Fraction from Wax Apple [Syzygium samarangense (Blume) Merrill and Perry] Fruit Extract Ameliorates Insulin Resistance via Modulating Insulin Signaling and Inflammation Pathway in Tumor Necrosis Factor α-Treated FL83B Mouse Hepatocytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
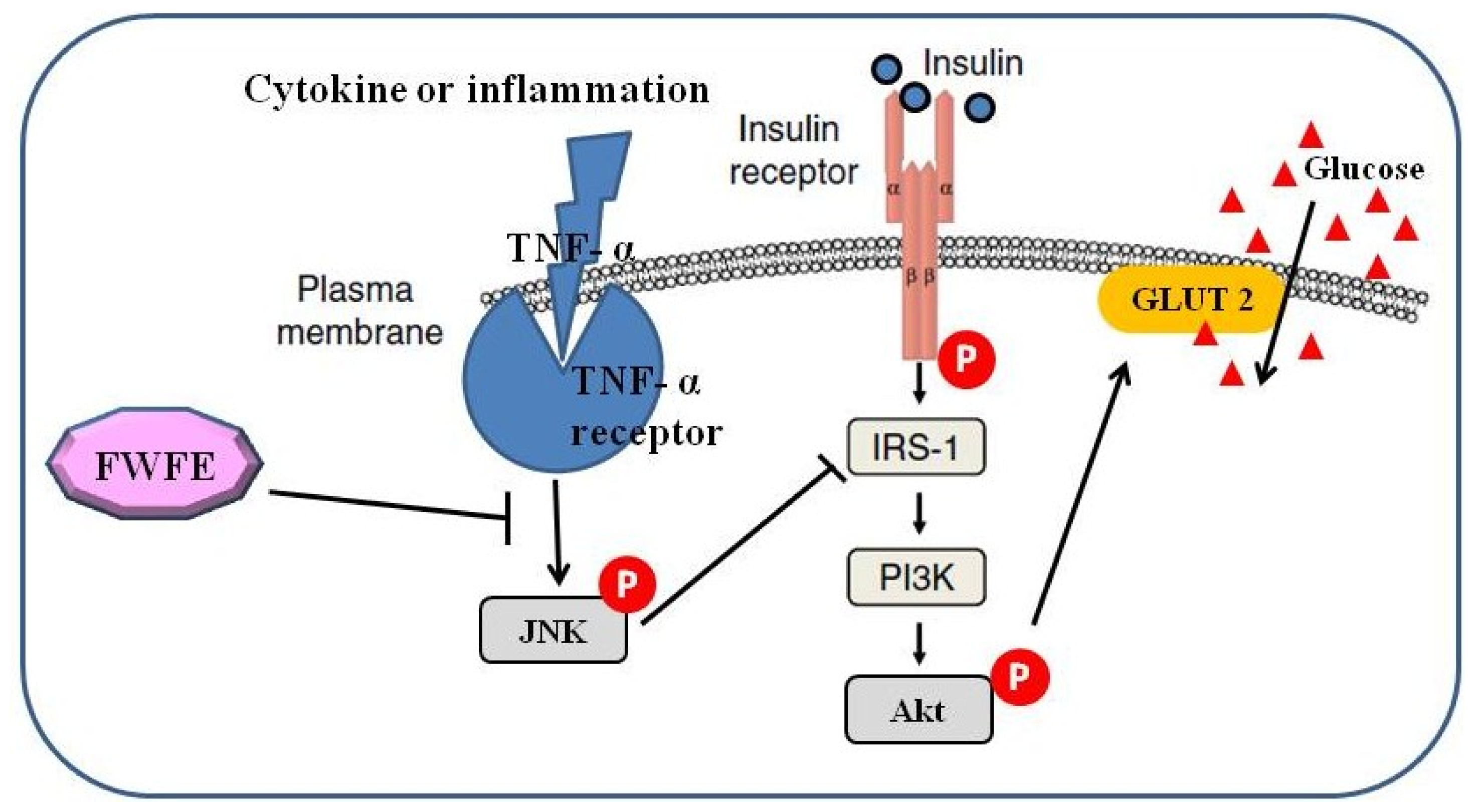
:1. Introduction
2. Results and Discussion
