Photochemical Production and Behavior of Hydroperoxyacids in Heterotrophic Bacteria Attached to Senescent Phytoplanktonic Cells

Abstract
:1. Introduction
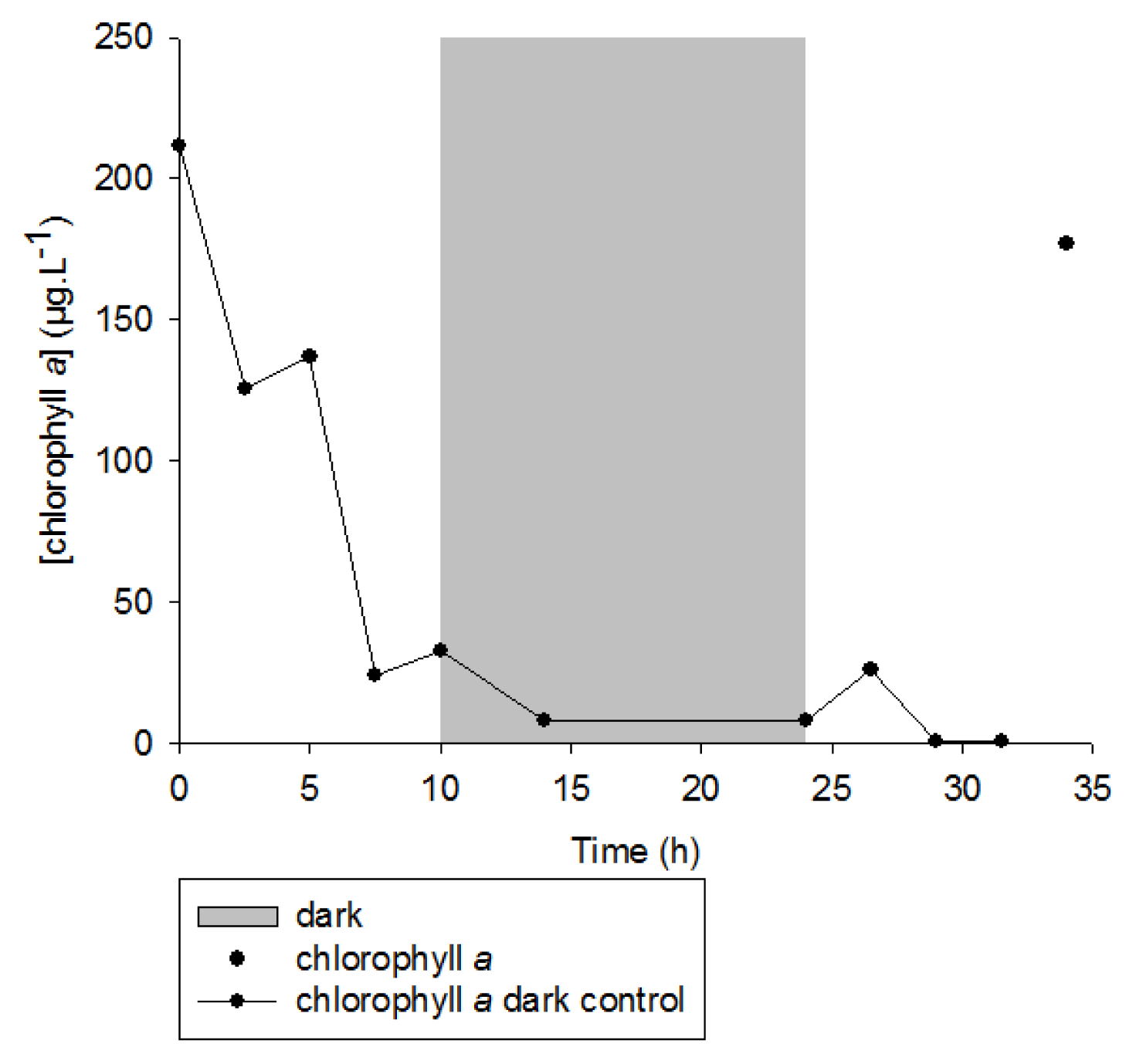
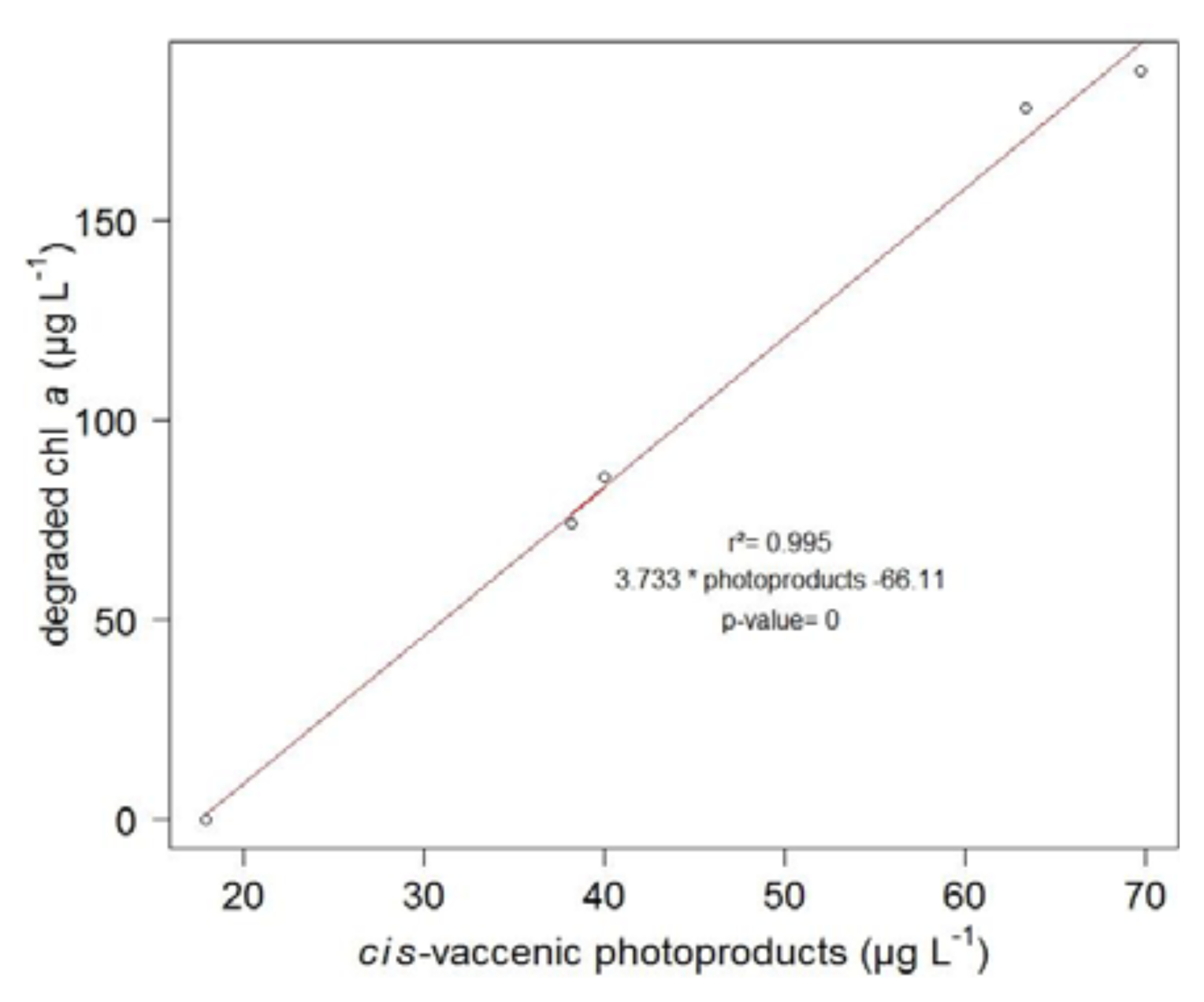
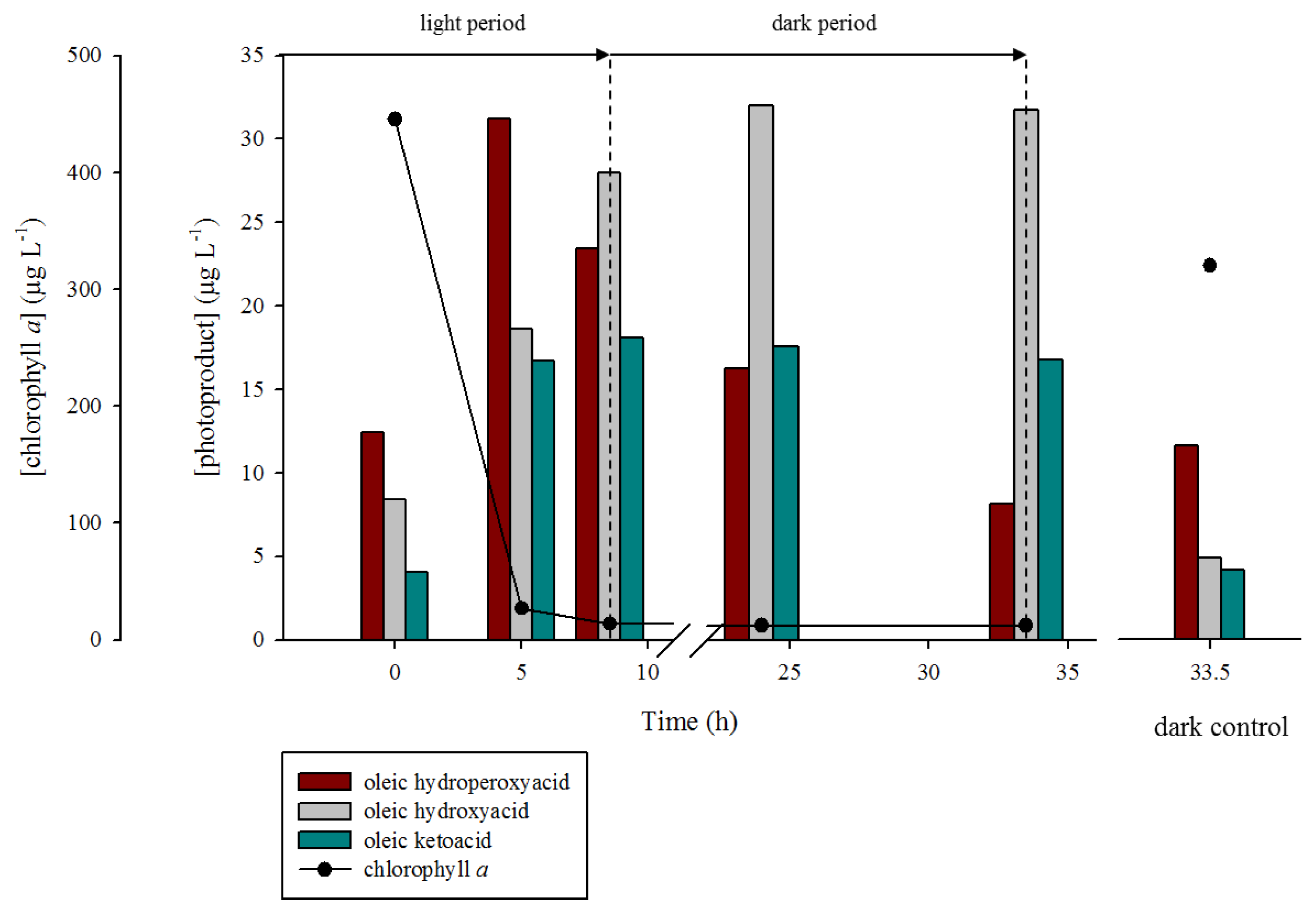
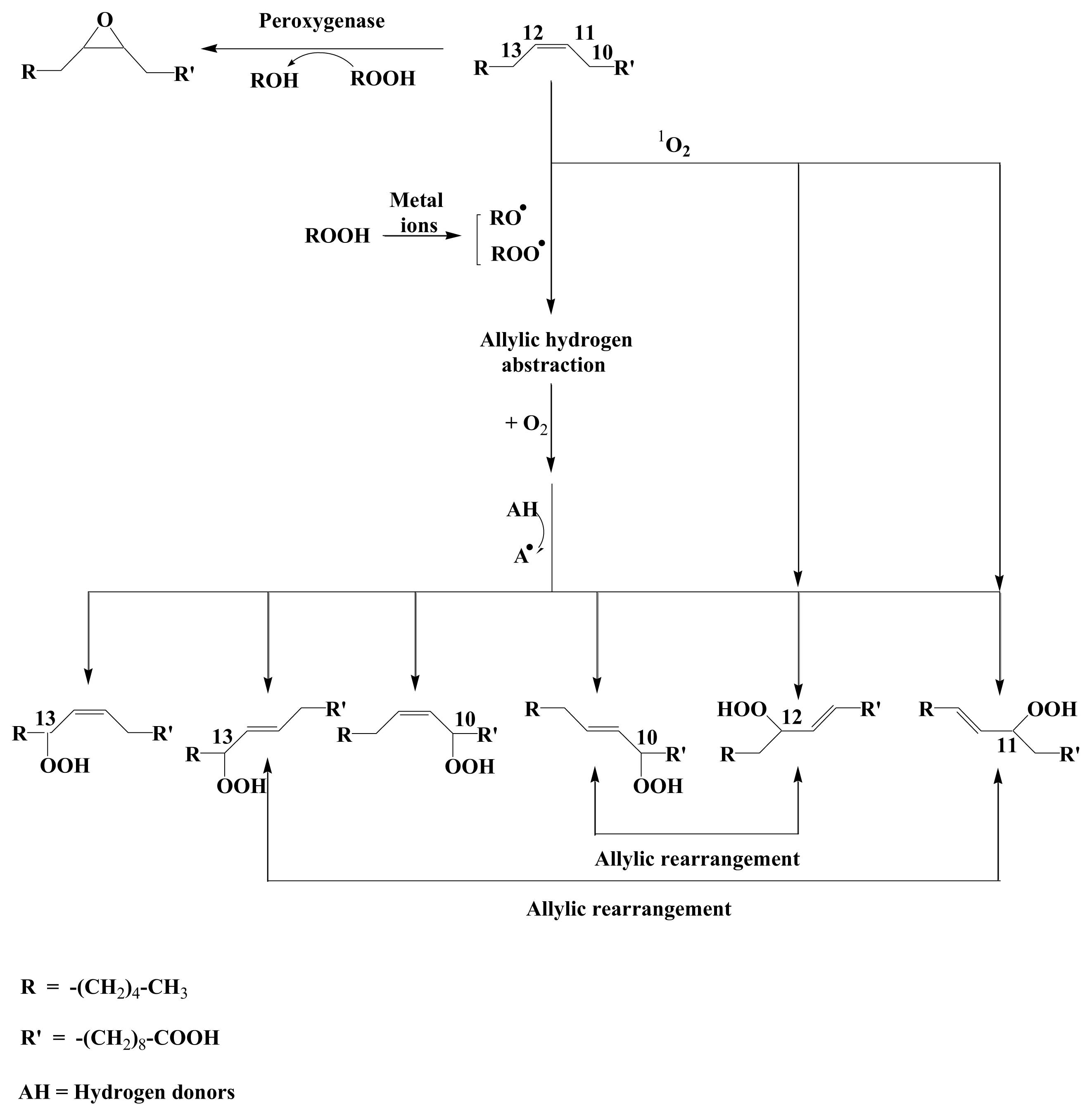
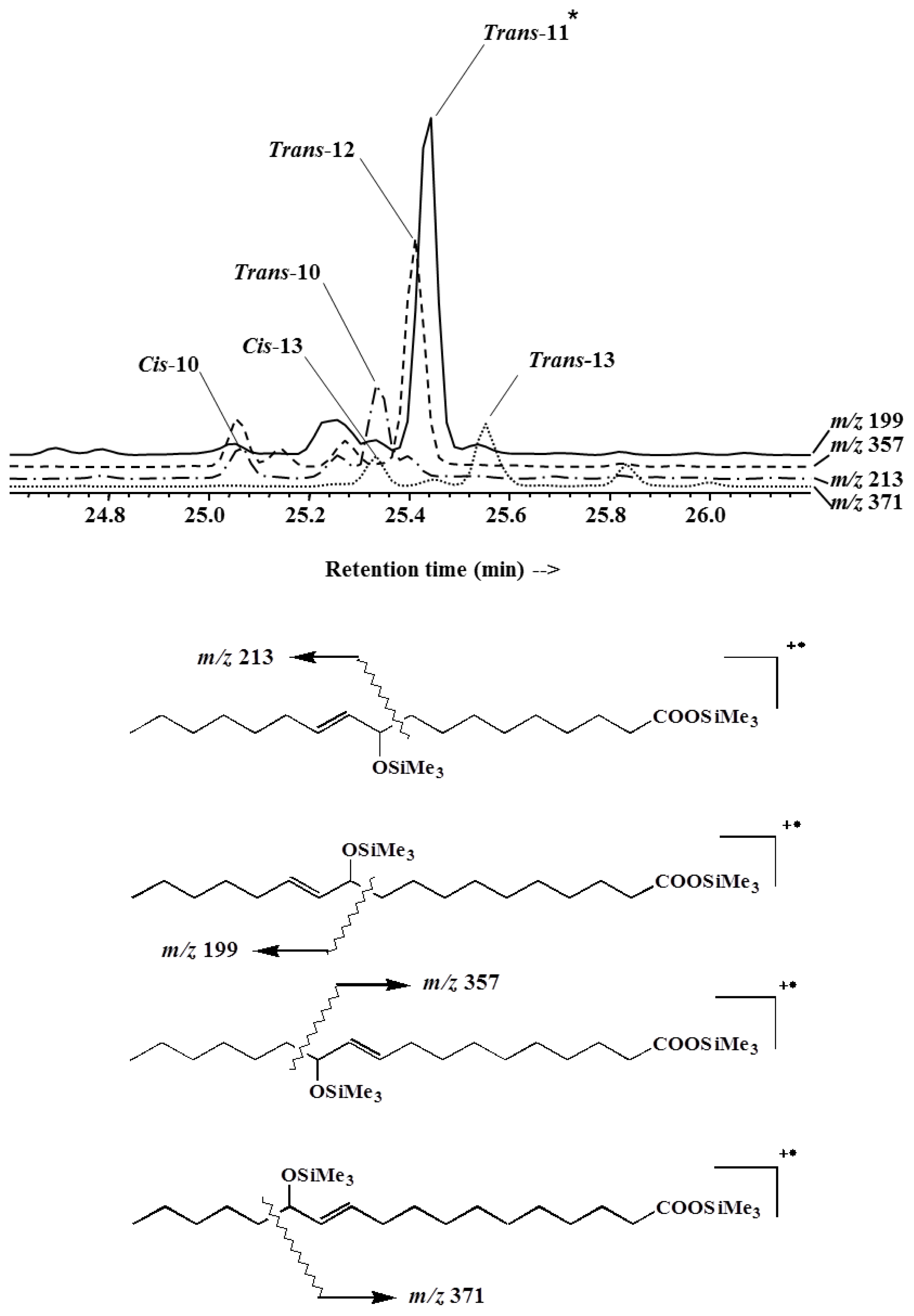
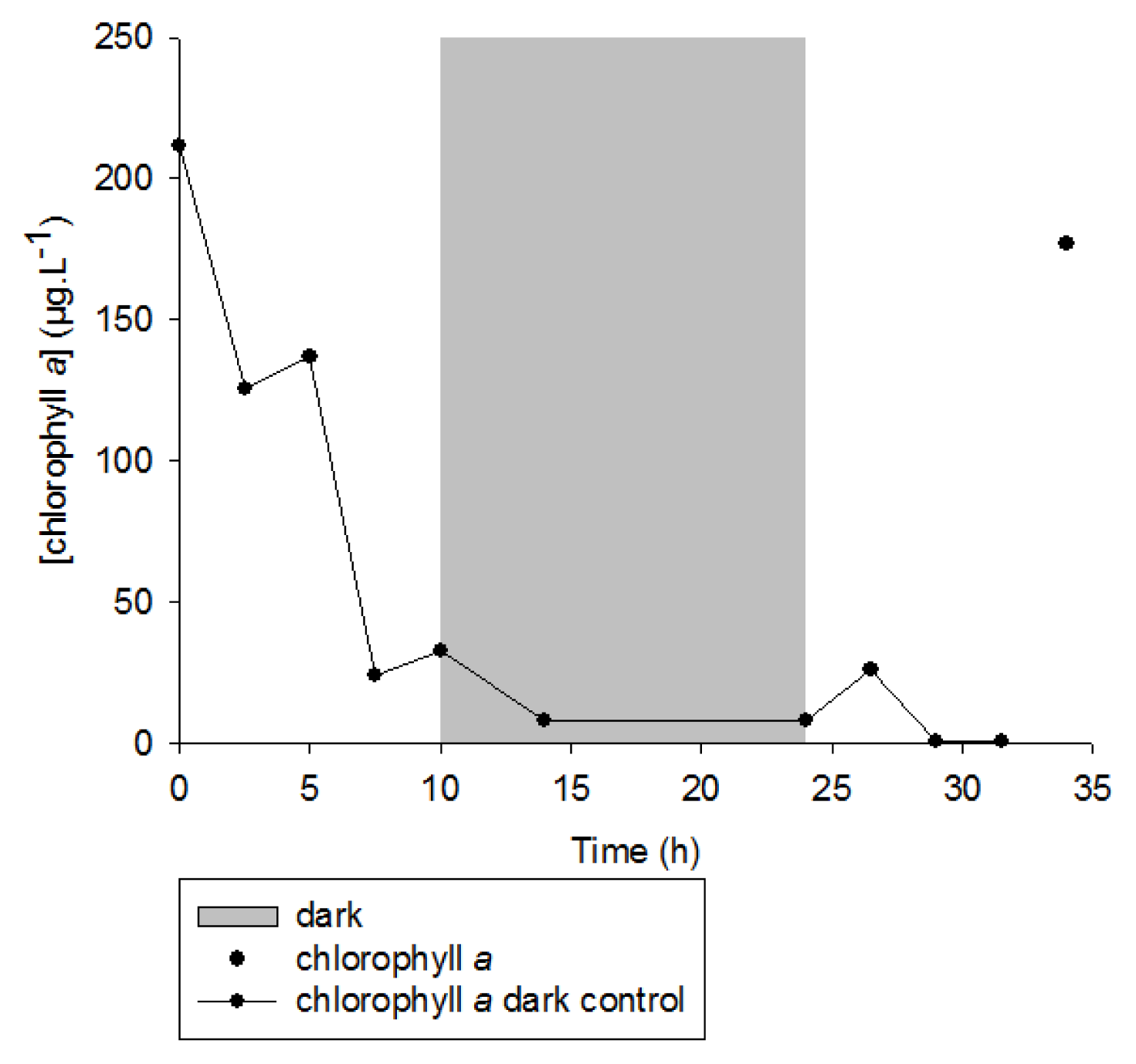
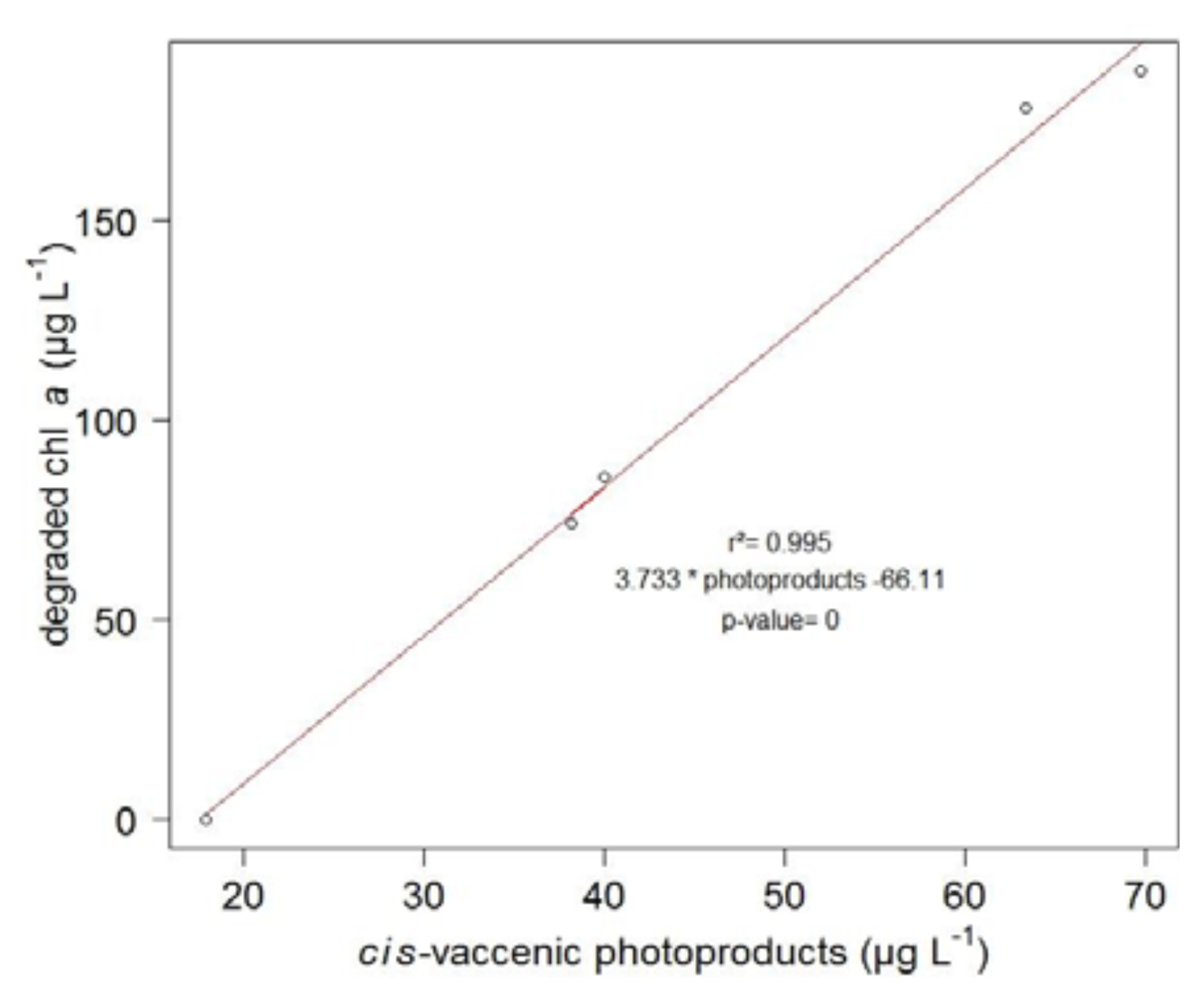
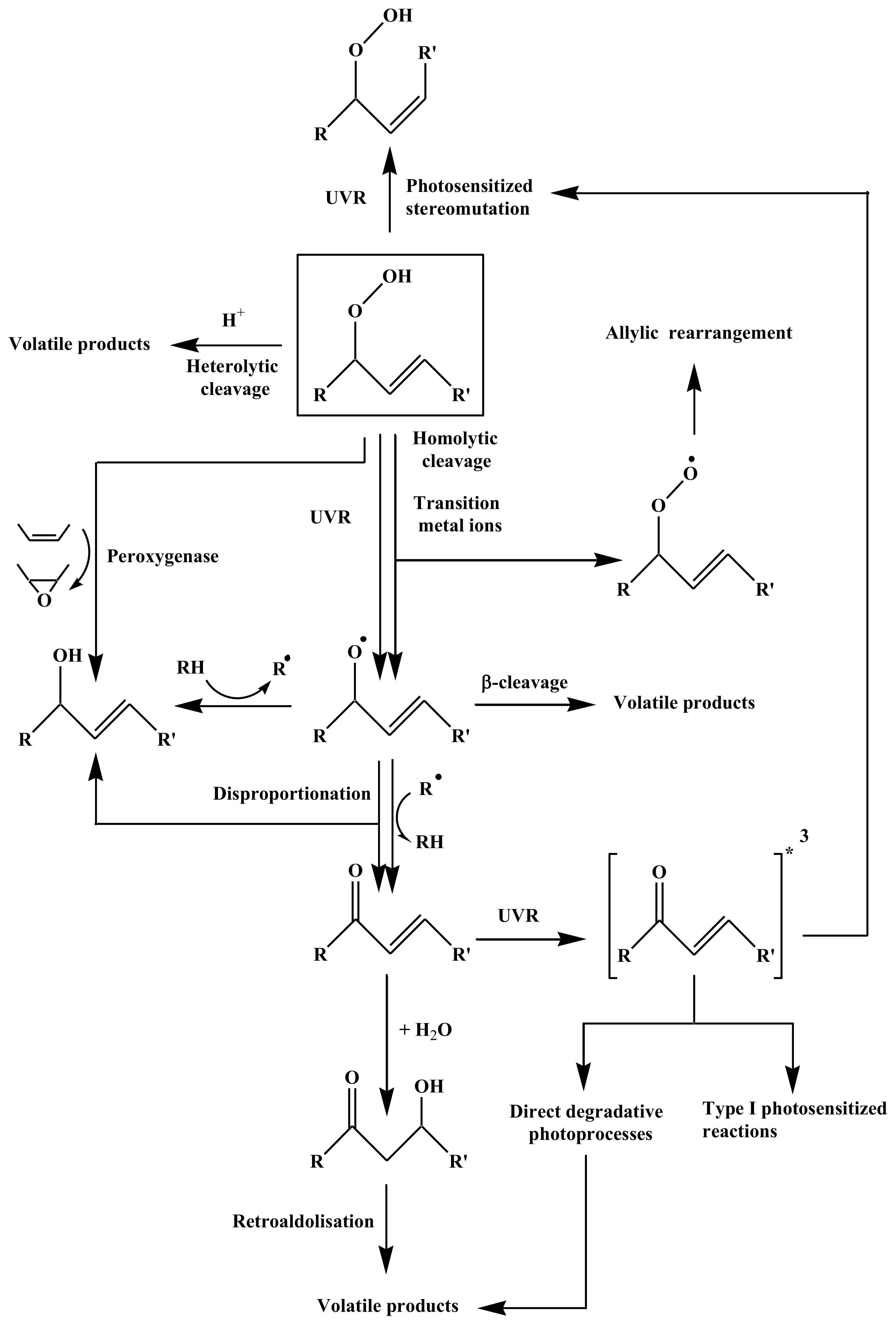
2. Results and Discussion
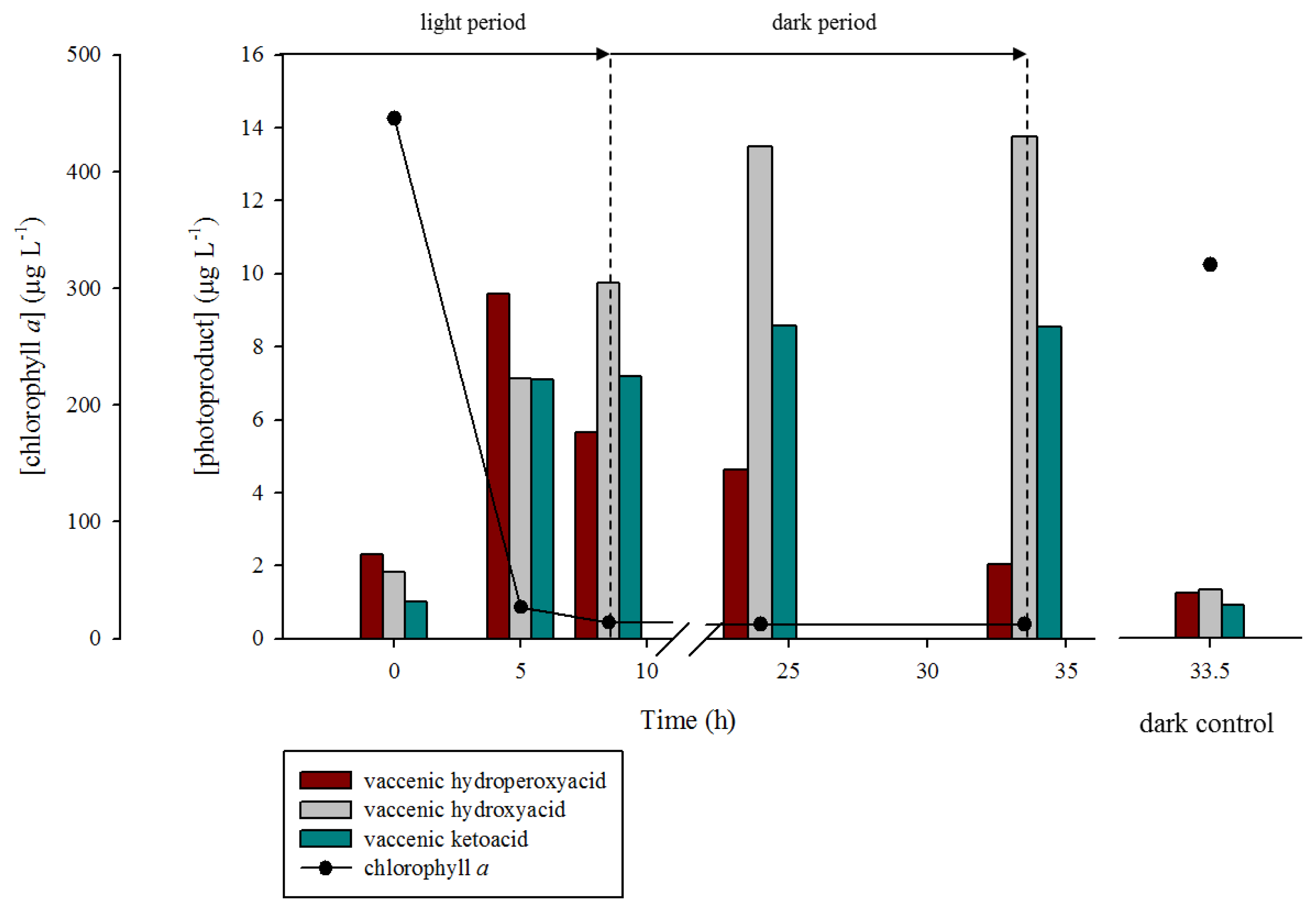
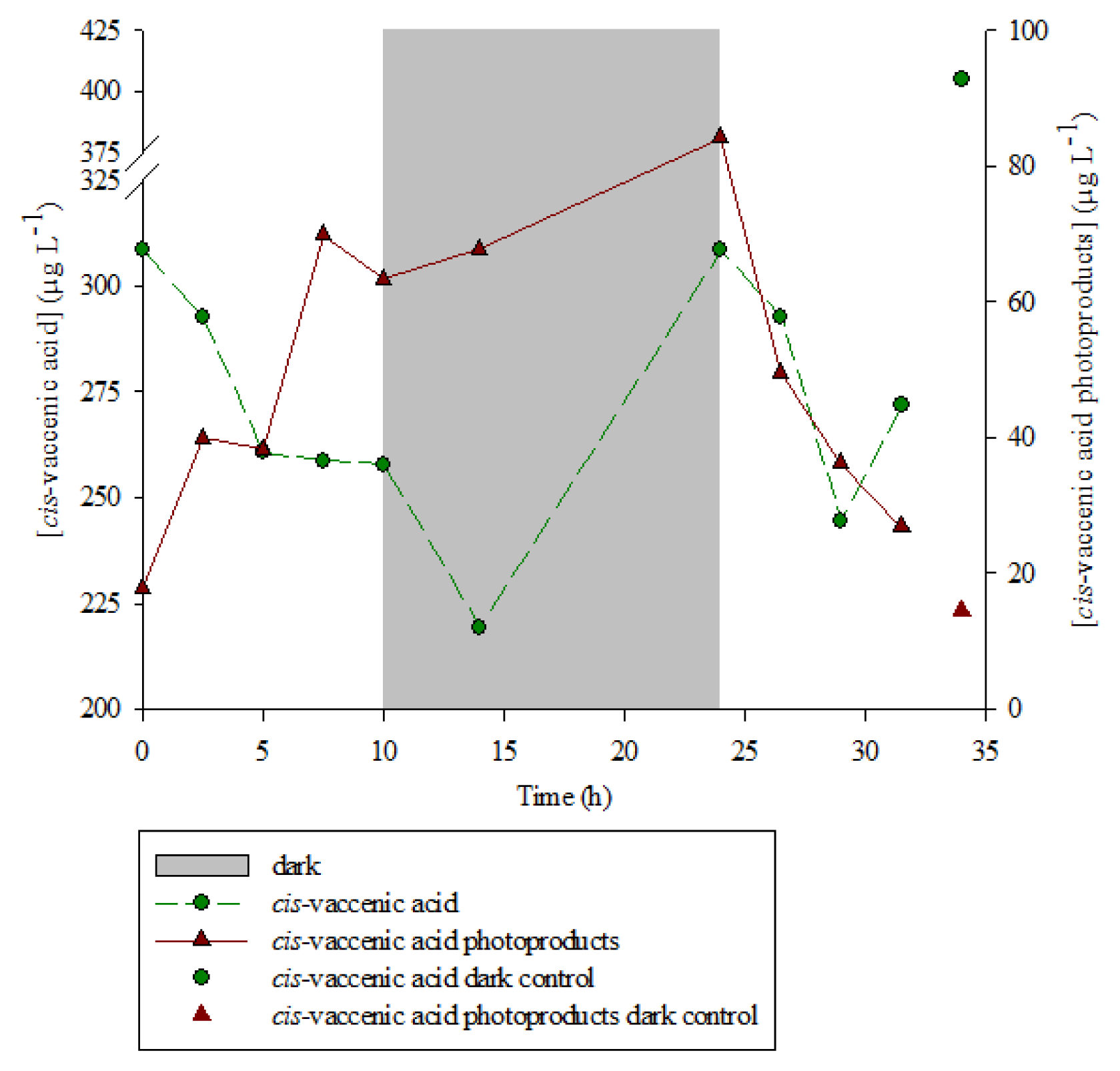
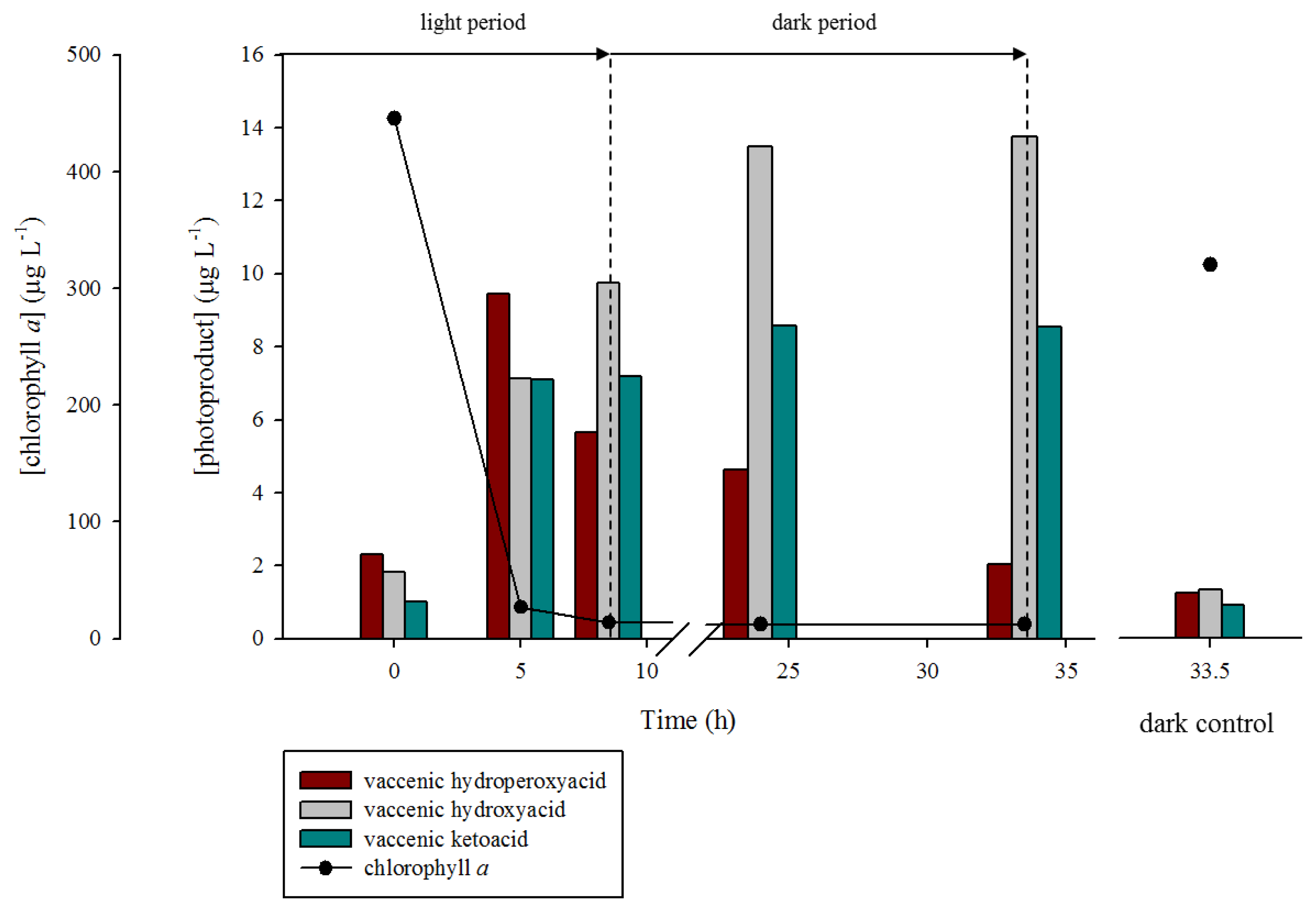
2.1. 1O2 Transfer
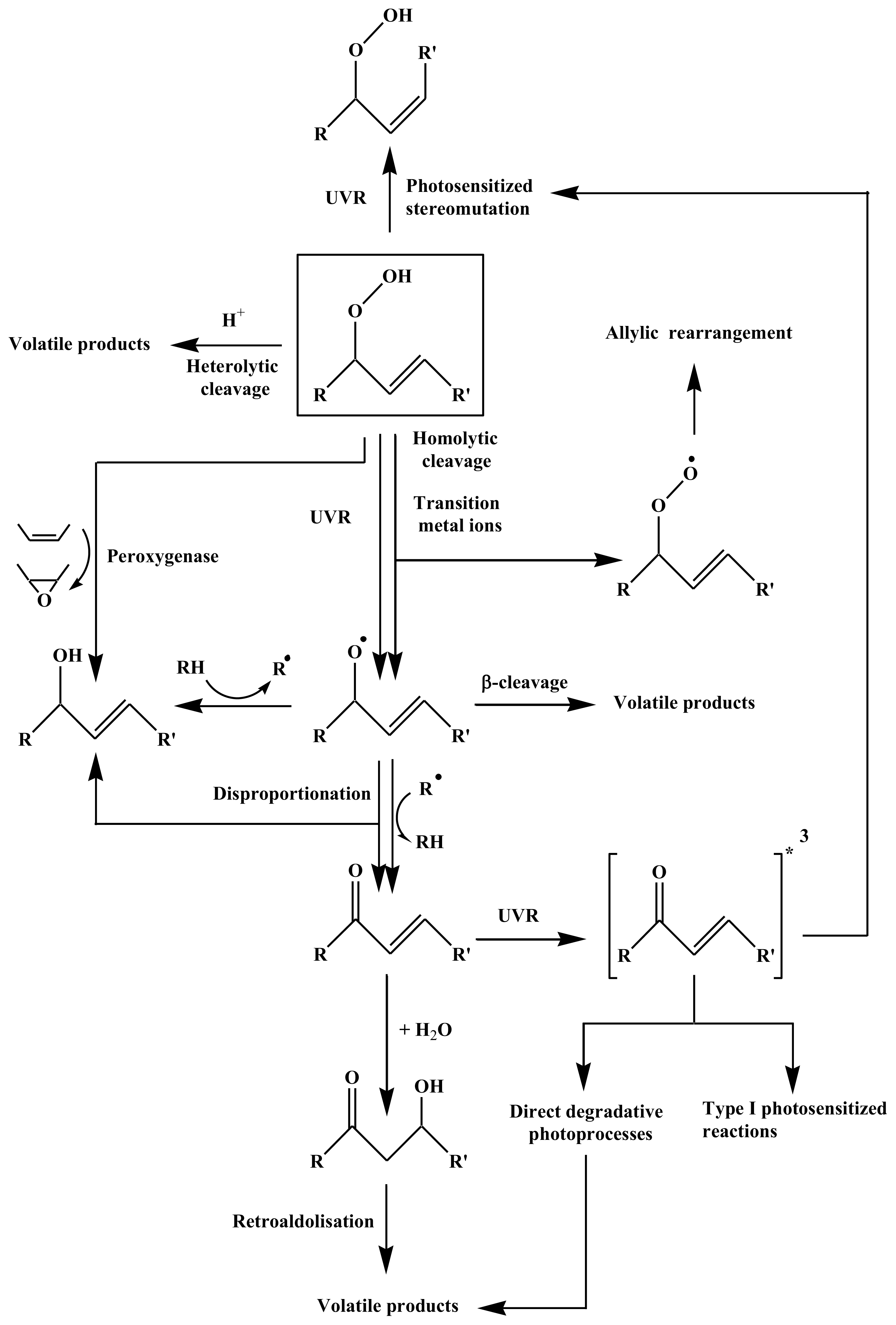
2.2. Degradation of Hydroperoxides
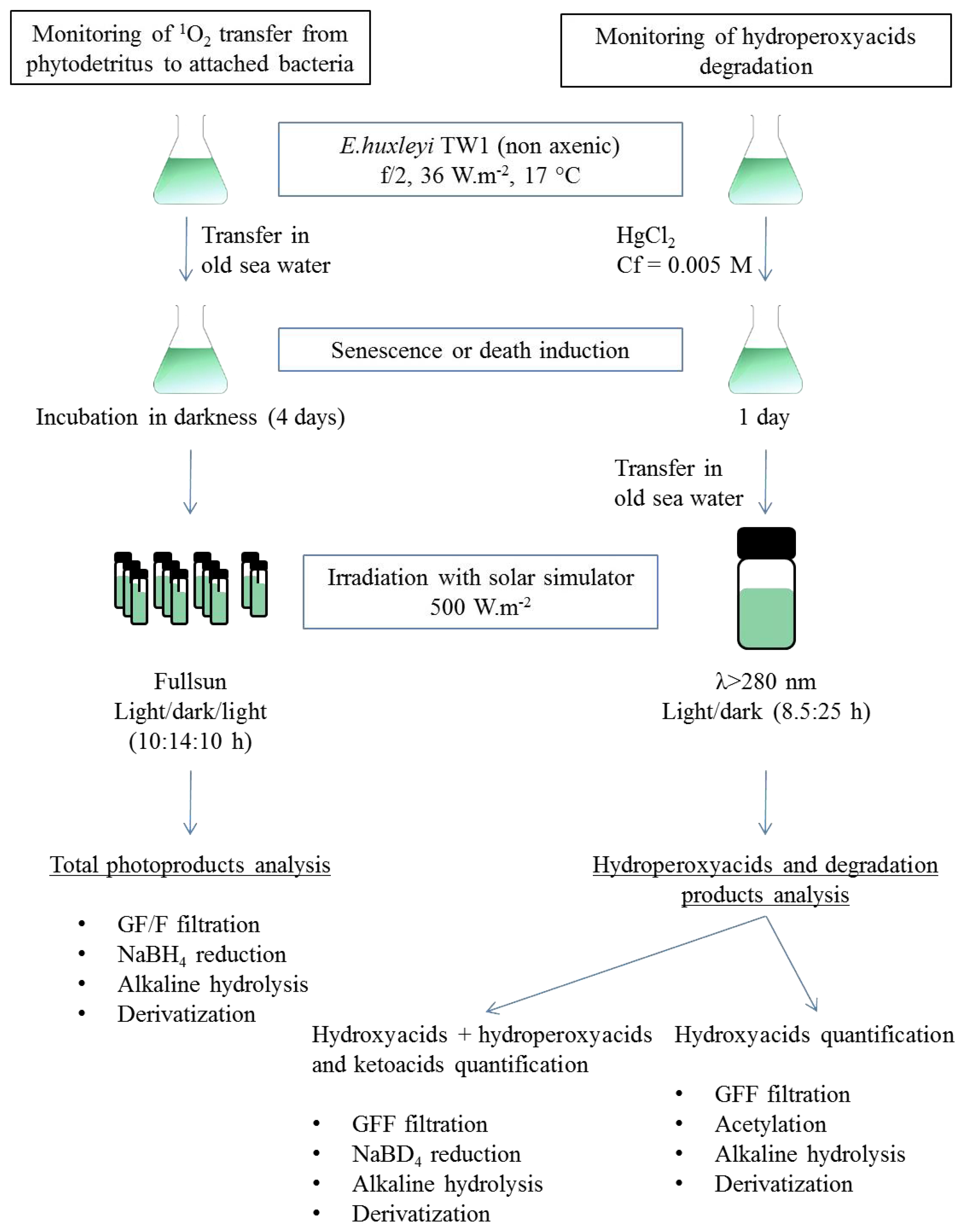
3. Experimental Section
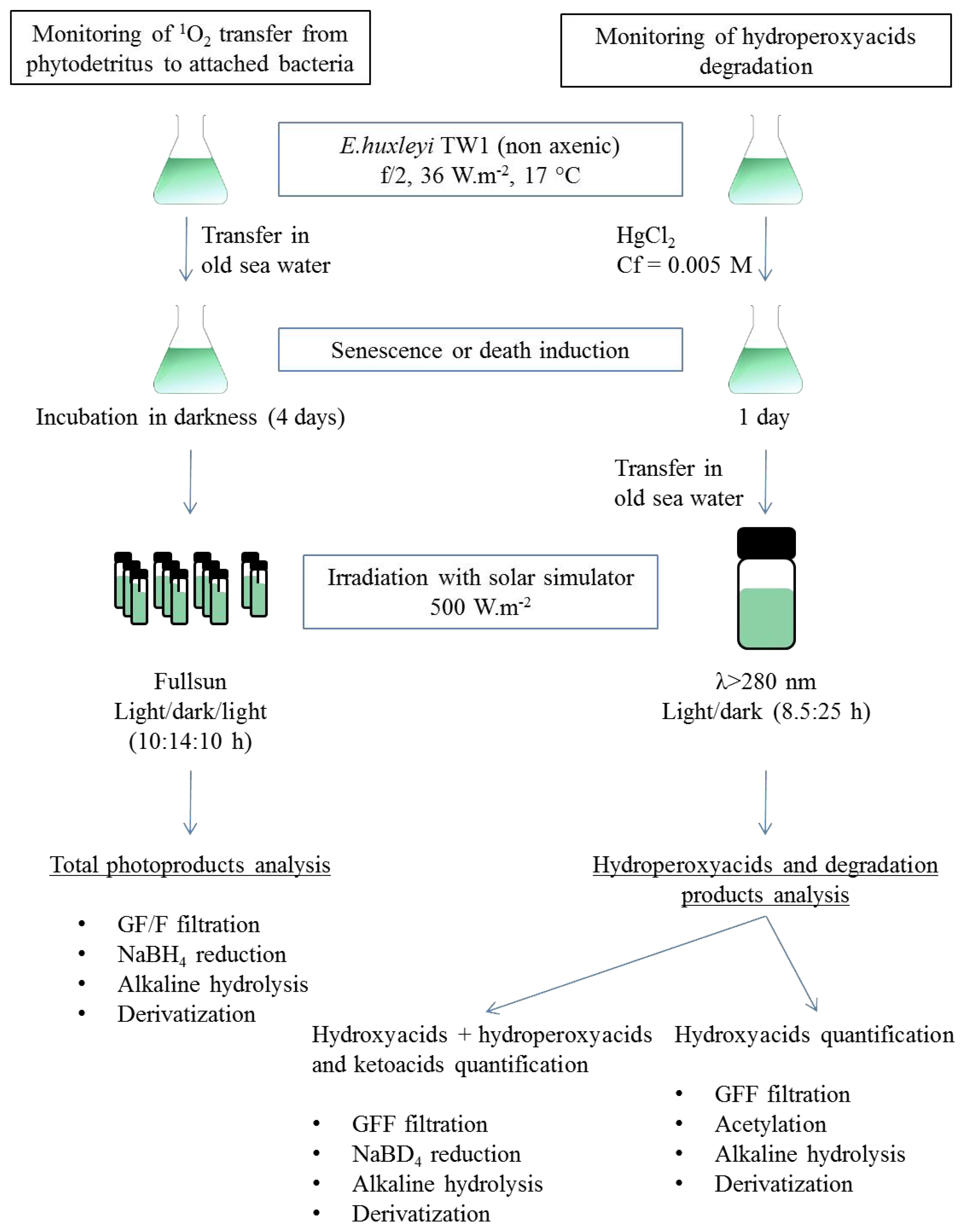
3.1. Algal and Bacterial Material Production
3.2. Photodegradation Experiments
3.3. Lipid Analyses
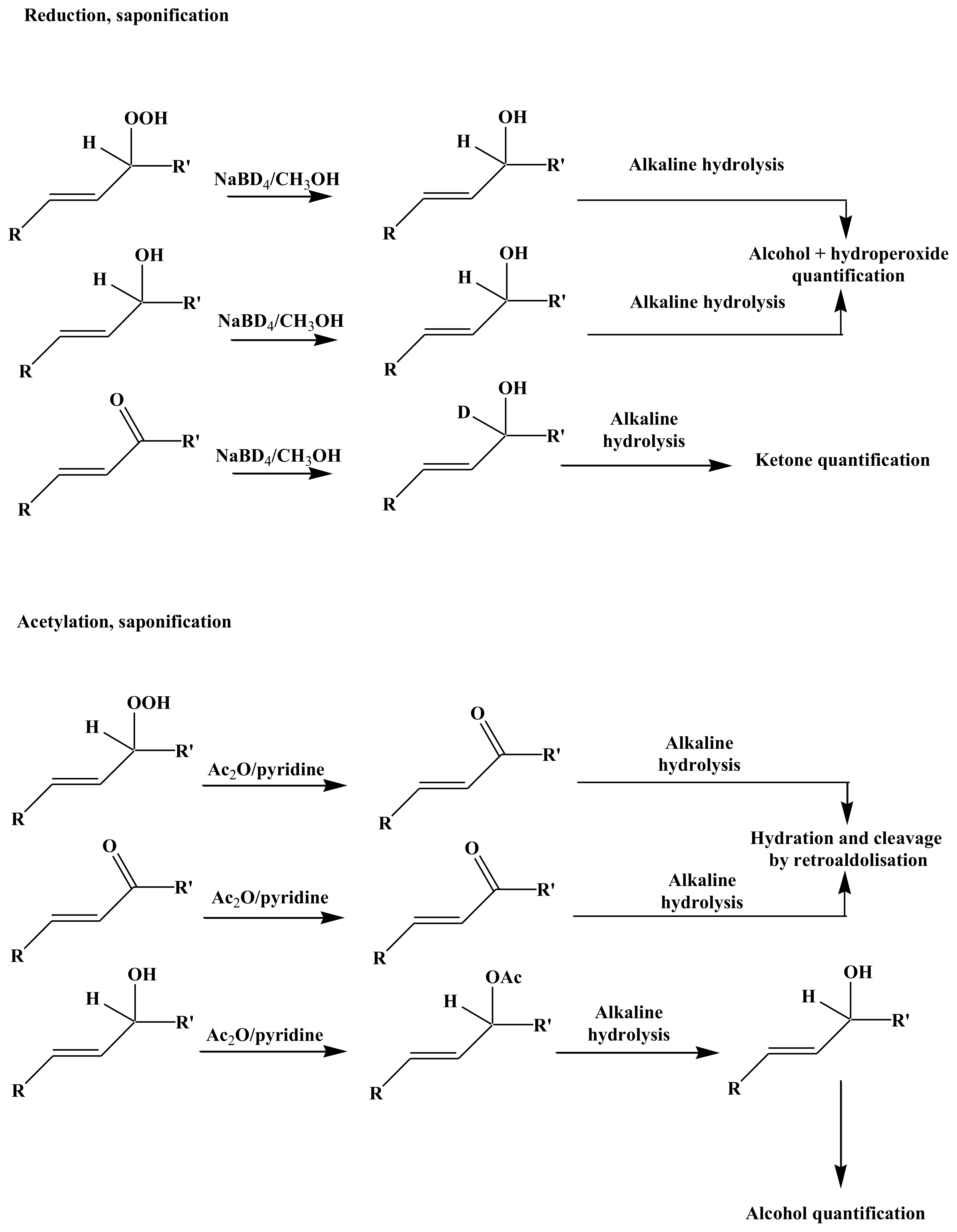
3.3.1. Reduction
3.3.2. Acetylation
3.3.3. Alkaline Hydrolysis
3.3.4. Derivatization
3.4. Identification and Quantification of Lipid Biomarkers and Their Degradation Products by Gas Chromatography—Electron Impact Mass Spectrometry
3.5. Chlorophyll a Analyses
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Cho, B.C.; Azam, F. Major role of bacteria in biogeochemical fluxes in the ocean’s interior. Nature 1988, 332, 441–443. [Google Scholar]
- Eppley, R.; Peterson, B.J. Particulate organic matter flux and planktonic new production in the deep ocean. Nature 1979, 282, 677–680. [Google Scholar]
- Boyd, P.W.; Trull, T.W. Understanding the export of biogenic particles in oceanic waters: Is there consensus? Progress Oceanogr 2007, 72, 276–312. [Google Scholar]
