Achillea millefolium L. Essential Oil Inhibits LPS-Induced Oxidative Stress and Nitric Oxide Production in RAW 264.7 Macrophages


Abstract
:1. Introduction
2. Results and Discussion
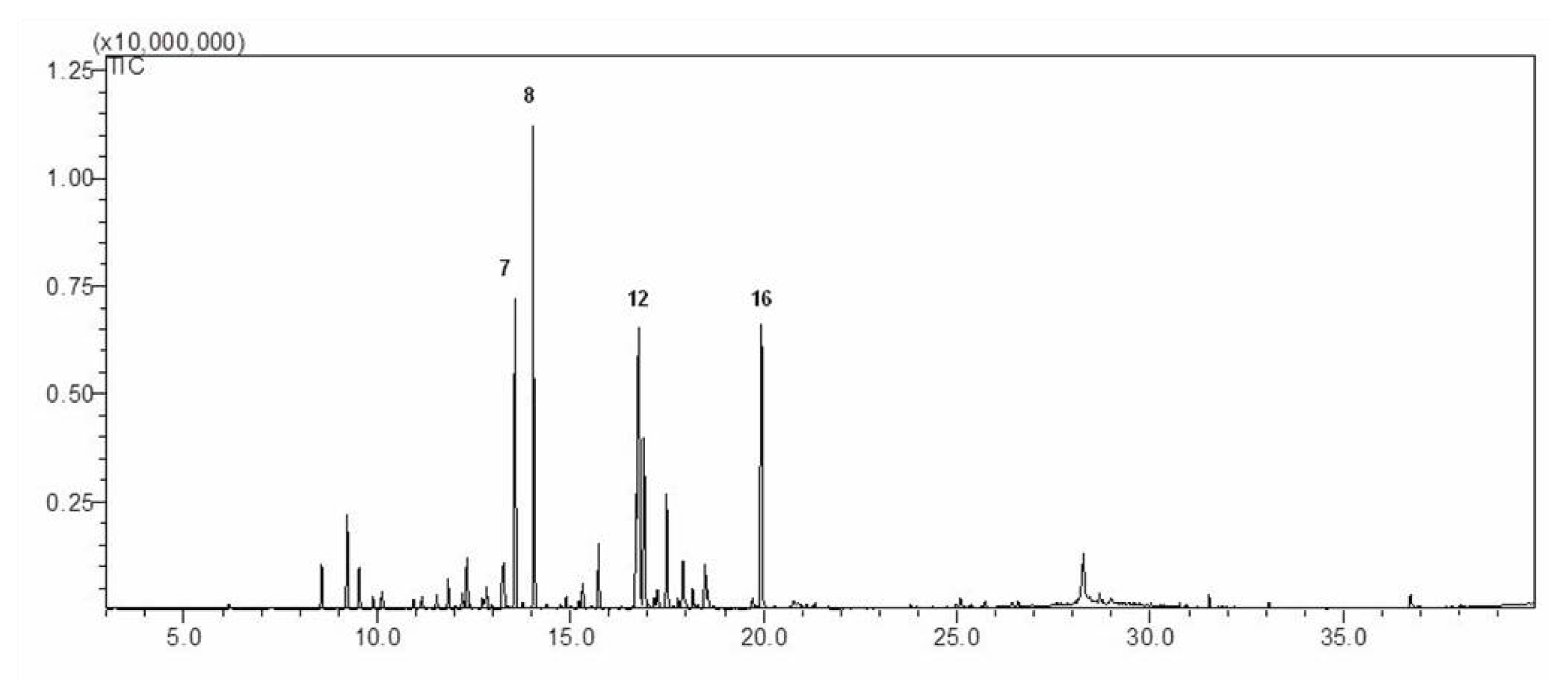
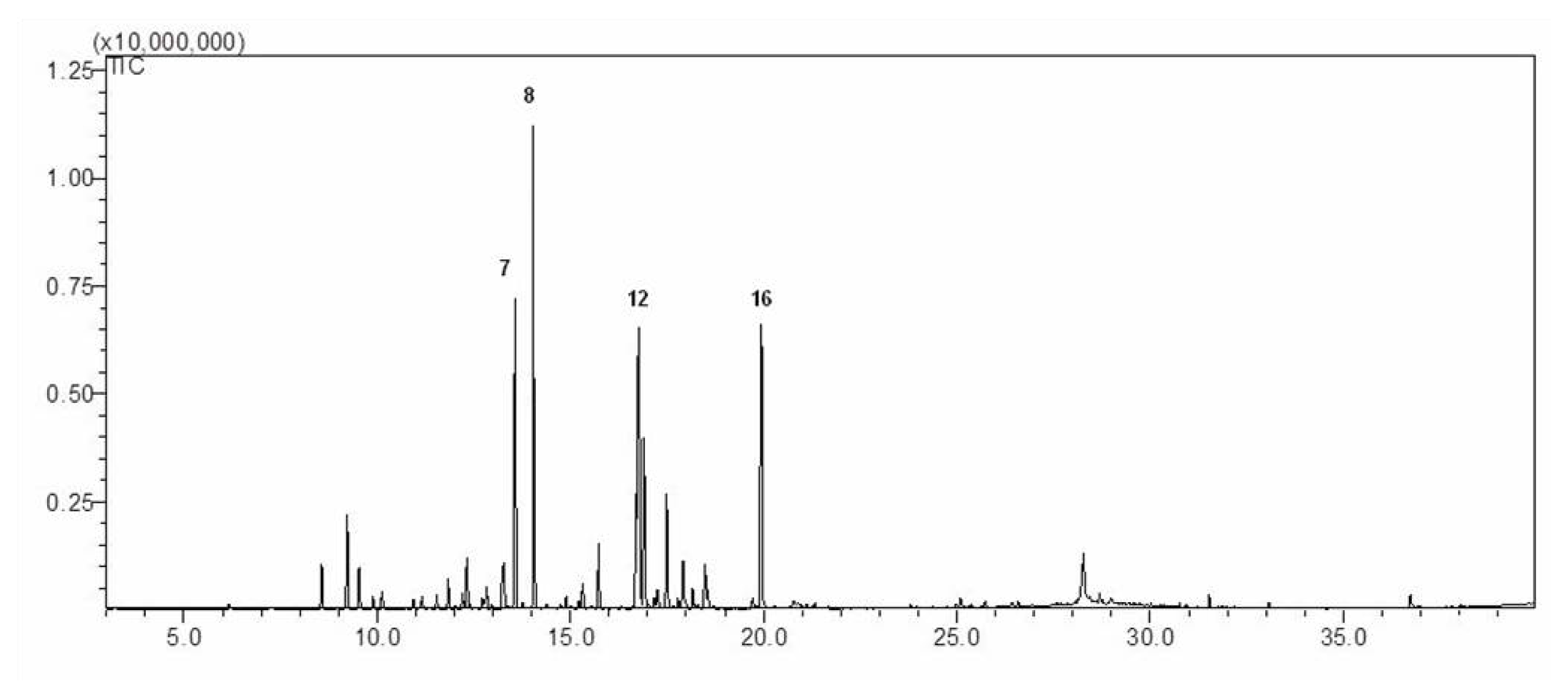
2.1. Chemical Composition of AM-EO
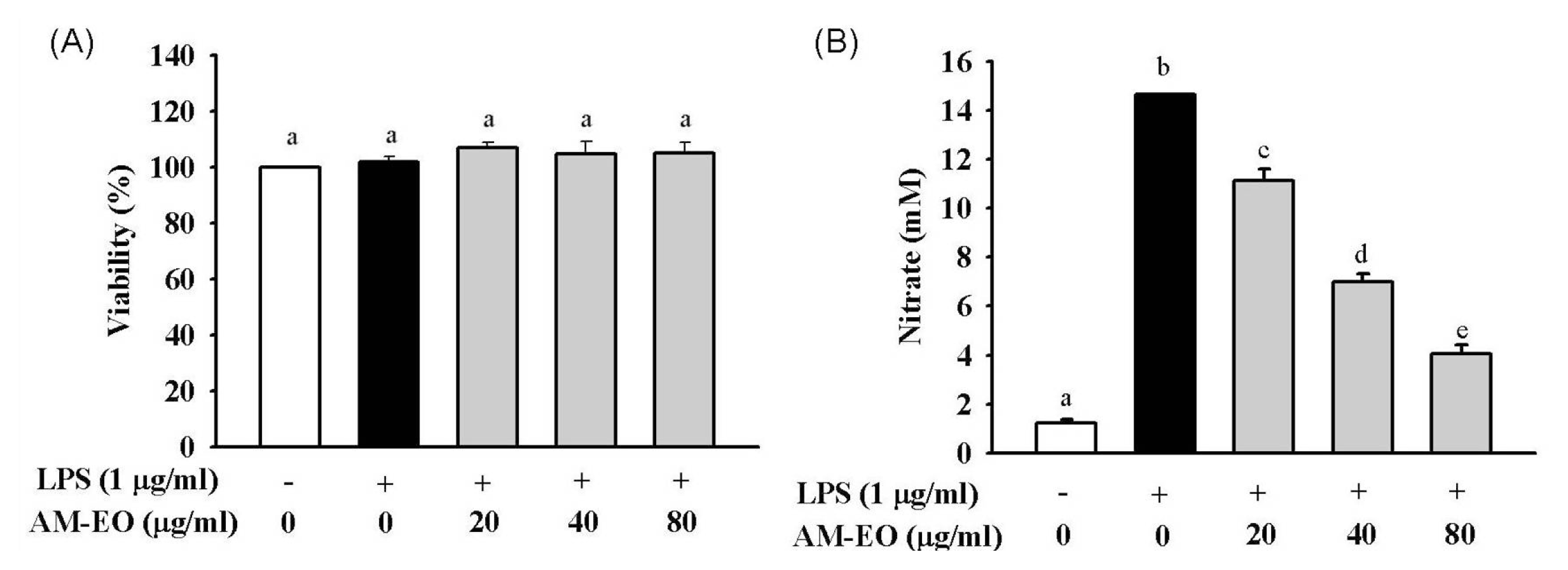
2.2. Effects of AM-EO on Cell Viability and NO Production
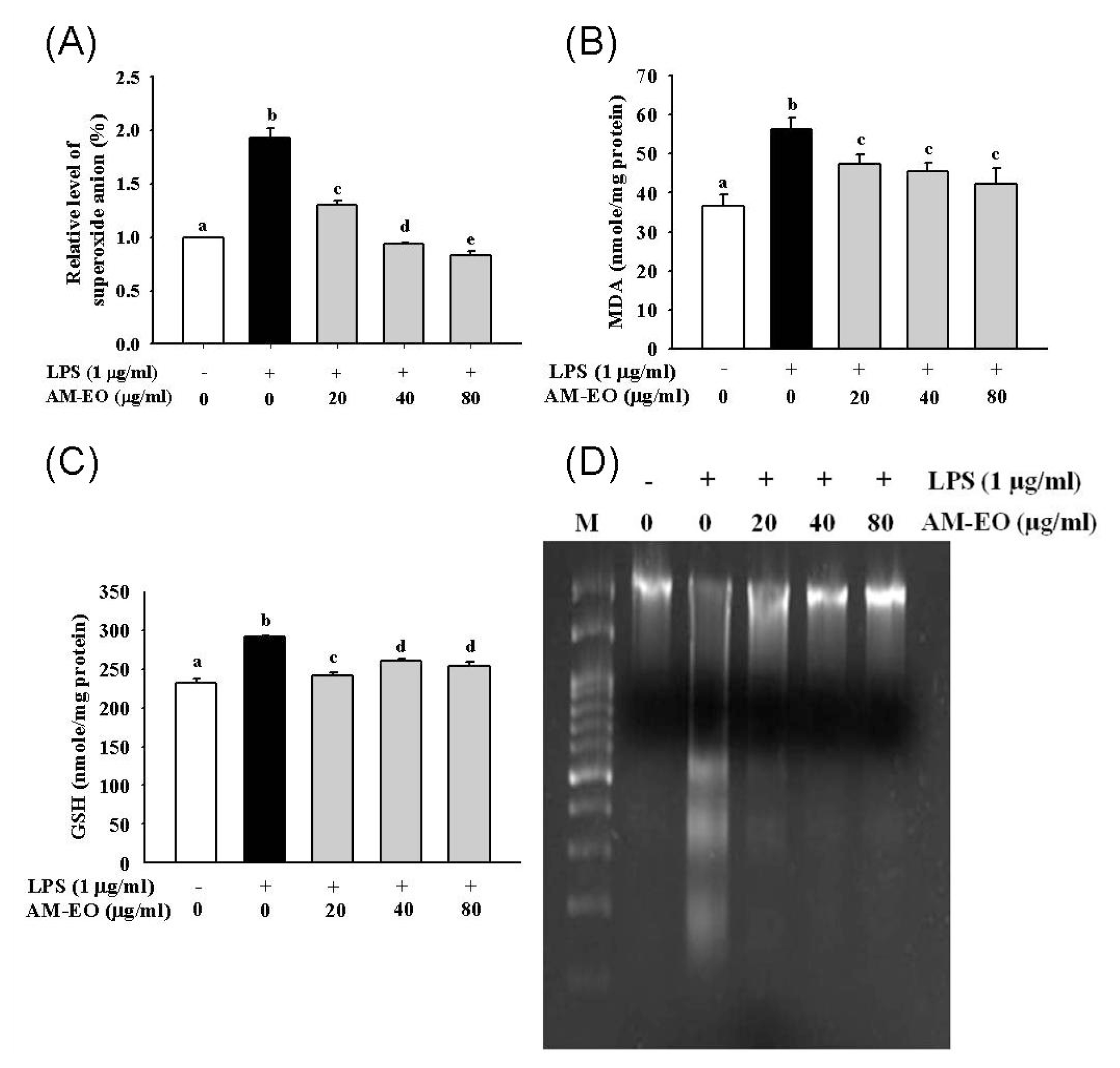
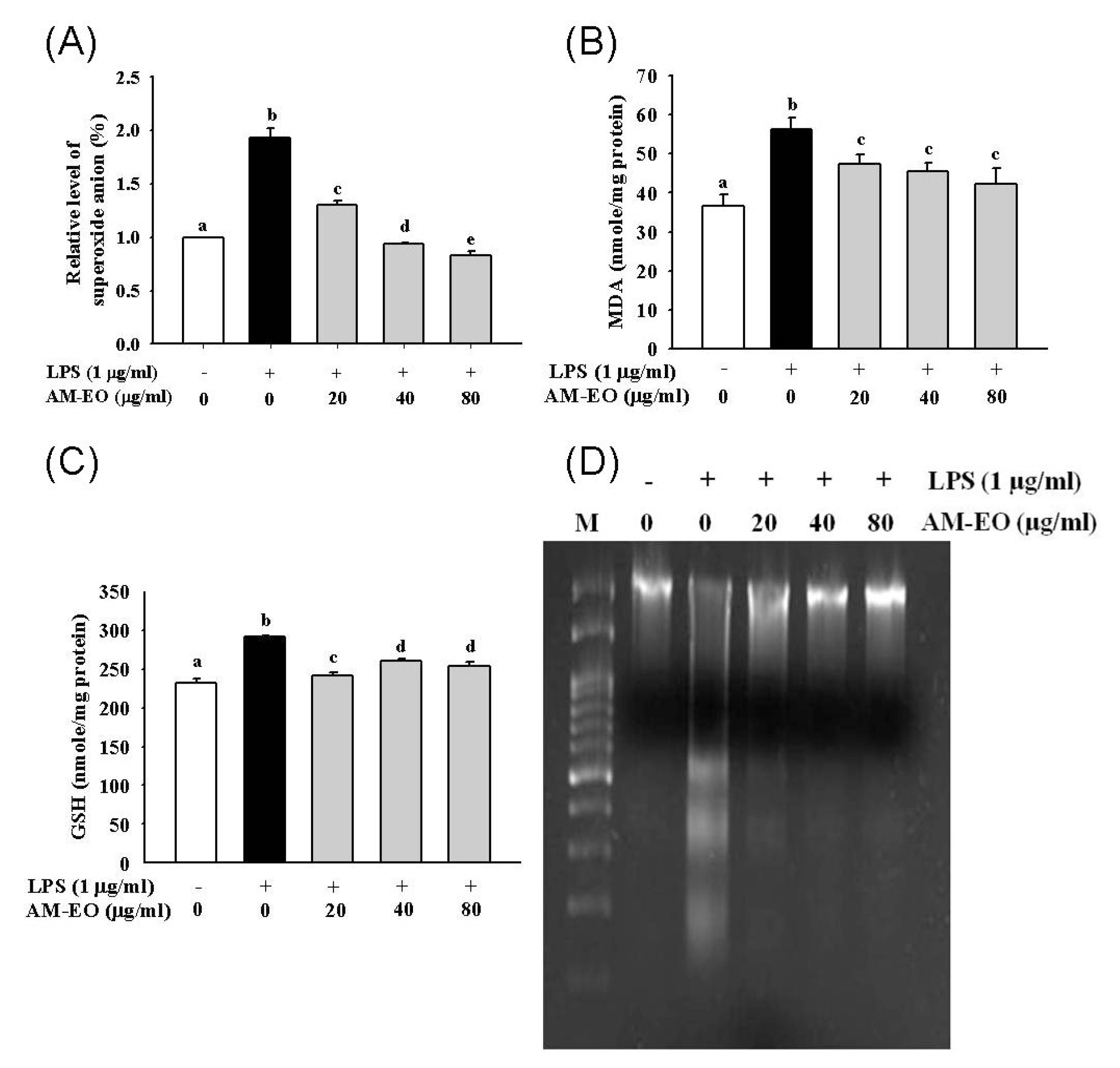
2.3. The Effect of AM-EO on Superoxide Anion and Malondialdehyde (MDA) Production, GSH Concentration and LPS-Induced DNA Damage
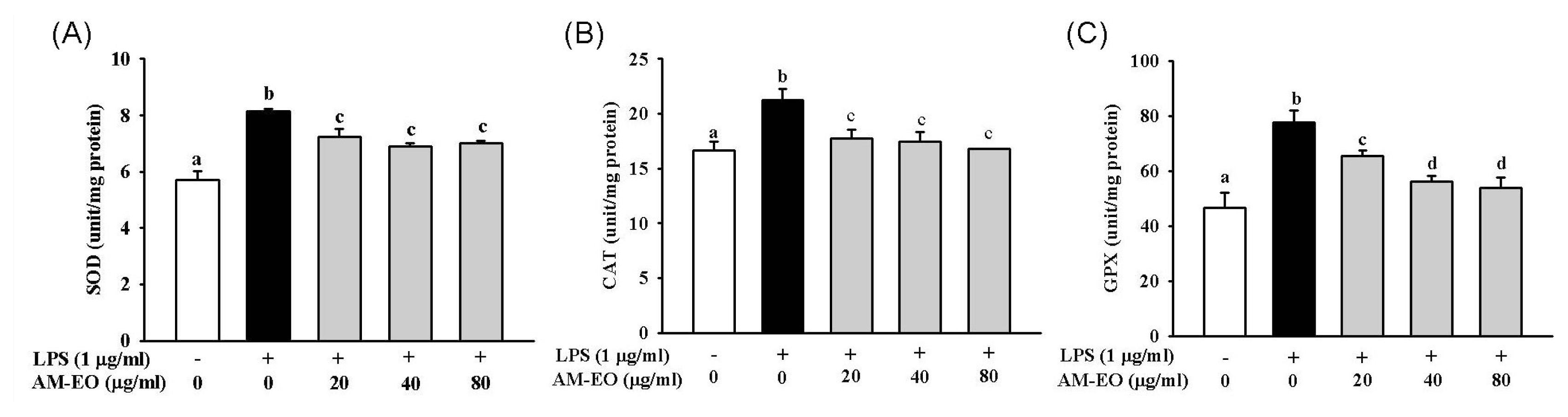
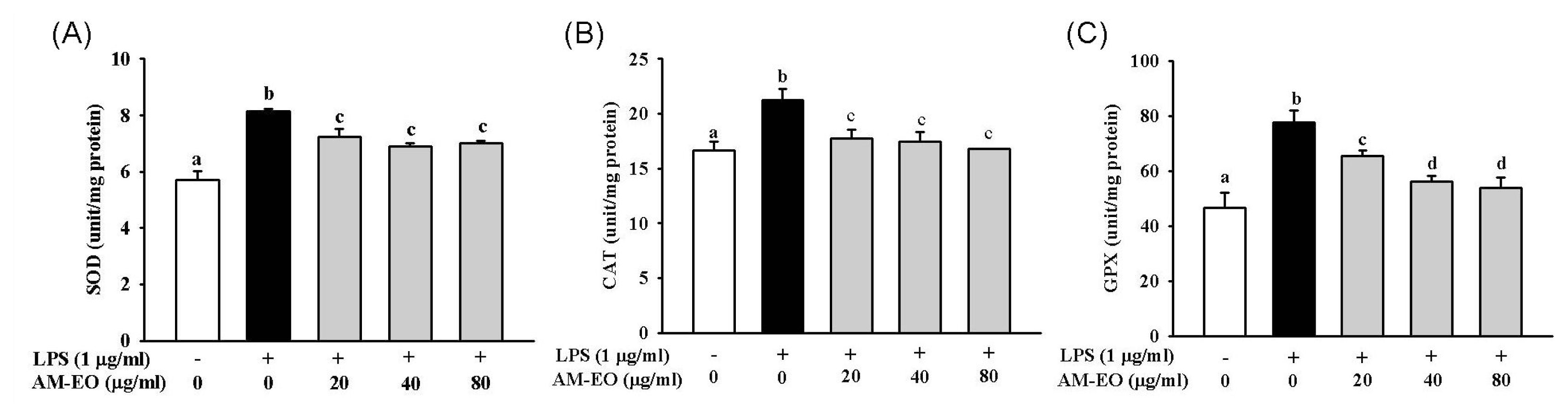
2.4. The Effect of AM-EO on Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPx) Activities
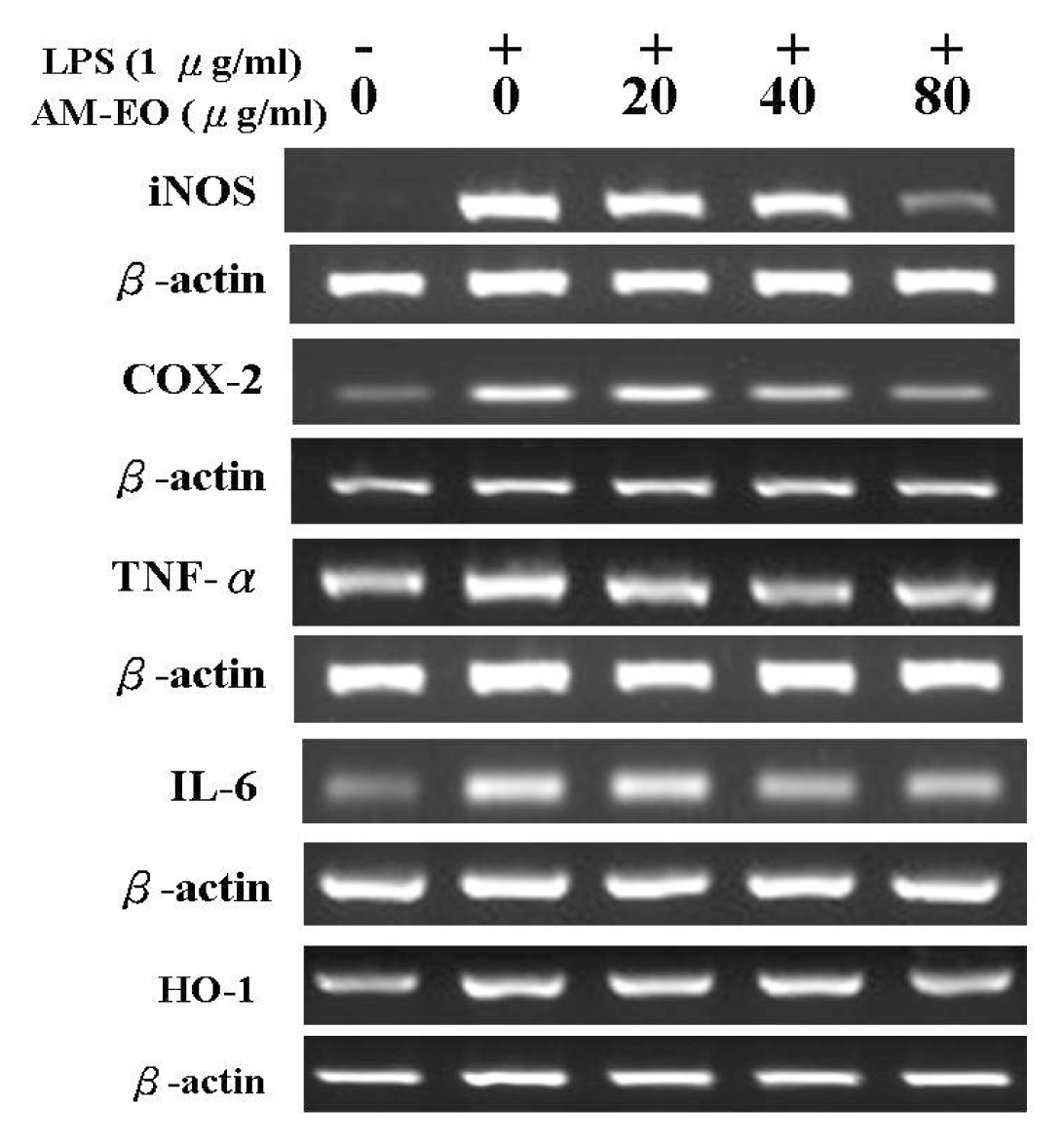
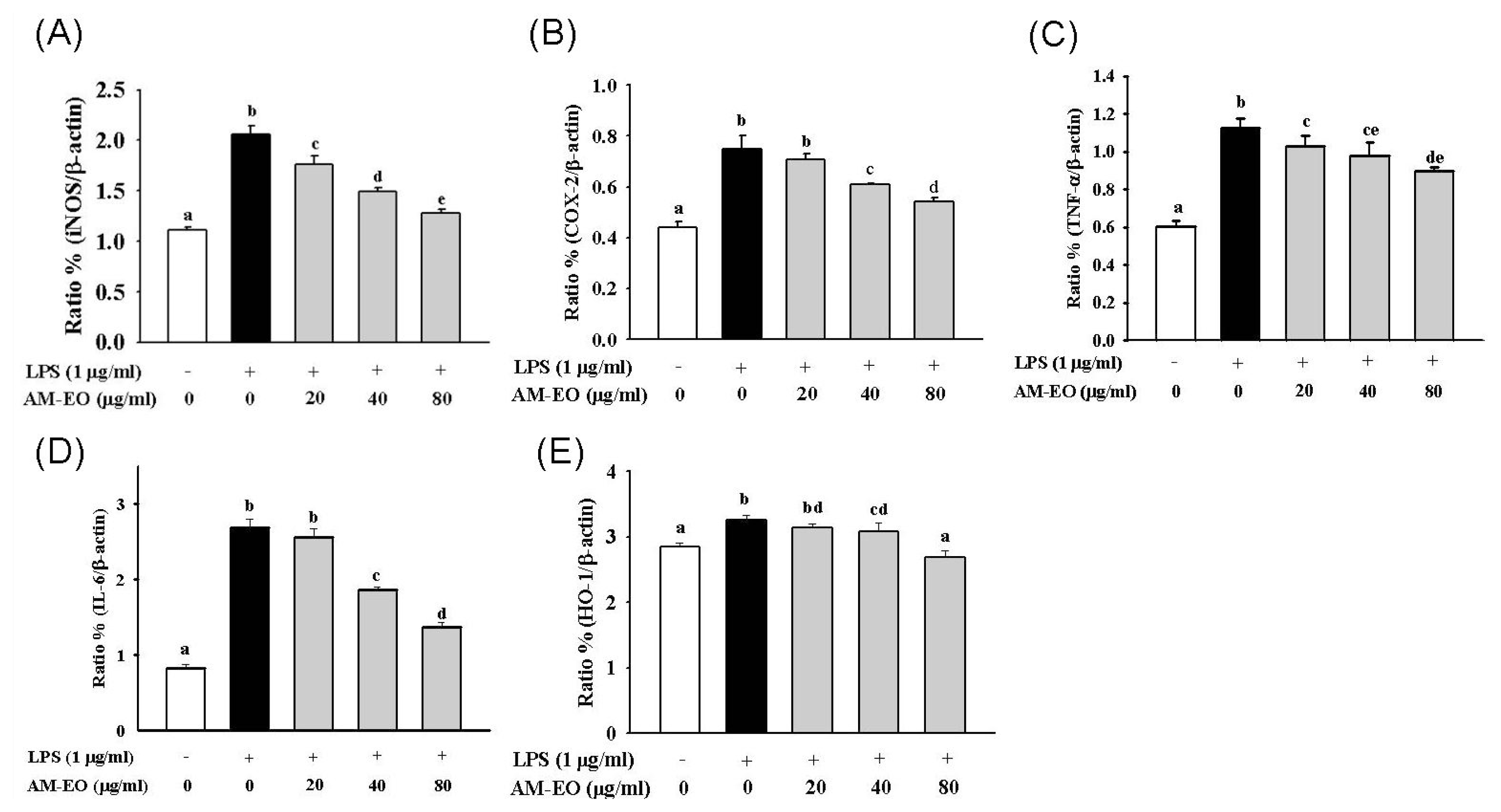
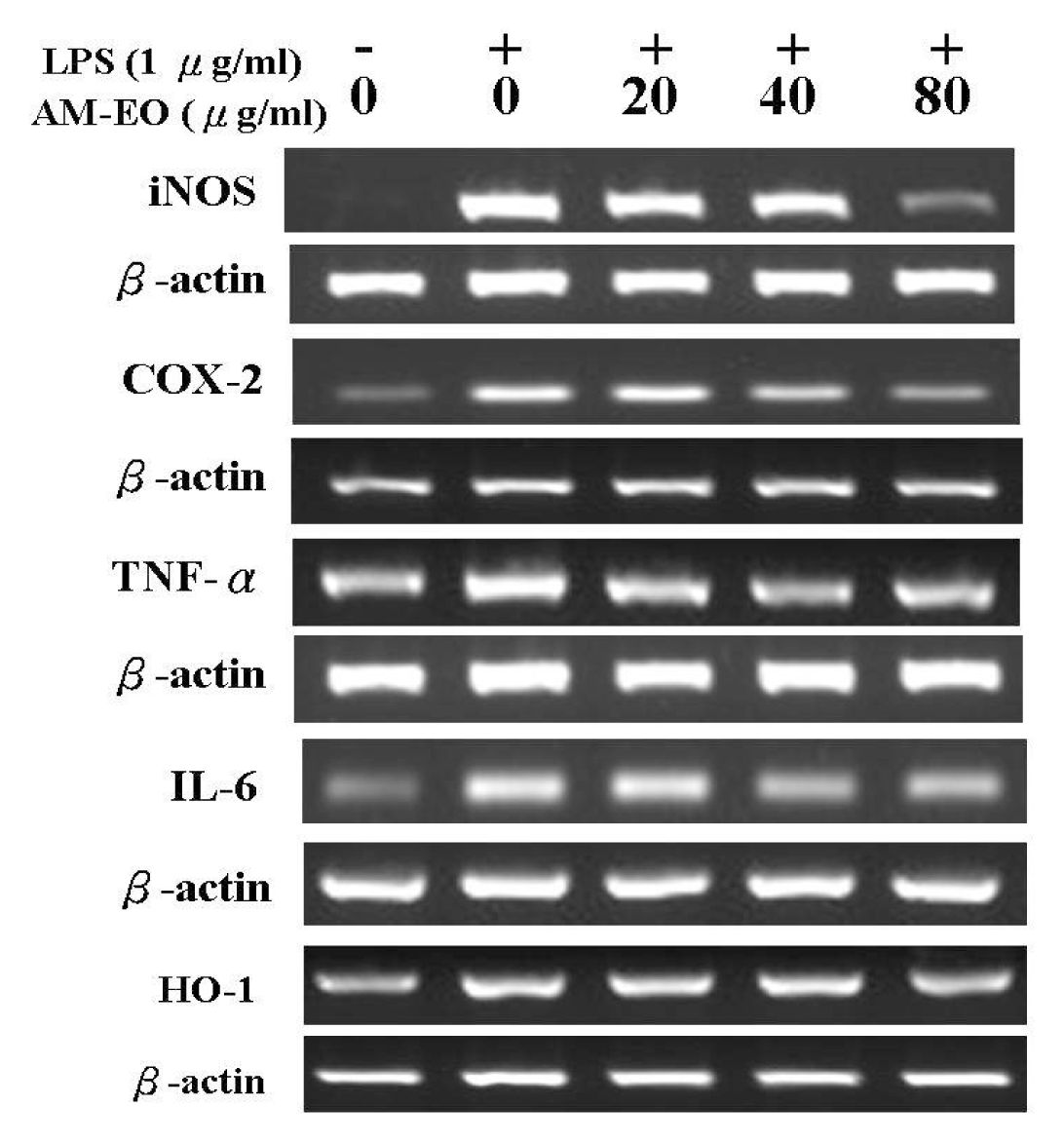
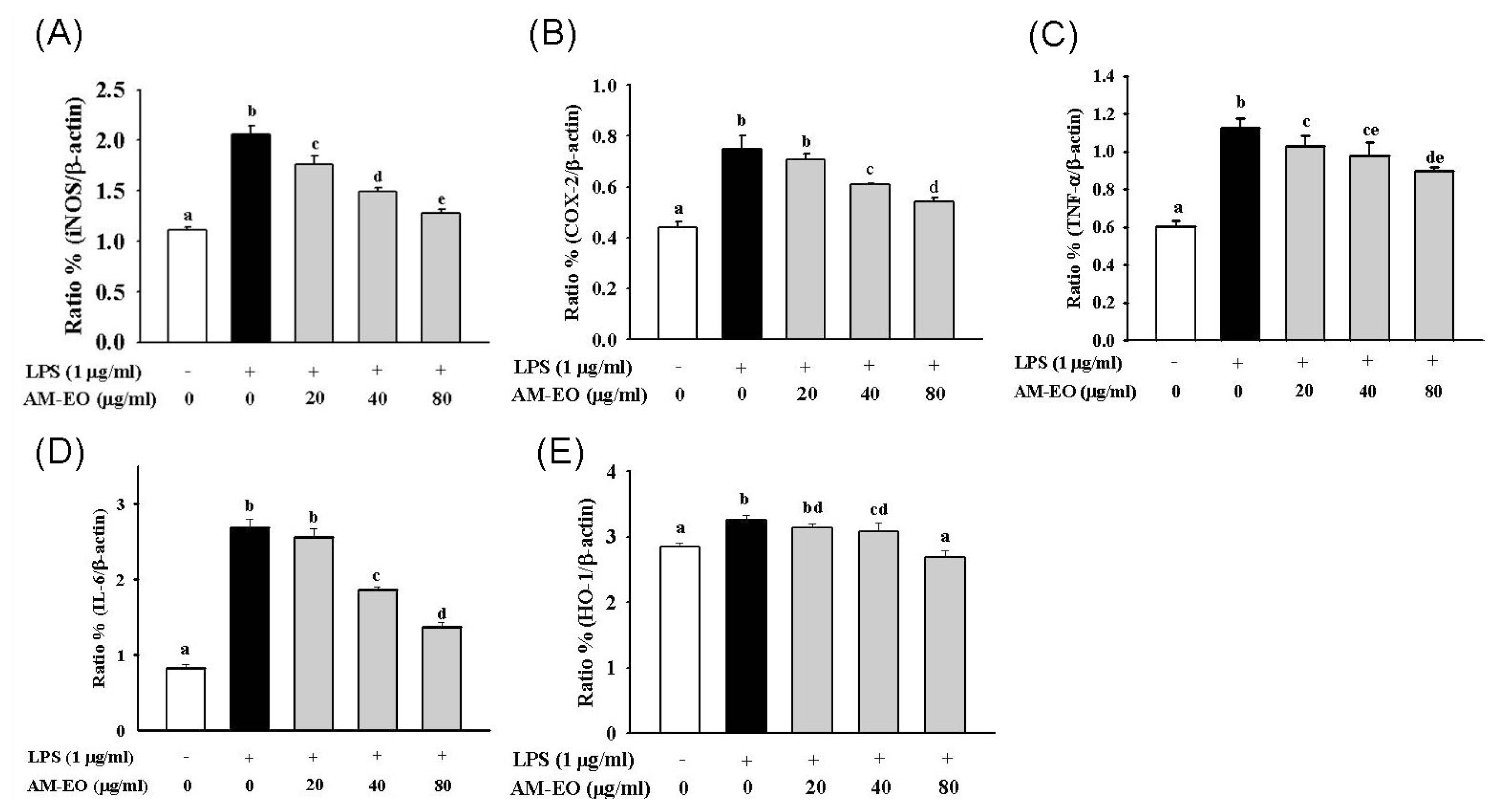
2.5. The Effects of AM-EO on the Expression Levels of iNOS, COX-2, TNF-α, IL-6 and HO-1 mRNA
3. Experimental Section
3.1. Essential Oil and Cell Line
3.2. Materials
3.3. Gas Chromatography and Mass Spectrometry Analysis
3.4. Cell Culture
3.5. Cell Viability
3.6. Nitrite Production
3.7. Measurement of Superoxide Anion Production, Lipid Peroxide and Glutathione (GSH) Levels
3.8. DNA Fragmentation Assay
3.9. Measurement of Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Peroxidase (GPx) Activity
3.10. Measurement of Messenger RNA (mRNA) Levels of Inducible nitric Oxide Synthase (iNOS), Cyclooxygenase-2 (COX-2), Tumour Necrosis Factor-α (TNF-α), Interleukin-6 (IL-6) and Heme Oxygenase-1 (HO-1)
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Ran, S.; Montgomery, K.E. Macrophage-mediated lymphangiogenesis: The emerging role of macrophages as lymphatic endothelial progenitors. Cancers 2012, 4, 618–657. [Google Scholar]
