Epigenetic Effects of Environmental Chemicals Bisphenol A and Phthalates


Abstract
:1. Introduction
2. Toxicities and Health Effects of Bisphenol A and Phthalates
3. Toxicogenomics of Bisphenol A and Phthalates
4. Epigenetic Effects of Bisphenol A and Phthalates
5. Conclusion and Remarks
Acknowledgement
- Conflict of InterestThe authors declare no competing financial interests.
Abbreviations
| BBP | butylbenzyl phthalate |
| BPA | bisphenol A |
| DBP | dibutyl phthalate |
| DEHP | diethylhexyl phthalate |
| EDCs | endocrine-disrupting chemicals |
| IPA | Ingenuity Pathways Analysis |
| MBP | monobutyl phthalate |
| MEHP | mono-(2-ethylhexyl)phthalate |
| PVC | polyvinyl chloride |
References
- Halden, R.U. Plastics and health risks. Annu. Rev. Public Health 2010, 31, 179–194. [Google Scholar]
- Heudorf, U.; Mersch-Sundermann, V.; Angerer, J. Phthalates: Toxicology and exposure. Int. J. Hyg. Environ. Health 2007, 210, 623–634. [Google Scholar]
- Wormuth, M.; Scheringer, M.; Vollenweider, M.; Hungerbühler, K. What are the sources of exposure to eight frequently used phthalic acid esters in Europeans? Risk Anal 2006, 26, 803–824. [Google Scholar]
- Thomas, J.A.; Thomas, M.J.; Gangolli, S.D. Biological effects of di-(2-ethylhexyl) phthalate and other phthalic acid esters. Crit. Rev. Toxicol 1984, 13, 283–317. [Google Scholar]
- Dolinoy, D.C.; Jirtle, R.L. Environmental epigenomics in human health and disease. Environ. Mol. Mutagen 2008, 49, 4–8. [Google Scholar]
- Anway, M.D.; Rekow, S.S.; Skinner, M.K. Transgenerational epigenetic programming of the embryonic testis transcriptome. Genomics 2008, 91, 30–40. [Google Scholar]
- Schwartz, D.; Collins, F. Environmental biology and human disease. Science 2007, 316, 695–696. [Google Scholar]
- Perera, F.; Herbstman, J. Prenatal environmental exposures, epigenetics, and disease. Reprod. Toxicol 2011, 31, 363–373. [Google Scholar]
- Liu, L.; Li, Y.; Tollefsbol, T.O. Gene-environment interactions and epigenetic basis of human diseases. Curr. Issues Mol. Biol 2008, 10, 25–36. [Google Scholar]
