Immunomodulatory Activity of Recombinant Ricin Toxin Binding Subunit B (RTB)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
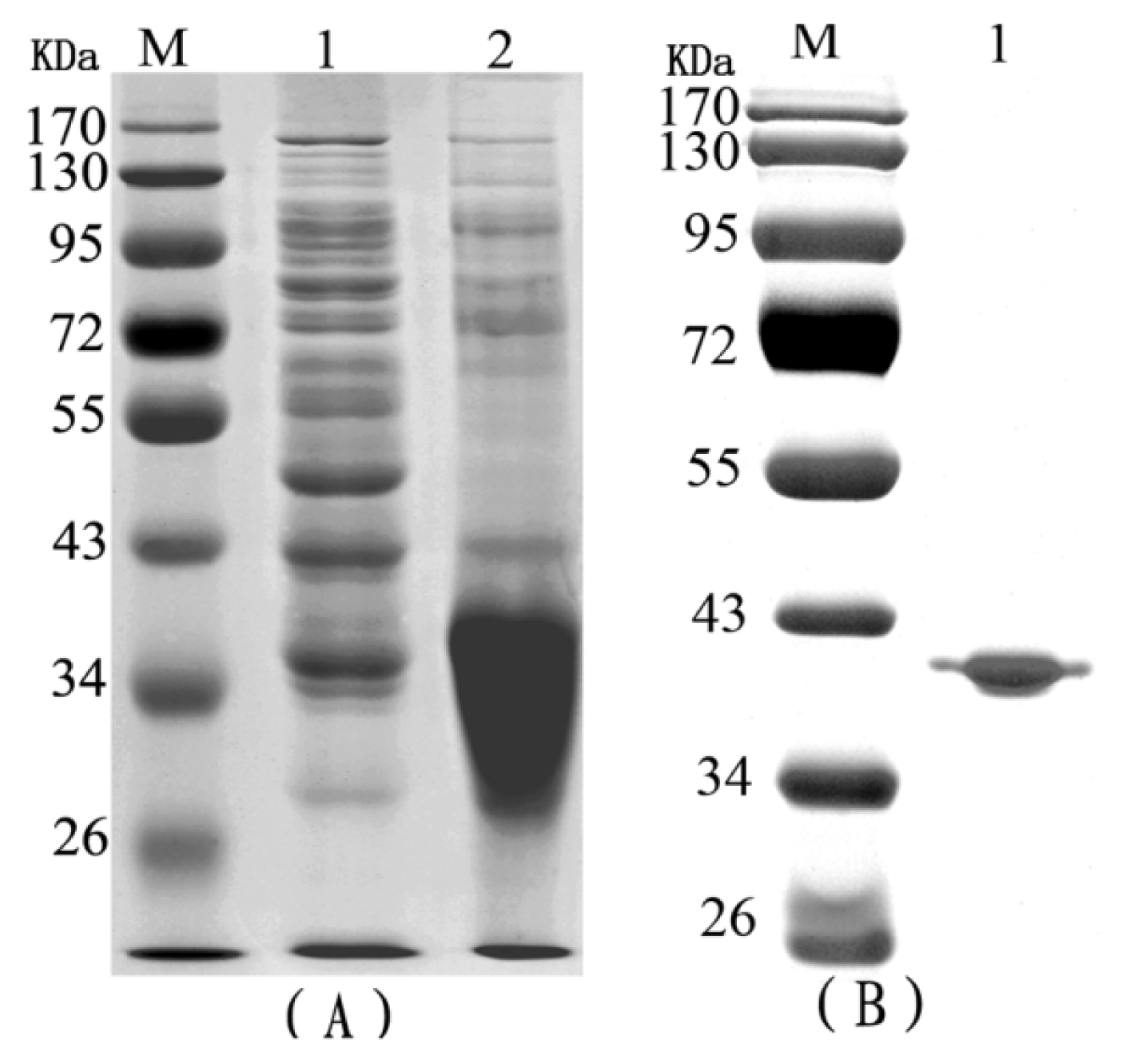
2.1. The Purification of Recombinant RTB
2.2. Cell Viability Assay
2.3. No Production
2.4. Cytokine Production of RAW264.7 Cells
2.5. The Proliferation of Mouse Splenocytes in Vitro
2.6. Spleen Cell Cytokine Production
3. Discussion
4. Experimental Section
4.1. Protein
4.2. Mice and Cell Line
4.3. Cell Viability Assay
4.4. NO Production Assay
4.5. Splenocyte Proliferation Assay
4.6. Cytokine Assays
4.7. Statistical Analysis
5. Conclusions
Conflict of Interest
References
- Stirpe, F.; Barbieri, L. Ribosome-inactivating proteins up to date. FEBS. Lett 1986, 195, 1–8. [Google Scholar]
- Zentz, C.; Frenoy, J.P.; Bourrillon, R. Bourrillon, binding of galactose and lactose to ricin. Equilibrium studies. Biochim. Biophys. Acta 1978, 536, 18–26. [Google Scholar]
- Rutenber, E.; Ready, M.; Robertus, J.D. Structure and evolution of ricin B chain. Nature 1987, 326, 624–626. [Google Scholar]
- Lord, M.J.; Jolliffe, N.A.; Marsden, C.J.; Pateman, C.S.; Smith, D.C.; Spooner, R.A.; Watson, P.D.; Roberts, L.M. Ricin: Mechanisms of cytotoxicity. Toxicol. Rev 2003, 22, 53–64. [Google Scholar]
- Medina-Bolivar, F.; Wright, R.; Funk, V.; Sentz, D.; Barroso, L.; Wilkins, T.D.; Petri, W., Jr; Cramer, C.L.A. Non-toxic lectin for antigen delivery of plant-based mucosal vaccines. Vaccine 2003, 21, 997–1005. [Google Scholar]
- Reed, D.G.; Nopo-Olazabal, L.H.; Funk, V.; Woffenden, B.J.; Reidy, M.J.; Dolan, M.C.; Cramer, C.L.; Medina-Bolivar, F. Expression of functional hexahistidine-tagged ricin B in tobacco. Plant Cell Rep 2005, 24, 15–24. [Google Scholar]
- Choi, N.W.; Estes, M.K.; Langridge, W.H. Mucosal immunization with a ricin toxin B subunit-rotavirus NSP4 fusion protein stimulates a Th1. J. Biotechnol 2006, 121, 272–283. [Google Scholar]
