Activation of Defense Mechanisms against Pathogens in Mosses and Flowering Plants

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Broad Host Range Pathogens Infect both Mosses and Flowering Plants
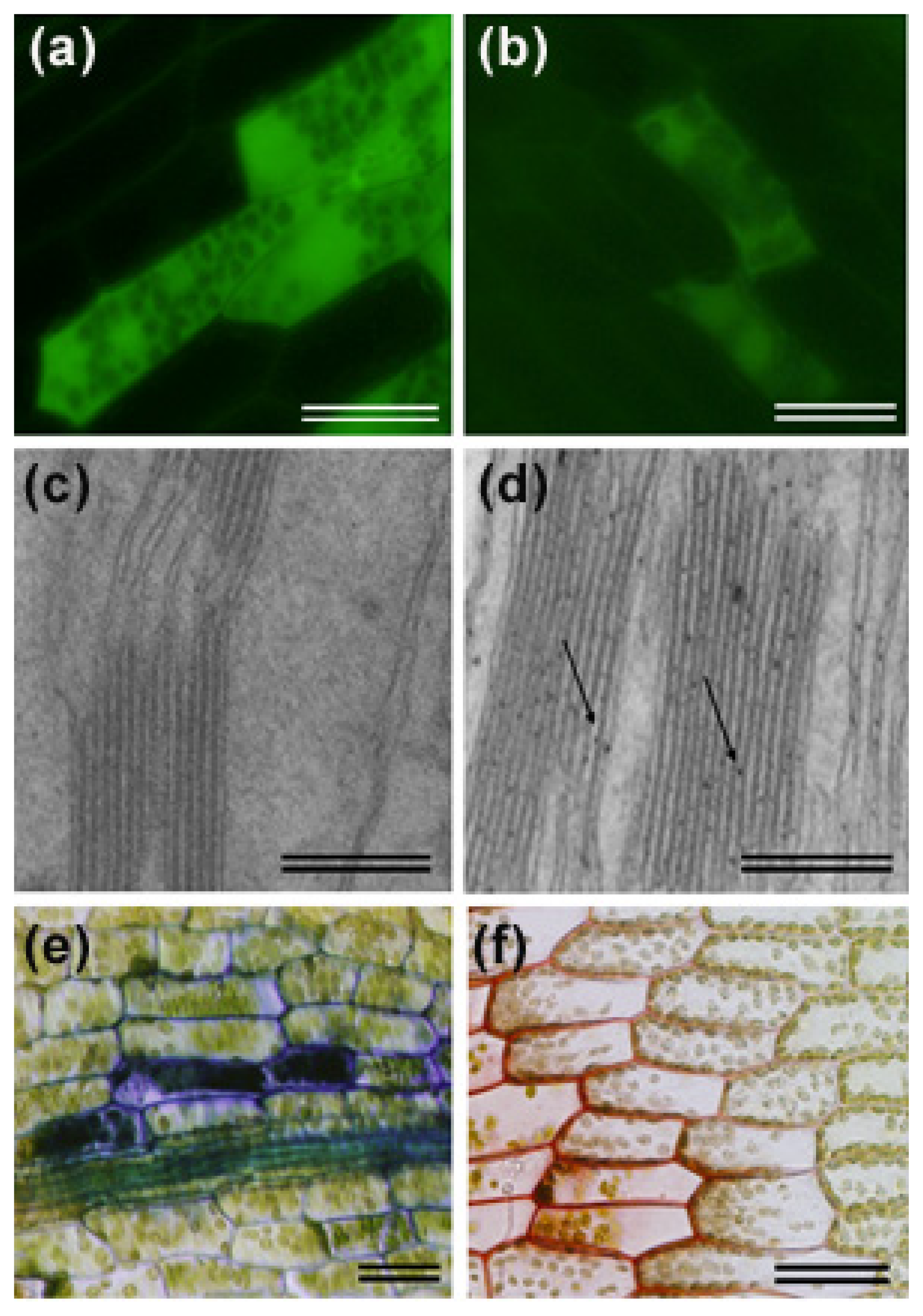
3. Activation of Cell Wall Associated Defense Responses
4. ROS Accumulation and Programmed Cell Death in Pathogen-Infected and Elicitor-Treated Plant Tissues
5. Induced Expression of Defense-Related Genes and Synthesis of Metabolites
6. Defense Hormones
7. Conclusions
Acknowledgments
Conflict of Interest
References
- Ausubel, F.M. Are innate immune signaling pathways in plants and animals conserved? Nat. Immunol 2005, 6, 973–979. [Google Scholar]
- Bittel, P.; Robatzek, S. Microbe-associated molecular patterns (MAMPs) probe plant immunity. Curr. Opin. Plant Biol 2007, 10, 335–341. [Google Scholar]
- Boller, T.; Felix, G. A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol 2009, 60, 379–406. [Google Scholar]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar]
- Abramovitch, R.B.; Janjusevic, R.; Stebbins, C.E.; Martin, G.B. Type III effector AvrPtoB requires intrinsic E3 ubiquitin ligase activity to suppress plant cell death and immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 2851–2856. [Google Scholar]
- Boller, T.; He, S.Y. Innate immunity in plants: An arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 2009, 324, 742–744. [Google Scholar]
- Cui, H.; Wang, Y.; Xue, L.; Chu, J.; Yan, C.; Fu, J.; Chen, M.; Innes, R.W.; Zhou, J.M. Pseudomonas syringae effector protein AvrB perturbs Arabidopsis hormone signaling by activating MAP kinase 4. Cell Host Microbe 2010, 7, 164–175. [Google Scholar]
- Grant, S.R.; Fisher, E.J.; Chang, J.H.; Mole, B.M.; Dangl, J.L. Subterfuge and manipulation: Type III effector proteins of phytopathogenic bacteria. Annu. Rev. Microbiol 2006, 60, 425–449. [Google Scholar]
- Zhou, J.M.; Chai, J.J. Plant pathogenic bacterial type III effectors subdue host responses. Curr. Opin. Microbiol 2008, 11, 179–185. [Google Scholar]
- Denoux, C.; Galletti, R.; Mammarella, N.; Gopalan, S.; Werck, D.; De Lorenzo, G.; Ferrari, S.; Ausubel, F.M.; Dewdney, J. Activation of defense response pathways by OGs and flg22 elicitors in Arabidopsis seedlings. Mol. Plant 2008, 1, 423–445. [Google Scholar]
- Lotze, M.T.; Zeh, H.J.; Rubartelli, A.; Sparvero, L.J.; Amoscato, A.A.; Washburn, N.R.; Devera, M.E.; Liang, X.; Tör, M.; Billiar, T. The grateful dead: damage associated molecular pattern molecules and reduction/oxidation regulate immunity. Immunol. Rev 2007, 220, 60–81. [Google Scholar]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol 2010, 13, 459–465. [Google Scholar]
- Thomma, BP.; Nurnberger, T.; Joosten, MH. Of PAMPs and effectors: the blurred PTI-ETI dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar]
- Nicaise, V.; Roux, M.; Zipfel, C. Recent advances in PAMP-triggered immunity against bacteria: Pattern recognition receptors watch over and raise the alarm. Plant Physiol 2009, 150, 1638–1647. [Google Scholar]
- van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol 2006, 44, 135–162. [Google Scholar]
- Bent, A.F.; Mackey, D. Elicitors, effectors, and R genes: The new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol 2007, 45, 399–436. [Google Scholar]
- Zipfel, C. Early molecular events in PAMP-triggered immunity. Curr. Opin. Plant Biol 2009, 12, 414–420. [Google Scholar]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar]
- Schaefer, D.G. A new moss genetics: targeted mutagenesis in Physcomitrella patens. Annu. Rev. Plant Biol 2002, 53, 477–501. [Google Scholar]
- Cove, D. The moss Physcomitrella patens. Annu. Rev. Genet 2005, 39, 339–358. [Google Scholar]
- Lewis, L.A.; McCourt, R.M. Green algae and the origin of land plants. Am. J. Bot 2004, 91, 1535–1556. [Google Scholar]
- Banks, J.A.; Nishiyama, T.; Hasebe, M.; Bowman, J.L.; Gribskov, M.; dePamphilis, C.; Albert, V.A.; Aono, N.; Aoyama, T.; Ambrose, B.A.; et al. The Selaginella genome identifies genetic changes associated with the evolution of vascular plants. Science 2011, 332, 960–963. [Google Scholar]
- Döbbeler, P. Biodiversity of bryophilous ascomycetes. Biodivers. Conserv 1997, 6, 721–738. [Google Scholar]
- Davey, M.L.; Currah, R.S. Interactions between mosses (Bryophyta) and fungi. Can. J. Bot 2006, 84, 1509–1519. [Google Scholar]
- Davey, M.L.; Tsuneda, A.; Currah, R.S. Pathogenesis of bryophyte hosts by the ascomycete Atradidymella muscivora. Am. J. Bot 2009, 96, 1274–1280. [Google Scholar]