- Choudhuri, S.; Cui, Y.; Klaassen, C.D. Molecular targets of epigenetic regulation and effectors of environmental influences. Toxicol. Appl. Pharmacol 2010, 245, 378–393. [Google Scholar]
- Kundakovic, M.; Champagne, F.A. Epigenetic perspective on the developmental effects of bisphenol A. Brain Behav. Immun 2011, 25, 1084–1093. [Google Scholar]
- Baccarelli, A.; Bollati, V. Epigenetics and environmental chemicals. Curr. Opin. Pediatr 2009, 21, 243–251. [Google Scholar]
- Shelby, M.D. NTP-CERHR monograph on the potential human reproductive and developmental effects of bisphenol A. NTP CERHR MON 2008, 22. [Google Scholar]
- Talsness, C.E.; Andrade, A.J.M.; Kuriyama, S.N.; Taylor, J.A.; vom Saal, F.S. Components of plastic: Experimental studies in animals and relevance for human health. Phil. Trans. Biol. Sci 2009, 364, 2079–2096. [Google Scholar]
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. Mol. Biol 2011, 127, 27–34. [Google Scholar]
- Lang, I.A.; Galloway, T.S.; Scarlett, A.; Henley, W.E.; Depledge, M.; Wallace, R.B.; Melzer, D. Association of urinary bisphenol a concentration with medical disorders and laboratory abnormalities in adults. J. Am. Med. Assoc 2008, 300, 1303–1310. [Google Scholar]
- Welshons, W.V.; Nagel, S.C.; vom Saal, F.S. Large effects from small exposures. III. Endocrine mechanisms mediating effects of bisphenol A at levels of human exposure. Endocrinology 2006, 147, s56–s69. [Google Scholar]
- Golub, M.S.; Wu, K.L.; Kaufman, F.L.; Li, L.-H.; Moran-Messen, F.; Zeise, L.; Alexeeff, G.V.; Donald, J.M. Bisphenol A: Developmental toxicity from early prenatal exposurea. Birth. Defects Res. B Dev. Reprod. Toxicol 2010, 89, 441–466. [Google Scholar]
- Shelby, M.D. NTP-CERHR monograph on the potential human reproductive and developmental effects of di-(2-ethylhexyl) phthalate (DEHP). NTP CERHR MON 2006, 18. [Google Scholar]