- Thunell, R.; Benitez-Nelson, C.; Varela, R.; Astor, Y.; Muller-Karger, F. Particulate organic carbon fluxes along upwelling-dominated continental margins: Rates and mechanisms. Glob. Biogeochem. Cycles 2007, 21, 1–12. [Google Scholar]
- Turley, C.; Mackie, P. Biogeochemical significance of attached and free-living bacteria and the flux of particles in the NE Atlantic Ocean. Mar. Ecol. Progress Ser 1994, 115, 191–203. [Google Scholar]
- Sempéré, R.; Yoro, S.C.; van Wambeke, F.; Charrière, B. Microbial decomposition of large organic particles in the northwestern Mediterranean Sea: An experimental approach. Mar. Ecol. Progress Ser 2000, 198, 61–72. [Google Scholar]
- Goutx, M.; Wakeham, S.G.; Lee, C.; Duflos, M. Composition and degradation of marine particles with different settling velocities in the northwestern Mediterranean Sea. Limnol. Oceanogr 2007, 52, 1645–1664. [Google Scholar]
- Tamburini, C.; Goutx, M.; Guigue, C.; Garel, M.; Lefèvre, D.; Charrière, B.; Sempéré, R.; Pepa, S.; Peterson, M.L.; Wakeham, S.G.; Lee, C. Effects of hydrostatic pressure on microbial alteration of sinking fecal pellets. Deep Sea Res. Part II 2009, 56, 1533–1546. [Google Scholar]
- Rontani, J.-F.; Zabeti, N.; Wakeham, S.G. Degradation of particulate organic matter in the equatorial Pacific Ocean: Biotic or abiotic? Limnol. Oceanogr 2011, 56, 333–349. [Google Scholar]
- Mayer, L.M.; Schick, L.L.; Bianchi, T.S.; Wysocki, L.A. Photochemical changes in chemical markers of sedimentary organic matter source and age. Mar. Chem 2009, 113, 123–128. [Google Scholar]
- Rontani, J.-F. Visible light-dependent degradation of lipidic phytoplanktonic components during senescence: A review. Phytochemistry 2001, 58, 187–202. [Google Scholar]