- Choi, N.W.; Estes, M.K.; Langridge, W.H. Synthesis of a ricin toxin B subunit-rotavirus VP7 fusion protein in potato. Mol. Biotechnol 2006, 32, 117–128. [Google Scholar]
- Choi, N.W.; Estes, M.K.; Langridge, W.H. Ricin toxin B subunit enhancement of rotavirus NSP4 immunogenicity in mice. Viral. Immunol 2006, 19, 54–63. [Google Scholar]
- Klimp, A.H.; de Vries, E.G.; Scherphof, G.L.; Daemen, T. A potential role of macrophage activation in the treatment of cancer. Crit. Rev. Oncol. Hematol 2002, 44, 143–161. [Google Scholar]
- Vijayan, M.; Chandra, N. Lectins. Curr. Opin 1999, 9, 707–714. [Google Scholar]
- Olsnes, S. The history of ricin, abrin and related toxins. Toxicon 2004, 44, 361–370. [Google Scholar]
- Sandvig, K.; Torgersen, M.L.; Engedal, N.; Skotland, T.; Iversen, T.G. Protein toxins from plants and bacteria: Probes for intracellular transport and tools in medicine. FEBS. Lett 2010, 584, 2626–2634. [Google Scholar]
- Yermakova, A.; Mantis, N.J. Protective immunity to ricin toxin conferred by antibodies against the toxin’s binding subunit (RTB). Vaccine 2011, 29, 7925–7935. [Google Scholar]
- Donayre-Torres, A.J.; Esquivel-Soto, E.; Gutiérrez-Xicoténcatl, M.L.; Esquivel-Guadarrama, F.R.; Gómez-Lim, M.A. Production and purification of immunologically active core protein P24 from HIV-1 fused to ricin toxin B subunit in E. coli. Virol. J 2007, 6, 17, :1–17:13.. [Google Scholar]
- Wang, Y.; Cui, X.; Tai, G.; Ge, J.; Li, N.; Chen, F.; Yu, F.; Liu, Z.A. Critical role of activin A in maturation of mouse peritoneal macrophages in vitro and in vivo. Cell Mol. Immunol 2009, 6, 387–392. [Google Scholar]
- Zoccal, K.F.; Bitencourt, C.S.; Secatto, A.; Sorgi, C.A.; Bordon, K.C.; Sampaio, S.V.; Arantes, E.C.; Faccioli, L.H. Tityus serrulatus venom and toxins Ts1, Ts2 and Ts6 induce macrophage activation and production of immune mediators. Toxicon 2011, 57, 1101–1108. [Google Scholar]
- Baugh, J.A.; Bucala, R. Mechanisms for modulating TNFa in immune and inflammatory disease. Curr. Opin. Drug. Discov. Devel 2001, 4, 635–650. [Google Scholar]