- Swan, S.H.; Main, K.M.; Liu, F.; Stewart, S.L.; Kruse, R.L.; Calafat, A.M.; Mao, C.S.; Redmon, J.B.; Ternand, C.L.; Sullivan, S.; et al. Decrease in anogenital distance among male infants with prenatal phthalate exposure. Environ. Health Perspect 2005, 113, 1056–1061. [Google Scholar]
- Martino-Andrade, A.J.; Chahoud, I. Reproductive toxicity of phthalate esters. Mol. Nutr. Food Res 2010, 54, 148–157. [Google Scholar]
- Christen, V.; Crettaz, P.; Oberli-Schrämmli, A.; Fent, K. Antiandrogenic activity of phthalate mixtures: Validity of concentration addition. Toxicol. Appl. Pharmacol 2012, 259, 169–176. [Google Scholar]
- Genuis, S.J.; Beesoon, S.; Birkholz, D.; Lobo, R.A. Human excretion of bisphenol A: Blood, urine, and sweat (BUS) study. J. Environ. Public Health 2012, 2012, 185731. [Google Scholar]
- Davis, A.P.; Murphy, C.G.; Saraceni-Richards, C.A.; Rosenstein, M.C.; Wiegers, T.C.; Mattingly, C.J. Comparative toxicogenomics database: A knowledgebase and discovery tool for chemical-gene-disease networks. Nucleic Acids Res 2009, 37, D786–D792. [Google Scholar]
- Singh, S.; Li, S.S.-L. Phthalates: Toxicogenomics and inferred human diseases. Genomics 2011, 97, 148–157. [Google Scholar]
- Singh, S.; Li, S.S.-L. Bisphenol A and phthalates exhibit similar toxicogenomics and health effects. Gene 2012, 494, 85–91. [Google Scholar]
- Dietert, R.R.; DeWitt, J.C.; Germolec, D.R.; Zelikoff, J.T. Breaking patterns of environmentally influenced disease for health risk reduction: Immune perspectives. Environ. Health Perspect 2010, 118, 1091–1099. [Google Scholar]
- Morgan, H.D.; Sutherland, H.G.E.; Martin, D.I.K.; Whitelaw, E. Epigenetic inheritance at the agouti locus in the mouse. Nat. Genet 1999, 23, 314–318. [Google Scholar]
- Dolinoy, D.C.; Huang, D.; Jirtle, R.L. Maternal nutrient supplementation counteracts bisphenol A-induced DNA hypomethylation in early development. Proc. Natl. Acad. Sci. USA 2007, 104, 13056–13061. [Google Scholar]
