Antioxidant Enzymatic Activities and Gene Expression Associated with Heat Tolerance in the Stems and Roots of Two Cucurbit Species (“Cucurbita maxima” and “Cucurbita moschata”) and Their Interspecific Inbred Line “Maxchata”

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Heat Stress on Reactive Oxygen Species Production
2.2. Effect of Heat Stress on Lipid Peroxidation
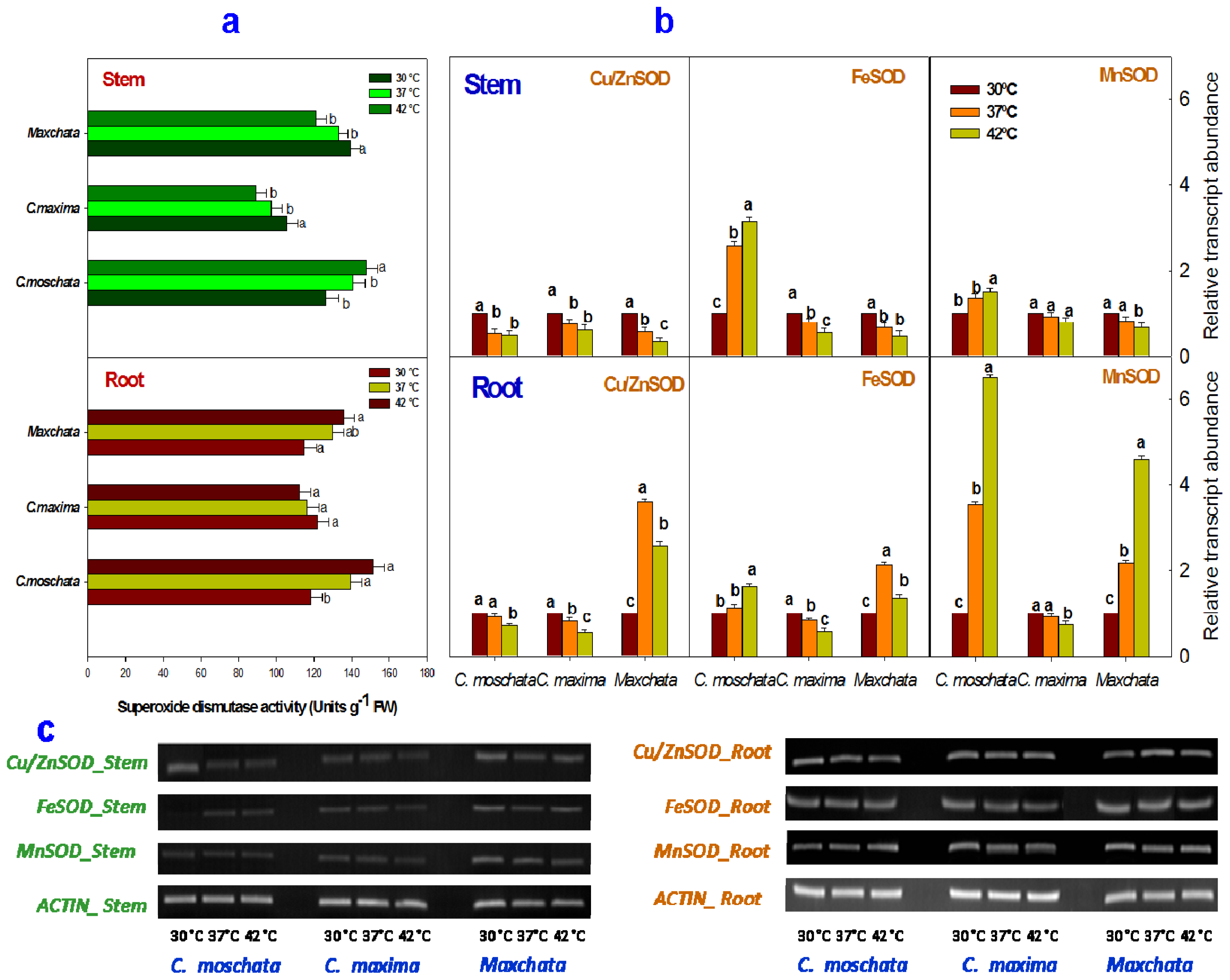
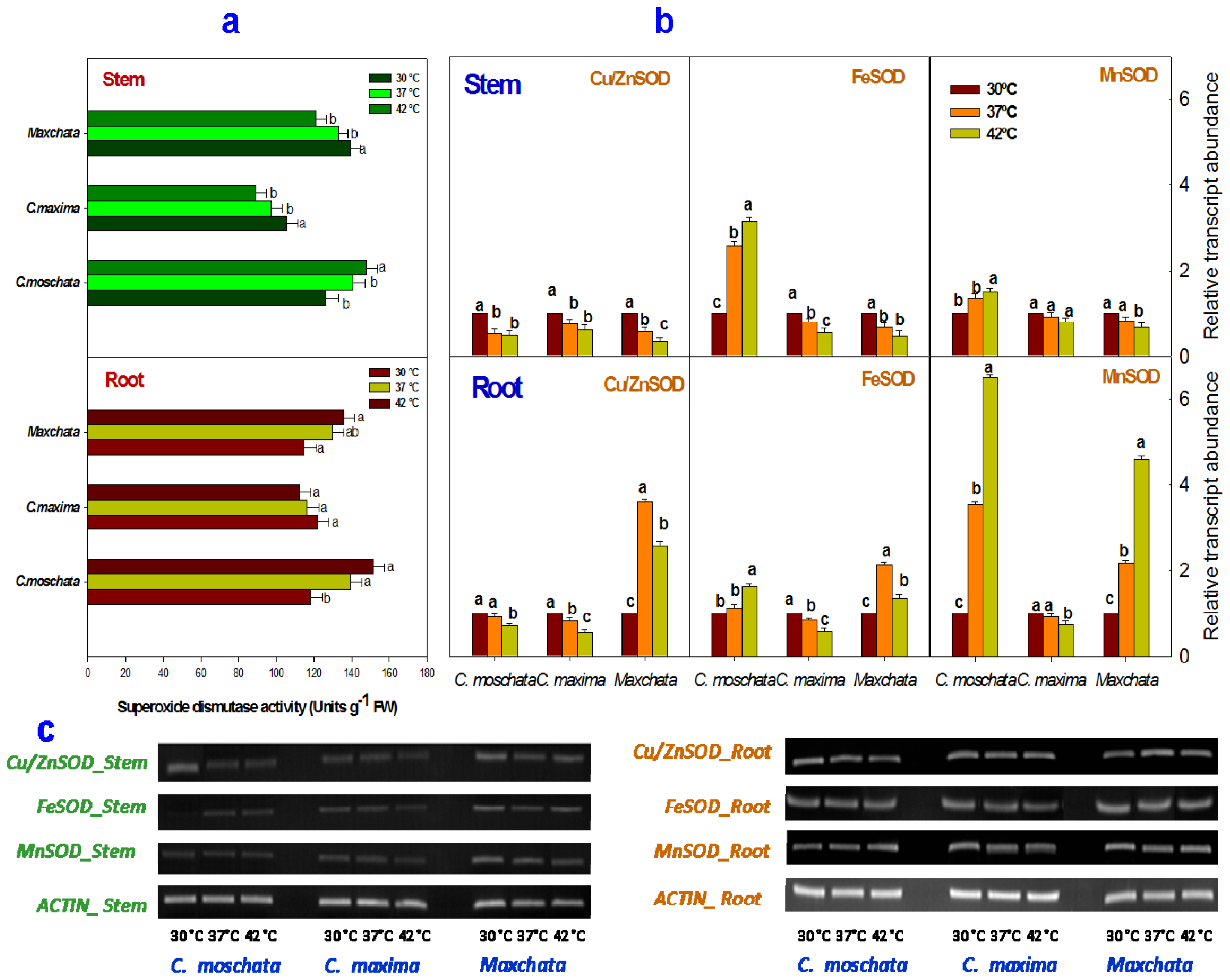
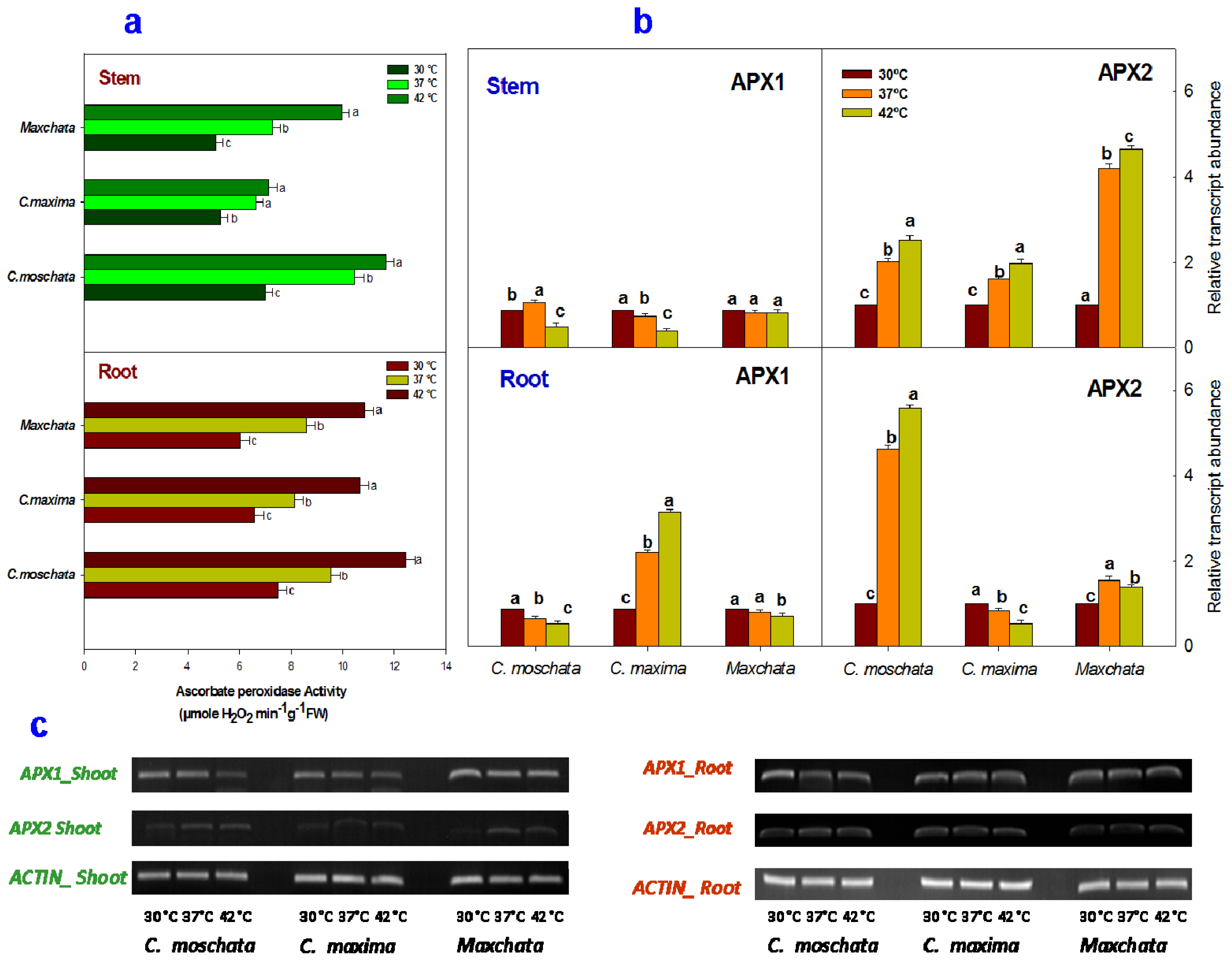
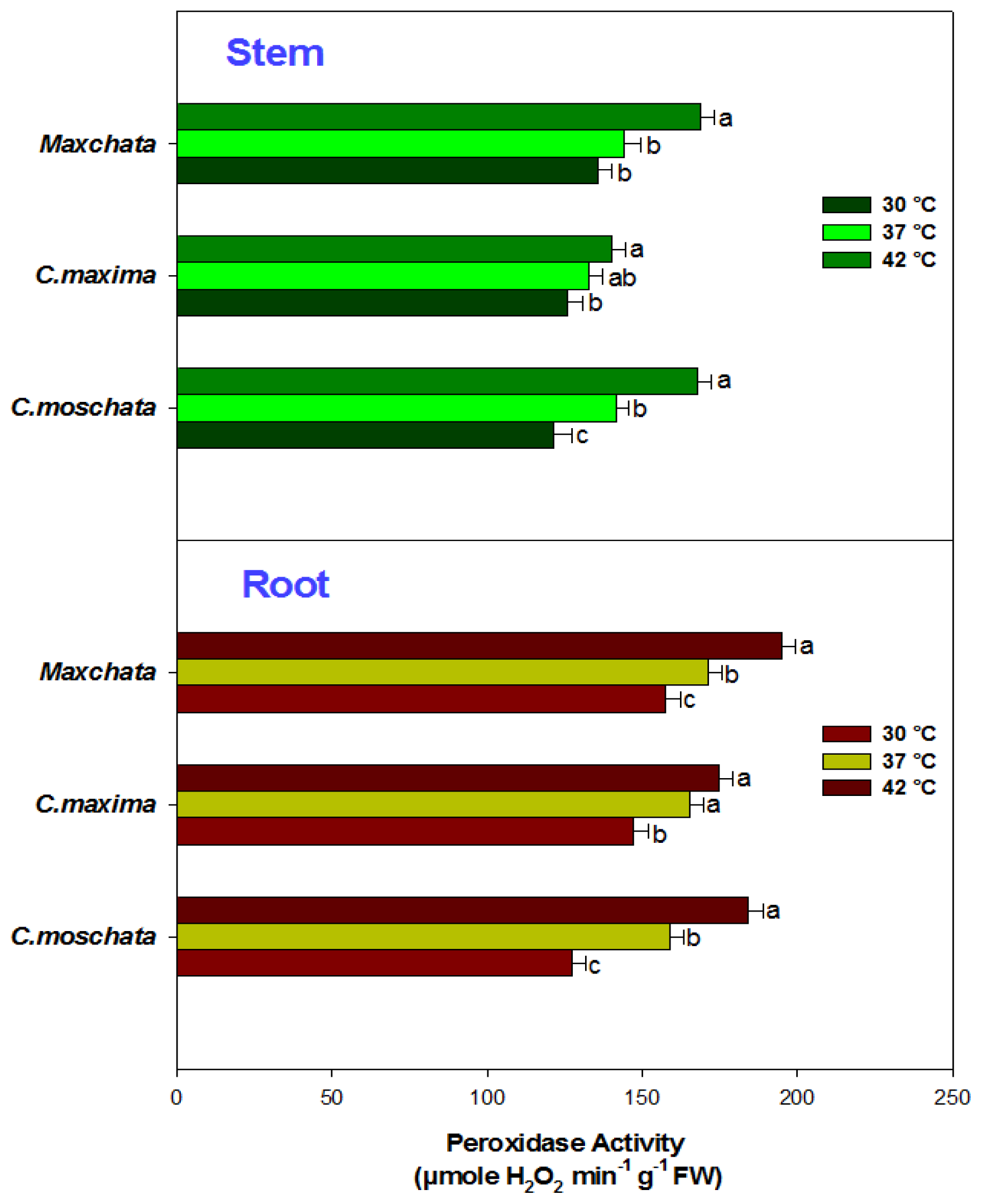
2.3. Activities of Enzymatic Antioxidants and Transcript Abundance Levels of Antioxidant Genes
3. Experimental Section
3.1. Plant Materials and Experimental Conditions
3.2. Quantification of ROS (H2O2 and O2−) Production
3.3. Determination of Lipid Peroxidation
3.4. Measurement of the Enzyme Activities
3.5. Total RNA Isolation and cDNA Synthesis
3.6. Quantitative Real Time-Polymerase Chain Reaction (qRT-PCR) Analysis
3.7. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Abbreviations
| APX | ascorbate peroxidase |
| C. | Cucurbita |
| CAT | catalase |
| SOD | superoxide dismutase |
| H2O2 | hydrogen peroxide |
| MDA | malondialdehyde |
| ELP | Electrolyte leakage percentage |
| O2− | superoxide |
| qRT-PCR | Quantitative Real Time-Polymerase Chain Reaction |
| ROS | reactive oxygen species |
| RT-PCR | Reverse transcriptase-Polymerase Chain Reaction |
Conflicts of Interest
References
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 2002, 7, 405–410. [Google Scholar]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol 2007, 58, 459–481. [Google Scholar]
- Khanna-Chopra, R.; Sabarinath, S. Heat-stable chloroplastic Cu/Zn superoxide dismutase in “Chenopodium murale”. Biochem. Biophys. Res. Commun 2004, 320, 1187–1192. [Google Scholar]
- Houghton, J.T.; Ding, Y.; Griggs, D.J.; Noguer, M.; van der Linden, P.J.; Dai, X.; Maskell, K.; Johnson, C. Climate Change 2001: The Scientific Basis; Cambridge University Press: Cambridge, UK, 2001; Volume 881. [Google Scholar]
