Towards a Molecular Understanding of the Biosynthesis of Amaryllidaceae Alkaloids in Support of Their Expanding Medical Use

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. The Amaryllidaceae Family and Their Medicinal Alkaloids
2.1. The Extent of the Amaryllidaceae Family
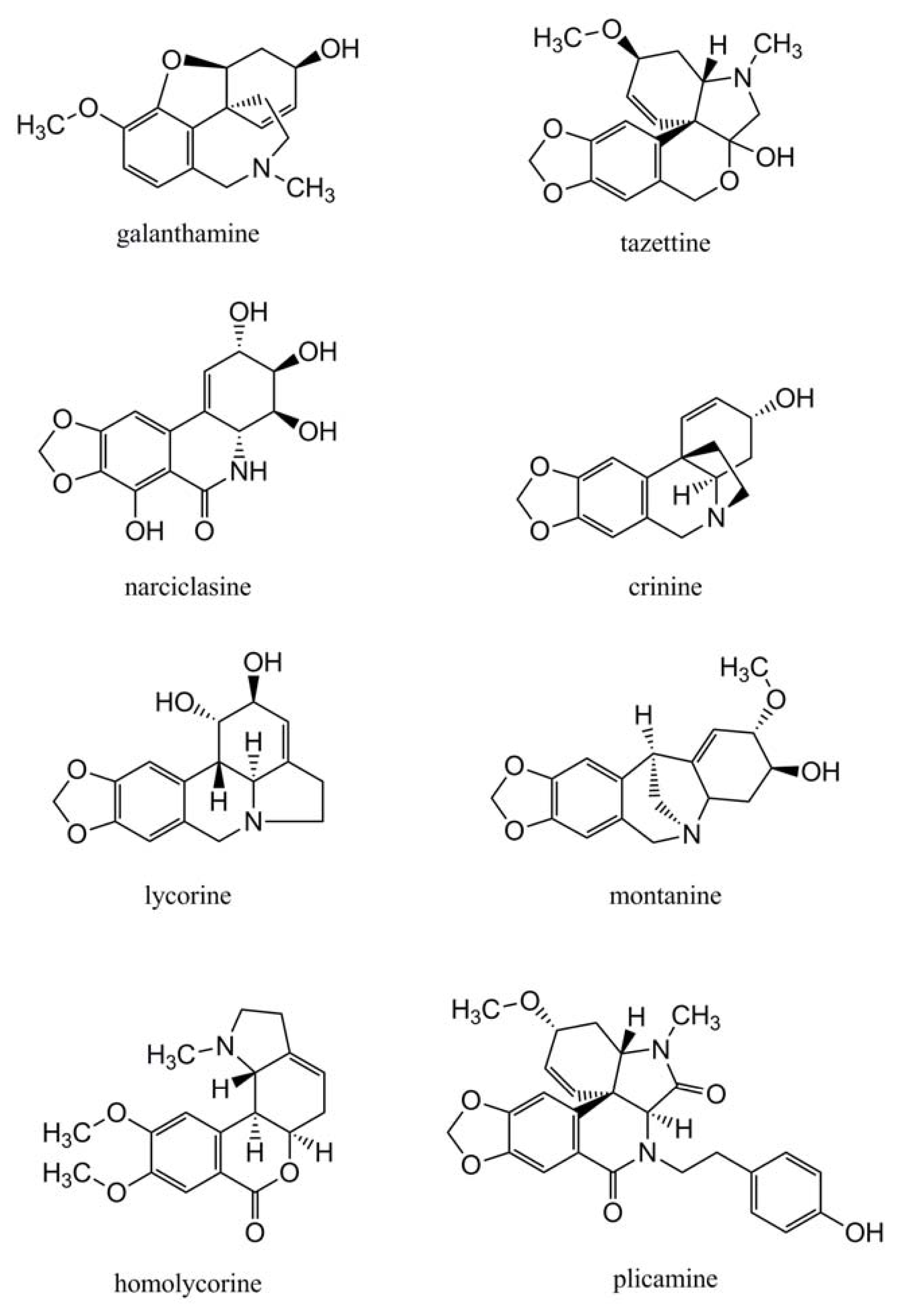
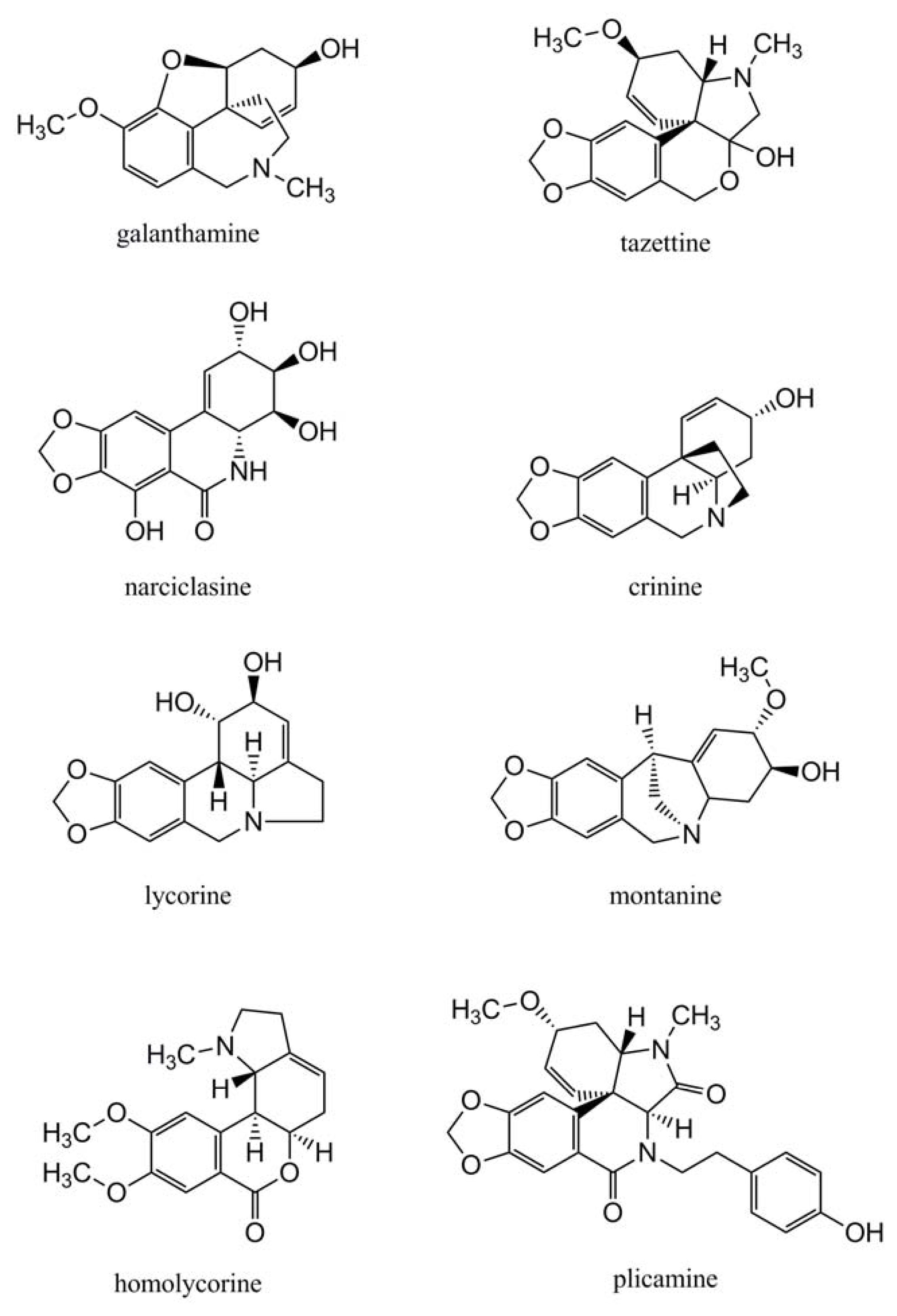
2.2. Amaryllidaceae Alkaloids
2.3. Galanthamine in the Treatment of Alzheimer’s Disease and Other Neurological Conditions
2.4. Amaryllidaceae Alkaloids and Their Pharmacological Activities
3. Ecological and Physiological Aspects of Amaryllidaceae Alkaloids
3.1. Chemical Ecology of Amaryllidaceae Alkaloids
3.2. Physiology of Amaryllidaceae Alkaloids
4. A Biochemical and Molecular Understanding of Amaryllidaceae Alkaloid Biosynthesis
4.1. Present Knowledge on Amaryllidaceae Alkaloid Biosynthesis
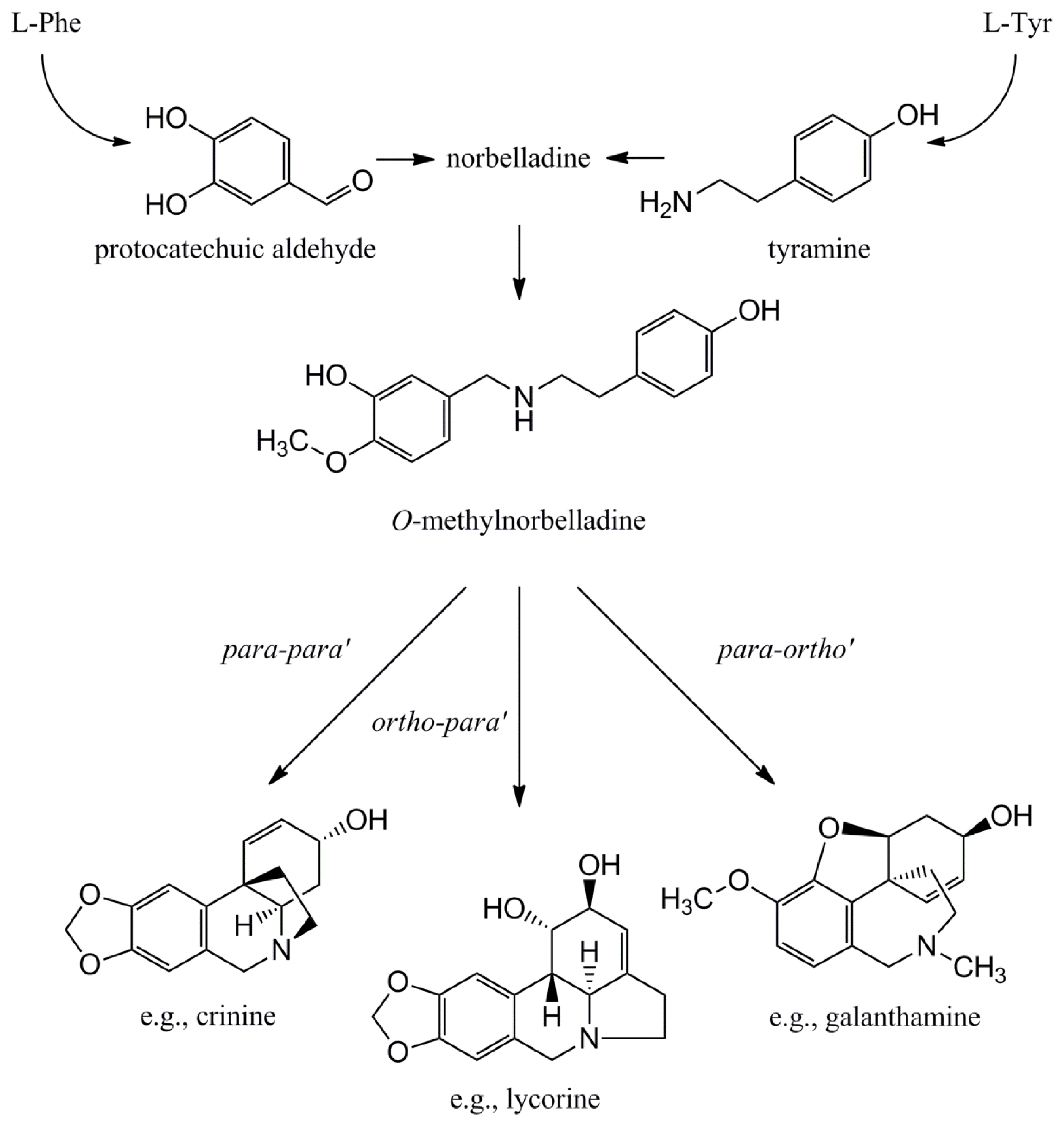
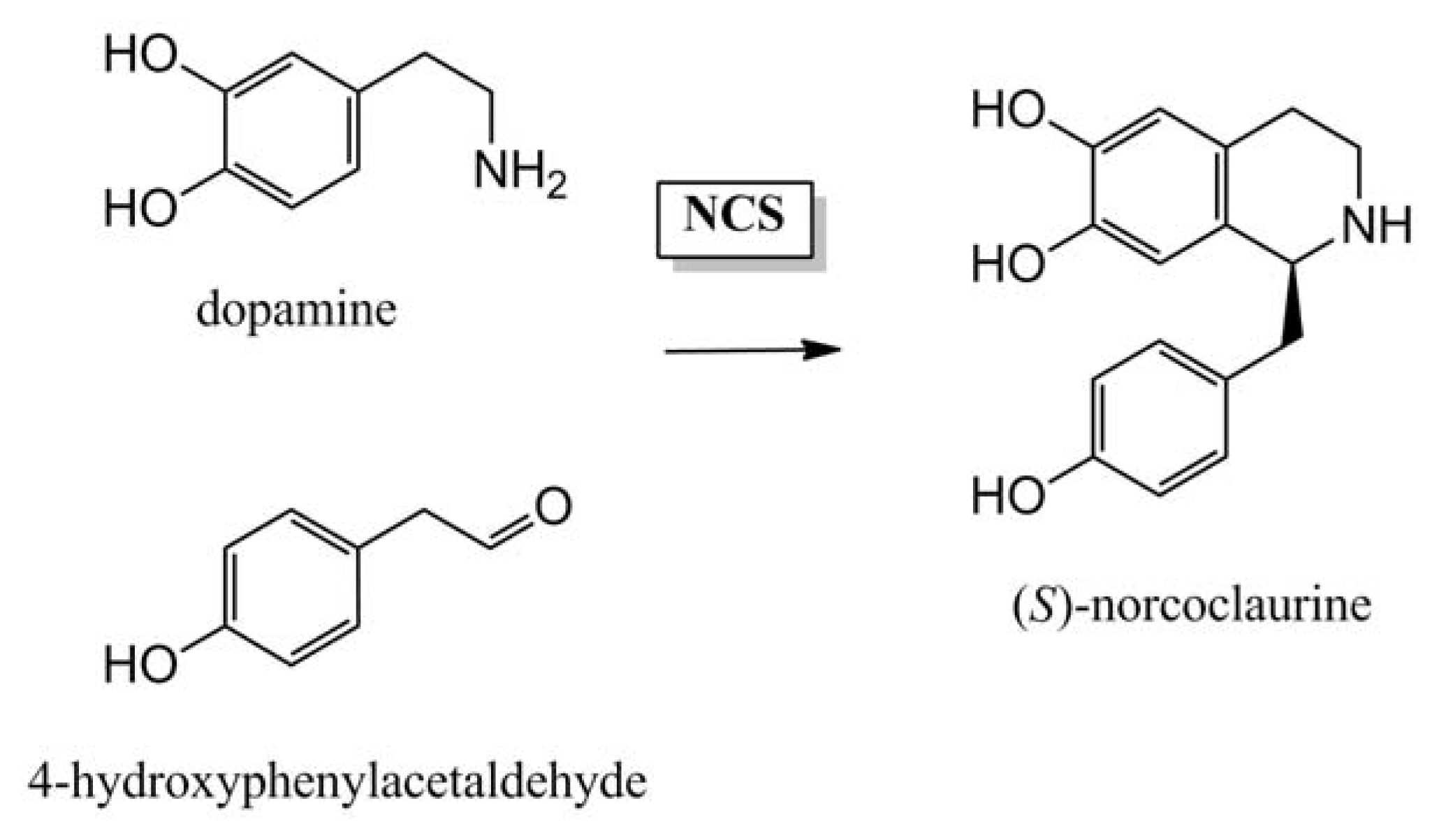
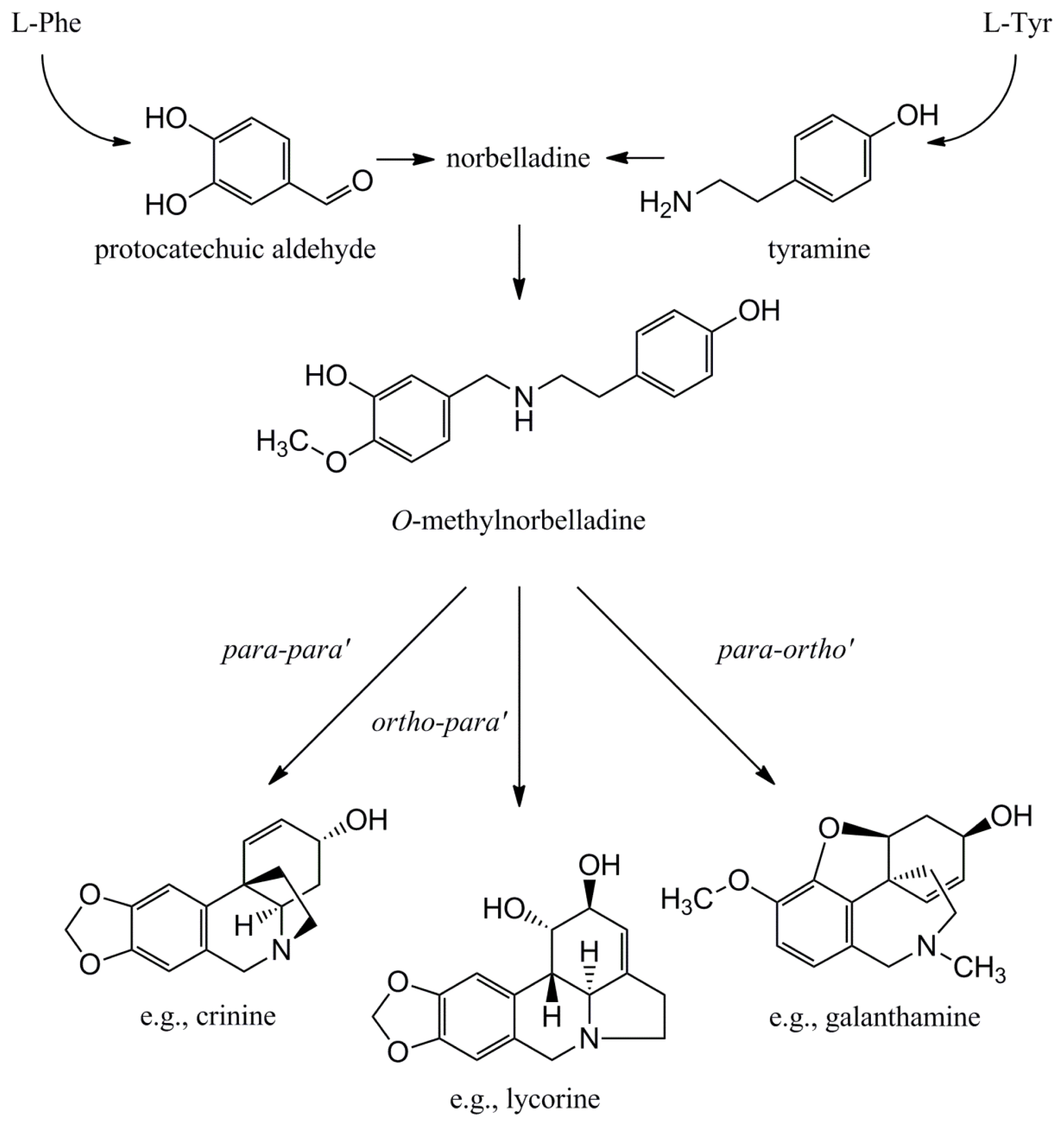