- Ho, S.-M.; Tang, W.-Y.; Belmonte de Frausto, J.; Prins, G.S. Developmental exposure to estradiol and bisphenol a increases susceptibility to prostate carcinogenesis and epigenetically regulates phosphodiesterase type 4 variant 4. Cancer Res 2006, 66, 5624–5632. [Google Scholar]
- Prins, G.S.; Tang, W.-Y.; Belmonte, J.; Ho, S.-M. Developmental exposure to bisphenol a increases prostate cancer susceptibility in adult rats: Epigenetic mode of action is implicated. Fertil. Steril 2008, 89, e41. [Google Scholar]
- Tang, W.-Y.; Morey, L.M.; Cheung, Y.Y.; Birch, L.; Prins, G.S.; Ho, S.-M. Neonatal exposure to estradiol/bisphenol A alters promoter methylation and expression of Nsbp1 and Hpcal1 genes and transcriptional programs of Dnmt3a/b and Mbd2/4 in the rat prostate gland throughout life. Endocrinology 2012, 153, 42–55. [Google Scholar]
- Doshi, T.; Mehta, S.S.; Dighe, V.; Balasinor, N.; Vanage, G. Hypermethylation of estrogen receptor promoter region in adult testis of rats exposed neonatally to bisphenol A. Toxicology 2011, 289, 74–82. [Google Scholar]
- Bromer, J.G.; Zhou, Y.; Taylor, M.B.; Doherty, L.; Taylor, H.S. Bisphenol—A exposure in utero leads to epigenetic alterations in the developmental programming of uterine estrogen response. FASEB J 2010, 24, 2273–2280. [Google Scholar]
- Yaoi, T.; Itoh, K.; Nakamura, K.; Ogi, H.; Fujiwara, Y.; Fushiki, S. Genome-wide analysis of epigenomic alterations in fetal mouse forebrain after exposure to low doses of bisphenol A. Biochem. Biophys. Res. Comm 2008, 376, 563–567. [Google Scholar]
- Weng, Y.-I.; Hsu, P.-Y.; Liyanarachchi, S.; Liu, J.; Deatherage, D.E.; Huang, Y.-W.; Zuo, T.; Rodriguez, B.; Lin, C.-H.; Cheng, A.-L.; et al. Epigenetic influences of low-dose bisphenol A in primary human breast epithelial cells. Toxicol. Appl. Pharmacol 2010, 248, 111–121. [Google Scholar]
- Doherty, L.; Bromer, J.; Zhou, Y.; Aldad, T.; Taylor, H. In utero exposure to diethylstilbestrol (DES) or bisphenol-A (BPA) increases EZH2 expression in the mammary gland: An epigenetic mechanism linking endocrine disruptors to breast cancer. Horm. Cancer 2010, 1, 146–155. [Google Scholar]