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing climate change adaptation needs for food security in 2030. Science 2008, 319, 607–610. [Google Scholar]
- Camejo, D.; Jiménez, A.; Alarcón, J.J.; Torres, W.; Gómez, J.M.; Sevilla, F. Changes in photosynthetic parameters and antioxidant activities following heat-shock treatment in tomato plants. Funct. Plant Biol 2006, 33, 177–187. [Google Scholar]
- Yin, H.; Chen, Q.; Yi, M. Effects of short-term heat stress on oxidative damage and responses of antioxidant system in Lilium longiflorum. Plant Growth Regul 2008, 54, 45–54. [Google Scholar]
- Paris, H. Genetic Analysis and Breeding of Pumpkins and Squash for High Carotene Content. In Vegetables and Vegetable Products; Springer-Verlag: Berlin, Germany, 1994; Volume 16, pp. 93–115. [Google Scholar]
- Bisognin, D.A. Origin and evolution of cultivated cucurbits. Ciênc. Rural 2002, 32, 715–723. [Google Scholar]
- Ara, N.; Yang, J.H.; Hu, Z.Y.; Zhang, M.F. Determining heat tolerance of interspecific (Cucurbita maxima × Cucurbita moschata) inbred line of squash “Maxchata” and its parents through photosynthetic response. Tarım Bilim. Derg.-J. Agric. Sci 2013, 19, 188–197. [Google Scholar]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ 2005, 28, 1056–1071. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem 2010, 48, 909–930. [Google Scholar]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot 2012, 217037, 1–26. [Google Scholar]
- Du, H.; Zhou, P.; Huang, B. Antioxidant enzymatic activities and gene expression associated with heat tolerance in a cool-season perennial grass species. Environ. Exp. Bot 2013, 87, 159–166. [Google Scholar]
- Chaitanya, K.; Sundar, D.; Masilamani, S.; Reddy, A.R. Variation in heat stress-induced antioxidant enzyme activities among three mulberry cultivars. Plant Growth Regul 2002, 36, 175–180. [Google Scholar]