4.1.1. Initial Biosynthetic Steps
4.1.2. Phenol Coupling
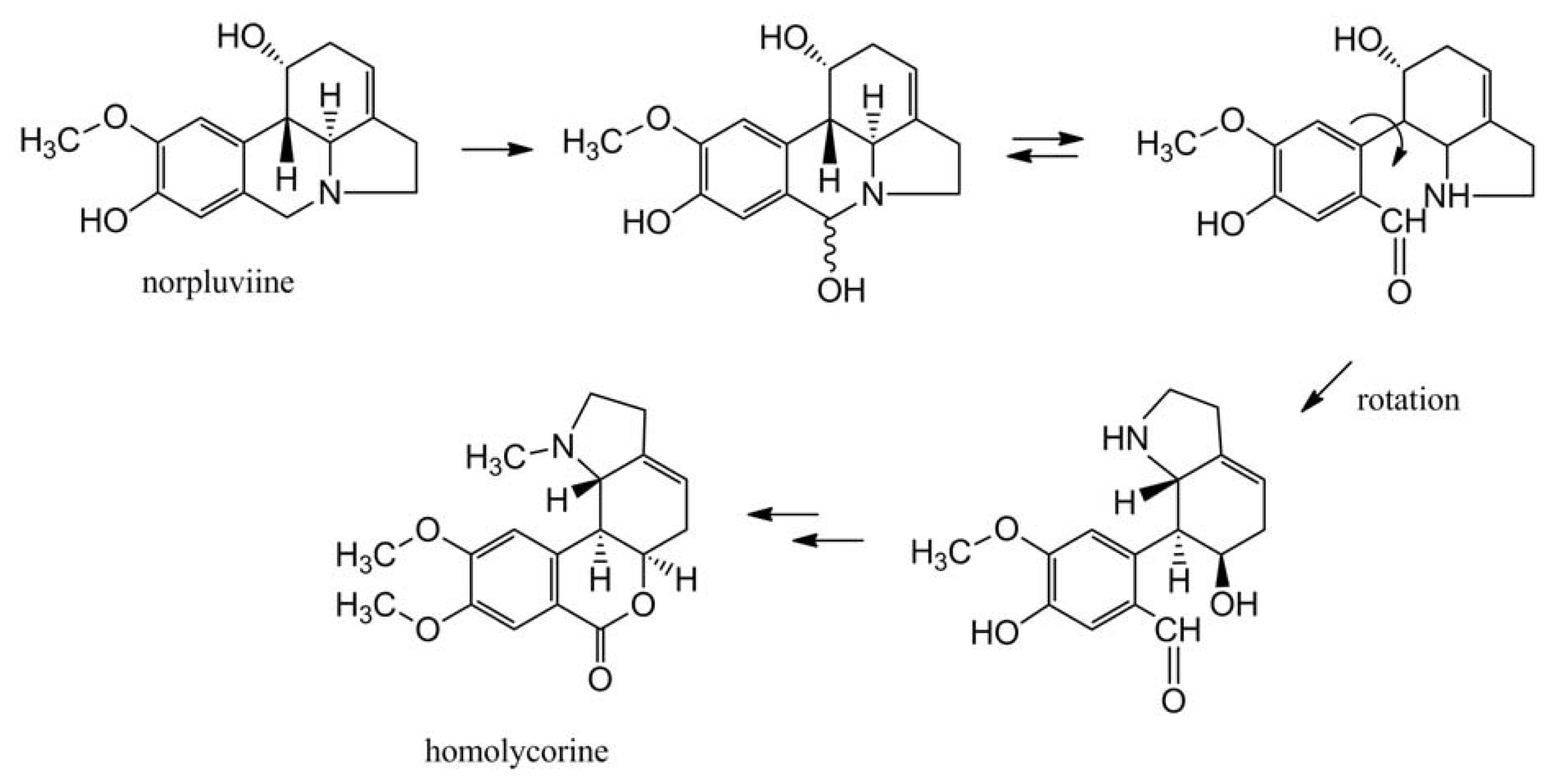
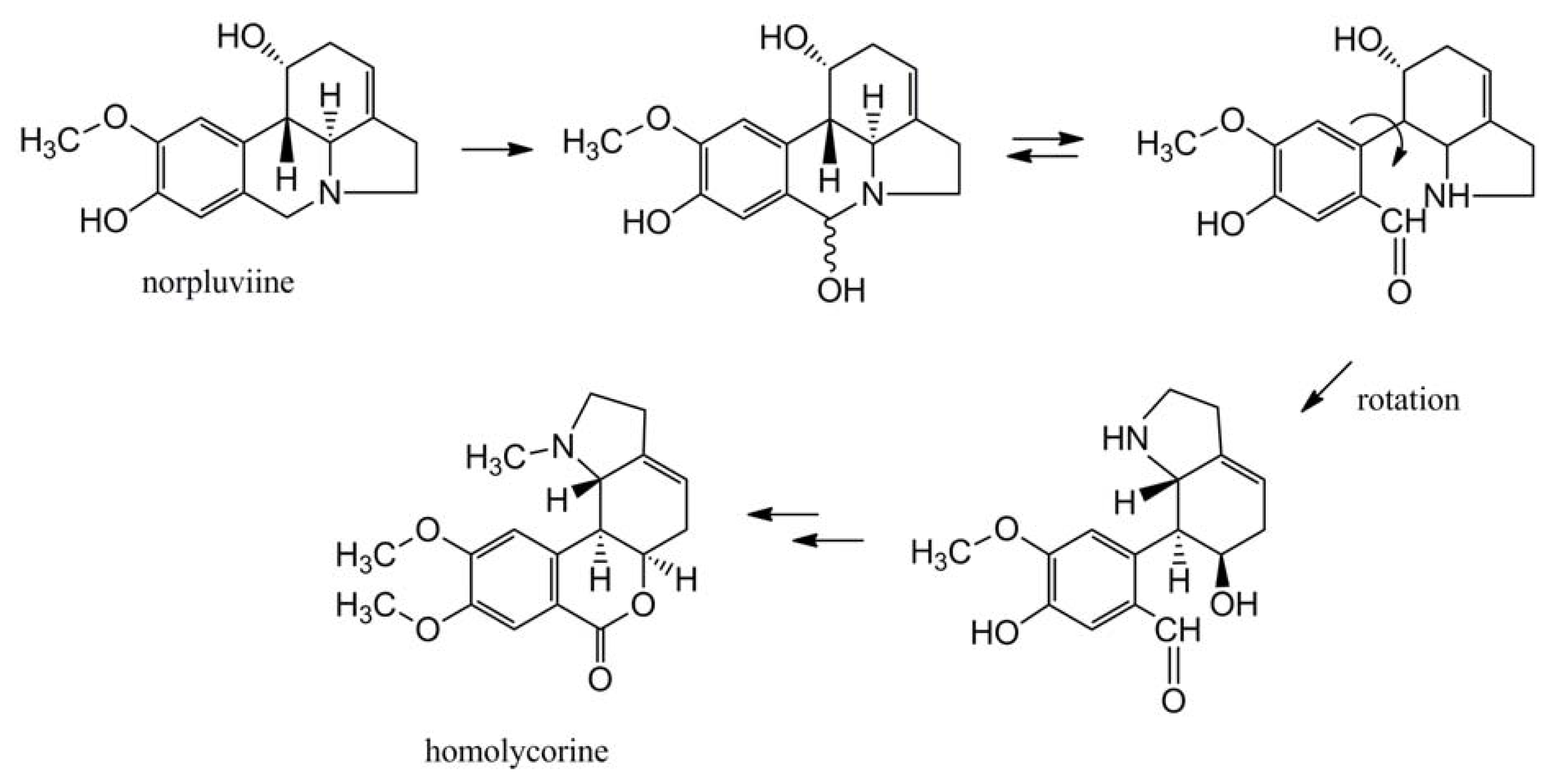
4.1.3. Further Skeleton Structures, Decorations and Modifications
4.2. Gene Discovery Using Transcriptomics
4.3. The Potential Use of Biosynthetic Gene Clusters in Gene Discovery
5. Perspectives on Amaryllidaceae Alkaloid Production
5.1. Examples of Present Commercial Alkaloid Production
5.2. Amaryllidaceae Species and Present Commercial Galanthamine Production
5.3. Perspectives on Amaryllidaceae Alkaloid Production
6. Conclusions
Acknowledgments
Conflict of Interest
References
- Ziegler, J.; Facchini, P.J. Alkaloid biosynthesis: Metabolism and trafficking. Annu. Rev. Plant Biol 2008, 59, 735–769. [Google Scholar]
- Nosov, A.M. Application of cell technologies for production of plant-derived bioactive substances of plant origin. Appl. Biochem. Microbiol 2012, 48, 609–624. [Google Scholar]