- Christodoulou, S.; Joux, F.; Marty, J.-C.; Sempéré, R.; Rontani, J.-F. Comparative study of UV and visible light induced degradation of lipids in non-axenic senescent cells of Emiliania huxleyi. Mar. Chem 2010, 119, 139–152. [Google Scholar]
- Foote, C. Photosensitized Oxidation and Singlet Oxygen: Consequences in Biological Systems; Pryor, W.A., Ed.; Academic Press: New York, NY, USA, 1976; pp. 85–131. [Google Scholar]
- Knox, J.P.; Dodge, A.D. Singlet oxygen and plants. Phytochemistry 1985, 24, 889–896. [Google Scholar]
- Halliwell, B. Oxidative damage, lipid peroxidation and antioxidant protection in chloroplasts. Chem. Phys. Lipids 1987, 44, 327–340. [Google Scholar]
- Nelson, J.R. Rates and possible mechanism of light-dependent degradation of pigments in detritus derived from phytoplankton. J. Mar. Res 1993, 51, 155–179. [Google Scholar]
- Merzlyak, M.N.; Hendry, G.A.F. Free radical metabolism, pigment degradation and lipid peroxidation in leaves during senescence. Proc. R. Soc. Edinb 1994, 102B, 459–471. [Google Scholar]
- Suwa, K.; Kimura, T.; Schaap, A.P. Reactivity of singlet molecular oxygen with cholesterol in a phospholipid membrane matrix. A model for oxidative damage of membranes. Biochem. Biophys. Res. Commun 1977, 75, 785–792. [Google Scholar]
- Rontani, J.-F. Photooxidative and Autoxidative Degradation of Lipid Components during the Senescence of Phototrophic Organisms; Matsumoto, T., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2008; pp. 115–144. [Google Scholar]
- Rontani, J.-F.; Rabourdin, A.; Pinot, F.; Kandel, S.; Aubert, C. Visible light-induced oxidation of unsaturated components of cutins: A significant process during the senescence of higher plants. Phytochemistry 2005, 66, 313–321. [Google Scholar]