- Mohamed, A.A.; Castagna, A.; Ranieri, A.; Sanità di Toppi, L. Cadmium tolerance in Brassica juncea roots and shoots is affected by antioxidant status and phytochelatin biosynthesis. Plant Physiol. Biochem 2012, 57, 15–22. [Google Scholar]
- Karabal, E.; Yücel, M.; Öktem, H.A. Antioxidant responses of tolerant and sensitive barley cultivars to boron toxicity. Plant Sci 2003, 164, 925–933. [Google Scholar]
- Verma, S.; Dubey, R. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci 2003, 164, 645–655. [Google Scholar]
- Liu, X.; Huang, B. Heat stress injury in relation to membrane lipid peroxidation in creeping bentgrass. Crop Sci 2000, 40, 503–510. [Google Scholar]
- Lin, K.H.; Huang, H.C.; Lin, C.Y. Cloning, expression and physiological analysis of broccoli catalase gene and chinese cabbage ascorbate peroxidase gene under heat stress. Plant Cell Rep 2010, 29, 575–593. [Google Scholar]
- Hao, H.P.; Jiang, C.-D.; Zhang, S.R.; Tang, Y.D.; Shi, L. Enhanced thermal-tolerance of photosystem II by elevating root zone temperature in Prunus mira koehne seedlings. Plant Soil 2012, 353, 367–378. [Google Scholar]
- Guo, Y.P.; Zhou, H.F.; Zhang, L.C. Photosynthetic characteristics and protective mechanisms against photooxidation during high temperature stress in two citrus species. Sci. Hortic 2006, 108, 260–267. [Google Scholar]
- Kumar, S.; Gupta, D.; Nayyar, H. Comparative response of maize and rice genotypes to heat stress: Status of oxidative stress and antioxidants. Acta Physiol. Plant 2012, 34, 75–86. [Google Scholar]
- Sato, T.; Kubo, M.; Watanabe, S. Heat shock induces a systemic acquired resistance (SAR)-related gene via salicylic acid accumulation in cucumber (Cucumis sativus L.). Jpn. J. Trop. Agric 2003, 47, 77–82. [Google Scholar]
- Dutilleul, C.; Garmier, M.; Noctor, G.; Mathieu, C.; Chétrit, P.; Foyer, C.H.; de Paepe, R. Leaf mitochondria modulate whole cell redox homeostasis, set antioxidant capacity, and determine stress resistance through altered signaling and diurnal regulation. Plant Cell 2003, 15, 1212–1226. [Google Scholar]