- Vire, E.; Brenner, C.; Deplus, R.; Blanchon, L.; Fraga, M.; Didelot, C.; Morey, L.; van Eynde, A.; Bernard, D.; Vanderwinden, J.-M.; et al. The polycomb group protein EZH2 directly controls DNA methylation. Nature 2006, 439, 871–874. [Google Scholar]
- Avissar-Whiting, M.; Veiga, K.R.; Uhl, K.M.; Maccani, M.A.; Gagne, L.A.; Moen, E.L.; Marsit, C.J. Bisphenol A exposure leads to specific microRNA alterations in placental cells. Reprod. Toxicol 2010, 29, 401–406. [Google Scholar]
- Cho, H.; Kim, S.; Park, H.-W.; Oh, M.-J.; Yu, S.; Lee, S.; Park, C.; Han, J.; Oh, J.-H.; Hwang, S.; et al. A relationship between miRNA and gene expression in the mouse sertoli cell line after exposure to bisphenol A. BioChip J 2010, 4, 75–81. [Google Scholar]
- Chan Kang, S.; Mu Lee, B. DNA methylation of estrogen receptor α gene by phthalates. J. Toxicol. Environ. Health A 2005, 68, 1995–2003. [Google Scholar]
- Wu, S.; Zhu, J.; Li, Y.; Lin, T.; Gan, L.; Yuan, X.; Xu, M.; Wei, G. Dynamic effect of di-2-(ethylhexyl) phthalate on testicular toxicity: Epigenetic changes and their impact on gene expression. Int. J. Toxicol 2010, 29, 193–200. [Google Scholar]
- Kundakovic, M.; Champagne, F.A. Epigenetic perspective on the developmental effects of bisphenol A. Brain Behav. Immun 2011, 25, 1084–1093. [Google Scholar]
- Manikkam, M.; Guerrero-Bosagna, C.; Tracey, R.; Haque, M.M.; Skinner, M.K. Transgenerational actions of environmental compounds on reproductive disease and identification of epigenetic biomarkers of ancestral exposures. PLoS One 2012, 7, e31901. [Google Scholar]
- Li, S.S.-L.; Liu, Y.H.; Tseng, C.N.; Chung, T.L.; Lee, T.Y.; Singh, S. Characterization and gene expression profiling of five new human embryonic stem cell lines derived in Taiwan. Stem Cells Dev 2006, 15, 532–555. [Google Scholar]
- Chen, B.Z.; Yu, S.L.; Singh, S.; Kao, L.P.; Tsai, Z.Y.; Yang, P.C.; Chen, B.H.; Li, S.S.-L. Identification of microRNAs expressed highly in pancreatic islet-like cell clusters differentiated from human embryonic stem cells. Cell Biol. Int 2011, 35, 29–37. [Google Scholar]
