The Emerging Roles of ATP-Dependent Chromatin Remodeling Enzymes in Nucleotide Excision Repair

Abstract
:1. Chromatin Structure and DNA Accessibility
2. Nucleotide Excision Repair (NER) in Chromatin
3. Modulation of NER by ATP-Dependent Chromatin Remodelers
4. SWI/SNF Subfamily
5. ISWI Subfamily
6. INO80 Subfamily
7. CSB (Cockayne Syndrome Group B)
8. Crosstalk between ACR and Histone Modifications
9. Concluding Remarks and Perspectives
Acknowledgments
Abbreviations
| ACR | ATP-dependent chromatin remodeler |
| ATP | Adenosine-5′-triphosphate |
| BRG1 | Brm-related gene 1 |
| BRM | Brahma protein |
| CHD | Chromodomain, Helicase, DNA binding |
| CPD | Cyclobutane pyrimidine dimer |
| DEAD/H | Asp-Glu-Ala-Asp/His helicases |
| ERCC1 | Excision Repair Cross Complementing |
| GG-NER | Global Genome Nucleotide Excision Repair |
| HML | Hidden Mat Left |
| ISWI | Imitation Switch |
| INO80 | Inositol Requiring 80 |
| NER | Nucleotide Excision Repair |
| PHD | Plant Homeo Domain |
| RSC | Remodel the Structure of Chromatin |
| SANT | DNA binding domain in SWI3, ADA2, N-CoR and TFIIIB |
| SLIDE | SANT like ISWI |
| SNF2 | Sucrose Non-Fermenting |
| SWI/SNF | Switching defective/Sucrose nonfermenting |
| TC-NER | Transcription Coupled Nucleotide Excision Repair |
| XPC | Xeroderma Pigmentosum Group C |
| XPF | Xeroderma Pigmentosum Group F |
| XPG | Xeroderma Pigmentosum Group G |
References
- Tan, S.; Davey, C.A. Nucleosome structural studies. Curr. Opin. Struct. Biol 2011, 21, 128–136. [Google Scholar]
- Fudenberg, G.; Mirny, L.A. Higher-order chromatin structure: Bridging physics and biology. Curr. Opin. Genet. Dev 2012, 22, 115–124. [Google Scholar]
- Schlick, T.; Hayes, J.; Grigoryev, S. Toward convergence of experimental studies and theoretical modeling of the chromatin fiber. J. Biol. Chem 2012, 287, 5183–5191. [Google Scholar]
- Gkikopoulos, T.; Schofield, P.; Singh, V.; Pinskaya, M.; Mellor, J.; Smolle, M.; Workman, J.L.; Barton, G.J.; Owen-Hughes, T. A role for Snf2-related nucleosome-spacing enzymes in genome-wide nucleosome organization. Science 2011, 333, 1758–1760. [Google Scholar]
- Lee, W.; Tillo, D.; Bray, N.; Morse, R.H.; Davis, R.W.; Hughes, T.R.; Nislow, C. A high-resolution atlas of nucleosome occupancy in yeast. Nat. Genet 2007, 39, 1235–1244. [Google Scholar]
- Richmond, T.J.; Davey, C.A. The structure of DNA in the nucleosome core. Nature 2003, 423, 145–150. [Google Scholar]
- Tamaru, H. Confining euchromatin/heterochromatin territory: Jumonji crosses the line. Genes Dev 2010, 24, 1465–1478. [Google Scholar]
- Lusser, A.; Kadonaga, J.T. Chromatin remodeling by ATP-dependent molecular machines. Bioessays 2003, 25, 1192–1200. [Google Scholar]
- Hopfner, K.P.; Gerhold, C.B.; Lakomek, K.; Wollmann, P. Swi2/Snf2 remodelers: Hybrid views on hybrid molecular machines. Curr. Opin. Struct. Biol 2012, 22, 225–233. [Google Scholar]
- Racki, L.R.; Narlikar, G.J. ATP-dependent chromatin remodeling enzymes: Two heads are not better, just different. Curr. Opin. Genet. Dev 2008, 18, 137–144. [Google Scholar]
- Buchler, N.E.; Bai, L. Chromatin: Bind at your own RSC. Curr. Biol 2011, 21, R223–R225. [Google Scholar]
- Wang, X.; Bai, L.; Bryant, G.O.; Ptashne, M. Nucleosomes and the accessibility problem. Trends Genet 2011, 27, 487–492. [Google Scholar]
- Wang, G.G.; Allis, C.D.; Chi, P. Chromatin remodeling and cancer, Part II: ATP-dependent chromatin remodeling. Trends Mol. Med 2007, 13, 373–380. [Google Scholar]