- Ghezzi, P.; Cerami, A. Tumor necrosis factor as a pharmacological target. Mol. Biotechnol 2005, 31, 239–244. [Google Scholar]
- Mosmann, T.R.; Cherwinski, H.; Bond, M.W.; Giedlin, M.A.; Coffman, R.L. Two types of murine helper T-cell clones. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol 1986, 136, 2348–2357. [Google Scholar]
- Mosmann, T.R.; Coffman, R.L. Heterogeneity of cytokine secretion patterns and functions of helper T cells. Adv. Immunol 1989, 46, 111–147. [Google Scholar]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev 2003, 8, 223–246. [Google Scholar]
- Krek, C.E.; Ladino, C.A.; Goldmacher, V.S.; Blättler, W.A.; Guild, B.C. Expression and secretion of a recombinant ricin immunotoxin from murine myeloma cells. Protein Eng 1995, 8, 481–489. [Google Scholar]
- Morgan, E.; Varro, R.; Sepulveda, H.; Ember, J.A.; Apgar, J.; Wilson, J.; Lowe, L.; Chen, R.; Shivraj, L.; Agadir, A.; et al. Cytometric bead array: A multiplexed assay platform with applications in various areas of biology. Clin. Immunol 2004, 110, 252–266. [Google Scholar]


© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, W.; Xu, N.; Yuan, H.; Li, S.; Liu, L.; Pu, Z.; Wan, J.; Wang, H.; Chang, Y.; Li, R. Immunomodulatory Activity of Recombinant Ricin Toxin Binding Subunit B (RTB). Int. J. Mol. Sci. 2013, 14, 12401-12410. https://doi.org/10.3390/ijms140612401
Liu W, Xu N, Yuan H, Li S, Liu L, Pu Z, Wan J, Wang H, Chang Y, Li R. Immunomodulatory Activity of Recombinant Ricin Toxin Binding Subunit B (RTB). International Journal of Molecular Sciences. 2013; 14(6):12401-12410. https://doi.org/10.3390/ijms140612401
Chicago/Turabian StyleLiu, Wensen, Na Xu, Hongyan Yuan, Songyan Li, Linna Liu, Zhaoyang Pu, Jiayu Wan, Huiwen Wang, Yaping Chang, and Ruisheng Li. 2013. "Immunomodulatory Activity of Recombinant Ricin Toxin Binding Subunit B (RTB)" International Journal of Molecular Sciences 14, no. 6: 12401-12410. https://doi.org/10.3390/ijms140612401