- Ponce de León, I.; Oliver, J.P.; Castro, A.; Gaggero, C.; Bentancor, M.; Vidal, S. Erwinia carotovora elicitors and Botrytis cinerea activate defense responses in Physcomitrella patens. BMC Plant Biol 2007, 7, 52. [Google Scholar]
- Ponce de León, I. The moss Physcomitrella patens as a model system to study interactions between plants and phytopathogenic fungi and oomycetes. J. Pathog. 2011. [Google Scholar] [CrossRef]
- Ponce De León, I.; Schmelz, E.A.; Gaggero, C.; Castro, A.; Álvarez, A.; Montesano, M. Physcomitrella patens activates reinforcement of the cell wall, programmed cell death and accumulation of evolutionary conserved defence signals, such as salicylic acid and 12-oxo-phytodienoic acid, but not jasmonic acid, upon Botrytis cinerea infection. Mol. Plant Pathol 2012, 13, 960–974. [Google Scholar]
- Oliver, J.P.; Castro, A.; Gaggero, C.; Cascón, T.; Schmelz, E.A.; Castresana, C.; Ponce de León, I. Pythium infection activates conserved plant defense responses in mosses. Planta 2009, 230, 569–579. [Google Scholar]
- Lehtonen, M.T.; Akita, M.; Kalkkinen, N.; Ahola-Iivarinen, E.; Rönnholm, G.; Somervuo, P.; Thelander, M.; Valkonen, J.P. Quickly-released peroxidase of moss in defense against fungal invaders. New Phytol 2009, 183, 432–443. [Google Scholar]
- Lawton, M.; Saidasan, H. Pathogenesis in mosses. Annu. Plant Rev 2009, 36, 298–339. [Google Scholar]
- Lehtonen, M.T.; Akita, M.; Frank, W.; Reski, R.; Valkonen, J.P. Involvement of a class III peroxidase and the mitochondrial protein TSPO in oxidative burst upon treatment of moss plants with a fungal elicitor. Mol. Plant-Microbe Interact 2012, 25, 363–371. [Google Scholar]
- Schwessinger, B.; Ronald, P.C. Plant innate immunity: Perception of conserved microbial signatures. Annu. Rev. Plant Biol 2012, 63, 451–482. [Google Scholar]
- Akita, M.; Valkonen, J.P.T. A novel gene family in moss (Physcomitrella patens) shows sequence homology and a phylogenetic relationship with the TIR-NBS class of plant disease resistance genes. J. Mol. Evol 2002, 55, 595–605. [Google Scholar]
- Xue, J.Y.; Wang, Y.; Wu, P.; Wang, Q.; Yang, L.T.; Pan, X.H.; Wang, B.; Chen, J.Q. A primary survey on bryophyte species reveals two novel classes of nucleotide-binding site (NBS) genes. PLoS One 2012, 7, e36700. [Google Scholar]
- Stone, J.K. Necrotroph. In Encyclopedia of Plant Pathology; Maloy, O.C., Murray, T.D., Eds.; Wiley: New York, NY, USA, 2001; Volume 2, pp. 676–677. [Google Scholar]
- Elad, Y.; Williamson, B.; Tudzynski, P.; Delen, N. Botrytis: Biology, Pathology and Control; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004. [Google Scholar]
- van Kan, J.A. Licensed to kill: The lifestyle of a necrotrophic plant pathogen. Trends Plant Sci 2006, 11, 247–253. [Google Scholar]
- Prins, T.W.; Tudzynski, P.; Tiedemann, A.V.; Tudzynski, B.; ten Have, A.; Hansen, M.E.; Tenberge, K.; van Kan, J.A.L. Infection strategies of Botrytis cinerea and related necrotrophic pathogens. In Fungal Pathology; Kronstad, J.W., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 33–64. [Google Scholar]
- Staats, M.; van Baarlen, P.; van Kan, J.A. Molecular phylogeny of the plant pathogenic genus Botrytis and the evolution of host specificity. Mol. Biol. Evol 2005, 22, 333–346. [Google Scholar]
- Perombelon, M.C.M.; Kelman, A. Ecology of the Soft Rot Erwinias. Annu Rev. Phytopathol 1980, 18, 361–387. [Google Scholar]
- Toth, I.K.; Birch, P.R. Rotting softly and stealthily. Curr. Opin. Plant Biol 2005, 8, 424–429. [Google Scholar]
- Palva, T.K.; Holmström, K.O.; Heino, P.; Palva, E.T. Induction of plant defense response by exoenzymes of Erwinia carotovora ssp. carotovora. Mol. Plant-Microbe Interact 1993, 6, 190–196. [Google Scholar]
- Norman, C.; Vidal, S.; Palva, E.T. Oligogalacturonide-mediated induction of a gene involved in jasmonic acid synthesis in response to the cell-wall-degrading enzymes of the plant pathogen Erwinia carotovora. Mol. Plant-Microbe Interact 1999, 12, 640–644. [Google Scholar]
- Norman-Setterblad, C.; Vidal, S.; Palva, E.T. Interacting signal pathways control defense gene expression in Arabidopsis in response to cell wall-degrading enzymes from Erwinia carotovora. Mol. Plant-Microbe Interact 2000, 13, 430–438. [Google Scholar]
- Vidal, S.; Ponce de León, I.; Denecke, J.; Palva, E.T. Salicylic acid and the plant pathogen Erwinia carotovora induce defense genes via antagonistic pathways. Plant J 1997, 11, 115–123. [Google Scholar]