- Marco-Contelles, J.; do Carmo Carreiras, M.; Rodriguez, C.; Villarroya, M.; Garcia, A.G. Synthesis and pharmacology of galanthamine. Chem. Rev 2006, 106, 116–133. [Google Scholar]
- De Luca, V.; Salim, V.; Atsumi, S.M.; Yu, F. Mining the biodiversity of plants: A revolution in the making. Science 2012, 336, 1658–1661. [Google Scholar]
- Gates, M.; Tschudi, G. The synthesis of morphine. J. Am. Chem. Soc 1952, 74, 1109–1110. [Google Scholar]
- Stork, G.; Yamashita, A.; Adams, J.; Schulte, G.R.; Chesworth, R.; Miyazaki, Y.; Farmer, J.J. Regiospecific and stereoselective syntheses of (±) morphine, codeine, and thebaine via a highly stereocontrolled intramolecular 4 + 2 cycloaddition leading to a phenanthrofuran system. J. Am. Chem. Soc 2009, 131, 11402–11406. [Google Scholar]
- Mascavage, L.M.; Wilson, M.L.; Dalton, D.R. Syntheses of morphine and codeine (1992–2002): templates for exploration of synthetic tools. Curr. Org. Synth 2006, 3, 99–120. [Google Scholar]
- Phyton Biotech. Available online: http://www.phytonbiotech.com (accessed on 20 April 2013).