- Lee, Y.P.; Kim, S.H.; Bang, J.W.; Lee, H.S.; Kwak, S.S.; Kwon, S.Y. Enhanced tolerance to oxidative stress in transgenic tobacco plants expressing three antioxidant enzymes in chloroplasts. Plant Cell Rep 2007, 26, 591–598. [Google Scholar]
- Hu, W.; Xiao, Y.; Zeng, J.; Hu, X. Photosynthesis, respiration and antioxidant enzymes in pepper leaves under drought and heat stresses. Biol. Plant 2010, 54, 761–765. [Google Scholar]
- Nagi, K.; Devaraj, V. Antioxidant and other biochemical defense responses of Macrotyloma uniflorum (lam) verdc. (horse gram) induced by high temperature and salt stress. Braz. J. Plant Physiol 2011, 23, 187–195. [Google Scholar]
- Sreenivasulu, N.; Grimm, B.; Wobus, U.; Weschke, W. Differential response of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet (Setaria italica). Physiol. Plant 2000, 109, 435–442. [Google Scholar]
- Sudhakar, C.; Lakshmi, A.; Giridarakumar, S. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (“Morus alba” L.) under NaCl salinity. Plant Sci 2001, 161, 613–619. [Google Scholar]
- Sairam, R.; Srivastava, G. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci 2002, 162, 897–904. [Google Scholar]
- Lu, P.; Sang, W.G.; Ma, K.P. Differential responses of the activities of antioxidant enzymes to thermal stresses between two invasive Eupatorium species in China. J. Integr. Plant Biol 2008, 50, 393–401. [Google Scholar]
- He, Y.; Huang, B. Differential responses to heat stress in activities and isozymes of four antioxidant enzymes for two cultivars of Kentucky bluegrass contrasting in heat tolerance. J. Am. Soci. Hortic. Sci 2010, 135, 116–124. [Google Scholar]