2.1. The Characterization of FWFE
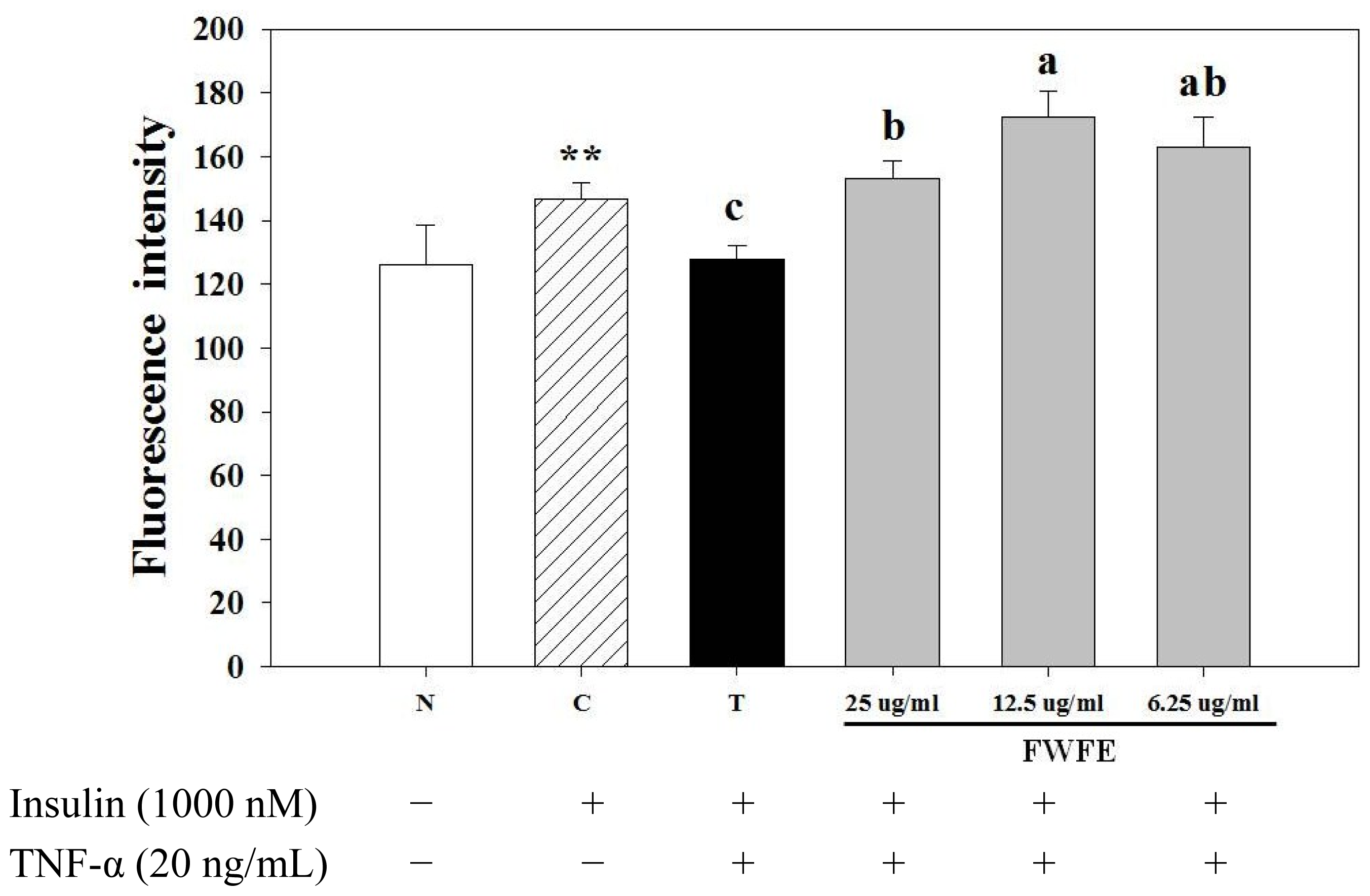
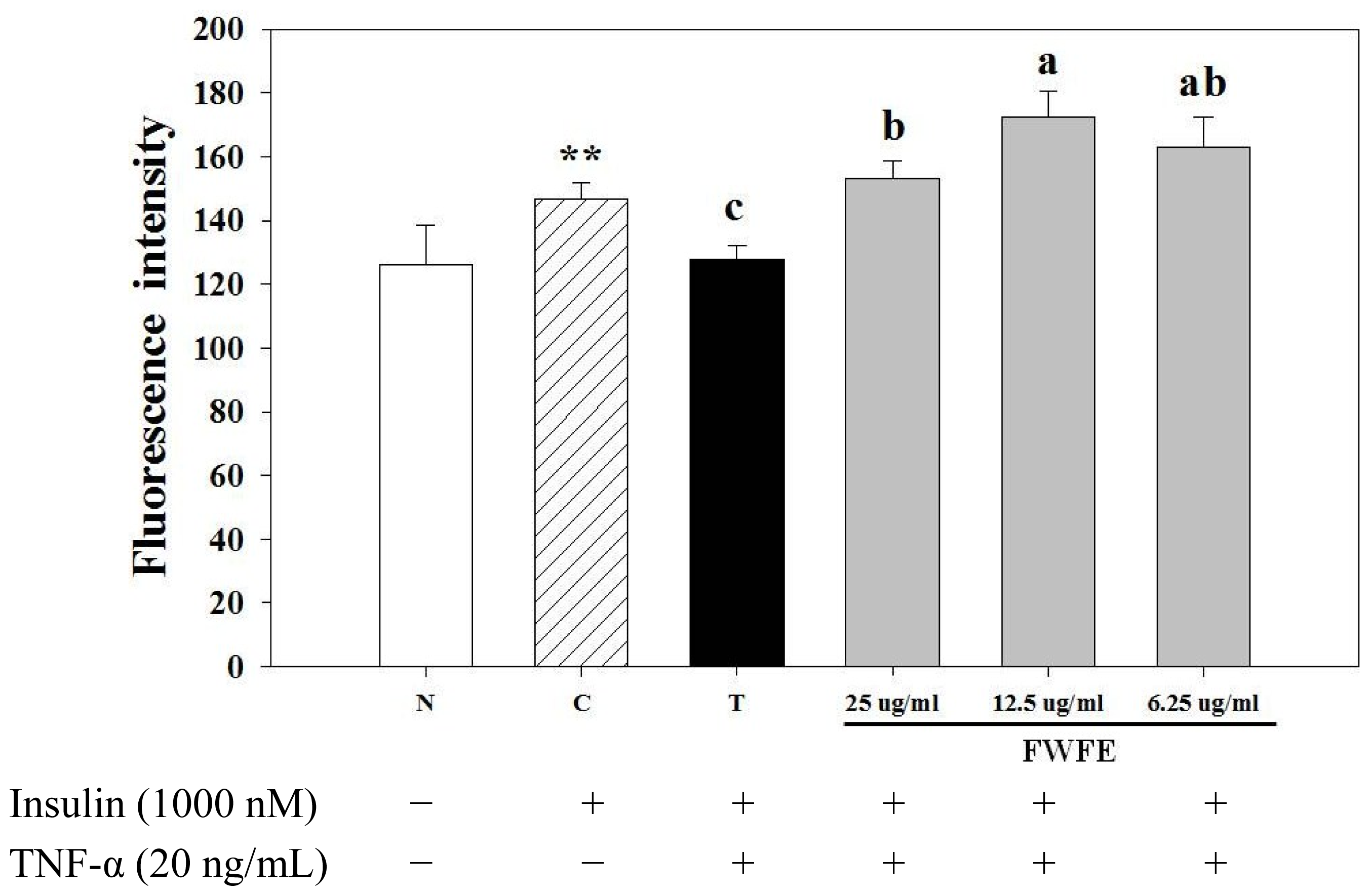
2.2. Effects of FWFE on Glucose Uptake in Insulin Resistant FL83B Mouse Hepatocytes