- Heinrich, M.; Teoh, H.L. Galanthamine from snowdrop—The development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmcol 2004, 92, 147–162. [Google Scholar]
- Lubbe, A.; Choi, Y.H.; Vreeburg, P.; Verpoorte, R. Effect of fertilizers on galanthamine and metabolite profiles in Narcissus bulbs by 1H NMR. J. Agric. Food Chem 2011, 59, 3155–3161. [Google Scholar]
- Berkov, S.; Georgieva, L.; Kondakova, V.; Atanassov, A.; Viladomat, F.; Bastida, J.; Codina, C. Plant sources of galanthamine: Phytochemical and biotechnological aspects. Biotechnol. Biotechnol. Eq 2009, 23, 1170–1176. [Google Scholar]
- Evidente, A.; Kornienko, A. Anticancer evaluation of structurally diverse Amaryllidaceae alkaloids and their synthetic derivatives. Phytochem. Rev 2009, 8, 449–459. [Google Scholar]
- Lefranc, F.; Sauvage, S.; van Goietsenoven, G.; Megalizzi, V.; Lamoral-Theys, D.; Debeir, O.; Spiegl-Kreinecker, S.; Berger, W.; Mathieu, V.; Decaestecker, C.; et al. Narciclasine, a plant growth modulator, activates Rho and stress fibers in glioblastoma cells. Mol. Cancer Ther 2009, 8, 1739–1750. [Google Scholar]
- Van Goietsenoven, G.; Hutton, J.; Becker, J.-P.; Lallemand, B.; Robert, F.; Lefranc, F.; Pirker, C.; Vandenbussche, G.; van Antwerpen, P.; Evidente, A.; et al. Targeting of eEF1a with Amaryllidaceae isocarbostyrils as a strategy to combat melanomas. FASEB J 2010, 24, 4574–4584. [Google Scholar]
- Van Goietsenoven, G.; Mathieu, V.; Lefranc, F.; Kornienko, A.; Evidente, A.; Kiss, R. Narciclasine as well as other Amaryllidaceae isocarbostyrils are promising GTP-ase targetting agents against brain cancers. Med. Res. Rev 2013, 33, 439–455. [Google Scholar]
- Chase, M.W.; Reveal, J.L.; Fay, M.F. A subfamilial classification for the expanded asparagalean families Amaryllidaceae, Asparagaceae and Xanthorrhoeaceae. Bot. J. Linn. Soc 2009, 161, 132–136. [Google Scholar]
- O’Donnell, G.; Gibbons, S. Antibacterial activity of two canthin-6-one alkaloids from Allium neapolitanum. Phytother. Res 2007, 21, 653–657. [Google Scholar]
- Louw, C.A.M.; Regnier, T.J.C.; Korsten, L. Medicinal bulbous plants of South Africa and their traditional relevance in the control of infectious diseases. J. Ethnopharmcol 2002, 82, 147–154. [Google Scholar]
- Rønsted, N.; Savolainen, V.; Mølgaard, P.; Jäger, A.K. Phylogenetic selection of Narcissus species for drug discovery. Biochem. Syst. Ecol 2008, 36, 417–422. [Google Scholar]
- Kornienko, A.; Evidente, A. Chemistry, biology, and medicinal potential of narciclasine and its congeners. Chem. Rev 2008, 108, 1982–2014. [Google Scholar]
- Fennell, C.W.; van Staden, J. Crinum species in traditional and modern medicine. J. Ethnopharmacol 2001, 78, 15–26. [Google Scholar]
- De Andrade, J.P.; Pigni, N.B.; Torras-Claveria, L.; Guo, Y.; Berkov, S.; Reyes-Chilpa, R.; El Amrani, A.; Zuanazzi, J.A.S.; Codina, C.; Viladomat, F.; et al. Alkaloids from the Hippeastrum genus: Chemistry and biological activity. Rev. Latinoamer. Quím 2012, 40, 83–98. [Google Scholar]
- Jin, Z. Amaryllidaceae and Sceletium alkaloids. Nat. Prod. Rep 2011, 28, 1126–1142. [Google Scholar]
- Bay-Smidt, M.G.K.; Jäger, A.K.; Krydsfeldt, K.; Meerow, A.W.; Stafford, G.I.; van Staden, J.; Rønsted, N. Phylogenetic selection of target species in Amaryllidaceae tribe Haemantheae for acetylcholinesterase inhibition and affinity to the serotonin reuptake transport protein. S. Afr. J. Bot 2011, 77, 175–183. [Google Scholar]
- Cedron, J.C.; Arco-Aguilar, M.D.; Estevez-Braun, A.; Ravelo, A.G. Chemistry and Biology of Pancratium Alkaloids. In The Alkaloids; Cordell, G.A., Ed.; Academic Press: Chennai, India, 2010; pp. 1–37. [Google Scholar]
- Tram, N.T.N.; Titorenkova, T.V.; Bankova, V.S.; Handjieva, N.V.; Popov, S.S. Crinum L. (Amaryllidaceae). Fitoterapia 2002, 73, 183–208. [Google Scholar]
- Shi, S.; Qiu, Y.; Wu, L.; Fu, C. Interspecific relationships of Lycoris (Amaryllidaceae) inferred from inter-simple sequence repeat data. Sci. Hortic 2006, 110, 285–291. [Google Scholar]
- Xuan, S.Q.; Zheng, J.Y.; Wang, H.; Gao, P.; Zhou, S.B.; Liu, K.; Zhu, G.-P. Polymorphic microsatellite loci for the genetic analysis of Lycoris radiate (Amaryllidaceae) and cross-amplification in other congeneric species. Genet. Mol. Res 2011, 110, 3141–3145. [Google Scholar]
- Berkov, S.; Georgieva, L.; Kondakova, V.; Viladomat, F.; Bastida, J.; Atanassov, A.; Codina, C. The geographic isolation of Leucojum aestivum populations leads to divergation of alkaloid biosynthesis. Biochem. Syst. Ecol 2013, 46, 152–161. [Google Scholar]
- Santos-Gally, R.; Vargas, P.; Arroyo, J. Insights into Neogene Mediterranean biogeography based on phylogenetic relationships of mountain and lowland lineages of Narcissus (Amaryllidaceae). J. Biogeogr 2012, 39, 782–798. [Google Scholar]
- Royal Horticultural Society, The Daffodil Register and Classified List. Available online: http://www.rhs.org.uk/plants/plant-science/plant-registration/daffodils (accessed on 19 March 2013).