- Heo, S.J.; Yoon, W.J.; Kim, K.N.; Oh, C.; Choi, Y.U.; Yoon, K.T.; Kang, D.H.; Qian, Z.J.; Choi, I.W.; Jung, W.K. Anti-inflammatory effect of fucoxanthin derivatives isolated from Sargassum siliquastrum in lipopolysaccharide-stimulated RAW 264.7 macrophage. Food Chem. Toxicol 2012, 50, 3336–3342. [Google Scholar]
- Rossol, M.; Heine, H.; Meusch, U.; Quandt, D.; Klein, C.; Sweet, M.J.; Hauschildt, S. LPS-induced cytokine production in human monocytes and macrophages. Crit. Rev. Immunol 2011, 31, 379–446. [Google Scholar]
- Ayroldi, E.; Bastianelli, A.; Cannarile, L.; Petrillo, M.G.; Delfino, D.V.; Fierabracci, A. A pathogenetic approach to autoimmune skin disease therapy: Psoriasis and biological drugs, unresolved issues, and future directions. Curr. Pharm. Des 2011, 17, 3176–3190. [Google Scholar]
- Vitalini, S.; Beretta, G.; Iriti, M.; Orsenigo, S.; Basilico, N.; Dall’Acqua, S.; Iorizzi, M.; Fico, G. Phenolic compounds from Achillea millefolium L. and their bioactivity. Acta Biochim. Pol 2011, 58, 203–209. [Google Scholar]
- Khan, A.U.; Gilani, A.H. Blood pressure lowering, cardiovascular inhibitory and bronchodilatory actions of Achillea millefolium. Phytother. Res 2011, 25, 577–583. [Google Scholar]
- Benedek, B.; Kopp, B. Achillea millefolium L. s.l. revisited: Recent findings confirm the traditional use. Wien. med. Wochenschr 2007, 157, 312–314. [Google Scholar]
- Cavalcanti, A.M.; Baggio, C.H.; Freitas, C.S.; Rieck, L.; de Sousa, R.S.; da Silva-Santos, J.E.; Mesia-Vela, S.; Marques, M.C. Safety and antiulcer efficacy studies of Achillea millefolium L. after chronic treatment in Wistar rats. J. Ethnopharmacol 2006, 107, 277–284. [Google Scholar]
- Benedek, B.; Kopp, B.; Melzig, M.F. Achillea millefolium L. s.l.—Is the anti-inflammatory activity mediated by protease inhibition? J. Ethnopharmacol 2007, 113, 312–317. [Google Scholar]
- Candan, F.; Unlu, M.; Tepe, B.; Daferera, D.; Polissiou, M.; Sokmen, A.; Akpulat, H.A. Antioxidant and antimicrobial activity of the essential oil and methanol extracts of Achillea millefolium subsp. millefolium Afan. (Asteraceae). J. Ethnopharmacol 2003, 87, 215–220. [Google Scholar]