- Rontani, J.-F.; Koblízek, M.; Beker, B.; Bonin, P.; Kolber, Z.S. On the origin of cis-vaccenic acid photodegradation products in the marine environment. Lipids 2003, 38, 8. [Google Scholar]
- Sicre, M.; Paillasseur, J.; Marty, J.-C.; Saliot, A. Characterization of seawater samples using chemometric methods applied to biomarker fatty acids. Org. Geochem 1988, 12, 281–288. [Google Scholar]
- Keweloh, H.; Heipieper, H.J. Trans unsaturated fatty acids in bacteria. Lipids 1996, 31, 129–37. [Google Scholar]
- Abboudi, M.; Surget, S.M.; Rontani, J.-F.; Sempéré, R.; Joux, F. Physiological alteration of the marine bacterium Vibrio angustum S14 exposed to simulated sunlight during growth. Curr. Microbiol 2008, 57, 412–417. [Google Scholar]
- Garcia-Pichel, F. A model for internal self-shading in planktonic organisms and its implications for the usefulness of ultraviolet sunscreens. Limnol. Oceanogr 1994, 39, 1704–1717. [Google Scholar]
- Marchand, D.; Rontani, J.-F. Characterisation of photo-oxidation and autoxidation products of phytoplanktonic monounsaturated fatty acids in marine particulate matter and recent sediments. Org. Geochem 2001, 32, 287–304. [Google Scholar]
- Marchand, D.; Marty, J.-C.; Miquel, J.-C.; Rontani, J.-F. Lipids and their oxidation products as biomarkers for carbon cycling in the northwestern Mediterranean Sea: Results from a sediment trap study. Mar. Chem 2005, 95, 129–147. [Google Scholar]
- Christodoulou, S.; Marty, J.-C.; Miquel, J.-C.; Volkman, J.K.; Rontani, J.-F. Use of lipids and their degradation products as biomarkers for carbon cycling in the northwestern Mediterranean Sea. Mar. Chem 2009, 113, 25–40. [Google Scholar]