- Vidal, S.; Eriksson, A.R.B.; Montesano, M.; Denecke, J.; Palva, E.T. Cell wall degrading enzymes from Erwinia carotovora cooperate in the salicylic acid-independent induction of a plant defense response. Mol. Plant-Microbe Interact. 1998, 11, 23–32. [Google Scholar]
- Montesano, M.; Brader, G.; Ponce de Leon, I.; Palva, ET. Multiple defense signals induced by Erwinia carotovora ssp. carotovora in potato. Mol. Plant Pathol. 2005, 6, 541–549. [Google Scholar]
- Montesano, M.; Kõiv, V.; Mãe, A.; Palva, E.T. Novel receptor-like protein kinases induced by Erwinia carotovora and short oligogalacturonides in potato. Mol. Plant Pathol 2001, 2, 339–346. [Google Scholar]
- Davis, K.R.; Lyon, G.D.; Darvill, A.G.; Albersheim, P. Host-pathogen interactions: XXV. Endopolygalacturonic acid lyase from Erwinia carotovora elicits phytoalexin accumulation by releasing plant cell wall fragments. Plant Physiol 1984, 74, 52–60. [Google Scholar]
- Nothnagel, E.A.; McNeil, M.; Albersheim, P.; Dell, A. Host-pathogen interactions: XXII. A galacturonic acid oligosaccharide from plant cell walls elicits phytoalexins. Plant Physiol 1983, 71, 916–926. [Google Scholar]
- Rantakari, A.; Virtaharju, O.; Vähämiko, S.; Taira, S.; Palva, E.T.; Saarilahti, H.T.; Romantschuk, M. Type III secretion contributes to the pathogenesis of the soft-rot pathogen Erwinia carotovora partial characterization of the hrp gene cluster. Mol. Plant-Microbe Interact 2001, 14, 962–968. [Google Scholar]
- Mattinen, L.; Tshuikina, M.; Mäe, A.; Pirhonen, M. Identification and characterization of Nip, necrosis-inducing virulence protein of Erwinia carotovora subsp. carotovora. Mol. Plant-Microbe Interact 2004, 17, 1366–1375. [Google Scholar]
- Martin, F. Pythium. In Pathogenesis and Host Specificity in Plant Diseases: Histopathological, Biochemical, Genetic and Molecular Bases; Komoto, K., Singh, U.S., Singh, R.P., Eds.; Pergamon Press: Oxford, UK, 1994; pp. 17–36. [Google Scholar]
- Campion, C.; Massiot, P.; Rouxel, F. Aggressiveness and production of cell-wall degrading enzymes by Pythium violae, Pythium sulcatum and Pythium ultimum, responsible for cavity spot on carrots. Eur. J. Plant Pathol 1997, 103, 725–735. [Google Scholar]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. Annu. Rev. Phytopathol 1996, 34, 367–386. [Google Scholar]
- Asselbergh, B.; Curvers, K.; Franca, S.C.; Audenaert, K.; Vuylsteke, M.; Van Breusegem, F.; Höfte, M. Resistance to Botrytis cinerea in sitiens, an abscisic acid-deficient tomato mutant, involves timely production of hydrogen peroxide and cell wall modifications in the epidermis. Plant Physiol 2007, 144, 1863–1877. [Google Scholar]
- Curvers, K.; Seifi, H.; Mouille, G.; de Rycke, R.; Asselbergh, B.; Van Hecke, A.; Vanderschaeghe, D.; Höfte, H.; Callewaert, N.; van Breusegem, F.; Höfte, M. Abscisic acid deficiency causes changes in cuticle permeability and pectin composition that influence tomato resistance to Botrytis cinerea. Plant Physiol 2010, 154, 847–860. [Google Scholar]
- Jacobs, A.K.; Lipka, V.; Burton, R.A.; Panstruga, R.; Strizhov, N.; Schulze-Lefert, P.; Fincher, G.B. An Arabidopsis callose synthase, GSL5, is required for wound and papillary callose formation. Plant Cell 2003, 15, 2503–2513. [Google Scholar]
- Ton, J.; Mauch-Mani, B. Beta-amino-butyric acid-induced resistance against necrotrophic pathogens is based on ABA-dependent priming for callose. Plant J 2004, 38, 119–130. [Google Scholar]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose deposition: A multifaceted plant defense response. Mol. Plant-Microbe Interact 2011, 24, 183–193. [Google Scholar]
- Adie, B.A.; Pérez-Pérez, J.; Pérez-Pérez, M.M.; Godoy, M.; Sánchez-Serrano, J.J.; Schmelz, E.A.; Solano, R. ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defenses in Arabidopsis. Plant Cell 2007, 19, 1665–1681. [Google Scholar]
- Benhamou, N.; Bélanger, R. Induction of systemic resistance to Pythium damping-off in cucumber plants by benzothiadiazole: ultrastructure and cytochemistry of the host response. Plant J 1998, 14, 13–21. [Google Scholar]
- Ramírez, V.; Agorio, A.; Coego, A.; García-Andrade, J.; Hernández, M.J.; Balaguer, B.; Ouwerkerk, P.B.; Zarra, I.; Vera, P. MYB46 modulates disease susceptibility to Botrytis cinerea in Arabidopsis. Plant Physiol 2011, 155, 1920–1935. [Google Scholar]
- Davin, L.B.; Lewis, N.G. Dirigent proteins and dirigent sites explain the mystery of specificity of radical precursor coupling in lignan and lignin biosynthesis. Plant Physiol 2000, 123, 453–462. [Google Scholar]
- Coram, T.E.; Wang, M.; Chen, X. Transcriptome analysis of the wheat–Puccinia striiformis f. sp. tritici interaction. Mol. Plant Pathol 2008, 9, 157–169. [Google Scholar]