- Konyalioglu, S.; Karamenderes, C. The protective effects of Achillea L. species native in Turkey against H2O2-induced oxidative damage in human erythrocytes and leucocytes. J. Ethnopharmacol 2005, 102, 221–227. [Google Scholar]
- Stojanovic, G.; Radulovic, N.; Hashimoto, T.; Palic, R. In vitro antimicrobial activity of extracts of four Achillea species: The composition of Achillea clavennae L. (Asteraceae) extract. J. Ethnopharmacol 2005, 101, 185–190. [Google Scholar]
- Potrich, F.B.; Allemand, A.; da Silva, L.M.; Dos Santos, A.C.; Baggio, C.H.; Freitas, C.S.; Mendes, D.A.; Andre, E.; Werner, M.F.; Marques, M.C. Antiulcerogenic activity of hydroalcoholic extract of Achillea millefolium L.: Involvement of the antioxidant system. J. Ethnopharmacol 2010, 130, 85–92. [Google Scholar]
- Innocenti, G.; Vegeto, E.; Dall’Acqua, S.; Ciana, P.; Giorgetti, M.; Agradi, E.; Sozzi, A.; Fico, G.; Tome, F. In vitro estrogenic activity of Achillea millefolium L. Phytomedicine 2007, 14, 147–152. [Google Scholar]
- Tozyo, T.; Yoshimura, Y.; Sakurai, K.; Uchida, N.; Takeda, Y.; Nakai, H.; Ishii, H. Novel antitumor sesquiterpenoids in Achillea millefolium. Chem. Pharm. Bull. (Tokyo) 1994, 42, 1096–1100. [Google Scholar]
- Csupor-Loffler, B.; Hajdu, Z.; Zupko, I.; Rethy, B.; Falkay, G.; Forgo, P.; Hohmann, J. Antiproliferative effect of flavonoids and sesquiterpenoids from Achillea millefolium s.l. on cultured human tumour cell lines. Phytother. Res 2009, 23, 672–676. [Google Scholar]
- Lin, L.T.; Liu, L.T.; Chiang, L.C.; Lin, C.C. In vitro anti-hepatoma activity of fifteen natural medicines from Canada. Phytother. Res 2002, 16, 440–444. [Google Scholar]
- Li, Y.; Zhang, M.L.; Cong, B.; Wang, S.M.; Dong, M.; Sauriol, F.; Huo, C.H.; Shi, Q.W.; Gu, Y.C.; Kiyota, H. Achillinin A, a cytotoxic guaianolide from the flower of Yarrow, Achillea millefolium. Biosci. Biotechnol. Biochem 2011, 75, 1554–1556. [Google Scholar]