- Minnock, A.; Vernon, D.; Schofield, J. Photoinactivation of bacteria. Use of a cationic water-soluble zinc phthalocyanine to photoinactivate both gram-negative and gram-positive bacteria. J. Photochem. Photobiol. B 1996, 32, 159–164. [Google Scholar]
- Merchat, M.; Bertolini, G.; Giacomini, P.; Villanueva, A; Jori, G. Meso-substituted cationic porphyrins as efficient photosensitizers of gram-positive and gram-negative bacteria. J. Photochem. Photobiol. B 1996, 32, 153–7. [Google Scholar]
- Merchat, M.; Spikes, J.D.; Bertoloni, G.; Jori, G. Studies on the mechanism of bacteria photosensitization by meso-substituted cationic porphyrins. J. Photochem. Photobiol. B 1996, 35, 149–57. [Google Scholar]
- Zanin, I.C.J.; Gonçalves, R.B.; Junior, A.B.; Keith Hope, C.; Pratten, J. Susceptibility of Streptococcus mutans biofilms to photodynamic therapy: An in vitro study. J. Antimicrob. Chemother 2005, 56, 324–330. [Google Scholar]
- Rontani, J.-F.; Charriere, B.; Forest, A.; Heussner, S.; Vaultier, F.; Petit, M.; Delsaut, N.; Fortier, L.; Sempéré, R. Intense photooxidative degradation of planktonic and bacterial lipids in sinking particles collected with sediment traps across the Canadian Beaufort Shelf (Arctic Ocean). Biogeosciences 2012, 9, 4787–4802. [Google Scholar]
- Frimer, A.A. The reaction of singlet oxygen with olefins: The question of mechanism. Chem. Rev 1979, 79, 359–387. [Google Scholar]
- Frankel, E.N. Lipid Oxidation; The Oily Press: Dundee, UK, 1998; pp. 1–302. [Google Scholar]
- Porter, N.A.; Caldwell, S.E.; Mills, K.A. Mechanisms of free radical oxidation of unsaturated lipids. Lipids 1995, 30, 277–290. [Google Scholar]
- Aebisher, D.; Azar, N.S.; Zamadar, M.; Gandra, N.; Gafney, H.D.; Gao, R.; Greer, A. Singlet oxygen chemistry in water: A porous vycor glass-supported photosensitizer. J. Phys. Chem. B 2008, 112, 1913–1917. [Google Scholar]