- Bhoomika, K.; Pyngrope, S.; Dubey, R.S. Differential responses of antioxidant enzymes to aluminum toxicity in two rice (Oryza sativa L.) cultivars with marked presence and elevated activity of Fe SOD and enhanced activities of Mn SOD and Catalase in aluminum tolerant cultivar. Plant Growth Regul 2013, 71, 235–252. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol 2004, 55, 373–399. [Google Scholar]
- Kaur, K.; Kaur, N.; Gupta, A.K.; Singh, I. Exploration of the antioxidative defense system to characterize chickpea genotypes showing differential response towards water deficit conditions. Plant Growth Regul 2013, 70, 49–60. [Google Scholar]
- Scandalios, J. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res 2005, 38, 995–1014. [Google Scholar]
- Esaka, M.; Yamada, N.; Kitabayashi, M.; Setoguchi, Y.; Tsugeki, R.; Kondo, M.; Nishimura, M. cDNA cloning and differential gene expression of three catalases in pumpkin. Plant Mol. Biol 1997, 33, 141–155. [Google Scholar]
- Devi, R.; Kaur, N.; Gupta, A.K. Potential of antioxidant enzymes in depicting drought tolerance of wheat (Triticum aestivum L.). Indian J. Biochem. Biophys 2012, 49, 257–265. [Google Scholar]
- Mittler, R.; Zilinskas, B. Molecular cloning and characterization of a gene encoding pea cytosolic ascorbate peroxidase. J. Biol. Chem 1992, 267, 21802–21807. [Google Scholar]
- Nahakpam, S.; Shah, K. Expression of key antioxidant enzymes under combined effect of heat and cadmium toxicity in growing rice seedlings. Plant Growth Regul 2011, 63, 23–35. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 1981, 22, 867–880. [Google Scholar]
- Panchuk, I.I.; Volkov, R.A.; Schöffl, F. Heat stress-and heat shock transcription factor-dependent expression and activity of ascorbate peroxidase in arabidopsis. Plant Physiol 2002, 129, 838–853. [Google Scholar]
- Örvar, B.L.; Ellis, B.E. Transgenic tobacco plants expressing antisense RNA for cytosolic ascorbate peroxidase show increased susceptibility to ozone injury. Plant J 1997, 11, 1297–1305. [Google Scholar]