- Bozin, B.; Mimica-Dukic, N.; Bogavac, M.; Suvajdzic, L.; Simin, N.; Samojlik, I.; Couladis, M. Chemical composition, antioxidant and antibacterial properties of Achillea collina Becker ex Heimerl s.l. and A. pannonica Scheele essential oils. Molecules 2008, 13, 2058–2068. [Google Scholar]
- Conti, B.; Canale, A.; Bertoli, A.; Gozzini, F.; Pistelli, L. Essential oil composition and larvicidal activity of six Mediterranean aromatic plants against the mosquito Aedes albopictus (Diptera: Culicidae). Parasitol. Res 2010, 107, 1455–1461. [Google Scholar]
- Orav, A.; Arak, E.; Raal, A. Phytochemical analysis of the essential oil of Achillea millefolium L. from various European Countries. Nat. Prod. Res 2006, 20, 1082–1088. [Google Scholar]
- Okoye, F.B.; Osadebe, P.O.; Nworu, C.S.; Okoye, N.N.; Omeje, E.O.; Esimone, C.O. Topical anti-inflammatory constituents of lipophilic leaf fractions of Alchornea floribunda and Alchornea cordifolia. Nat. Prod. Res 2011, 25, 1941–1949. [Google Scholar]
- Verma, M.; Singh, S.K.; Bhushan, S.; Pal, H.C.; Kitchlu, S.; Koul, M.K.; Thappa, R.K.; Saxena, A.K. Induction of mitochondrial-dependent apoptosis by an essential oil from Tanacetum gracile. Planta Med 2008, 74, 515–520. [Google Scholar]
- Setzer, W.N.; Vogler, B.; Schmidt, J.M.; Leahy, J.G.; Rives, R. Antimicrobial activity of Artemisia douglasiana leaf essential oil. Fitoterapia 2004, 75, 192–200. [Google Scholar]
- Bastos, V.P.; Gomes, A.S.; Lima, F.J.; Brito, T.S.; Soares, P.M.; Pinho, J.P.; Silva, C.S.; Santos, A.A.; Souza, M.H.; Magalhaes, P.J. Inhaled 1,8-cineole reduces inflammatory parameters in airways of ovalbumin-challenged Guinea pigs. Basic Clin. Pharmacol. Toxicol 2010, 108, 34–39. [Google Scholar]
- Ehrnhöfer-Ressler, M.M.; Fricke, K.; Pignitter, M.; Walker, J.M.; Walker, J.; Rychlik, M.; Somoza, V. Identification of 1,8-cineole, borneol, camphor, and thujone as anti-inflammatory compounds in a Salvia Officinalis L. infusion using human gingival fibroblasts. J. Agr. Food Chem 2013, 61, 3451–3459. [Google Scholar]