- Wilson, B.G.; Roberts, C.W. SWI/SNF nucleosome remodellers and cancer. Nat. Rev. Cancer 2011, 11, 481–492. [Google Scholar]
- Jones, K.L.; Zhang, L.; Seldeen, K.L.; Gong, F. Detection of bulky DNA lesions: DDB2 at the interface of chromatin and DNA repair in eukaryotes. IUBMB Life 2010, 62, 803–811. [Google Scholar]
- Lans, H.; Marteijn, J.A.; Vermeulen, W. ATP-dependent chromatin remodeling in the DNA-damage response. Epigenet. Chromatin 2012, 5, 4. [Google Scholar]
- Palomera-Sanchez, Z.; Zurita, M. Open, repair and close again: Chromatin dynamics and the response to UV-induced DNA damage. DNA Repair 2011, 10, 119–125. [Google Scholar]
- Waters, R.; Teng, Y.; Yu, Y.; Yu, S.; Reed, S.H. Tilting at windmills? The nucleotide excision repair of chromosomal DNA. DNA Repair 2009, 8, 146–152. [Google Scholar]
- Yu, S.; Teng, Y.; Waters, R.; Reed, S.H. How chromatin is remodelled during DNA repair of UV-induced DNA damage in Saccharomyces cerevisiae. PLoS Genet 2011, 7, e1002124. [Google Scholar]
- Zhang, L.; Jones, K.; Gong, F. The molecular basis of chromatin dynamics during nucleotide excision repair. Biochem. Cell Biol 2009, 87, 265–272. [Google Scholar]
- Smerdon, M.J.; Conconi, A. Modulation of DNA damage and DNA repair in chromatin. Prog. Nucleic Acid Res. Mol. Biol 1999, 62, 227–255. [Google Scholar]
- Smerdon, M.J.; Lieberman, M.W. Nucleosome rearrangement in human chromatin during UV-induced DNA- reapir synthesis. Proc. Natl. Acad. Sci. USA 1978, 75, 4238–4241. [Google Scholar]
- Misteli, T.; Soutoglou, E. The emerging role of nuclear architecture in DNA repair and genome maintenance. Nat. Rev. Mol. Cell Biol 2009, 10, 243–254. [Google Scholar]
- Hanawalt, P.C. Genomic instability: Environmental invasion and the enemies within. Mutat. Res 1998, 400, 117–125. [Google Scholar]
- Hoeijmakers, J.H. Genome maintenance mechanisms for preventing cancer. Nature 2001, 411, 366–374. [Google Scholar]
- Cadet, J.; Sage, E.; Douki, T. Ultraviolet radiation-mediated damage to cellular DNA. Mutat. Res 2005, 571, 3–17. [Google Scholar]
- Park, H.; Zhang, K.; Ren, Y.; Nadji, S.; Sinha, N.; Taylor, J.S.; Kang, C. Crystal structure of a DNA decamer containing a cis-syn thymine dimer. Proc. Natl. Acad. Sci. USA 2002, 99, 15965–15970. [Google Scholar]
- Friedberg, E.C. How nucleotide excision repair protects against cancer. Nat. Rev. Cancer 2001, 1, 22–33. [Google Scholar]
- Leibeling, D.; Laspe, P.; Emmert, S. Nucleotide excision repair and cancer. J. Mol. Histol 2006, 37, 225–238. [Google Scholar]
- Hanawalt, P.C. Subpathways of nucleotide excision repair and their regulation. Oncogene 2002, 21, 8949–8956. [Google Scholar]
- Hanawalt, P.C.; Spivak, G. Transcription-coupled DNA repair: Two decades of progress and surprises. Nat. Rev. Mol. Cell Biol 2008, 9, 958–970. [Google Scholar]
- Sarker, A.H.; Tsutakawa, S.E.; Kostek, S.; Ng, C.; Shin, D.S.; Peris, M.; Campeau, E.; Tainer, J.A.; Nogales, E.; Cooper, P.K. Recognition of RNA polymerase II and transcription bubbles by XPG, CSB, and TFIIH: Insights for transcription-coupled repair and Cockayne Syndrome. Mol. Cell 2005, 20, 187–198. [Google Scholar]
- Nakazawa, Y.; Sasaki, K.; Mitsutake, N.; Matsuse, M.; Shimada, M.; Nardo, T.; Takahashi, Y.; Ohyama, K.; Ito, K.; Mishima, H.; et al. Mutations in UVSSA cause UV-sensitive syndrome and impair RNA polymerase IIo processing in transcription-coupled nucleotide-excision repair. Nat. Genet 2012, 44, 586–592. [Google Scholar]
