in walnut seeds. In Tryptophan, Seronin and Melatonin: Basic Aspects and Applications; Huether, G., Kochen, W., Simat, T.J., Steinhart, H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; Volume 467, pp. 637–644. [Google Scholar]
- Koyama, F.C.; Ribeiro, R.Y.; Garcia, J.L.; Azevedo, M.F.; Chakrabarti, D.; Garcia, C.R. Ubiquitin proteasome system and the atypical kinase PfPK7 are involved in melatonin signaling in Plasmodium falciparum. J. Pineal. Res 2012, 165, 1–7. [Google Scholar]
- Dorin, D.; Semblat, J.P.; Poullet, P.; Alano, P.; Goldring, J.P.; Whittle, C.; Patterson, S.; Chakrabarti, D.; Doerig, C. PfPK7, an atypical MEK-related protein kinase, reflects the absence of classical three-component MAPK pathways in the human malaria parasite Plasmodium falciparum. Mol. Microbiol 2005, 55, 184–196. [Google Scholar]
- Doerig, C.; Abdi, A.; Bland, N.; Eschenlauer, S.; Dorin-Semblat, D.; Fennell, C.; Halbert, J.; Holland, Z.; Nivez, M.P.; Semblat, J.P.; et al. Malaria: Targeting parasite and host cell kinomes. Biochim. Biophys. Acta 2010, 1804, 604–612. [Google Scholar]
- Chang, C.F.; Huang, H.J.; Lee, H.C.; Hung, K.C.; Wu, R.T.; Lin, A.M. Melatonin attenuates kainic acid-induced neurotoxicity in mouse hippocampus via inhibition of autophagy and alpha-synuclein aggregation. J. Pineal Res 2012, 52, 312–321. [Google Scholar]
- Luchetti, F.; Canonico, B.; Betti, M.; Arcangeletti, M.; Pilolli, F.; Piroddi, M.; Canesi, L.; Papa, S.; Galli, F. Melatonin signaling and cell protection function. FASEB J 2010, 24, 3603–3624. [Google Scholar]
- Tomas-Zapico, C.; Coto-Montes, A. A proposed mechanism to explain the stimulatory effect of melatonin on antioxidative enzymes. J. Pineal. Res 2005, 39, 99–104. [Google Scholar]
- Yoo, Y.M.; Jung, E.M.; Choi, K.C.; Jeung, E.B. Effect of melatonin on mRNA expressions of transcription factors in murine embryonic stem cells. Brain Res 2011, 1385, 1–7. [Google Scholar]
- Lima, W.R.; Moraes, M.; Alves, E.; Azevedo, M.F.; Passos, D.O.; Garcia, C.R. The PfNF-YB transcription factor is a downstream target of melatonin and cAMP signalling in the human malaria parasite Plasmodium falciparum. J. Pineal. Res 2013, 54, 145–153. [Google Scholar]
- Dorn, A.; Bollekens, J.; Staub, A.; Benoist, C.; Mathis, D. A multiplicity of CCAAT box-binding proteins. Cell 1987, 50, 863–872. [Google Scholar]
- Maity, S.N.; de Crombrugghe, B. Role of the CCAAT-binding protein CBF/NF-Y in transcription. Trends Biochem. Sci 1998, 23, 174–178. [Google Scholar]
- Kao, C.Y.; Tanimoto, A.; Arima, N.; Sasaguri, Y.; Padmanabhan, R. Transactivation of the human CDC2 promoter by adenovirus E1A. E1A induces the expression and assembly of a heteromeric complex consisting of the CCAAT box binding factor, CBF/NF-Y, and a 110-kDa DNA-binding protein. J. Biol. Chem 1999, 274, 23043–23051. [Google Scholar]
- Hu, Q.; Bhattacharya, C.; Maity, S.N. CCAAT binding factor (CBF) binding mediates cell cycle activation of topoisomerase IIalpha. Conventional CBF activation domains are not required. J. Biol. Chem 2002, 277, 37191–37200. [Google Scholar]