- Miguel, M.G. Antioxidant and anti-inflammatory activities of essential oils: A short review. Molecules 2010, 15, 9252–9287. [Google Scholar]
- Peana, A.T.; D’Aquila, P.S.; Panin, F.; Serra, G.; Pippia, P.; Moretti, M.D. Anti-inflammatory activity of linalool and linalyl acetate constituents of essential oils. Phytomedicine 2002, 9, 721–726. [Google Scholar]
- Silva, C.R.; Oliveira, S.M.; Rossato, M.F.; Dalmolin, G.D.; Guerra, G.P.; da Silveira Prudente, A.; Cabrini, D.A.; Otuki, M.F.; Andre, E.; Ferreira, J. The involvement of TRPA1 channel activation in the inflammatory response evoked by topical application of cinnamaldehyde to mice. Life Sci 2011, 88, 1077–1087. [Google Scholar]
- Ahmad, R.; Rasheed, Z.; Ahsan, H. Biochemical and cellular toxicology of peroxynitrite: Implications in cell death and autoimmune phenomenon. Immunopharmacol. Immunotoxicol 2009, 31, 388–396. [Google Scholar]
- Kunwar, A.; Sandur, S.K.; Krishna, M.; Priyadarsini, K.I. Curcumin mediates time and concentration dependent regulation of redox homeostasis leading to cytotoxicity in macrophage cells. Eur. J. Pharmacol 2009, 611, 8–16. [Google Scholar]
- Bak, M.J.; Jun, M.; Jeong, W.S. Antioxidant and hepatoprotective effects of the red ginseng essential oil in H2O2-treated HepG2 cells and CCl4-treated mice. Int. J. Mol. Sci 2012, 13, 2314–2330. [Google Scholar]
- Sreelatha, S.; Inbavalli, R. Antioxidant, antihyperglycemic, and antihyperlipidemic effects of Coriandrum sativum leaf and stem in alloxan-induced diabetic rats. J. Food Sci 2012, 77, T119–T123. [Google Scholar]
- Ben-Shaul, V.; Lomnitski, L.; Nyska, A.; Carbonatto, M.; Peano, S.; Zurovsky, Y.; Bergman, M.; Eldridge, S.R.; Grossman, S. Effect of natural antioxidants and apocynin on LPS-induced endotoxemia in rabbit. Hum. Exp. Toxicol 2000, 19, 604–614. [Google Scholar]