- Jin, N.; Lahaye, D.E.; Groves, J.T. A “push-pull” mechanism for heterolytic O–O bond cleavage in hydroperoxo manganese porphyrins. Inorg. Chem 2010, 49, 11516–11524. [Google Scholar]
- Dahl, T.A.; Midden, W.R.; Hartman, P.E. Comparison of killing of gram-negative and gram-positive bacteria by pure singlet oxygen. J. Bacteriol 1989, 171, 2188–2194. [Google Scholar]
- Horspool, W.M.; Armesto, D. Organic Photochemistry: A Comprehensive Treatment; Horwood, E., Ed.; Prentice Hall: Chichester, UK, 1992; p. 521. [Google Scholar]
- Schaich, K.M. Lipid Oxidation: Theoretical Aspects. In Bailey’s Industrial Oil and Fat Products; Jonh Wiley & Sons: Hoboken, NJ, USA, 2005; pp. 269–355. [Google Scholar]
- Berti, G. Stereochemical aspects of the synthesis of 1,2-epoxides. Top. Stereochem 1973, 7, 95–234. [Google Scholar]
- Fossey, J.; Lefort, D.; Sorba, J. Free Radicals in Organic Chemistry; Masson, Ed.; Wiley; Paris, France, 1995; p. 307. [Google Scholar]
- Blée, E.; Schuber, F. Efficient epoxidation of unsaturated fatty acids by a hydroperoxide-dependent oxygenase. J. Biol. Chem 1990, 265, 12887–12894. [Google Scholar]
- Pokorny, J. Major Factors Affecting the Autoxidation of Lipids; Chan, H.W., Ed.; Academic Press: London, UK, 1987; pp. 141–206. [Google Scholar]
- Schaich, K.M. Metals and lipid oxidation. Contemporary issues. Lipids 1992, 27, 209–218. [Google Scholar]
- Testa, A.C. Photosensitized cis-trans isomerization of methyl oleate. J. Org. Chem 1964, 29, 2461–2462. [Google Scholar]
- Jacobson, F.S.; Morgan, R.W.; Christmanlf, M.F.; Amesll, B.N. An alkyl hydroperoxide reductase from Salmonella typhimurium involved in the defense of DNA against oxidative damage. J. Biol. Chem 1989, 264, 1488–1989. [Google Scholar]