- Schwertman, P.; Lagarou, A.; Dekkers, D.H.; Raams, A.; van der Hoek, A.C.; Laffeber, C.; Hoeijmakers, J.H.; Demmers, J.A.; Fousteri, M.; Vermeulen, W.; et al. UV-sensitive syndrome protein UVSSA recruits USP7 to regulate transcription-coupled repair. Nat. Genet 2012, 44, 598–602. [Google Scholar]
- Zhang, X.; Horibata, K.; Saijo, M.; Ishigami, C.; Ukai, A.; Kanno, S.; Tahara, H.; Neilan, E.G.; Honma, M.; Nohmi, T.; et al. Mutations in UVSSA cause UV-sensitive syndrome and destabilize ERCC6 in transcription-coupled DNA repair. Nat. Genet 2012, 44, 593–597. [Google Scholar]
- Sugasawa, K.; Okuda, Y.; Saijo, M.; Nishi, R.; Matsuda, N.; Chu, G.; Mori, T.; Iwai, S.; Tanaka, K.; Tanaka, K.; Hanaoka, F. UV-induced ubiquitylation of XPC protein mediated by UV-DDB-ubiquitin ligase complex. Cell 2005, 121, 387–400. [Google Scholar]
- Min, J.H.; Pavletich, N.P. Recognition of DNA damage by the Rad4 nucleotide excision repair protein. Nature 2007, 449, 570–575. [Google Scholar]
- Oksenych, V.; Coin, F. The long unwinding road: XPB and XPD helicases in damaged DNA opening. Cell Cycle 2010, 9, 90–96. [Google Scholar]
- Wood, R.D. Nucleotide excision repair in mammalian cells. J. Biol. Chem 1997, 272, 23465–23468. [Google Scholar]
- Gong, F.; Kwon, Y.; Smerdon, M.J. Nucleotide excision repair in chromatin and the right of entry. DNA Repair 2005, 4, 884–896. [Google Scholar]
- Hara, R.; Mo, J.; Sancar, A. DNA damage in the nucleosome core is refractory to repair by human excision nuclease. Mol. Cell Biol 2000, 20, 9173–9181. [Google Scholar]
- Green, C.M.; Almouzni, G. When repair meets chromatin. EMBO Rep 2002, 3, 28–33. [Google Scholar]
- Kamileri, I.; Karakasilioti, I.; Garinis, G.A. Nucleotide excision repair: New tricks with old bricks. Trends Genet 2012, in press. [Google Scholar]
- Wang, H.; Zhai, L.; Xu, J.; Joo, H.Y.; Jackson, S.; Erdjument-Bromage, H.; Tempst, P.; Xiong, Y.; Zhang, Y. Histone H3 and H4 ubiquitylation by the CUL4-DDB-ROC1 ubiquitin ligase facilitates cellular response to DNA damage. Mol. Cell 2006, 22, 383–394. [Google Scholar]
- Kapetanaki, M.G.; Guerrero-Santoro, J.; Bisi, D.C.; Hsieh, C.L.; Rapic-Otrin, V.; Levine, A.S. The DDB1-CUL4ADDB2 ubiquitin ligase is deficient in xeroderma pigmentosum group E and targets histone H2A at UV-damaged DNA sites. Proc. Natl. Acad. Sci. USA 2006, 103, 2588–2593. [Google Scholar]
- Le, M.N.; Mota-Fernandes, D.; Velez-Cruz, R.; Iltis, I.; Biard, D.; Egly, J.M. NER factors are recruited to active promoters and facilitate chromatin modification for transcription in the absence of exogenous genotoxic attack. Mol. Cell 2010, 38, 54–66. [Google Scholar]
- Fousteri, M.; Mullenders, L.H. Transcription-coupled nucleotide excision repair in mammalian cells: Molecular mechanisms and biological effects. Cell Res 2008, 18, 73–84. [Google Scholar]
- Huang, J.C.; Sancar, A. Determination of minimum substrate size for human excinuclease. J. Biol. Chem 1994, 269, 19034–19040. [Google Scholar]
- Hara, R.; Sancar, A. The SWI/SNF chromatin-remodeling factor stimulates repair by human excision nuclease in the mononucleosome core particle. Mol. Cell Biol 2002, 22, 6779–6787. [Google Scholar]
- Hara, R.; Sancar, A. Effect of damage type on stimulation of human excision nuclease by SWI/SNF chromatin remodeling factor. Mol. Cell Biol 2003, 23, 4121–4125. [Google Scholar]