- Klein, D.C.; Coon, S.L.; Roseboom, P.H.; Weller, J.L.; Bernard, M.; Gastel, J.A.; Zatz, M.; Iuvone, P.M.; Rodriguez, I.R.; Begay, V.; et al. The melatonin rhythm-generating enzyme: Molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog. Horm. Res 1997, 52, 307–357, ;. [Google Scholar]
- Johnston, J.D.; Bashforth, R.; Diack, A.; Andersson, H.; Lincoln, G.A.; Hazlerigg, D.G. Rhythmic melatonin secretion does not correlate with the expression of arylalkylamine N-acetyltransferase, inducible cyclic amp early repressor, period1 or cryptochrome1 mRNA in the sheep pineal. Neuroscience 2004, 124, 789–795. [Google Scholar]
- Sugden, L.A.; Sugden, D.; Klein, D.C. Alpha 1-adrenoceptor activation elevates cytosolic calcium in rat pinealocytes by increasing net influx. J. Biol. Chem 1987, 262, 741–745. [Google Scholar]
- Schomerus, C.; Laedtke, E.; Korf, H.W. Calcium responses of isolated, immunocytochemically identified rat pinealocytes to noradrenergic, cholinergic and vasopressinergic stimulations. Neurochem. Int 1995, 27, 163–175. [Google Scholar]
- Gastel, J.A.; Roseboom, P.H.; Rinaldi, P.A.; Weller, J.L.; Klein, D.C. Melatonin production: Proteasomal proteolysis in serotonin N-acetyltransferase regulation. Science 1998, 279, 1358–1360. [Google Scholar]
- Srinivasan, V.; Spence, D.W.; Moscovitch, A.; Pandi-Perumal, S.R.; Trakht, I.; Brown, G.M.; Cardinali, D.P. Malaria: Therapeutic implications of melatonin. J. Pineal. Res 2010, 48, 1–8. [Google Scholar]
- Reppert, S.M.; Weaver, D.R.; Ebisawa, T. Cloning and characterization of a mammalian melatonin receptor that mediates reproductive and circadian responses. Neuron 1994, 13, 1177–1185. [Google Scholar]
- Reppert, S.M.; Godson, C.; Mahle, C.D.; Weaver, D.R.; Slaugenhaupt, S.A.; Gusella, J.F. Molecular characterization of a second melatonin receptor expressed in human retina and brain: The Mel1b melatonin receptor. Proc. Natl. Acad. Sci. USA 1995, 92, 8734–8738. [Google Scholar]
- Reppert, S.M.; Weaver, D.R.; Ebisawa, T.; Mahle, C.D.; Kolakowski, L.F., Jr. Cloning of a melatonin-related receptor from human pituitary. FEBS Lett. 1996, 386, 219–224. [Google Scholar]
- Levoye, A.; Dam, J.; Ayoub, M.A.; Guillaume, J.L.; Couturier, C.; Delagrange, P.; Jockers, R. The orphan GPR50 receptor specifically inhibits MT1 melatonin receptor function through heterodimerization. EMBO J 2006, 25, 3012–3023. [Google Scholar]
- Dubocovich, M.L.; Yun, K.; Al-Ghoul, W.M.; Benloucif, S.; Masana, M.I. Selective MT2 melatonin receptor antagonists block melatonin-mediated phase advances of circadian rhythms. FASEB J 1998, 12, 1211–1220. [Google Scholar]
- Smirnov, A.N. Nuclear melatonin receptors. Biochem. Biokhimiia 2001, 66, 19–26. [Google Scholar]
- Garcia-Perganeda, A.; Guerrero, J.M.; Rafii-El-Idrissi, M.; Paz Romero, M.; Pozo, D.; Calvo, J.R. Characterization of membrane melatonin receptor in mouse peritoneal macrophages: Inhibition of adenylyl cyclase by a pertussis toxin-sensitive G protein. J. Neuroimmunol 1999, 95, 85–94. [Google Scholar]