- Bastida, J.; Berkov, S.; Torras, L.; Pigni, N.B.; de Andrade, J.P.; Martinez, V.; Codina, C.; Viladomat, F. Chemical and Biological Aspects of Amaryllidaceae Alkaloids. In Recent Advances in Pharmaceutical Sciences; Munoz-Torrero, D., Ed.; Transworld Research Network: Kerala, India, 2011; pp. 65–100. [Google Scholar]
- Ghosal, S.; Saini, K.S.; Razdan, S. Crinum alkaloids: Their chemistry and biology. Phytochemistry 1985, 24, 2141–2156. [Google Scholar]
- Unver, N. New skeletons and new concepts in Amaryllidaceae alkaloids. Phytochem. Rev 2007, 6, 125–135. [Google Scholar]
- Jin, Z. Amaryllidaceae and Sceletium alkaloids. Nat. Prod. Rep 2009, 26, 363–381. [Google Scholar]
- Miyakado, M.; Kato, T.; Ohno, N.; Koshimizu, K. Alkaloids of Urginea altissima and their antimicrobial activity against Phytophthora capsici. Phytochemistry 1975, 14, 2717. [Google Scholar]
- Pohl, T.; Koorbanally, C.; Crouch, N.R.; Mulholland, D.A. Bufadienolides from Drimia robusta and Urginea altissima (Hyacinthaceae). Phytochemistry 2001, 58, 557–561. [Google Scholar]
- Mulholland, D.A.; Crouch, N.; Decker, B.; Smith, M.T. The isolation of the Amaryllidaceae alkaloid crinamine from Dioscorea dregeana (Dioscoreaceae). Biochem. Syst. Ecol 2002, 30, 183–185. [Google Scholar]
- Bjarnholt, N.; Rook, F.; Motawia, M.S.; Cornett, C.; Jørgensen, C.; Olsen, C.E.; Jaroszewski, J.W.; Bak, S.; Møller, B.L. Diversification of an ancient theme: Hydroxynitrile glucosides. Phytochemistry 2008, 69, 1507–1516. [Google Scholar]
- Takos, A.M.; Knudsen, C.; Lai, D.; Kannangara, R.; Mikkelsen, L.; Motawia, M.S.; Olsen, C.E.; Sato, S.; Tabata, S.; Jørgensen, K.; et al. Genomic clustering of cyanogenic glucoside biosynthetic genes aids their identification in Lotus japonicus and suggests the repeated evolution of this chemical defence pathway. Plant J 2011, 68, 273–286. [Google Scholar]
- Pichersky, E.; Lewinsohn, E. Convergent evolution in plant specialized metabolism. Annu. Rev. Plant Biol 2011, 62, 549–566. [Google Scholar]
- Reimann, A.; Nurhayati, N.; Backenköhler, A.; Ober, D. Repeated evolution of the pyrrolizidine alkaloid-mediated defense system in separate angiosperm lineages. Plant Cell 2004, 16, 2772–2784. [Google Scholar]
- Frey, M.; Schullehner, K.; Dick, R.; Fiesselmann, A.; Gierl, A. Benzoxazinoid biosynthesis, a model for evolution of secondary metabolic pathways in plants. Phytochemistry 2009, 70, 1645–1651. [Google Scholar]
- Dick, R.; Rattei, T.; Haslbeck, M.; Schwab, W.; Gierl, A.; Frey, M. Comparative analysis of benzoxazinoid biosynthesis in monocots and dicots: Independent recruitment of stabilization and activation functions. Plant Cell 2012, 24, 915–928. [Google Scholar]
- Jirschitzka, J.; Schmidt, G.W.; Reichelt, M.; Schneider, B.; Gershenzon, J.; D’Auria, J.C. Plant tropane alkaloid biosynthesis evolved independently in the Solanaceae and Erythroxylaceae. Proc. Natl. Acad. Sci. USA 2012, 109, 10304–10309. [Google Scholar]
- Leete, E.; Michelson, R.H. Biosynthesis of dioscorine from trigonelline in Dioscorea hispida. Phytochemistry 1998, 27, 3793–3798. [Google Scholar]
- Wang, Y.-H.; Zhang, Z.-K.; Yang, F.-M.; Sun, Q.-Y.; He, H.-P.; Di, Y.-T.; Mu, S.-Z.; Ly, Y.; Chang, Y.; Zheng, Q.-T.; et al. Benzylphenethylamine alkaloids from Hosta plantaginea with inhibitory activity against Tobacco Mosaic Virus and acetylcholinesterase. J. Nat. Prod 2007, 70, 1458–1461. [Google Scholar]
- Wang, Y.-H.; Gao, S.; Yang, F.-M.; Sun, Q.-Y.; Wang, J.-S.; Liu, H.-Y.; Li, C.-S.; Di, Y.-T.; Li, S.-L.; He, H.-P.; et al. Structure elucidation and biomimetic synthesis of hostasinine A, a new benzylphenethylamine alkaloid from Hosta plantaginea. Org. Lett 2007, 9, 5279–5281. [Google Scholar]
- Keller, C.; Kadir, A.; Forsberg, A.; Porras, O.; Nordberg, A. Long-term effects of galantamine treatment on brain functional activities as measured by PET in Alzheimer’s disease patients. J. Alzheimers Dis 2011, 24, 109–123. [Google Scholar]
- U.S. Food and Drug Administration. Available online: http://www.fda.gov/Drugs/default.htm (accessed on 9 March 2013).
- Santos, M.D.; Alkondon, M.; Pereira, E.F.R.; Aracava, Y.; Eisenberg, H.M.; Maelicke, A.; Albuquerque, E.X. The nicotinic allosteric potentiating ligand galanthamine facilitates synaptic transmission in the mammalian central nervous system. Mol. Pharmacol 2002, 61, 1222–1234. [Google Scholar]
- Egea, J.; Martin-de-Saavedra, M.D.; Parada, E.; Romero, A.; del Barrio, L.; Rosa, A.O.; Garcia, A.G.; Lopez, M.G. Galanthamine elicits neuroprotection by inhibiting iNOS, NADPH oxidase and ROS in hippocampal slices stressed with anoxia/reoxygenation. Neuropharmacology 2012, 62, 1082–1090. [Google Scholar]
- Lorrio, S.; Sobrado, M.; Aria, E.; Roda, J.M.; Garcia, A.G.; Lopez, M.G. Galanthamine postischemia provides neuroprotection and memory recovery against transient global cerebral ischemia in gerbils. J. Pharmacol. Exp. Ther 2007, 322, 591–599. [Google Scholar]
- Hilmas, C.J.; Poole, M.J.; Finneran, K.; Clark, M.G.; Williams, P.T. Galanthamine is a novel post-exposure therapeutic against lethal VX challenge. Toxicol. Appl. Pharm 2009, 240, 166–173. [Google Scholar]
- Pereira, E.F.R.; Aracava, Y.; Alkondon, M.; Akkerman, M.; Merchenthaler, I.; Albuquerque, E.X. Molecular and cellular actions of galanthamine: Clinical implications for treatment of organophosphorus poisoning. J. Mol. Neurosci 2010, 40, 196–203. [Google Scholar]
- Wimo, A.; Price, M. World Alzheimer Report 2010: The global economic impact of dementia. Alzheimer’s Disease International. 2010. Available online: http://www.alz.co.uk (accessed on 20 March 2013).