- Chakravarthy, S.; Velásquez, A.C.; Ekengren, S.K.; Collmer, A.; Martin, G.B. Identification of Nicotiana benthamiana genes involved in pathogen associated molecular pattern-triggered immunity. Mol. Plant-Microbe Interact 2010, 23, 715–726. [Google Scholar]
- Ferrari, S.; Gallettim, R.; Denoux, C.; De Lorenzo, G.; Ausubel, F.M.; Dewdney, J. Resistance to Botrytis cinerea induced in Arabidopsis by elicitors is independent of salicylic acid, ethylene or jasmonate signaling but requires PAD3. Plant Physiol 2007, 144, 367–379. [Google Scholar]
- Xu, Z.; Zhang, D.; Hu, J.; Zhou, X.; Ye, X.; Reichel, K.L.; Stewart, N.R.; Syrenne, R.D.; Yang, X.; et al. Comparative genome analysis of lignin biosynthesis gene families across the plant kingdom. BMC Bioinformatics 2009, 10, S3. [Google Scholar]
- Popper, Z.A. Evolution and diversity of green plant cell walls. Curr. Opin. Plant Biol 2008, 11, 286–292. [Google Scholar]
- Weng, J.K.; Chapple, C. The origin and evolution of lignin biosynthesis. New Phytol 2010, 187, 273–285. [Google Scholar]
- Lloyd, A.J.; William Allwood, J.; Winder, C.L.; Dunn, W.B.; Heald, J.K.; Cristescu, S.M.; Sivakumaran, A.; Harren, F.J.; Mulema, J.; Denby, K.; et al. Metabolomic approaches reveal that cell wall modifications play a major role in ethylene-mediated resistance against. Botrytis cinerea. Plant J. 2011, 67, 852–868. [Google Scholar]
- Torres, M.A.; Jones, J.D.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol 2006, 141, 373–378. [Google Scholar]
- Tiedemann, A.V. Evidence for a primary role of active oxygen species in induction of host cell death during infection of bean leaves with Botrytis cinerea. Physiol. Mol. Plant Pathol 1997, 50, 151–166. [Google Scholar]
- Schouten, A.; Tenberge, K.B.; Vermeer, J.; Stewart, J.; Wagemakers, L.; Williamson, B.; van Kan, J.A. Functional analysis of an extracellular catalase of Botrytis cinerea. Mol. Plant Pathol 2002, 3, 227–238. [Google Scholar]
- Choquer, M.; Fournier, E.; Kunz, C.; Levis, C.; Pradier, J.M.; Simon, A.; Viaud, M. Botrytis cinerea virulence factors: New insights into a necrotrophic and polyphageous pathogen. FEMS Microbiol. Lett 2007, 277, 1–10. [Google Scholar]
- Rolke, Y.; Liu, S.; Quidde, T.; Williamson, B.; Schouten, A.; Weltring, K.M.; Siewers, V.; Tenberge, K.B.; Tudzynski, B.; Tudzynski, P. Functional analysis of H2O2-generating systems in Botrytis cinerea: The major Cu-Zn-superoxide dismutase (BCSOD1) contributes to virulence on French bean, whereas a glucose oxidase (BCGOD1) is dispensable. Mol. Plant Pathol. 2004, 5, 17–27. [Google Scholar]
- Govrin, E.M.; Levine, A. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr. Biol 2000, 10, 751–757. [Google Scholar]
- Asai, S.; Yoshioka, H. Nitric oxide as a partner of reactive oxygen species participates in disease resistance to necrotrophic pathogen Botrytis cinerea in Nicotiana benthamiana. Mol. Plant-Microbe Interact 2009, 22, 619–629. [Google Scholar]
- Montesano, M.; Scheller, H.V.; Wettstein, R.; Palva, E.T. Down-regulation of photosystem I by Erwinia carotovora-derived elicitors correlates with H2O2 accumulation in chloroplasts of potato. Mol. Plant Pathol 2004, 5, 115–123. [Google Scholar]
- Ghosh, M. Antifungal properties of haem peroxidase from Acorus calamus. Ann. Bot 2006, 98, 1145–1153. [Google Scholar]
- Hemetsberger, C.; Herrberger, C.; Zechmann, B.; Hillmer, M.; Doehlemann, G. The Ustilago maydis effector Pep1 suppresses plant immunity by inhibition of host peroxidase activity. PLoS Pathog 2012, 8, e1002684. [Google Scholar]
- Frank, W.; Baar, K.; Qudeimat, E.; Woriedh, M.; Alawady, A.; Ratnadewi, D.; Gremillon, L.; Grimm, B.; Reski, R. A mitochondrial protein homologous to the mammalian peripheral-type benzodiazepine receptor is essential for stress adaptation in plants. Plant J 2007, 51, 1004–1018. [Google Scholar]
- van Doorn, W.G.; Beers, E.P.; Dangl, J.L.; Franklin-Tong, V.E.; Gallois, P.; Hara-Nishimura, I.; Jones, A.M.; Kawai-Yamada, M.; Lam, E.; Mundy, J.; et al. Morphological classification of plant cell deaths. Cell Death Differ. 2011, 18, 1241–1246. [Google Scholar]
- Kjemtrup, S.; Nimchuk, Z.; Dangl, J.L. Effector proteins of phytopathogenic bacteria: Bifunctional signals in virulence and host recognition. Curr. Opin. Microbiol 2000, 3, 73–78. [Google Scholar]
- Kachroo, P.; Shanklin, J.; Shah, J.; Whittle, E.J.; Klessig, D.F. A fatty acid desaturase modulates the activation of defense signaling pathways in plants. Proc. Natl. Acad. Sci. USA 2001, 98, 9448–9453. [Google Scholar]