- Morimura, Y.; Iwamoto, K.; Ohya, T.; Igarashi, T.; Nakamura, Y.; Kubo, A.; Tanaka, K.; Ikawa, T. Light-enhanced induction of ascorbate peroxidase in japanese radish roots during postgerminative growth. Plant Sci 1999, 142, 123–132. [Google Scholar]
- Yoshimura, K.; Yabuta, Y.; Ishikawa, T.; Shigeoka, S. Expression of spinach ascorbate peroxidase isoenzymes in response to oxidative stresses. Plant Physiol 2000, 123, 223–234. [Google Scholar]
- Takahama, U. Oxidation of vacuolar and apoplastic phenolic substrates by peroxidase: Physiological significance of the oxidation reactions. Phytochem. Rev 2004, 3, 207–219. [Google Scholar]
- Havey, M.; McCreight, J.; Rhodes, B.; Taurick, G. Differential transmission of the Cucumis organellar genomes. Theor. Appl. Genet 1998, 97, 122–128. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci 2000, 151, 59–66. [Google Scholar]
- Jiang, M.; Zhang, J. Effect of abscisic acid on active oxygen species, antioxidative defense system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol 2001, 42, 1265–1273. [Google Scholar]
- Zhou, W.; Leul, M. Uniconazole-induced tolerance of rape plants to heat stress in relation to changes in hormonal levels, enzyme activities and lipid peroxidation. Plant Growth Regul 1999, 27, 99–104. [Google Scholar]
- Zhang, X. The Measurement and Mechanism of Lipid Peroxidation and SOD, POD and CAT Activities in Biological System. In Research Methodology of Crop Physiology; Agriculture Press: Beijing, China, 1992; pp. 208–211. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol 1984, 105, 121–126. [Google Scholar]
- Zhang, W.; Zhang, F.; Raziuddin, R.; Gong, H.; Yang, Z.; Lu, L.; Ye, Q.; Zhou, W. Effects of 5-aminolevulinic acid on oilseed rape seedling growth under herbicide toxicity stress. J. Plant Growth Regul 2008, 27, 159–169. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; Volume 2. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in Real-Time RT-PCR. Nucleic Acids Res 2001, 29, e45. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant organ | Genotypes | Temperature (°C) | H2O2 content | O2− content | MDA content | |||