- Ura, K.; Araki, M.; Saeki, H.; Masutani, C.; Ito, T.; Iwai, S.; Mizukoshi, T.; Kaneda, Y.; Hanaoka, F. ATP-dependent chromatin remodeling facilitates nucleotide excision repair of UV-induced DNA lesions in synthetic dinucleosomes. EMBO J 2001, 20, 2004–2014. [Google Scholar]
- Chai, B.; Huang, J.; Cairns, B.R.; Laurent, B.C. Distinct roles for the RSC and Swi/Snf ATP-dependent chromatin remodelers in DNA double-strand break repair. Genes Dev 2005, 19, 1656–1661. [Google Scholar]
- Gospodinov, A.; Vaissiere, T.; Krastev, D.B.; Legube, G.; Anachkova, B.; Herceg, Z. Mammalian Ino80 mediates double-strand break repair through its role in DNA end strand resection. Mol. Cell Biol 2011, 31, 4735–4745. [Google Scholar]
- Lukas, J.; Lukas, C.; Bartek, J. More than just a focus: The chromatin response to DNA damage and its role in genome integrity maintenance. Nat. Cell Biol 2011, 13, 1161–1169. [Google Scholar]
- Papamichos-Chronakis, M.; Krebs, J.E.; Peterson, C.L. Interplay between Ino80 and Swr1 chromatin remodeling enzymes regulates cell cycle checkpoint adaptation in response to DNA damage. Genes Dev 2006, 20, 2437–2449. [Google Scholar]
- Polo, S.E.; Kaidi, A.; Baskcomb, L.; Galanty, Y.; Jackson, S.P. Regulation of DNA-damage responses and cell-cycle progression by the chromatin remodelling factor CHD4. EMBO J 2010, 29, 3130–3139. [Google Scholar]
- Sinha, M.; Watanabe, S.; Johnson, A.; Moazed, D.; Peterson, C.L. Recombinational repair within heterochromatin requires ATP-dependent chromatin remodeling. Cell 2009, 138, 1109–1121. [Google Scholar]
- Van Attikum, H.; Fritsch, O.; Hohn, B.; Gasser, S.M. Recruitment of the INO80 complex by H2A phosphorylation links ATP-dependent chromatin remodeling with DNA double-strand break repair. Cell 2004, 119, 777–788. [Google Scholar]
- Gong, F.; Fahy, D.; Smerdon, M.J. Rad4-Rad23 interaction with SWI/SNF links ATP-dependent chromatin remodeling with nucleotide excision repair. Nat. Struct. Mol. Biol 2006, 13, 902–907. [Google Scholar]
- Sarkar, S.; Kiely, R.; McHugh, P.J. The Ino80 chromatin-remodeling complex restores chromatin structure during UV DNA damage repair. J. Cell Biol 2010, 191, 1061–1068. [Google Scholar]
- Winston, F.; Carlson, M. Yeast SNF/SWI transcriptional activators and the SPT/SIN chromatin connection. Trends Genet 1992, 8, 387–391. [Google Scholar]
- Chiba, H.; Muramatsu, M.; Nomoto, A.; Kato, H. Two human homologues of Saccharomyces cerevisiae SWI2/SNF2 and Drosophila brahma are transcriptional coactivators cooperating with the estrogen receptor and the retinoic acid receptor. Nucleic Acids Res 1994, 22, 1815–1820. [Google Scholar]
- Dingwall, A.K.; Beek, S.J.; McCallum, C.M.; Tamkun, J.W.; Kalpana, G.V.; Goff, S.P.; Scott, M.P. The Drosophila snr1 and brm proteins are related to yeast SWI/SNF proteins and are components of a large protein complex. Mol. Biol. Cell 1995, 6, 777–791. [Google Scholar]
- Clapier, C.R.; Cairns, B.R. The biology of chromatin remodeling complexes. Annu. Rev. Biochem 2009, 78, 273–304. [Google Scholar]
- Awad, S.; Hassan, A.H. The Swi2/Snf2 bromodomain is important for the full binding and remodeling activity of the SWI/SNF complex on H3- and H4-acetylated nucleosomes. Ann. N. Y. Acad. Sci 2008, 1138, 366–375. [Google Scholar]
- Mohrmann, L.; Verrijzer, C.P. Composition and functional specificity of SWI2/SNF2 class chromatin remodeling complexes. Biochim. Biophys. Acta 2005, 1681, 59–73. [Google Scholar]