- Veronese, P.; Chen, X.; Bluhm, B.; Salmeron, J.; Dietrich, R.; Mengiste, T. The BOS loci of Arabidopsis are required for resistance to Botrytis cinerea infection. Plant J 2004, 40, 558–574. [Google Scholar]
- Dickman, M.B.; Park, Y.K.; Oltersdorf, T.; Li, W.; Clemente, T.; French, R. Abrogation of disease development in plants expressing animal antiapoptotic genes. Proc. Natl. Acad. Sci. USA 2001, 98, 6957–6962. [Google Scholar]
- van Baarlen, P.; Woltering, E.J.; Staats, M.; van Kan, J.A.L. Histochemical and genetic analysis of host and non-host interactions of Arabidopsis with three Botrytis species: an important role for cell death control. Mol. Plant Pathol 2007, 8, 41–54. [Google Scholar]
- Wei, Z.M.; Laby, R.J.; Zumoff, C.H.; Bauer, D.W.; He, S.Y.; Collmer, A.; Beer, S.V. Harpin, elicitor of the hypersensitive response produced by the plant pathogen Erwinia amylovora. Science 1992, 257, 85–88. [Google Scholar]
- Kariola, T.; Palomäki, T.A.; Brader, G.; Palva, E.T. Erwinia carotovora subsp. carotovora and Erwinia -derived elicitors HrpN and PehA trigger distinct but interacting defense responses and cell death in Arabidopsis. Mol. Plant-Microbe Interact 2003, 16, 179–187. [Google Scholar]
- Kim, J.G.; Jeon, E.; Oh, J.; Moon, J.S.; Hwang, I. Mutational analysis of Xanthomonas harpin HpaG identifies a key functional region that elicits the hypersensitive response in nonhost plants. J. Bacteriol 2004, 186, 6239–6247. [Google Scholar]
- Alfano, J.R.; Bauer, D.W.; Milos, T.M.; Collmer, A. Analysis of the role of the Pseudomonas syringae pv. syringae HrpZ harpin in elicitation of the hypersensitive response in tobacco using functionally non-polar hrpZ deletion mutations, truncated HrpZ fragments, and hrmA mutations. Mol. Microbiol 1996, 19, 715–728. [Google Scholar]
- Zhu, Q.; Dröge-Laser, W.; Dixon, R.A.; Lamb, C. Transcriptional activation of plant defense genes. Curr. Opin. Genet. Dev 1996, 6, 624–630. [Google Scholar]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar]
- Windram, O.; Madhou, P.; McHattie, S.; Hill, C.; Hickman, R.; Cooke, E.; Jenkins, D.J.; Penfold, C.A.; Baxter, L.; Breeze, E.; et al. Arabidopsis Defense against Botrytis cinerea: Chronology and Regulation Deciphered by High-Resolution Temporal Transcriptomic Analysis. Plant Cell 2012, 24, 3530–3557. [Google Scholar]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol 2002, 53, 275–297. [Google Scholar]
- Rensing, S.A.; Ick, J.; Fawcett, J.A.; Lang, D.; Zimmer, A.; Van de Peer, Y.; Reski, R. An ancient genome duplication contributed to the abundance of metabolic genes in the moss Physcomitrella patens. BMC Evol. Biol 2007, 7, 130. [Google Scholar]
- Wolf, L.; Rizzini, L.; Stracke, R.; Ulm, R.; Rensing, S.A. The molecular and physiological responses of Physcomitrella patens to ultraviolet-B radiation. Plant Physiol 2010, 153, 1123–1134. [Google Scholar]
- Koduri, P.K.; Gordon, G.S.; Barker, E.I.; Colpitts, C.C.; Ashton, N.W.; Suh, D.Y. Genome-wide analysis of the chalcone synthase superfamily genes of Physcomitrella patens. Plant Mol. Biol 2010, 72, 247–263. [Google Scholar]
- Jia, Z.; Zou, B.; Wang, X.; Qiu, J.; Ma, H.; Gou, Z.; Song, S.; Dong, H. Quercetin-induced H(2)O(2) mediates the pathogen resistance against Pseudomonas syringae pv. Tomato DC3000 in Arabidopsis thaliana. Biochem. Biophys. Res. Commun 2010, 28, 522–527. [Google Scholar]
- Wichard, T.; Göbel, C.; Feussner, I.; Pohnert, V. Unprecedented lipoxygenase/hydroperoxide lyase pathways in the moss Physcomitrella patens. Angew. Chem. Int. Edit 2004, 44, 158–161. [Google Scholar]
- Senger, T.; Wichard, T.; Kunze, S.; Gobel, C.; Lerchl, J.; Pohnert, G.; Feussner, I. A multifunctional lipoxygenase with fatty acid hydroperoxide cleaving activity from the moss Physcomitrella patens. J. Biol. Chem 2005, 280, 7588–7596. [Google Scholar]
- Anterola, A.; Göbel, C.; Hornung, E.; Sellhorn, G.; Feussner, I.; Grimes, H. Physcomitrella patens has lipoxygenases for both eicosanoid and octadecanoid pathways. Phytochemistry 2009, 70, 40–52. [Google Scholar]
- Stumpe, M.; Bode, J.; Gobel, C.; Wichard, T.; Schaaf, A.; Frank, W.; Frank, M.; Reski, R.; Pohnert, G.; Feussner, I. Biosynthesis of C9-aldehydes in the moss Physcomitrella patens. Biochim. Biophys. Acta 2006, 1761, 301–312. [Google Scholar]
- Prost, I.; Dhondt, S.; Rothe, G.; Vicente, J.; Rodriguez, M.J.; Kift, N.; Carbonne, F.; Griffiths, G.; Esquerré-Tugayé, M.T.; Rosahl, S.; et al. Evaluation of the antimicrobial activities of plant oxylipins supports their involvement in defense against pathogens. Plant Physiol. 2005, 139, 1902–1913. [Google Scholar]