|---|---|---|---|---|---|---|---|---|
| (μmol g−1 FW) | % Change compared to the control | (nmol min−1 g−1 FW) | % Change compared to the control | (nmol g−1 FW) | % Change compared to the control | |||
| Stem | C. moschata | 30 | 3.32 ± 0.25 b | - | 4.46 ± 0.24 b | - | 53.55 ± 5.55 a | - |
| 37 | 3.80 ± 0.21 ab | 14.9 | 5.65 ± 0.48 a | 27.0 | 65.15 ± 6.33 a | 21.7 | ||
| 42 | 4.04 ± 0.23 a | 22.3 | 6.39 ± 0.29 a | 43.5 | 77.26 ± 6.15 b | 44.8 | ||
| C. maxima | 30 | 3.11 ±0.25 c | - | 5.13 ± 0.18 c | - | 49.73 ± 5.72 a | - | |
| 37 | 4.40 ± 0.35 b | 41.8 | 7.43 ± 0.31 b | 44.7 | 70.61 ± 4.04 b | 42.9 | ||
| 42 | 5.21 ± 0.30 a | 67.4 | 9.26 ± 0.320 a | 80.4 | 89.12 ± 5.37 c | 81.4 | ||
| Maxchata | 30 | 3.18 ± 0.1 b | - | 4.97 ± 0.333 c | - | 51.41 ± 3.85 a | - | |
| 37 | 3.76 ± 0.27 ab | 18.5 | 6.43 ± 0.34 b | 29.5 | 66.11 ± 5.26 b | 29.0 | ||
| 42 | 4.46 ± 0.16 a | 40.3 | 7.52 ± 0.32 a | 21.7 | 82.75 ± 5.89 c | 61.4 | ||
| Root | C. moschata | 30 | 2.37 ± 0.24 a | - | 3.68 ± 0.35 b | - | 42.68 ± 3.02 b | - |
| 37 | 2.65 ± 0.20 a | 12.7 | 4.34 ± 0.22 b | 18.0 | 50.13 ± 3.44 ab | 17.5 | ||
| 42 | 2.80 ± 0.28 a | 18.4 | 5.14 ± 0.26 a | 40.0 | 57.74 ± 4.85 a | 35.7 | ||
| C. maxima | 30 | 2.28 ± 0.17b | - | 3.91 ± 0.20 c | - | 41.74 ± 3.94 c | - | |
| 37 | 2.98 ± 0.36 ab | 31.7 | 5.45 ± 0.17 b | 39.6 | 56.52 ± 3.63 b | 35.8 | ||
| 42 | 3.67 ± 0.27 b | 61.0 | 6.68 ± 0.31 a | 70.7 | 69.87 ± 4.54 a | 68.5 | ||
| Maxchata | 30 | 2.29 ± 0.16 c | - | 3.87 ± 0.18 c | - | 43.99 ± 4.51 b | - | |
| 37 | 2.62 ± 0.29 b | 15.0 | 4.69 ± 0.32 b | 51.5 | 54.53 ± 4.94 ab | 25.3 | ||
| 42 | 3.05 ± 0.12 a | 33.6 | 5.70 ± 0.31 a | 47.3 | 65.38 ± 5.16 a | 50.4 | ||
| Gene | GenBank Accession No | Forward primer (5′→3′) | Reverse primer (5′→3′) | Amplicon size (bp) |
|---|---|---|---|---|
| Cu/ZnSOD | KF831053, KF831056 | GCATGTCAACTGGACCACAT | CCTTTCCATCTTCACCAACA | 95 |