- Chou, S.-T.; Lai, C.-P.; Lin, C.-C.; Shih, Y. Study of the chemical composition, antioxidant activity and anti-inflammatory activity of essential oil from Vetiveria zizanioides. Food Chem 2012, 134, 262–268. [Google Scholar]
- Shyur, L.F.; Huang, C.C.; Hsu, Y.Y.; Cheng, Y.W.; Yang, S.D. A sesquiterpenol extract potently suppresses inflammation in macrophages and mice skin and prevents chronic liver damage in mice through JNK-dependent HO-1 expression. Phytochemistry 2011, 72, 391–399. [Google Scholar]
- Wang, H.; Khor, T.O.; Saw, C.L.; Lin, W.; Wu, T.; Huang, Y.; Kong, A.N. Role of Nrf2 in suppressing LPS-induced inflammation in mouse peritoneal macrophages by polyunsaturated fatty acids docosahexaenoic acid and eicosapentaenoic acid. Mol. Pharmacol 2010, 7, 2185–2193. [Google Scholar]
- Lee, S.-M.; Liu, K.-H.; Liu, Y.-Y.; Chang, Y.-P.; Lin, C.-C.; Chen, Y.-S. Chitosonic® acid as a novel cosmetic ingredient: Evaluation of its antimicrobial, antioxidant and hydration activities. Materials 2013, 6, 1391–1402. [Google Scholar]
- Chen, Y.C.; Li, Y.C.; You, B.J.; Chang, W.T.; Chao, L.K.; Lo, L.C.; Wang, S.Y.; Huang, G.J.; Kuo, Y.H. Diterpenoids with anti-inflammatory activity from the wood of Cunninghamia konishii. Molecules 2013, 18, 682–689. [Google Scholar]
- Freire, A.C.; de Assis, C.F.; Frick, A.O.; da Silva Melo, P.; Haun, M.; Aoyama, H.; Duran, N.; Sauer, M.M.; Kallas, E.G.; Ferreira, C.V. Influence of protein phosphatase inhibitors on HL60 cells death induction by dehydrocrotonin. Leuk. Res 2003, 27, 823–829. [Google Scholar]
- Huang, C.S.; Hu, H.H.; Tsai, Y.M.; Chang, W.T. In vitro effects of Monascus purpureus on antioxidation activity during fermentation of Kinmen sorghum liquor waste. J. Biosci. Bioeng 2013, 115, 418–423. [Google Scholar]
- Park, C.M.; Park, J.Y.; Noh, K.H.; Shin, J.H.; Song, Y.S. Taraxacum officinale Weber extracts inhibit LPS-induced oxidative stress and nitric oxide production via the NF-kappaB modulation in RAW 264.7 cells. J. Ethnopharmacol 2011, 133, 834–842. [Google Scholar]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem 1985, 150, 76–85. [Google Scholar]