- Fan, H.Y.; Trotter, K.W.; Archer, T.K.; Kingston, R.E. Swapping function of two chromatin remodeling complexes. Mol. Cell 2005, 17, 805–815. [Google Scholar]
- Ghaemmaghami, S.; Huh, W.K.; Bower, K.; Howson, R.W.; Belle, A.; Dephoure, N.; O’Shea, E.K.; Weissman, J.S. Global analysis of protein expression in yeast. Nature 2003, 425, 737–741. [Google Scholar]
- Euskirchen, G.M.; Auerbach, R.K.; Davidov, E.; Gianoulis, T.A.; Zhong, G.; Rozowsky, J.; Bhardwaj, N.; Gerstein, M.B.; Snyder, M. Diverse roles and interactions of the SWI/SNF chromatin remodeling complex revealed using global approaches. PLoS Genet 2011, 7, e1002008. [Google Scholar]
- Whitehouse, I.; Flaus, A.; Cairns, B.R.; White, M.F.; Workman, J.L.; Owen-Hughes, T. Nucleosome mobilization catalysed by the yeast SWI/SNF complex. Nature 1999, 400, 784–787. [Google Scholar]
- Saha, A.; Wittmeyer, J.; Cairns, B.R. Chromatin remodeling through directional DNA translocation from an internal nucleosomal site. Nat. Struct. Mol. Biol 2005, 12, 747–755. [Google Scholar]
- Gaillard, H.; Fitzgerald, D.J.; Smith, C.L.; Peterson, C.L.; Richmond, T.J.; Thoma, F. Chromatin remodeling activities act on UV-damaged nucleosomes and modulate DNA damage accessibility to photolyase. J. Biol. Chem 2003, 278, 17655–17663. [Google Scholar]
- Ray, A.; Mir, S.N.; Wani, G.; Zhao, Q.; Battu, A.; Zhu, Q.; Wang, Q.E.; Wani, A.A. Human SNF5/INI1, a component of the human SWI/SNF chromatin remodeling complex, promotes nucleotide excision repair by influencing ATM recruitment and downstream H2AX phosphorylation. Mol. Cell Biol 2009, 29, 6206–6219. [Google Scholar]
- Zhao, Q.; Wang, Q.E.; Ray, A.; Wani, G.; Han, C.; Milum, K.; Wani, A.A. Modulation of nucleotide excision repair by mammalian SWI/SNF chromatin-remodeling complex. J. Biol. Chem 2009, 284, 30424–30432. [Google Scholar]
- Gong, F.; Fahy, D.; Liu, H.; Wang, W.; Smerdon, M.J. Role of the mammalian SWI/SNF chromatin remodeling complex in the cellular response to UV damage. Cell Cycle 2008, 7, 1067–1074. [Google Scholar]
- Tsukiyama, T.; Wu, C. Purification and properties of an ATP-dependent nucleosome remodeling factor. Cell 1995, 83, 1011–1020. [Google Scholar]
- Boyer, L.A.; Latek, R.R.; Peterson, C.L. The SANT domain: A unique histone-tail-binding module? Nat. Rev. Mol. Cell Biol 2004, 5, 158–163. [Google Scholar]
- Dang, W.; Bartholomew, B. Domain architecture of the catalytic subunit in the ISW2-nucleosome complex. Mol. Cell Biol 2007, 27, 8306–8317. [Google Scholar]
- Erdel, F.; Rippe, K. Binding kinetics of human ISWI chromatin-remodelers to DNA repair sites elucidate their target location mechanism. Nucleus 2011, 2, 105–112. [Google Scholar]
- Grune, T.; Brzeski, J.; Eberharter, A.; Clapier, C.R.; Corona, D.F.; Becker, P.B.; Muller, C.W. Crystal structure and functional analysis of a nucleosome recognition module of the remodeling factor ISWI. Mol. Cell 2003, 12, 449–460. [Google Scholar]
- Corona, D.F.; Langst, G.; Clapier, C.R.; Bonte, E.J.; Ferrari, S.; Tamkun, J.W.; Becker, P.B. ISWI is an ATP-dependent nucleosome remodeling factor. Mol. Cell 1999, 3, 239–245. [Google Scholar]
- Arancio, W.; Onorati, M.C.; Burgio, G.; Collesano, M.; Ingrassia, A.M.; Genovese, S.I.; Fanto, M.; Corona, D.F. The nucleosome remodeling factor ISWI functionally interacts with an evolutionarily conserved network of cellular factors. Genetics 2010, 185, 129–140. [Google Scholar]