- Rontani, J.-F.; Harji, R.; Guasco, S.; Prahl, F.G.; Volkman, J.K.; Bhosle, N.B.; Bonin, P. Degradation of alkenones by aerobic heterotrophic bacteria: Selective or not? Org. Geochem 2008, 39, 34–51. [Google Scholar]
- Volkman, J.K.; Farmer, C.L.; Barrett, S.M.; Sikes, E.L. Unusual dihydroxysterols as chemotaxonomic markers for microalgae from the order pavlovales (haptophyceae). J. Phycol 1997, 33, 1016–1023. [Google Scholar]
- Marchand, D.; Rontani, J.-F. Visible light-induced oxidation of lipid components of purple sulfur bacteria: A significant process in microbial mats. Org. Geochem 2003, 34, 61–79. [Google Scholar]
- Mihara, S.; Tateba, H. Photosensitized oxygenation reactions of phytol and its derivatives. J. Org. Chem 1986, 51, 1142–1144. [Google Scholar]
- Pierce, A.E. Silylation of Organic Compounds; Pierce Chemical Co.: Rockford, IL, USA, 1982. [Google Scholar]
- Marker, A.F.H. The use of acetone and methanol in the estimation of chlorophyll in the presence of phaeophytin. Freshw. Biol 1972, 2, 361–385. [Google Scholar]
- Holm-Hansen, O.; Lorenzen, C.J.; Holmes, R.W.; Strickland, J.D.H. Fluorometric determination of chlorophyll. ICES J. Mar. Sci 1965, 30, 3–15. [Google Scholar]
- Halliwell, B.; Gutteridget, J.M.C. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J 1984, 219, 1–14. [Google Scholar]
- Dias Cavalcante, A.K.; Martinez, G.R.; di Mascio, P.; Martins Menck, C.F.; Lucymara Fassarella, A.-L. Cytotoxicity and mutagenesis induced by singlet oxygen in wild type and DNA repair deficient Escherichia coli strains. DNA Repair 2002, 1, 1051–1056. [Google Scholar]
- Davies, M.J. The oxidative environment and protein damage. Biochim. Biophys. Acta 2005, 1703, 93–109. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cis-vaccenic acid concentration (μg·L−1) | Cis-vaccenic acid oxidation products concentration (μg·L−1) | Sum (μg·L−1) |