- Lu, H.F.; Lai, T.Y.; Hsia, T.C.; Tang, Y.J.; Yang, J.S.; Chiang, J.H.; Lu, C.C.; Liu, C.M.; Wang, H.L.; Chung, J.G. Danthron induces DNA damage and inhibits DNA repair gene expressions in GBM 8401 human brain glioblastoma multiforms cells. Neurochem. Res 2010, 35, 1105–1110. [Google Scholar]
- Yin, H.; Miao, J.; Ma, C.; Sun, G.; Zhang, Y. Beta-Casomorphin-7 cause decreasing in oxidative stress and inhibiting NF-kappaB-iNOS-NO signal pathway in pancreas of diabetes rats. J. Food Sci 2012, 77, C278–C282. [Google Scholar]
- Yao, X.; Zhu, L.; Chen, Y.; Tian, J.; Wang, Y. In vivo and in vitro antioxidant activity and alpha-glucosidase, alpha-amylase inhibitory effects of flavonoids from Cichorium glandulosum seeds. Food Chem 2013, 139, 59–66. [Google Scholar]
- Afolayan, A.J.; Sunmonu, T.O. Artemisia afra Jacq. ameliorates oxidative stress in the pancreas of streptozotocin-induced diabetic Wistar rats. Biosci. Biotechnol. Biochem 2011, 75, 2083–2086. [Google Scholar]
- Lee, S.J.; Bai, S.K.; Lee, K.S.; Namkoong, S.; Na, H.J.; Ha, K.S.; Han, J.A.; Yim, S.V.; Chang, K.; Kwon, Y.G.; et al. Astaxanthin inhibits nitric oxide production and inflammatory gene expression by suppressing I(kappa)B kinase-dependent NF-kappaB activation. Mol. Cells 2003, 16, 97–105. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds | KI a | Peak area (%) |
|---|---|---|---|
| 01 | Camphene | 929 | 1.79 |
| 02 | alpha-Pinene | 939 | 1.18 |
| 03 | beta-Pinene | 943 | 2.38 |
| 04 | Yomogi alcohol | 1002 | 6.36 |
| 05 | o-Cymene | 1021 | 5.26 |
| 06 | D-Limonene | 1042 | 7.39 |
| 07 | 1,8-Cineole | 1059 | 10.15 |
| 08 | Artemisia ketone | 1065 | 14.92 |
| 09 | Artemisia alcohol | 1068 | 2.41 |
| 10 | Linalool | 1082 | 6.55 |
| 11 | Thujone | 1097 | 1.68 |
| 12 | Camphor | 1121 | 11.64 |
| 13 | Borneol | 1138 | 5.37 |
| 14 | Terpinenol-4-ol | 1139 | 3.69 |