- Sala, A.; Toto, M.; Pinello, L.; Gabriele, A.; di Benedetto, V.; Ingrassia, A.M.; Lo Bosco, G.; di Gesù, V.; Giancarlo, R.; Corona, D.F. Genome-wide characterization of chromatin binding and nucleosome spacing activity of the nucleosome remodelling ATPase ISWI. EMBO J 2011, 30, 1766–1777. [Google Scholar]
- Gangaraju, V.K.; Bartholomew, B. Mechanisms of ATP dependent chromatin remodeling. Mutat. Res 2007, 618, 3–17. [Google Scholar]
- Fazzio, T.G.; Tsukiyama, T. Chromatin remodeling in vivo: Evidence for a nucleosome sliding mechanism. Mol. Cell 2003, 12, 1333–1340. [Google Scholar]
- Corona, D.F.; Siriaco, G.; Armstrong, J.A.; Snarskaya, N.; McClymont, S.A.; Scott, M.P.; Tamkun, J.W. ISWI regulates higher-order chromatin structure and histone H1 assembly in vivo. PLoS Biol 2007, 5, e232. [Google Scholar]
- Lans, H.; Marteijn, J.A.; Schumacher, B.; Hoeijmakers, J.H.; Jansen, G.; Vermeulen, W. Involvement of global genome repair, transcription coupled repair, and chromatin remodeling in UV DNA damage response changes during development. PLoS Genet 2010, 6, e1000941. [Google Scholar]
- Shen, X.; Mizuguchi, G.; Hamiche, A.; Wu, C. A chromatin remodelling complex involved in transcription and DNA processing. Nature 2000, 406, 541–544. [Google Scholar]
- Papamichos-Chronakis, M.; Watanabe, S.; Rando, O.J.; Peterson, C.L. Global regulation of H2A.Z localization by the INO80 chromatin-remodeling enzyme is essential for genome integrity. Cell 2011, 144, 200–213. [Google Scholar]
- Cai, Y.; Jin, J.; Gottschalk, A.J.; Yao, T.; Conaway, J.W.; Conaway, R.C. Purification and assay of the human INO80 and SRCAP chromatin remodeling complexes. Methods 2006, 40, 312–317. [Google Scholar]
- Hogan, C.J.; Aligianni, S.; Durand-Dubief, M.; Persson, J.; Will, W.R.; Webster, J.; Wheeler, L.; Mathews, C.K.; Elderkin, S.; Oxley, D.; et al. Fission yeast Iec1-ino80-mediated nucleosome eviction regulates nucleotide and phosphate metabolism. Mol. Cell Biol 2010, 30, 657–674. [Google Scholar]
- Ebbert, R.; Birkmann, A.; Schuller, H.J. The product of the SNF2/SWI2 paralogue INO80 of Saccharomyces cerevisiae required for efficient expression of various yeast structural genes is part of a high-molecular-weight protein complex. Mol. Microbiol 1999, 32, 741–751. [Google Scholar]
- Morrison, A.J.; Kim, J.A.; Person, M.D.; Highland, J.; Xiao, J.; Wehr, T.S.; Hensley, S.; Bao, Y.; Shen, J.; Collins, S.R.; et al. Mec1/Tel1 phosphorylation of the INO80 chromatin remodeling complex influences DNA damage checkpoint responses. Cell 2007, 130, 499–511. [Google Scholar]
- Papamichos-Chronakis, M.; Peterson, C.L. The Ino80 chromatin-remodeling enzyme regulates replisome function and stability. Nat. Struct. Mol. Biol 2008, 15, 338–345. [Google Scholar]
- Jiang, Y.; Wang, X.; Bao, S.; Guo, R.; Johnson, D.G.; Shen, X.; Li, L. INO80 chromatin remodeling complex promotes the removal of UV lesions by the nucleotide excision repair pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 17274–17279. [Google Scholar]
- Czaja, W.; Bespalov, V.A.; Hinz, J.M.; Smerdon, M.J. Proficient repair in chromatin remodeling defective ino80 mutants of Saccharomyces cerevisiae highlights replication defects as the main contributor to DNA damage sensitivity. DNA Repair 2010, 9, 976–984. [Google Scholar]
- Eisen, J.A.; Sweder, K.S.; Hanawalt, P.C. Evolution of the SNF2 family of proteins: Subfamilies with distinct sequences and functions. Nucleic Acids Res 1995, 23, 2715–2723. [Google Scholar]