- Von Schwartzenberg, K.; Schultze, W.; Kassner, H. The moss Physcomitrella patens releases a tetracyclic diterpene. Plant Cell Rep 2004, 22, 780–786. [Google Scholar]
- Peters, R.J. Uncovering the complex metabolic network underlying diterpenoid phytoalexin biosynthesis in rice and other cereal crop plants. Phytochemistry 2006, 67, 2307–2317. [Google Scholar]
- Schmelz, E.A.; Kaplan, F.; Huffaker, A.; Dafoe, N.J.; Vaughan, M.M.; Ni, X.; Rocca, J.R.; Alborn, H.T.; Teal, P.E. Identity, regulation, and activity of inducible diterpenoid phytoalexins in maize. Proc. Natl. Acad. Sci. USA 2011, 108, 5455–5460. [Google Scholar]
- López, M.A.; Bannenberg, G.; Castresana, C. Controlling hormone signaling is a plant and pathogen challenge for growth and survival. Curr. Opin. Plant Biol 2008, 11, 420–427. [Google Scholar]
- Feys, B.J.; Parker, J.E. Interplay of signaling pathways in plant disease resistance. Trends Genet 2000, 16, 449–455. [Google Scholar]
- Lund, S.T.; Stall, R.E.; Klee, H.J. Ethylene regulates the susceptible response to pathogen infection in tomato. Plant Cell 1998, 10, 371–382. [Google Scholar]
- Greenberg, J.T.; Silverman, F.P.; Liang, H. Uncoupling salicylic acid-dependent cell death and defense-related responses from disease resistance in the Arabidopsis mutant acd5. Genetics 2000, 156, 341–350. [Google Scholar]
- Pilloff, R.K.; Devadas, S.K.; Enyedi, A.; Raina, R. The Arabidopsis gain-of-function mutant Dll1 spontaneously develops lesions mimicking cell death associated with disease. Plant J 2002, 30, 61–70. [Google Scholar]
- Mur, L.A.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol 2006, 140, 249–262. [Google Scholar]
- Koornneef, A.; Pieterse, C.M. Cross talk in defense signaling. Plant Physiol 2008, 146, 839–844. [Google Scholar]
- Komatsu, K.; Nishikawa, Y.; Ohtsuka, T.; Taji, T.; Quatrano, R.S.; Tanaka, S.; Sakata, Y. Functional analyses of the ABI1-related protein phosphatase type 2C reveal evolutionarily conserved regulation of abscisic acid signaling between Arabidopsis and the moss Physcomitrella patens. Plant Mol. Biol 2009, 70, 327–340. [Google Scholar]
- Bierfreund, N.M.; Reski, R.; Decker, E.L. Use of an inducible reporter gene system for the analysis of auxin distribution in the moss Physcomitrella patens. Plant Cell Rep 2003, 21, 1143–1152. [Google Scholar]
- Schwartzenberg, K.V.; Nunez, M.F.; Blaschke, H.; Dobrev, P.I.; Novak, D.O.; Motyka, V.; Strnad, M. Cytokinins in the bryophyte Physcomitrella patens: analyses of activity, distribution, and cytokinin oxidase/dehydrogenase overexpression reveal the role of extracellular cytokinins. Plant Physiol 2007, 145, 786–800. [Google Scholar]
- Khandelwal, A.; Cho, S.H.; Marella, H.; Sakata, Y.; Perroud, P.F.; Pan, A.; Quatrano, R.S. Role of ABA and ABI3 in desiccation tolerance. Science 2010, 327, 546. [Google Scholar]
- Bhyan, S.B.; Minami, A.; Kaneko, Y.; Suzuki, S.; Arakawa, K.; Sakata, Y.; Takezawa, D. Cold acclimation in the moss Physcomitrella patens involves abscisic acid-dependent signaling. J. Plant Physiol 2012, 169, 137–145. [Google Scholar]
- Prigge, M.J.; Bezanilla, M. Evolutionary crossroads in developmental biology: Physcomitrella patens. Development 2010, 137, 3535–3543. [Google Scholar]
- Jang, G.; Dolan, L. Auxin promotes the transition from chloronema to caulonema in moss protonema by positively regulating PpRSL1and PpRSL2 in Physcomitrella patens. New Phytol 2011, 192, 319–327. [Google Scholar]
- Saleh, O.; Issman, N.; Seumel, G.I.; Stav, R.; Samach, A.; Reski, R.; Frank, W.; Arazi, T. MicroRNA534a control of BLADE-ON-PETIOLE 1 and 2 mediates juvenile-to-adult gametophyte transition in Physcomitrella patens. Plant J 2011, 65, 661–674. [Google Scholar]
- Ton, J.; Flors, V.; Mauch-Mani, B. The multifaceted role of ABA in disease resistance. Trends Plant Sci 2009, 14, 310–317. [Google Scholar]
- Mauch-Mani, B.; Mauch, F. The role of abscisic acid in plant-pathogen interactions. Curr. Opin. Plant Biol 2005, 8, 409–414. [Google Scholar]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol 2009, 69, 473–488. [Google Scholar]
- Grant, M.R.; Jones, J.D. Hormone (dis)harmony moulds plant health and disease. Science 2009, 324, 750–752. [Google Scholar]
- AbuQamar, S.; Chen, X.; Dhawan, R.; Bluhm, B.; Salmeron, J.; Lam, S.; Dietrich, R.A.; Mengiste, T. Expression profiling and mutant analysis reveals complex regulatory networks involved in Arabidopsis response to Botrytis infection. Plant J 2006, 48, 28–44. [Google Scholar]