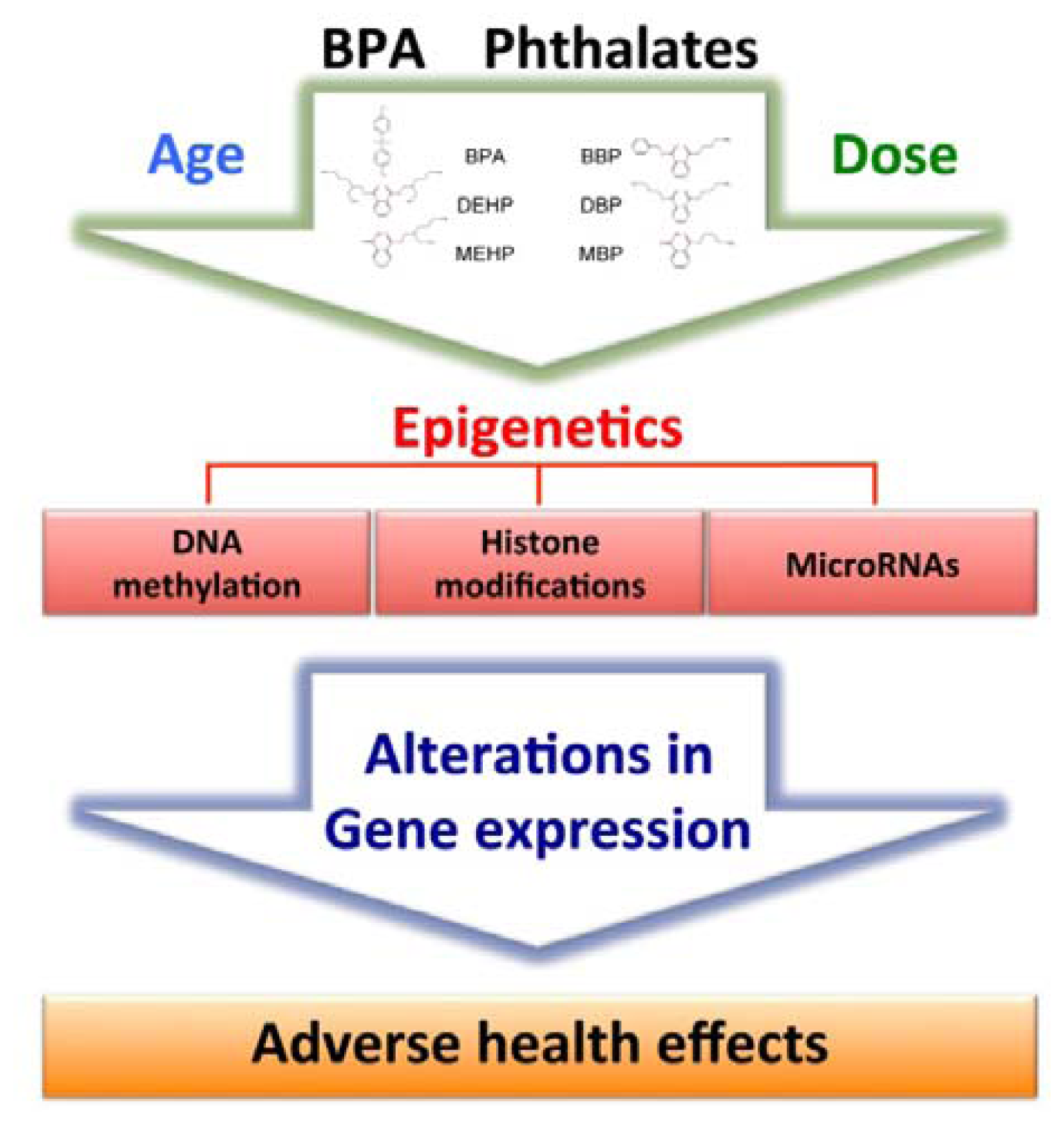

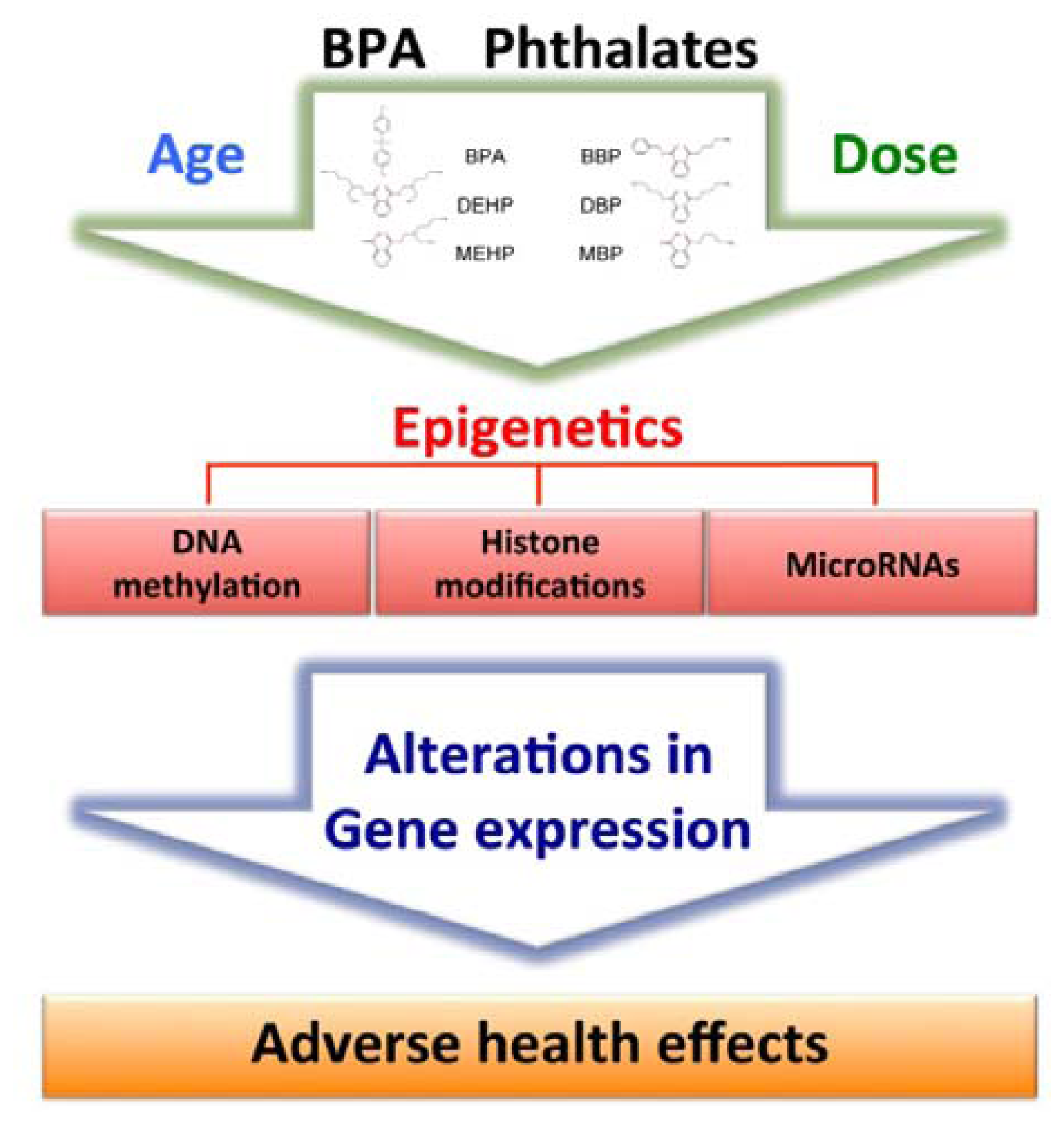{kind=link}
| Chemicals | Epignetic alterations | Genes | Organisms | References |
|---|---|---|---|---|
| BPA | DNA hypomethylation | Agouti | Mouse | [28,29] |
| BPA | DNA hypomethylation | Pde4d4 | Rat | [30,31] |
| BPA | DNA hypomethylation | Nsbp1 | Rat | [32] |
| DNA hypermethylation | Hpcal1 | |||
| BPA | DNA hypermethylaton | ESR1&2 | Rat | [33] |
| BPA | DNA hypomethylation | Hoxa10 | Mouse | [34] |
| BPA | DNA hypomethylation or hypermethylation | 13 genes | Mouse | [35] |
| BPA | DNA hypermethylation | LAMP3 | Human | [36] |
| BPA | Histone modification | H3K27me3 | Human Mouse | [37,38] |
| BPA | Induction of microRNA | miR-146a | Human | [39] |
| BPA | Up/down-regulated miRNAs | 37 miRNAs | Mouse | [40] |
| Phthalate BBP | DNA hypomethylation | ESR1 | Human | [41] |
| Phthalate DEHP | DNA hypermethylation | INSL3 | Mouse | [42] |
| Mixture of BPA & phthalates | Differential DNA methylation Transgeneration (F3) | Rat | [43] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Singh, S.; Li, S.S.-L. Epigenetic Effects of Environmental Chemicals Bisphenol A and Phthalates. Int. J. Mol. Sci. 2012, 13, 10143-10153. https://doi.org/10.3390/ijms130810143
Singh S, Li SS-L. Epigenetic Effects of Environmental Chemicals Bisphenol A and Phthalates. International Journal of Molecular Sciences. 2012; 13(8):10143-10153. https://doi.org/10.3390/ijms130810143
Chicago/Turabian StyleSingh, Sher, and Steven Shoei-Lung Li. 2012. "Epigenetic Effects of Environmental Chemicals Bisphenol A and Phthalates" International Journal of Molecular Sciences 13, no. 8: 10143-10153. https://doi.org/10.3390/ijms130810143
APA StyleSingh, S., & Li, S. S.-L. (2012). Epigenetic Effects of Environmental Chemicals Bisphenol A and Phthalates. International Journal of Molecular Sciences, 13(8), 10143-10153. https://doi.org/10.3390/ijms130810143