| FeSOD | KF831054, KF831057 | AGATCTGGAACCATGATTTTCTG | GACTTGAATTCTTCCAAAAACTTTT | 110 |
| MnSOD | KF831055, KF831058 | TCTGGATGGGTGTGGCTTGCTCT | GCATGCTCCCAAACATCGATC | 120 |
| APX1 | KF831043, KF831045 | TGAGCTCGCCCATGGCGCCAA | ATCTCAACAGCAACAACACCAG | 112 |
| APX2 | KF831044, KF831046 | TGCTGTCGAGATCACTGGA | CCCTCAAATGATCAGAACCC | 101 |
| CAT1 | KF831047, KF831050 | TACTCAGAGGCACCGTCTTG | CTCCTCATCTCGGTGCATAA | 105 |
| CAT2 | KF831048, KF831051 | TTCGTGAATCGGTTCGTAGA | CAGAGACCTGTCTGCCTGAG | 98 |
| CAT3 | KF831049, KF831052 | GTGGAAGCGTTGTCGGATCC | GTTCGGCCTCACGTTCAGTC | 100 |
| ACTIN | KF831059, KF831060 | CCGCTCTTGCTCCGAGCAG | ATCCACATCTGTTGGAAGGTAC | 120 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ara, N.; Nakkanong, K.; Lv, W.; Yang, J.; Hu, Z.; Zhang, M. Antioxidant Enzymatic Activities and Gene Expression Associated with Heat Tolerance in the Stems and Roots of Two Cucurbit Species (“Cucurbita maxima” and “Cucurbita moschata”) and Their Interspecific Inbred Line “Maxchata”. Int. J. Mol. Sci. 2013, 14, 24008-24028. https://doi.org/10.3390/ijms141224008
Ara N, Nakkanong K, Lv W, Yang J, Hu Z, Zhang M. Antioxidant Enzymatic Activities and Gene Expression Associated with Heat Tolerance in the Stems and Roots of Two Cucurbit Species (“Cucurbita maxima” and “Cucurbita moschata”) and Their Interspecific Inbred Line “Maxchata”. International Journal of Molecular Sciences. 2013; 14(12):24008-24028. https://doi.org/10.3390/ijms141224008
Chicago/Turabian StyleAra, Neelam, Korakot Nakkanong, Wenhui Lv, Jinghua Yang, Zhongyuan Hu, and Mingfang Zhang. 2013. "Antioxidant Enzymatic Activities and Gene Expression Associated with Heat Tolerance in the Stems and Roots of Two Cucurbit Species (“Cucurbita maxima” and “Cucurbita moschata”) and Their Interspecific Inbred Line “Maxchata”" International Journal of Molecular Sciences 14, no. 12: 24008-24028. https://doi.org/10.3390/ijms141224008