- Selby, C.P.; Sancar, A. Cockayne syndrome group B protein enhances elongation by RNA polymerase II. Proc. Natl. Acad. Sci. USA 1997, 94, 11205–11209. [Google Scholar]
- Weidenheim, K.M.; Dickson, D.W.; Rapin, I. Neuropathology of Cockayne syndrome: Evidence for impaired development, premature aging, and neurodegeneration. Mech. Ageing Dev 2009, 130, 619–636. [Google Scholar]
- Malik, S.; Chaurasia, P.; Lahudkar, S.; Durairaj, G.; Shukla, A.; Bhaumik, S.R. Rad26p, a transcription-coupled repair factor, is recruited to the site of DNA lesion in an elongating RNA polymerase II-dependent manner in vivo. Nucleic Acids Res 2010, 38, 1461–1477. [Google Scholar]
- Tantin, D.; Kansal, A.; Carey, M. Recruitment of the putative transcription-repair coupling factor CSB/ERCC6 to RNA polymerase II elongation complexes. Mol. Cell Biol 1997, 17, 6803–6814. [Google Scholar]
- Citterio, E.; van den Boom, V.; Schnitzler, G.; Kanaar, R.; Bonte, E.; Kingston, R.E.; Hoeijmakers, J.H.; Vermeulen, W. ATP-dependent chromatin remodeling by the Cockayne syndrome B DNA repair-transcription-coupling factor. Mol. Cell Biol 2000, 20, 7643–7653. [Google Scholar]
- Lake, R.J.; Geyko, A.; Hemashettar, G.; Zhao, Y.; Fan, H.Y. UV-induced association of the CSB remodeling protein with chromatin requires ATP-dependent relief of N-terminal autorepression. Mol. Cell 2010, 37, 235–246. [Google Scholar]
- Suganuma, T.; Workman, J.L. Signals and combinatorial functions of histone modifications. Annu. Rev. Biochem 2011, 80, 473–499. [Google Scholar]
- Chandrasekharan, M.B.; Huang, F.; Sun, Z.W. Ubiquitination of histone H2B regulates chromatin dynamics by enhancing nucleosome stability. Proc. Natl. Acad. Sci. USA 2009, 106, 16686–16691. [Google Scholar]
- Batta, K.; Zhang, Z.; Yen, K.; Goffman, D.B.; Pugh, B.F. Genome-wide function of H2B ubiquitylation in promoter and genic regions. Genes Dev 2011, 25, 2254–2265. [Google Scholar]
- Shogren-Knaak, M.; Ishii, H.; Sun, J.M.; Pazin, M.J.; Davie, J.R.; Peterson, C.L. Histone H4-K16 acetylation controls chromatin structure and protein interactions. Science 2006, 311, 844–847. [Google Scholar]
- Yun, M.; Wu, J.; Workman, J.L.; Li, B. Readers of histone modifications. Cell Res 2011, 21, 564–578. [Google Scholar]
- Dhalluin, C.; Carlson, J.E.; Zeng, L.; He, C.; Aggarwal, A.K.; Zhou, M.M. Structure and ligand of a histone acetyltransferase bromodomain. Nature 1999, 399, 491–496. [Google Scholar]
- Hassan, A.H.; Neely, K.E.; Workman, J.L. Histone acetyltransferase complexes stabilize swi/snf binding to promoter nucleosomes. Cell 2001, 104, 817–827. [Google Scholar]
- Hassan, A.H.; Prochasson, P.; Neely, K.E.; Galasinski, S.C.; Chandy, M.; Carrozza, M.J.; Workman, J.L. Function and selectivity of bromodomains in anchoring chromatin-modifying complexes to promoter nucleosomes. Cell 2002, 111, 369–379. [Google Scholar]
- Nag, R.; Kyriss, M.; Smerdon, J.W.; Wyrick, J.J.; Smerdon, M.J. A cassette of N-terminal amino acids of histone H2B are required for efficient cell survival, DNA repair and Swi/Snf binding in UV irradiated yeast. Nucleic Acids Res 2010, 38, 1450–1460. [Google Scholar]
- Kasten, M.; Szerlong, H.; Erdjument-Bromage, H.; Tempst, P.; Werner, M.; Cairns, B.R. Tandem bromodomains in the chromatin remodeler RSC recognize acetylated histone H3 Lys14. EMBO J 2004, 23, 1348–1359. [Google Scholar]