- Abbott, A. A problem for our age. Nature 2011, 475, S1–S4. [Google Scholar]
- Lamoral-Theys, D.; Andolfi, A.; van Goietsenoven, G.; Cimmino, A.; Le Calve, B.; Wauthoz, N.; Megalizzi, V.; Gras, T.; Bruyere, C.; Dubois, J.; et al. Lycorine, the main phenanthridine Amaryllidaceae alkaloid, exhibits significant antitumor activity in cancer cells that display resistance to proapoptotic stimuli: An investigation of structure-activity relationship and mechanistic insight. J. Med. Chem 2009, 52, 6244–6256. [Google Scholar]
- Hayden, R.E.; Pratt, G.; Drayson, M.T.; Bunce, C.M. Lycorine sensitizes CD40 ligand-protected chronic lymphocytic leukemia cells to bezafibrate- and medroxyprogesterone acetate- induced apoptosis but dasatanib does not overcome reported CD40-mediated drug resistance. Haematologica 2010, 95, 1889–1896. [Google Scholar]
- Luchetti, G.; Johnston, R.; Mathieu, V.; Lefranc, F.; Hayden, K.; Andolfi, A.; Lamoral-Theys, D.; Reisenauer, M.R.; Champion, C.; Pelly, S.C.; et al. Bulbispermine: A crinine-type Amaryllidaceae alkaloid exhibiting cytostatic activity towards apoptosis-resistant glioma cells. ChemMedChem 2012, 7, 815–822. [Google Scholar]
- Likhitwitayawuid, K.; Angerhofer, C.K.; Chai, H.; Pezzuto, J.M.; Cordell, G.A. Cytotoxic and antimalarial alkaloids from the bulbs of Crinum amabile. J. Nat. Prod 1993, 56, 1331–1338. [Google Scholar]
- Giordani, R.B.; Junior, C.O.R.; de Andrade, J.P.; Bastida, J.; Zuanazzi, J.A.S.; Tasca, T.; de Almeida, M.V. Lycorine derivatives against Trichomonas vaginalis. Chem. Biol. Drug. Des 2012, 80, 129–133. [Google Scholar]
- Casida, J.E. Pest toxicology: The primary mechanisms of pesticide action. Chem. Res. Toxicol 2009, 22, 609–619. [Google Scholar]
- Mutero, A.; Pralavorio, M.; Bride, J.-M.; Fournier, D. Resistance-associated point mutations in insecticide-insensitive acetylcholinesterase. Proc. Natl. Acad. Sci. USA 1994, 91, 5922–5926. [Google Scholar]
- Ilg, T.; Schmalz, S.; Werr, M.; Cramer, J. Acetylcholinesterase of the cat flea Ctenocephalides felis: Identification of two distinct genes and biochemical characterization of recombinant and in vivo enzyme activities. Insect Biochem. Mol 2010, 40, 153–164. [Google Scholar]
- Piiroinen, S.; Lindström, L.; Lyytinen, A.; Mappes, J.; Chen, Y.H.; Izzo, V.; Grapputo, A. Pre-invasion history and demography shape the genetic variation in the insecticide resistance-related acetylcholinesterase 2 gene in the invasive Colorado potato beetle. BMC Evol. Biol 2013, 13, 13. [Google Scholar]
- Santana, O.; Reina, M.; Anaya, A.L.; Hernandez, F.; Izquierdo, M.E; Gonzalez-Coloma, A. 3-O-Acetyl-narcissidine, a bioactive alkaloid from Hippeastrum puniceum Lam. (Amaryllidaceae). Z. Naturforsch. 2008, 63c, 639–643. [Google Scholar]
- Opitz, S.E.W.; Müller, C. Plant chemistry and insect sequestration. Chemoecology 2009, 19, 117–154. [Google Scholar]
- Chichiricco, G. Infestation of Narcissus poeticus by a scathophagid fly, Norellia melaleuca (Diptera: Scathophagidae). Eur. J. Entomol 2010, 107, 677–680. [Google Scholar]
- Iqbal, Z.; Nasir, H.; Hiradate, S.; Fujii, Y. Plant growth inhibitory activity of Lycoris radiate Herb. and the possible involvement of lycorine as an allelochemical. Weed Biol. Manag 2006, 6, 221–227. [Google Scholar]
- Ghosal, S.; Kumar, Y.; Chakrabarti, D.K.; Lal, J.; Singh, S.K. Parasitism of Imperata cylindrica on Pancratium biflorum and the concomitant chemical changes in the host species. Phytochemistry 1986, 25, 1097–1102. [Google Scholar]
- Ceriotti, G. Narciclasine: An antimitotic substance from Narcissus bulbs. Nature 1967, 213, 595–596. [Google Scholar]
- Hu, Y.; Yang, L.; Na, X.; You, J.; Hu, W.; Liang, X.; Liu, J.; Mao, L.; Wang, X.; Wang, H.; et al. Narciclasine inhibits the responses of Arabidopsis roots to auxin. Planta 2012, 236, 597–612. [Google Scholar]
- Lu, H.; Wan, Q.; Wang, H.; Na, X.; Wang, X.; Bi, Y. Oxidative stress and mitochondrial dysfunctions are early events in narciclasine-induced programmed cell death in tobacco Bright Yellow-2 cells. Physiol. Plantarum 2012, 144, 48–58. [Google Scholar]
- Bruynzeel, D.P. Bulb dermatitis. Dermatological problems in the flower bulb industries. Contact Dermatitis 1997, 37, 70–77. [Google Scholar]
- Julian, C.G.; Bowers, P.W. The nature and distribution of daffodil pickers’ rash. Contact Dermatitis 1997, 37, 259–262. [Google Scholar]
- Franceschi, V.R.; Nakata, P.A. Calcium oxalate in plants: Formation and function. Annu. Rev. Plant Biol 2005, 56, 41–71. [Google Scholar]
- Scepankova, I.; Hudak, J. Lead and tepal anatomy, plastid ultrastructure and chlorophyll content in Galanthus nivalis L. and Leucojum aestivum L. Plant Syst. Evol 2004, 243, 211–219. [Google Scholar]
- Ghosal, S.; Kumar, Y.; Singh, S. Glucosyloxy alkaloids from Pancratium biflorum. Phytochemistry 1984, 23, 1167–1171. [Google Scholar]
- Ruiters, C. Biomass and resource allocation patterns within the bulb of the perennial geophyte Haemanthus pubescens L. subsp. pubescens (Amaryllidaceae) in a periodic arid environment of lowland fynbos, South Africa. J. Arid Environ 1995, 31, 311–323. [Google Scholar]
- Hanks, G.R. The Biology of Narcissus. In Narcissus and Daffodil. The Genus Narcissus; Hanks, G.R., Ed.; Taylor & Francis: London, UK, 2002; pp. 1–18. [Google Scholar]
- Ruamrungsri, S.; Ruamrungsri, S.; Ikarashi, T.; Ohyama, T. Carbohydrate metabolism in Narcissus. J. Hortic. Sci. Biotech 1999, 74, 395–400. [Google Scholar]
- Ruiz, N.; Ward, D.; Saltz, D. Responses of Pancratium sickenbergeri to simulated bulb herbivory: Combining defence and tolerance strategies. J. Ecol 2002, 90, 472–479. [Google Scholar]
- Lambrechts, H.; Rook, F.; Kollöffel, C. Carbohydrate status of tulip bulbs during cold-induced flower stalk elongation and flowering. Plant Physiol 1994, 104, 515–520. [Google Scholar]
- Herranz, J.M.; Copete, M.A.; Ferrandis, P. Environmental regulation of embryo growth, dormancy breaking and germination in Narcissus alcaracensis (Amaryllidaceae), a threatened endemic Iberian daffodil. Am. Midl. Nat 2013, 169, 147–167. [Google Scholar]
- Hanks, G.R. Commercial Production of Narcissus Bulbs. In Narcissus and Daffodil. The Genus Narcissus; Hanks, G.R., Ed.; Taylor & Francis: London, UK, 2002; pp. 53–130. [Google Scholar]
- Chen, J.; Ziv, M. The effects of storage conditions on starch metabolism and regeneration potentials of twin-scales and inflorescence stem explants of Narcissus tazetta. In Vitro Cell. Dev. Biol. Plant 2005, 41, 816–821. [Google Scholar]
- Lubbe, A.; Verpoorte, R.; Choi, Y.H. Effects of fungicides on galanthamine and metabolite profiles in Narcissus bulbs. Plant Physiol. Biochem 2012, 58, 116–123. [Google Scholar]
- Ru, Q.; Wang, X.; Liu, T.; Zheng, H. Physiological and comparative proteomic analyses in response to nitrogen application in an Amaryllidaceae plant, Lycoris aurea. Acta Physiol. Plant 2013, 35, 271–282. [Google Scholar]
- Colque, R.; Viladomat, F.; Bastida, J.; Codina, C. Improved production of galanthamine and related alkaloids by methyl jasmonate in Narcissus confusus shoot-clumps. Planta Med 2004, 70, 1180–1188. [Google Scholar]
- Lubbe, A.; Gude, H.; Verpoorte, R.; Choi, Y.H. Seasonal accumulation of major alkaloids in organs of pharmaceutical crop Narcissus Carlton. Phytochemistry 2013, 88, 43–53. [Google Scholar]
- Battersby, A.R.; Fales, H.M.; Wildman, W.C. Biosynthesis in the Amaryllidaceae. Tyrosine and norbelladine as precursors of haemanthamine. J. Amer. Chem. Soc 1961, 83, 4098–4099. [Google Scholar]
- Barton, D.H.R.; Kirby, G.W.; Taylor, J.B.; Thomas, G.M. Phenol oxidation and biosynthesis. Part VI. The biogenesis of Amaryllidaceae alkaloids. J. Chem. Soc. 1963, 4545–4558. [Google Scholar]