- Siewers, V.; Smedsgaard, J.; Tudzynski, P. The P450 monooxygenase BcABA1 is essential for abscisic acid biosynthesis in Botrytis cinerea. Appl. Environ. Microbiol 2004, 70, 3868–3876. [Google Scholar]
- Audenaert, K.; De Meyer, G.B.; Höfte, M.M. Abscisic acid determines basal susceptibility of tomato to Botrytis cinerea and suppresses salicylic acid dependent signaling mechanisms. Plant Physiol 2002, 128, 491–501. [Google Scholar]
- Yasuda, M.; Ishikawa, A.; Jikumaru, Y.; Seki, M.; Umezawa, T.; Asami, T.; Maruyama-Nakashita, A.; Kudo, T.; Shinozaki, K.; Yoshida, S.; et al. Antagonistic interaction between systemic acquired resistance and the abscisic acid-mediated abiotic stress response in. Arabidopsis. Plant Cell 2008, 20, 1678–1692. [Google Scholar]
- Rohwer, F.; Bopp, M. Ethylene Synthesis in Moss Protonema. J. Plant Physiol 1985, 117, 331–338. [Google Scholar]
- Osborne, D.J.; Walters, J.; Milborrow, B.V.; Norville, A.; Stange, L.M.C. Evidence for a non-ACC ethylene biosynthesis pathway in lower plants. Phytochemistry 1995, 42, 51–60. [Google Scholar]
- Ishida, K.; Yamashino, T.; Nakanishi, H.; Mizuno, T. Classification of the genes involved in the two-component system of the moss Physcomitrella patens. Biosci. Biotechnol. Biochem 2010, 74, 2542–2545. [Google Scholar]
- Yasumura, Y.; Pierik, R.; Fricker, M.D.; Voesenek, L.A.; Harberd, N.P. Studies of Physcomitrella patens reveal that ethylene-mediated submergence responses arose relatively early in land-plant evolution. Plant J 2012, 72, 947–959. [Google Scholar]
- Díaz, J.; ten Have, A.; van Kan, J.A. The role of ethylene and wound signaling in resistance of tomato to Botrytis cinerea. Plant Physiol 2002, 129, 1341–1351. [Google Scholar]
- Penninckx, I.A.M.A.; Eggermont, K.; Terras, F.R.G.; Thomma, B.P.H.J.; De Samblanx, G.W.; Buchala, A.; Métraux, J.P.; Manners, J.M.; Broekaert, W.F. Pathogen-induced systemic activation of a plant defensin gene in Arabidopsis follows a salicylic acid-independent pathway. Plant Cell 1996, 8, 2309–2323. [Google Scholar]
- Thomma, B.P.H.J.; Eggermont, K.; Tierens, K.F.M.J.; Broekaert, W.F. Requirement of functional ethylene-insensitive 2 gene for efficient resistance of Arabidopsis to infection by Botrytis cinerea. Plant Physiol 1999, 121, 1093–1101. [Google Scholar]
- Berrocal-Lobo, M.; Molina, A.; Solano, R. Constitutive expression of ETHYLENE-RESPONSEFACTOR1 in Arabidopsis confers resistance to several necrotrophic fungi. Plant J 2002, 29, 23–32. [Google Scholar]
- Chague, V.; Elad, Y.; Barakat, R.; Tudzynski, P.; Sharon, A. Ethylene biosynthesis in Botrytis cinerea. FEMS Microbiol. Ecol 2002, 40, 143–149. [Google Scholar]
- Han, L.; Li, G.J.; Yang, K.Y.; Mao, G.; Wang, R.; Liu, Y.; Zhang, S. Mitogen-activated protein kinase 3 and 6 regulate Botrytis cinerea-induced ethylene production in Arabidopsis. Plant J 2010, 64, 114–127. [Google Scholar]
- Bandara, P.K.; Takahashi, K.; Sato, M.; Matsuura, H.; Nabeta, K. Cloning and functional analysis of an allene oxide synthase in Physcomitrella patens. Biosci. Biotechnol. Biochem 2009, 73, 2356–2359. [Google Scholar]
- Scholz, J.; Dickmanns, A.; Feussner, I.; Ficner, R. Crystal Structures of Physcomitrella patens AOC1 and AOC2: Insights into the Enzyme Mechanism and Differences in Substrate Specificity. Plant Physiol 2012, 160, 1251–1266. [Google Scholar]
- Stumpe, M.; Göbel, C.; Faltin, B.; Beike, A.K.; Hause, B.; Himmelsbach, K.; Bode, J.; Kramell, R.; Wasternack, C.; Frank, W.; et al. The moss Physcomitrella patens contains cyclopentenones but no jasmonates: mutations in allene oxide cyclase lead to reduced fertility and altered sporophyte morphology. New Phytol. 2010, 188, 740–749. [Google Scholar]
- Hashimoto, T.; Takahashi, K.; Sato, M.; Bandara, P.K.G.S.S.; Nabeta, K. Cloning and characterization of an allene oxide cyclase, PpAOC3, in Physcomitrella patens. Plant Growth Regul. 2011, 65, 239–245. [Google Scholar]
- Breithaupt, C.; Kurzbauer, R.; Schaller, F.; Stintzi, A.; Schaller, A.; Huber, R.; Macheroux, P.; Clausen, T. Structural basis of substrate specificity of plant 12-oxophytodienoate reductases. J. Mol. Biol 2009, 392, 1266–1277. [Google Scholar]
- Li, W.; Liu, B.; Yu, L.; Feng, D.; Wang, H.; Wang, J. Phylogenetic analysis, structural evolution and functional divergence of the 12-oxophytodienoate acid reductase gene family in plants. BMC Evol. Biol 2009, 9, 90. [Google Scholar]