- Browning, C.; Beresford, I.; Fraser, N.; Giles, H. Pharmacological characterization of human recombinant melatonin MT(1) and MT(2) receptors. Br. J. Pharmacol 2000, 129, 877–886. [Google Scholar]
- Dubocovich, M.L.; Cardinali, D.P.; Guardiola-Lemaitre, B.; Hagan, R.M.; Krause, D.N.; Sugden, D. Melatonin receptors. In The IUPHAR Compendium of Receptor Characterization and Classification, 2nd ed; IUPHAR Media: London, UK, 2000; pp. 271–277. [Google Scholar]
- Bagnaresi, P.; Markus, R.P.; Hotta, C.T.; Pozzan, T.; Garcia, C.R. Desynchronizing Plasmodium cell cycle increases chloroquine protection at suboptimal doses. Open Parasitol. J 2008, 2, 55–58. [Google Scholar]
- Gazarini, M.L.; Garcia, C.R. The malaria parasite mitochondrion senses cytosolic Ca2+ fluctuations. Biochem. Biophys. Res. Commun 2004, 321, 138–144. [Google Scholar]
- Guha, M.; Maity, P.; Choubey, V.; Mitra, K.; Reiter, R.J.; Bandyopadhyay, U. Melatonin inhibits free radical-mediated mitochondrial-dependent hepatocyte apoptosis and liver damage induced during malarial infection. J. Pineal. Res 2007, 43, 372–381. [Google Scholar]
- Bagnaresi, P.; Alves, E.; da Silva, H.B.; Epiphanio, S.; Mota, M.M.; Garcia, C.R. Unlike the synchronous Plasmodium falciparum and P. chabaudi infection, the P. berghei and P. yoelii asynchronous infections are not affected by melatonin. Int. J. Gen. Med 2009, 2, 47–55. [Google Scholar]
- O’Donnell, A.J.; Schneider, P.; McWatters, H.G.; Reece, S.E. Fitness costs of disrupting circadian rhythms in malaria parasites. Proc. Biol. Sci 2011, 278, 2429–2436. [Google Scholar]
- Mideo, N.; Reece, S.E.; Smith, A.L.; Metcalf, C.J. The Cinderella syndrome: Why do malaria-infected cells burst at midnight? Trends Parasitol 2013, 29, 10–16. [Google Scholar]
- Srinivasan, V.; Mohamed, M.; Zakaria, R.; Ahmad, A.H. Malaria, anti malarial drugs and the role of melatonin. Infect. Disord. Drug Targets 2012, 12, 371–379. [Google Scholar]
- Srinivasan, V.; Ahmad, A.H.; Mohamed, M.; Zakaria, R. Melatonin effects on Plasmodium life cycle: New avenues for therapeutic approach. Recent Pat. Endocr. Metab. Immune Drug Discov 2012, 6, 139–147. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lima, W.R.; Holder, A.A.; Garcia, C.R.S. Melatonin Signaling and Its Modulation of PfNF-YB Transcription Factor Expression in Plasmodium falciparum. Int. J. Mol. Sci. 2013, 14, 13704-13718. https://doi.org/10.3390/ijms140713704
Lima WR, Holder AA, Garcia CRS. Melatonin Signaling and Its Modulation of PfNF-YB Transcription Factor Expression in Plasmodium falciparum. International Journal of Molecular Sciences. 2013; 14(7):13704-13718. https://doi.org/10.3390/ijms140713704
Chicago/Turabian StyleLima, Wânia Rezende, Anthony A. Holder, and Célia R. S. Garcia. 2013. "Melatonin Signaling and Its Modulation of PfNF-YB Transcription Factor Expression in Plasmodium falciparum" International Journal of Molecular Sciences 14, no. 7: 13704-13718. https://doi.org/10.3390/ijms140713704