- Eichhorn, J.; Takada, T.; Kita, Y.; Zenk, M.H. Biosynthesis of the Amaryllidaceae alkaloid galanthamine. Phytochemistry 1988, 49, 1037–1047. [Google Scholar]
- Ghosal, S.; Shanthy, A.; Singh, S.K. Isocraugsodine, an N-arylidenephenethylamine from Crinum asiaticum and its E-Z isomerism. Phytochemistry 1988, 27, 1849–1852. [Google Scholar]
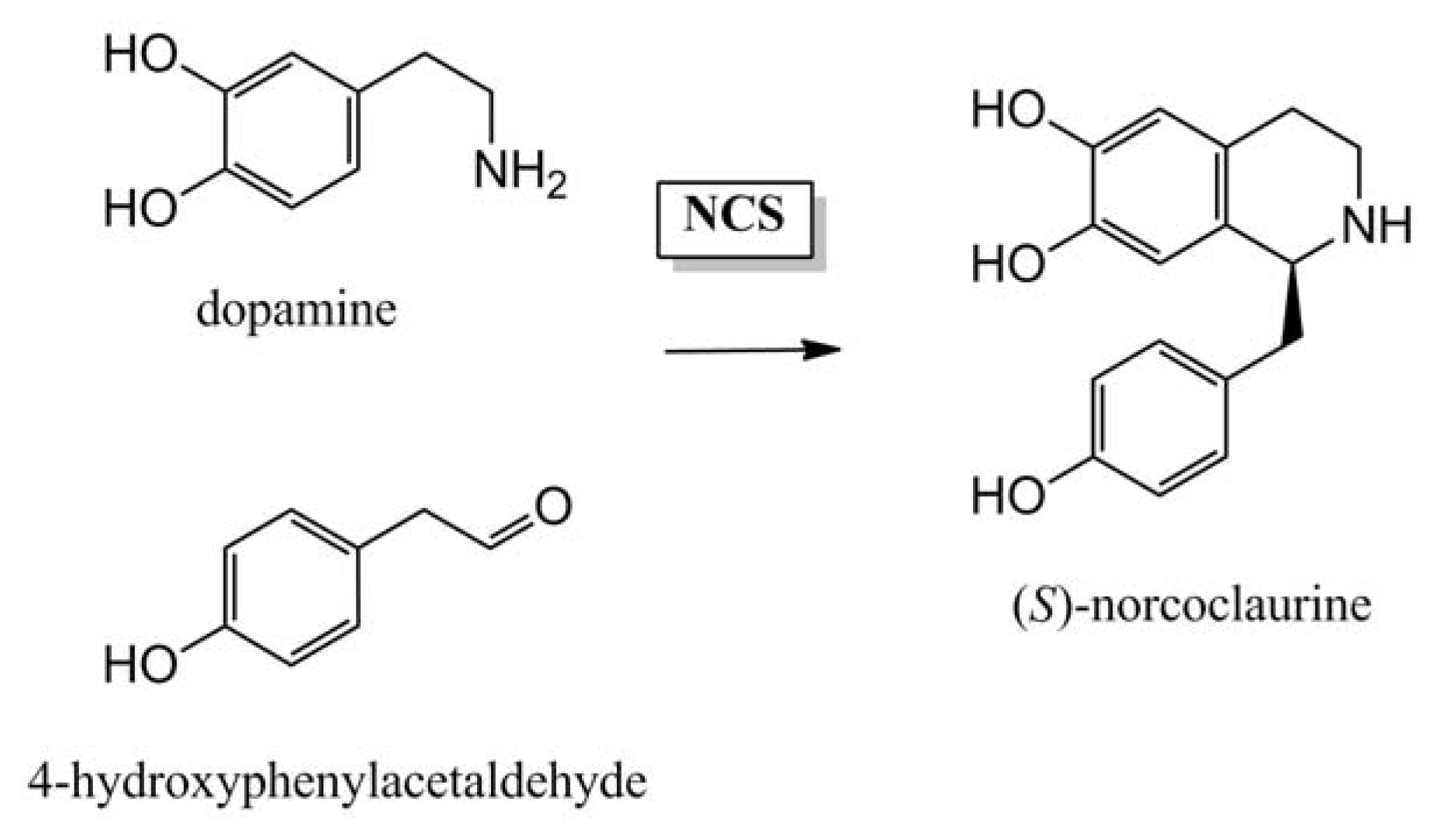
- Luk, L.Y.P.; Bunn, S.; Liscombe, D.K.; Facchini, P.J.; Tanner, M.E. Mechanistic studies on norclaurine synthase of benzylisoquinoline alkaloid biosynthesis: An enzymatic Pictet-Spengler reaction. Biochemistry 2007, 46, 10153–10161. [Google Scholar]
- Samanani, N.; Liscombe, D.K.; Facchini, P.J. Molecular cloning and characterization of norcoclaurine synthase, an enzyme catalyzing the first committed step in benzylisoquinoline alkaloid biosynthesis. Plant J 2004, 40, 302–313. [Google Scholar]
- Lee, E.-J.; Facchini, P. Norcoclaurine synthase is a member of the Pathogenesis-Related 10/Bet v1 protein family. Plant Cell 2010, 22, 3489–3503. [Google Scholar]
- Kutchan, T.M. Strictosidine: From alkaloid to enzyme to gene. Phytochemistry 1993, 32, 493–506. [Google Scholar]
- Kutchan, T.M.; Hampp, N.; Lottspeich, F.; Beyreuther, K.; Zenk, M.H. The cDNA clone for strictosidine synthase from Rauvolfia serpentina. DNA sequence determination and expression in Escherichia coli. FEBS Lett 1988, 237, 40–44. [Google Scholar]
- Ma, X.; Panjikar, S.; Koepke, J.; Loris, E.; Stöckigt, J. The structure of Rauvolfia serpentina strictosidine synthase is a novel six-bladed β-propeller fold in plant proteins. Plant Cell 2006, 18, 907–920. [Google Scholar]
- Weber, B.; Hoesch, L.; Rast, D.M. Protocatechualdehyde and other phenols as cell wall components of grapevine leaves. Phytochemistry 1995, 40, 433–437. [Google Scholar]
- Prachayasittikul, S.; Buraparuangsang, P.; Worachartcheewan, A.; Isarankura-Na-Ayudhya, C.; Ruchirawat, S.; Prachayasittikul, V. Antimicrobial and antioxidative activities of bioactive constituents from Hydnophytum formicarum Jack. Molecules 2008, 13, 904–921. [Google Scholar]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace amines: Identification of a family of mammalian G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar]
- Miller, G.M. The emerging role of trace amine-associated receptor 1 in the functional regulation of monoamine transporters and dopaminergic activity. J. Neurochem 2011, 116, 164–176. [Google Scholar]
- Berkov, S.; Bastida, J.; Sidjimova, B.; Viladomat, F.; Codina, C. Alkaloid diversity in Galanthus elwesii and Galanthus nivalis. Chem. Biodivers 2011, 8, 115–130. [Google Scholar]
- Nair, J.J.; Campbell, W.E.; Brun, R.; Viladomat, F.; Codina, C.; Bastida, J. Alkaloids from Nerine filifolia. Phytochemistry 2005, 66, 373–382. [Google Scholar]
- Cahlikova, L.; Zavadil, S.; Macakova, K.; Valterova, I.; Kulhankova, A.; Hostalkova, A.; Kunes, J.; Opletal, L. Isolation and cholinesterase activity of Amaryllidaceae alkaloids from Nerine bowdenii. Nat. Prod. Commun 2011, 6, 1827–1830. [Google Scholar]
- Kita, Y.; Arisawa, M.; Gyoten, M.; Nakajima, M.; Hamada, R.; Tohma, H.; Takada, T. Oxidative intramolecular phenolic coupling reaction induced by a hypervalent iodine(III) reagent: Leading to galanthamine-type Amaryllidaceae alkaloids. J. Org. Chem 1998, 63, 6625–6633. [Google Scholar]
- Ikezawa, N.; Iwasa, K.; Sato, F. Molecular cloning and characterization of CYP80G2, a cytochrome P450 that catalyzes an intramolecular C–C phenol coupling of (S)-reticuline in magnoflorine biosynthesis, from cultured Coptis japonica cells. J. Biol. Chem 2008, 283, 8810–8821. [Google Scholar]
- Gesell, A.; Rolf, M.; Ziegler, J.; Diaz Chavez, M.L.; Huang, F.-C.; Kutchan, T.M. CYP719B1 is salutaridine synthase, the C–C phenol-coupling enzyme of morphine biosynthesis in opium poppy. J. Biol. Chem 2009, 284, 24432–24442. [Google Scholar]
- Grobe, N.; Zhang, B.; Fisinger, U.; Kutchan, T.M.; Zenk, M.H.; Guengerich, F.P. Mammalian cytochrome P450 enzymes catalyze the phenol-coupling step in endogenous morphine biosynthesis. J. Biol. Chem 2009, 284, 24425–24431. [Google Scholar]
- Belin, P.; Le Du, M.H.; Fielding, A.; Lequin, O.; Jacquet, M.; Charbonnier, J.-B.; Lecoq, A.; Thai, R.; Courcon, M.; Masson, C.; et al. Identification and structural basis of the reaction catalyzed by CYP121, an essential cytochrome P450 in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2009, 106, 7426–7431. [Google Scholar]
- Nelson, D.; Werck-Reichhart, D. A P450-centric view of plant evolution. Plant J 2011, 66, 194–211. [Google Scholar]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Daffodils as potential crops of galanthamine. Assessment of more than 100 ornamental varieties for their alkaloid content and acetylcholinesterase inhibitory activity. Ind. Crop. Prod 2013, 43, 237–244. [Google Scholar]
- Wildman, W.C.; Bailey, D.T. Amaryllidaceae interconversions. Partial syntheses of [2]benzopyrano[3,4,-c]indoles. J. Am. Chem. Soc 1969, 91, 150–157. [Google Scholar]
- Goossens, A.; Rischer, H. Implementation of functional genomics for gene discovery in alkaloid producing plants. Phytochem. Rev 2007, 6, 35–49. [Google Scholar]
- Jennewein, S.; Wildung, M.R.; Chau, M.; Walker, K.; Croteau, R. Random sequencing of an induced Taxus cell cDNA library for identification of clones involved in taxol biosynthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 9149–9154. [Google Scholar]
- Desgagné-Penix, I.; Farrow, S.C.; Cram, D.; Nowak, J.; Facchini, P.J. Integration of deep transcript and targeted metabolite profiles for eight cultivars of opium poppy. Plant Mol. Biol 2012, 79, 295–313. [Google Scholar]
- Wang, R.; Xu, S.; Jiang, Y.; Jiang, J.; Li, X.; Liang, L.; He, J.; Peng, F.; Xia, B. De novo sequence assembly and characterization of Lycoris aurea transcriptome using GS FLX titanium platform of 454 pyrosequencing. PLoS One 2013, 8, e60449. [Google Scholar]
- Ziegler, J.; Voigtländer, S.; Schmidt, J.; Kramell, R.; Miersch, O.; Ammer, C.; Gesell, A.; Kutchan, T.M. Comparative transcript and alkaloid profiling in Papaver species identifies a short chain dehydrogenase/reductase involved in morphine biosynthesis. Plant J 2006, 48, 177–192. [Google Scholar]
- Winzer, T.; Gazda, V.; He, Z.; Kaminski, F.; Kern, M.; Larson, T.R.; Li, Y.; Meade, F.; Teodor, R.; Vaistij, F.E.; et al. A Papaver somniferum 10-gene cluster for synthesis of the anticancer alkaloid noscapine. Science 2012, 336, 1704–1708. [Google Scholar]
- Chu, H.Y.; Wegel, E.; Osbourn, A. From hormones to secondary metabolism: The emergence of metabolic gene clusters in plants. Plant J 2011, 66, 66–79. [Google Scholar]
- Takos, A.M.; Rook, F. Why biosynthetic genes for chemical defense compounds cluster. Trends Plant Sci 2012, 17, 383–388. [Google Scholar]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol 2005, 3, 937–947. [Google Scholar]
- Zonneveld, B.J.M. The systematic value of nuclear DNA content for all species of Narcissus L. (Amaryllidaceae). Plant Syst. Evol 2008, 275, 109–132. [Google Scholar]
- Marienhagen, J.; Bott, M. Metabolic engineering of microorganisms for the synthesis of plant natural products. J. Biotechnol 2013, 163, 166–178. [Google Scholar]
- Keasling, J.D. Manufacturing molecules through metabolic engineering. Science 2010, 330, 1355–1358. [Google Scholar]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. Innovation for sustainable growth: A bioeconomy for Europe. Available online: http://ec.europa.eu/research/bioeconomy/pdf/201202_innovating_sustainable_growth.pdf (accessed on 9 March 2013).