| 15 | (Z)-Chrysanthenyl acetate | 1276 | 1.44 |
| 16 | Linalyl acetate | 1279 | 11.51 |
| 17 | Caryophyllene | 1494 | 1.83 |
| 18 | Germacrene D | 1515 | 2.76 |
| 19 | Viridiflorol | 1530 | 1.69 |
| Monoterpene hydrocarbons | 18.00 | ||
| Oxygenated monoterpenes | 75.72 | ||
| Sesquiterpene hydrocarbons | 4.59 | ||
| Oxygenated sesquiterpenes | 1.69 | ||
| Total identified | 100.00 |
| Target | Type | Sequences |
|---|---|---|
| β-actin | Sense | 5′-TGGAATCCTGTGGCATCCATGAAAC-3′ |
| Anti-sense | 5′-TAAAACGCAGCTCAGTAACAGTCCG-3′ | |
| iNOS | Sense | 5′-AGACTGGATTTGGCTGGTCCCTCC-3′ |
| Anti-sense | 5′-AGAACTGAGGGTACATGCTGGAGCC-3′ | |
| COX-2 | Sense | 5′-GGAGAGACTATCAAGATAGT-3′ |
| Anti-sense | 5′-ATGGTCAGTAGACTTTTACA-3′ | |
| TNF-α | Sense | 5′-GGCAGGTCTACTTTGGAGTCATTGC-3′ |
| Anti-sense | 5′-ACATTCGAGGCTCCAGTGAATTCGG-3′ | |
| IL-6 | Sense | 5′-GAGGATACCACTCCCAACAGA-3′ |
| Anti-sense | 5′-AAGTGCATCATCGTTGTTCATACA-3′ | |
| HO-1 | Sense | 5′-TGAAGGAGGCCACCAAGGAGG-3′ |
| Anti-sense | 5′-AGAGGTCACCCAGGTAGCGGG-3′ |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chou, S.-T.; Peng, H.-Y.; Hsu, J.-C.; Lin, C.-C.; Shih, Y. Achillea millefolium L. Essential Oil Inhibits LPS-Induced Oxidative Stress and Nitric Oxide Production in RAW 264.7 Macrophages. Int. J. Mol. Sci. 2013, 14, 12978-12993. https://doi.org/10.3390/ijms140712978
Chou S-T, Peng H-Y, Hsu J-C, Lin C-C, Shih Y. Achillea millefolium L. Essential Oil Inhibits LPS-Induced Oxidative Stress and Nitric Oxide Production in RAW 264.7 Macrophages. International Journal of Molecular Sciences. 2013; 14(7):12978-12993. https://doi.org/10.3390/ijms140712978
Chicago/Turabian StyleChou, Su-Tze, Hsin-Yi Peng, Jaw-Cherng Hsu, Chih-Chien Lin, and Ying Shih. 2013. "Achillea millefolium L. Essential Oil Inhibits LPS-Induced Oxidative Stress and Nitric Oxide Production in RAW 264.7 Macrophages" International Journal of Molecular Sciences 14, no. 7: 12978-12993. https://doi.org/10.3390/ijms140712978