|---|---|---|
| 308.7 | 18.0 | 326.7 |
| 292.7 | 40.0 | 323.7 |
| 260.8 | 38.2 | 299.0 |
| 258.7 | 69.8 | 328.6 |
| 257.9 | 63.4 | 321.2 |
| Average: 321.6 ± 13.3 |
| E. huxleyi strain | Reaction rate constants (k) of cis-vaccenic photoproducts formation (h−1) |
|---|---|
| TW1 | 0.152 |
| CS814 | 0.138 |
| CS57 | 0.173 |
| Average : 0.153 ± 0.018 |
| Percentage of D2O | Increase of cis-vaccenic photooxidation products relative to the pure H2O sample (%) |
|---|---|
| 0 | 0 |
| 25 | 2.5 |
| 50 | 76.5 |
| 75 | 50.2 |
| 100 | 70.4 |
| Irradiation time (h) | Non-axenic senescent cells of E. huxleyi | Bacterial community isolated from E. huxleyi cells |
|---|---|---|
| 0 | 5.8 | 0 |
| 7.5 | 27.0 | 0 |
| 31.5 | 9.9 | 0.6 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Petit, M.; Sempéré, R.; Vaultier, F.; Rontani, J.-F. Photochemical Production and Behavior of Hydroperoxyacids in Heterotrophic Bacteria Attached to Senescent Phytoplanktonic Cells. Int. J. Mol. Sci. 2013, 14, 11795-11815. https://doi.org/10.3390/ijms140611795
Petit M, Sempéré R, Vaultier F, Rontani J-F. Photochemical Production and Behavior of Hydroperoxyacids in Heterotrophic Bacteria Attached to Senescent Phytoplanktonic Cells. International Journal of Molecular Sciences. 2013; 14(6):11795-11815. https://doi.org/10.3390/ijms140611795
Chicago/Turabian StylePetit, Morgan, Richard Sempéré, Frédéric Vaultier, and Jean-François Rontani. 2013. "Photochemical Production and Behavior of Hydroperoxyacids in Heterotrophic Bacteria Attached to Senescent Phytoplanktonic Cells" International Journal of Molecular Sciences 14, no. 6: 11795-11815. https://doi.org/10.3390/ijms140611795