- Yu, Y.; Teng, Y.; Liu, H.; Reed, S.H.; Waters, R. UV irradiation stimulates histone acetylation and chromatin remodeling at a repressed yeast locus. Proc. Natl. Acad. Sci. USA 2005, 102, 8650–8655. [Google Scholar]
- Teng, Y.; Liu, H.; Gill, H.W.; Yu, Y.; Waters, R.; Reed, S.H. Saccharomyces cerevisiae Rad16 mediates ultraviolet-dependent histone H3 acetylation required for efficient global genome nucleotide-excision repair. EMBO Rep 2008, 9, 97–102. [Google Scholar]
- Bang, D.D.; Verhage, R.; Goosen, N.; Brouwer, J.; van de Putte, P. Molecular cloning of RAD16, a gene involved in differential repair in Saccharomyces cerevisiae. Nucleic Acids Res 1992, 20, 3925–3931. [Google Scholar]
- Yu, S.; Owen-Hughes, T.; Friedberg, E.C.; Waters, R.; Reed, S.H. The yeast Rad7/Rad16/Abf1 complex generates superhelical torsion in DNA that is required for nucleotide excision repair. DNA Repair 2004, 3, 277–287. [Google Scholar]
- Lee, H.S.; Park, J.H.; Kim, S.J.; Kwon, S.J.; Kwon, J. A cooperative activation loop among SWI/SNF, gamma-H2AX and H3 acetylation for DNA double-strand break repair. EMBO J 2010, 29, 1434–1445. [Google Scholar]
- Morrison, A.J.; Highland, J.; Krogan, N.J.; Rbel-Eden, A.; Greenblatt, J.F.; Haber, J.E.; Shen, X. INO80 and gamma-H2AX interaction links ATP-dependent chromatin remodeling to DNA damage repair. Cell 2004, 119, 767–775. [Google Scholar]
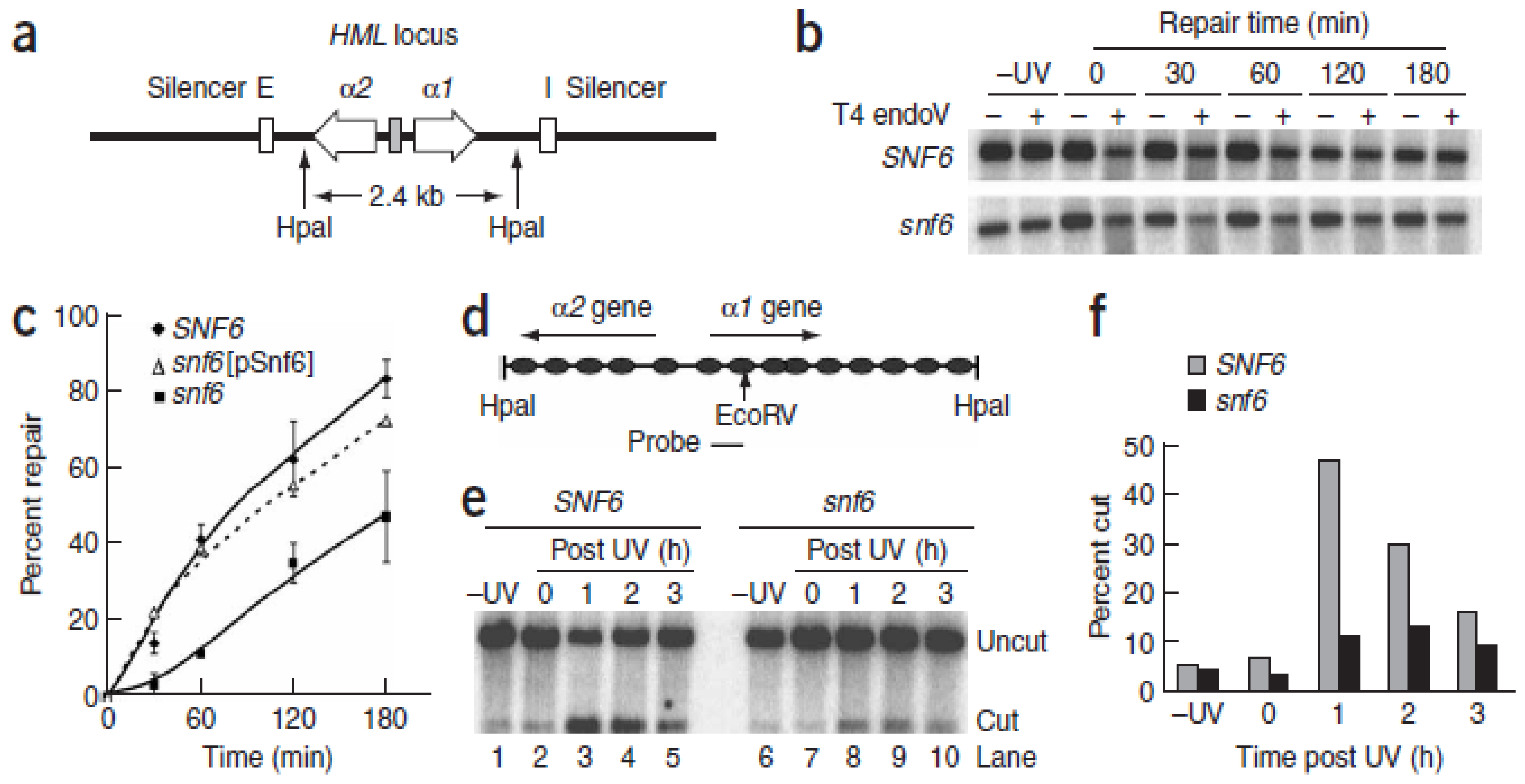

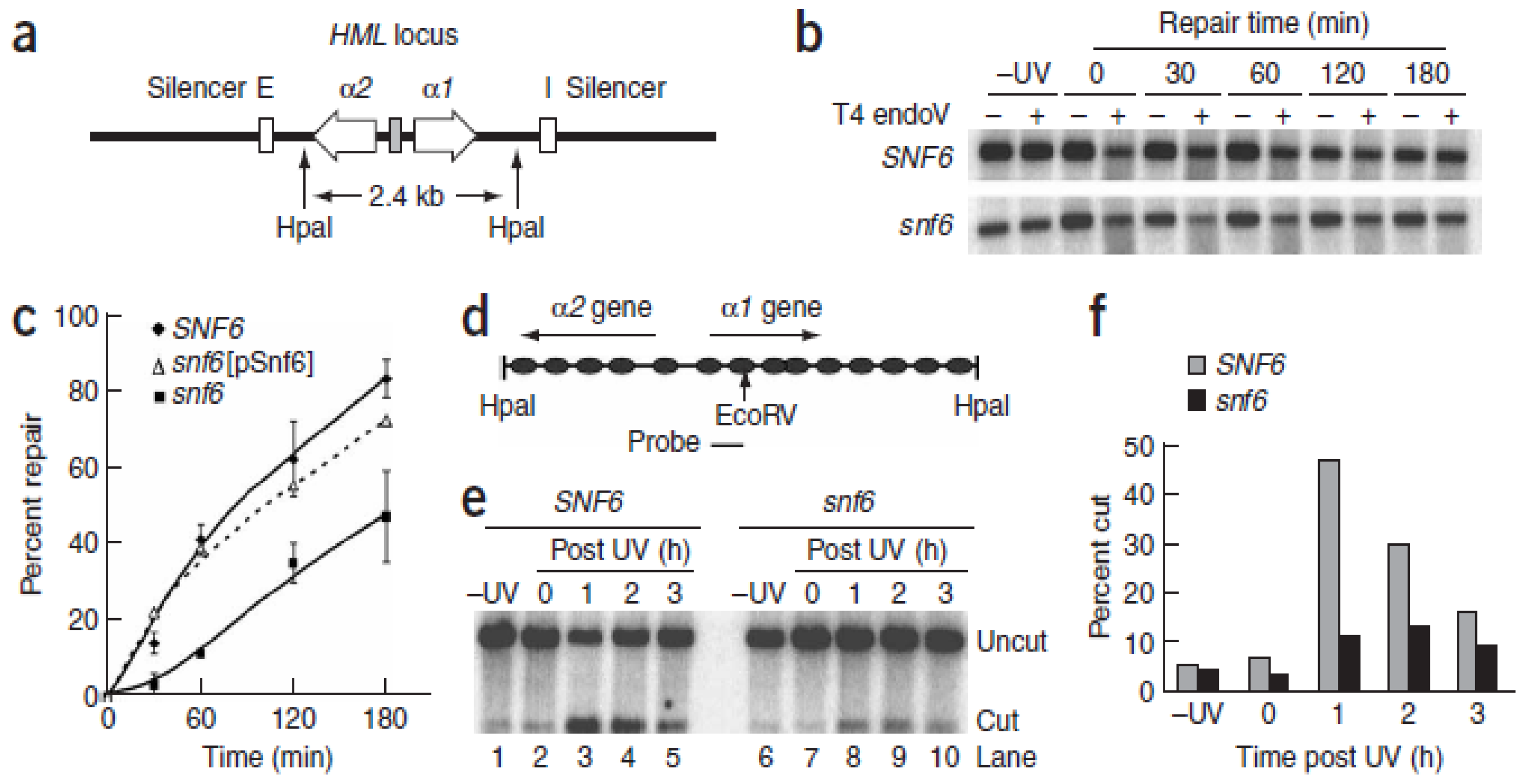{kind=link}
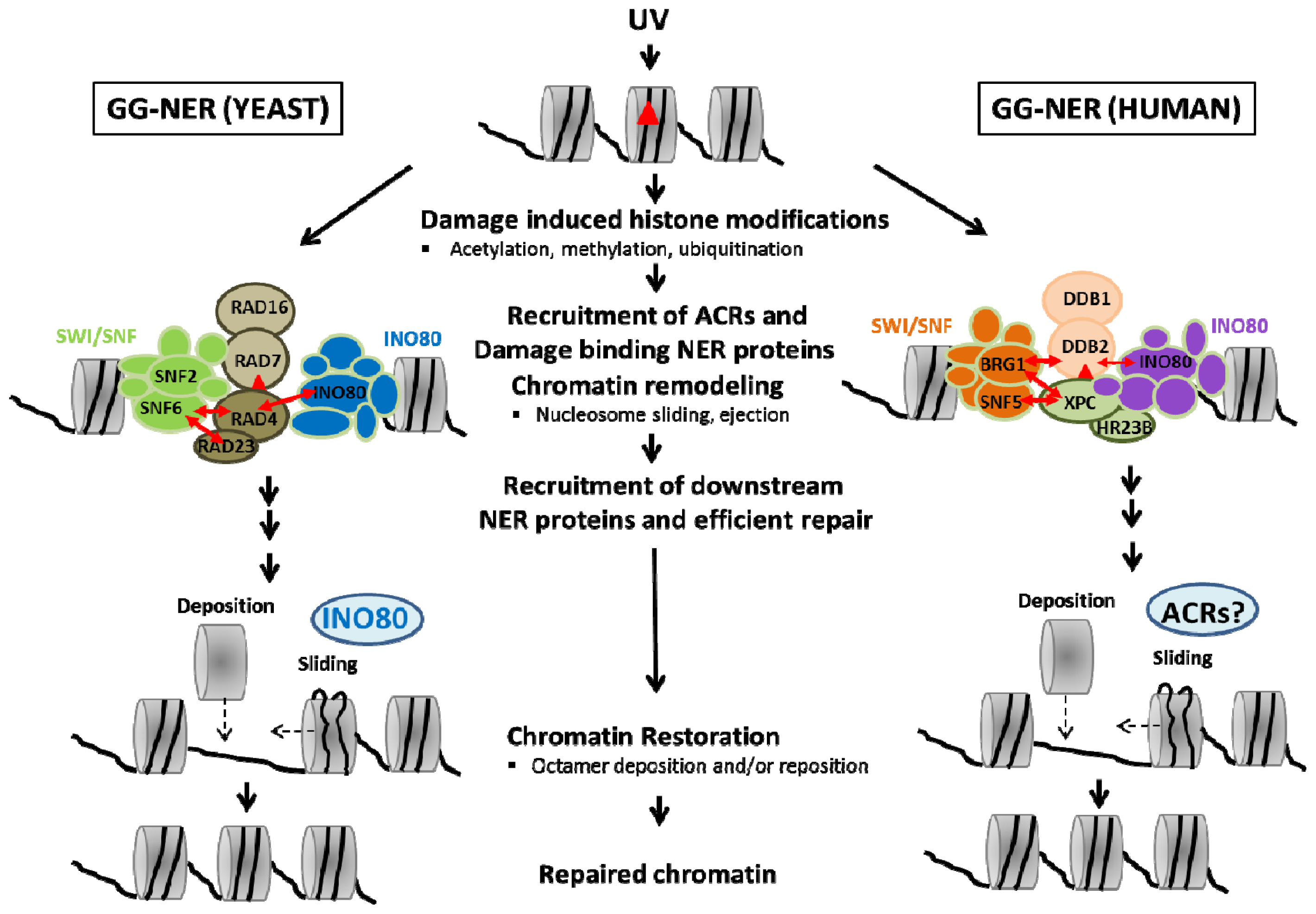
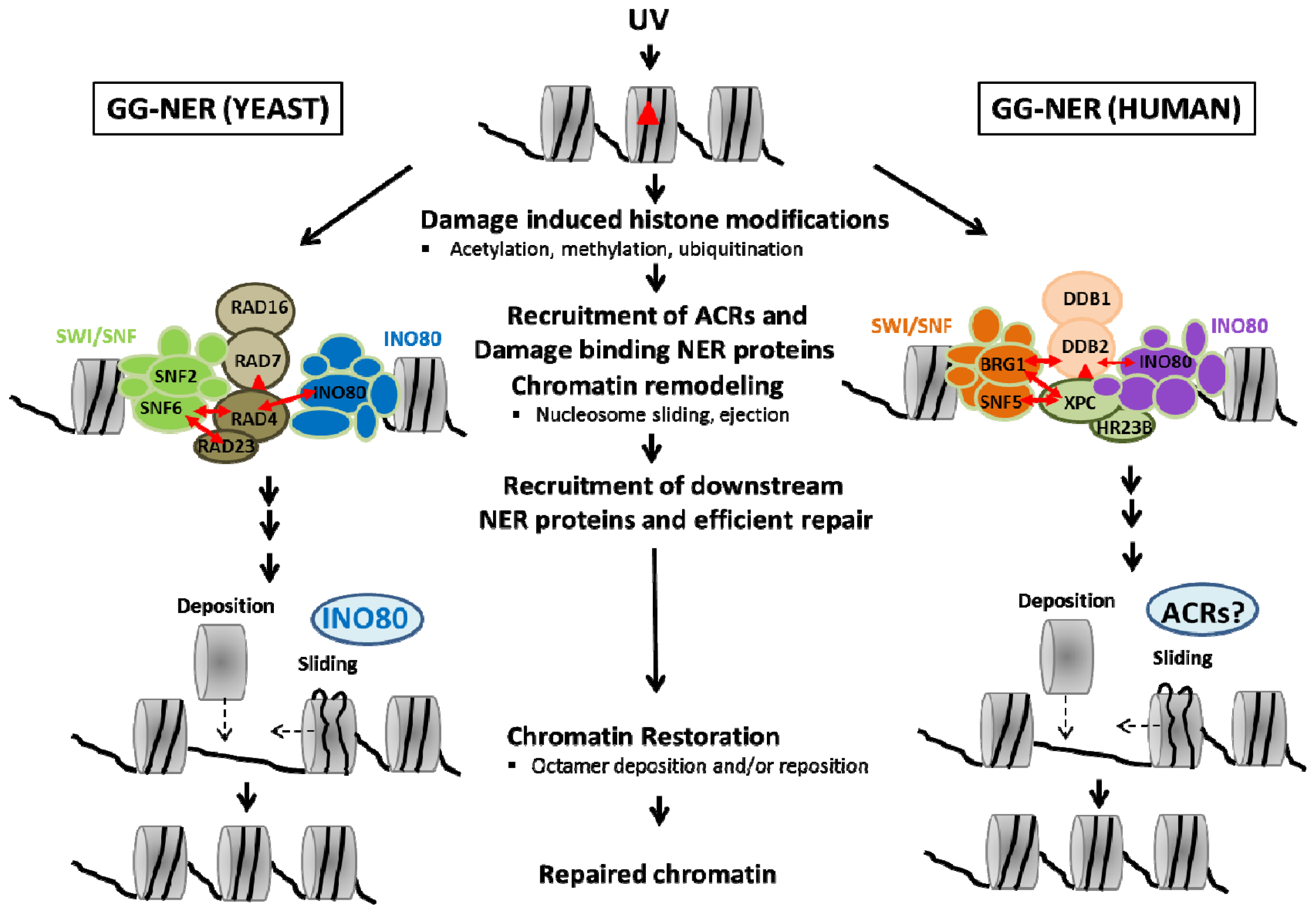{kind=link}
| ACR complex (Number of subunits) | ACR abundance (Molecules per cell) | DNA-dependent ATPase subunit in Yeast and human ACR complexes | Chromatin remodeling activity of ACR complex | Role of ACR in NER | |
|---|---|---|---|---|---|
| Yeast | Human | ||||
| SWI/SNF (9–13) [14,64] | SWI/SNF (~200) [68] | Swi2/Snf2 | BRG1, hBRM | Nucleosome sliding, displacement in trans [70] | Enhances NER in yeast and mammalian cells [59,73,74] |
| ISWI (2–5) [64] | ISWI (~1500) [68] | Isw1 | hSNF2L | Nucleosome sliding, spacing and assembly [79,84]. Promotes chromatin compaction and H1 association with linker [86] | Facilitates NER in nucleosomes in vitro [51] Implicated in UV DNA damage response [87] |
| INO80 (13–16) [64] | INO80 (~6000) [68] | Ino80 | hIno80 | Nucleosome sliding [90] and eviction [91] | Enhances NER in yeast cells [60] Promotes NER in mammalian cells [95] |
| CSB (1) [103] | - | - | CSB | Nucleosome core remodeling, nucleosome repositioning [102] | Enhances TC-NER in mammalian cells [47,103] |
| Rad26 (1) [100] | Rad26 (<50) [68] | Rad26 | - | - | Stimulates TC-NER in yeast cells [100] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Czaja, W.; Mao, P.; Smerdon, M.J. The Emerging Roles of ATP-Dependent Chromatin Remodeling Enzymes in Nucleotide Excision Repair. Int. J. Mol. Sci. 2012, 13, 11954-11973. https://doi.org/10.3390/ijms130911954
Czaja W, Mao P, Smerdon MJ. The Emerging Roles of ATP-Dependent Chromatin Remodeling Enzymes in Nucleotide Excision Repair. International Journal of Molecular Sciences. 2012; 13(9):11954-11973. https://doi.org/10.3390/ijms130911954
Chicago/Turabian StyleCzaja, Wioletta, Peng Mao, and Michael J. Smerdon. 2012. "The Emerging Roles of ATP-Dependent Chromatin Remodeling Enzymes in Nucleotide Excision Repair" International Journal of Molecular Sciences 13, no. 9: 11954-11973. https://doi.org/10.3390/ijms130911954