- Vicedo, B.; Flors, V.; de la O Leyva, M.; Finiti, I.; Kravchuk, Z.; Real, M.D.; García-Agustín, P.; González-Bosch, C. Hexanoic acid-induced resistance against Botrytis cinerea in tomato plants. Mol. Plant-Microbe Interact 2009, 22, 1455–1465. [Google Scholar]
- Browse, J. Jasmonate passes muster: A receptor and targets for the defense hormone. Annu. Rev. Plant Biol 2009, 60, 183–205. [Google Scholar]
- Stintzi, A.; Weber, H.; Reymond, P.; Browse, J.; Farmer, E.E. Plant defense in the absence of jasmonic acid: The role of cyclopentenones. Proc. Natl. Acad. Sci. USA 2001, 98, 12837–12842. [Google Scholar]
- Taki, N.; Sasaki-Sekimoto, Y.; Obayashi, T.; Kikuta, A.; Kobayashi, K.; Ainai, T.; Yagi, K.; Sakurai, N.; Suzuki, H.; Masuda, T.; et al. 12-Oxo-phytodienoic acid triggers expression of a distinct set of genes and plays a role in wound-induced gene expression in Arabidopsis. Plant Physiol. 2005, 139, 1268–1283. [Google Scholar]
- Mueller, S.; Hilbert, B.; Dueckershoff, K.; Roitsch, T.; Krischke, M.; Mueller, M.J.; Berger, S. General detoxification and stress responses are mediated by oxidized lipids through TGA transcription factors in Arabidopsis. Plant Cell 2008, 20, 768–785. [Google Scholar]
- Staswick, P.E.; Su, W.P.; Howell, S.H. Methyl jasmonate inhibition of root-growth and induction of a leaf protein are decreased in an Arabidopsis thaliana mutant. Proc. Natl. Acad. Sci. USA 1992, 89, 6837–6840. [Google Scholar]
- Vellosillo, T.; Martinez, M.; Lopez, M.A.; Vicente, J.; Cascon, T.; Dolan, L.; Hamberg, M.; Castresana, C. Oxylipins produced by the 9-lipoxygenase pathway in Arabidopsis regulate lateral root development and defense responses through a specific signaling cascade. Plant Cell 2007, 19, 831–846. [Google Scholar]
- Yan, Y.; Stolz, S.; Chételat, A.; Reymond, P.; Pagni, M.; Dubugnon, L.; Farmer, E.E. A downstream mediator in the growth repression limb of the jasmonate pathway. Plant Cell 2007, 19, 2470–2483. [Google Scholar]
- Chico, J.M.; Chini, A.; Fonseca, S.; Solano, R. JAZ repressors set the rhythm in jasmonate signaling. Curr. Opin. Plant Biol 2008, 11, 486–494. [Google Scholar]
- El Oirdi, M.; El Rahman, T.A.; Rigano, L.; El Hadrami, A.; Rodriguez, M.C.; Daayf, F.; Vojnov, A.; Bouarab, K. Botrytis cinerea manipulates the antagonistic effects between immune pathways to promote disease development in tomato. Plant Cell 2011, 23, 2405–2421. [Google Scholar]
- Veronese, P.; Nakagami, H.; Bluhm, B.; Abuqamar, S.; Chen, X.; Salmeron, J.; Dietrich, R.A.; Hirt, H.; Mengiste, T. The membrane-anchored Botrytis induced kinase1 plays distinct roles in Arabidopsis resistance to necrotrophic and biotrophic pathogens. Plant Cell 2006, 18, 257–273. [Google Scholar]
- Andersson, R.A.; Akita, M.; Pirhonen, M.; Gammelgård, E.; Valkonen, J.P.T. Moss-Erwinia pathosystem reveals possible similarities in pathogenesis and pathogen defense in vascular and nonvascular plants. J. Gen. Plant Pathol 2005, 71, 23–28. [Google Scholar]
- Palva, TK.; Hurtig, M.; Saindrenan, P.; Palva, ET. Salicylic Acid Induced Resistance to Erwinia carotovora subsp, carotovora in tobacco. Mol. Plant-Microbe Interact. 1994, 7, 356–363. [Google Scholar]
- Dangl, J.L.; Dietrich, R.A.; Richberg, M.H. Death Don’t Have No Mercy: Cell Death Programs in Plant-Microbe Interactions. Plant Cell 1996, 8, 1793–1807. [Google Scholar]
- Alvarez, M.E. Salicylic acid in the machinery of hypersensitive cell death and disease resistance. Plant Mol. Biol 2000, 44, 429–442. [Google Scholar]
- Yue, J.; Hu, X.; Sun, H.; Yang, Y.; Huang, J. Widespread impact of horizontal gene transfer on plant colonization of land. Nat. Commun. 2012, 3. [Google Scholar] [CrossRef]
- Wang, B.; Yeun, L.H.; Xue, J.Y.; Liu, Y.; Ané, J.M.; Qiu, Y.L. Presence of three mycorrhizal genes in the common ancestor of land plants suggests a key role of mycorrhizas in the colonization of land by plants. New Phytol 2010, 186, 514–525. [Google Scholar]



© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ponce de León, I.; Montesano, M. Activation of Defense Mechanisms against Pathogens in Mosses and Flowering Plants. Int. J. Mol. Sci. 2013, 14, 3178-3200. https://doi.org/10.3390/ijms14023178
Ponce de León I, Montesano M. Activation of Defense Mechanisms against Pathogens in Mosses and Flowering Plants. International Journal of Molecular Sciences. 2013; 14(2):3178-3200. https://doi.org/10.3390/ijms14023178
Chicago/Turabian StylePonce de León, Inés, and Marcos Montesano. 2013. "Activation of Defense Mechanisms against Pathogens in Mosses and Flowering Plants" International Journal of Molecular Sciences 14, no. 2: 3178-3200. https://doi.org/10.3390/ijms14023178