- The White House. National Bioeconomy Blueprint. Available online: http://www.whitehouse.gov/sites/default/files/microsites/ostp/national_bioeconomy_blueprint_april_2012.pdf (accessed on 9 March 2013).
- Berkov, S.; Pavlov, A.; Georgiev, V.; Weber, J.; Bley, T.; Viladomat, F.; Bastida, J.; Codina, C. Changes in apolar metabolites during in vivo organogenesis of Pancratium maritimum. Plant Physiol. Biochem 2010, 48, 827–835. [Google Scholar]
- Ro, D.-K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engigeered yeast. Nature 2006, 440, 940–943. [Google Scholar]
- Levesque, F.; Seeberger, P.H. Continuous-flow synthesis of the anti-malaria drug artemisinin. Angew. Chem. Int. Ed 2012, 51, 1706–1709. [Google Scholar]
- Paddon, C.J.; Westfall, P.J.; Pitera, D.J.; Benjamin, K.; Fisher, K.; McPhee, D.; Leavell, M.D.; Tai, A.; Main, A.; Eng, D.; et al. High-level semi-synthetic production of the potent antimalarial artemisinin. Nature 2013, 496, 528–536. [Google Scholar]
- De Lorenzo, V. Towards a global biotechnology scenario. Curr. Opin. Biotechnol 2012, 23, 827–829. [Google Scholar]
- Hagel, J.M.; Krizevski, R.; Marsolais, F.; Lewinsohn, E.; Facchini, P.J. Biosynthesis of amphetamine analogs in plants. Trends Plant Sci 2012, 17, 404–412. [Google Scholar]
- Hawkins, K.M.; Smolke, C.D. Production of benzylisoquinoline alkaloids in Saccharomyces cerevisiae. Nat. Chem. Biol 2008, 4, 564–573. [Google Scholar]
- Tattersall, D.B.; Bak, S.; Jones, P.R.; Olsen, C.E.; Nielsen, J.K.; Hansen, M.L.; Høj, P.B.; Møller, B.L. Resistance to an herbivore through engineered cyanogenic glucoside synthesis. Science 2001, 293, 1826–1828. [Google Scholar]
- Mikkelsen, M.D.; Olsen, C.E.; Halkier, B.A. Production of the cancer-preventive glucoraphanin in tobacco. Mol. Plant 2010, 3, 751–759. [Google Scholar]
- Nakagawa, A.; Minami, H.; Kim, J.-S.; Koyanagi, T.; Katayama, T.; Sato, F.; Kumagai, H. A bacterial platform for fermentative production of plant alkaloids. Nat. Commun 2011, 2, 326. [Google Scholar]
- Dai, Z.; Liu, Y.; Huang, L.; Zhang, X. Production of miltiradiene by metabolically engineered Saccharomyces cerevisiae. Biotechnol. Bioeng 2012, 109, 2845–2853. [Google Scholar]
- Van der Heijden, R.; Jacobs, D.I.; Snoeijer, W.; Hallard, D.; Verpoorte, R. The Catharanthus alkaloids: Pharmacognosy and biotechnology. Curr. Med. Chem 2004, 11, 607–628. [Google Scholar]
- Allen, R.S.; Millgate, A.G.; Chitty, J.A.; Thisleton, J.; Miller, J.A.C.; Fist, A.J.; Gerlach, W.L.; Larkin, P.J. RNAi-mediated replacement of morphine with the nonnarcotic alkaloid reticuline in opium poppy. Nat. Biotechnol 2004, 22, 1559–1566. [Google Scholar]
- Frick, S.; Kramell, R.; Kutchan, T.M. Metabolic engineering with a morphine biosynthetic P450 in opium poppy surpasses breeding. Metab. Eng 2007, 9, 169–176. [Google Scholar]
- GlaxoSmithKline. Available online: http://www.gsk.com.au/about-us_poppy-production.aspx (accessed on 20 April 2013).
- Kreh, M. Studies on Galanthamine Extraction from Narcissus and Other Amaryllidaceae. In Narcissus and Daffodil. The Genus Narcissus; Hanks, G.R., Ed.; Taylor & Francis: London, UK, 2002; pp. 256–272. [Google Scholar]
- Brandham, P.E.; West, J.P. Correlation between nuclear DNA values and differing optimal ploidy levels in Narcissus, Hyacinthus and Tulipa cultivars. Genetica 1993, 90, 1–8. [Google Scholar]
- Bloembollenkeuringsdienst (BKD). Voorlopige statistiek voorjaarsbloeiers 2012–2013. Available online: http://www.bkd.eu/index.php/over-de-bkd/statistieken/voorjaarsbloeiende-bolgewassen (accessed on 6 April 2013).
- Howes, M.-J.R.; Houghton, P.J. Plants used in Chinese and Indian traditional medicine for improvement of memory and cognitive function. Pharmacol. Biochem. Behav 2003, 75, 513–527. [Google Scholar]
- Zhejiang Yixin Pharmaceutical Co., Ltd. Available online: http://www.herb-tech.com/index.asp (accessed on 20 February 2013).
- Jiang, Y.; Xia, N.; Li, X.; Shen, W.; Liang, L.; Wang, C.; Wang, R.; Peng, F.; Xia, B. Molecular cloning and characterization of a phenylalanine ammonia-lyase gene (LrPAL) from Lycoris radiate. Mol. Biol. Rep 2011, 38, 1935–1940. [Google Scholar]
- Ortiz, J.E.; Berkov, S.; Pigni, N.B.; Theoduloz, C.; Roitman, G.; Tapia, A.; Bastida, J.; Feresin, G.E. Wild Argentinian Amaryllidaceae, a new renewable source of the acetylcholinesterase inhibitor galanthamine and other alkaloids. Molecules 2012, 17, 13473–13482. [Google Scholar]
- Schumann, A.; Berkov, S.; Claus, D.; Gerth, A.; Bastida, J.; Codina, C. Production of galanthamine by Leucojum aestivum shoots grown in different bioreactor systems. Appl. Biochem. Biotechnol. 2012, 167, 1907–1920. [Google Scholar]
- Georgiev, V.; Ivanov, I.; Berkov, S.; Ilieva, M.; Georgiev, M.; Gocheva, T.; Pavlov, A. Galanthamine production by Leucojum aestivum L. shoot culture in a modified bubble column bioreactor with internal sections. Eng. Life Sci 2012, 12, 534–543. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Takos, A.M.; Rook, F. Towards a Molecular Understanding of the Biosynthesis of Amaryllidaceae Alkaloids in Support of Their Expanding Medical Use. Int. J. Mol. Sci. 2013, 14, 11713-11741. https://doi.org/10.3390/ijms140611713
Takos AM, Rook F. Towards a Molecular Understanding of the Biosynthesis of Amaryllidaceae Alkaloids in Support of Their Expanding Medical Use. International Journal of Molecular Sciences. 2013; 14(6):11713-11741. https://doi.org/10.3390/ijms140611713
Chicago/Turabian StyleTakos, Adam M., and Fred Rook. 2013. "Towards a Molecular Understanding of the Biosynthesis of Amaryllidaceae Alkaloids in Support of Their Expanding Medical Use" International Journal of Molecular Sciences 14, no. 6: 11713-11741. https://doi.org/10.3390/ijms140611713
APA StyleTakos, A. M., & Rook, F. (2013). Towards a Molecular Understanding of the Biosynthesis of Amaryllidaceae Alkaloids in Support of Their Expanding Medical Use. International Journal of Molecular Sciences, 14(6), 11713-11741. https://doi.org/10.3390/ijms140611713