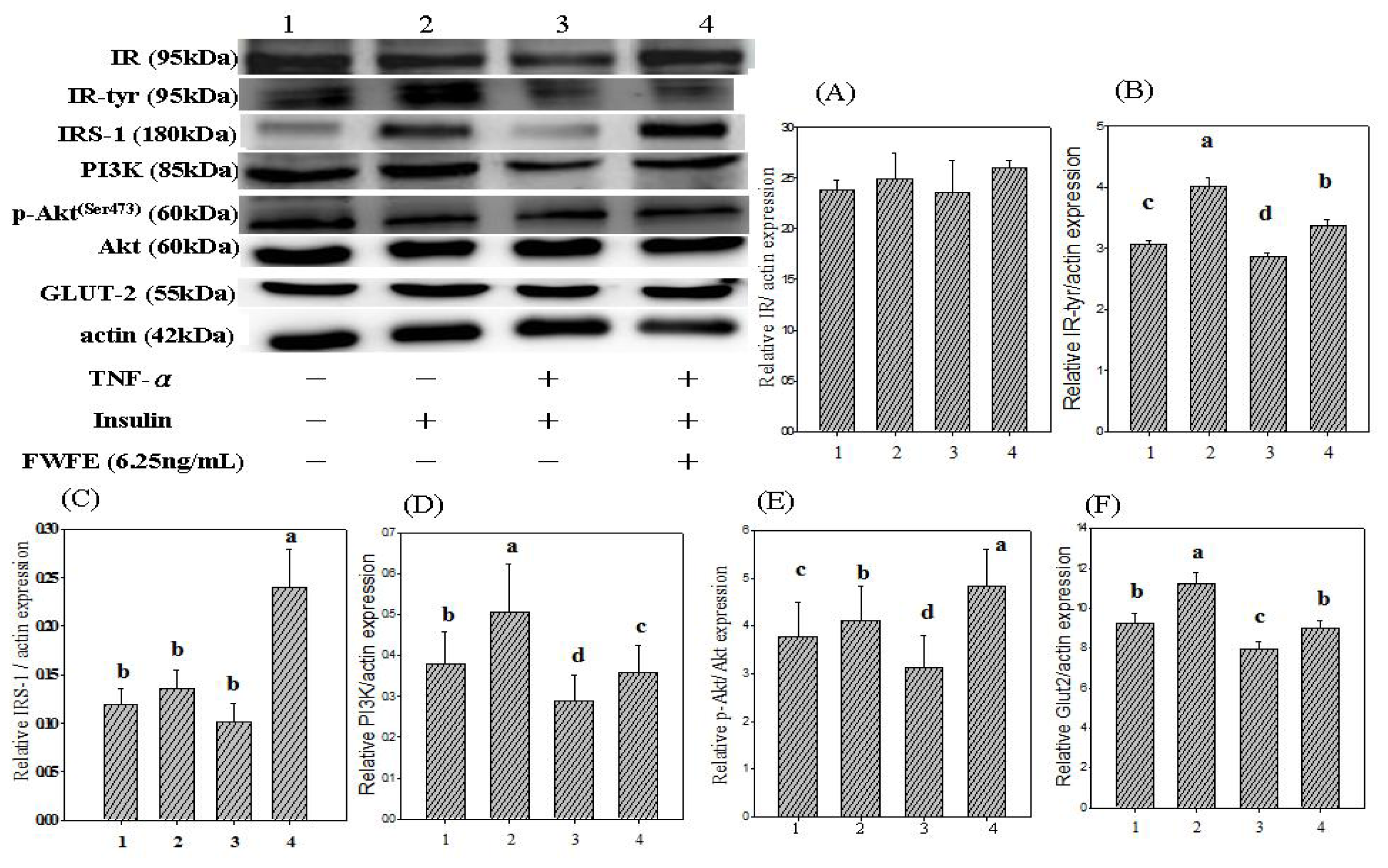
2.3. Insulin Receptor Expression and IR Tyrosyl Phosphorylation
2.4. Insulin Receptor Substrate Expression
2.5. Phosphatidylinositol-3 Kinase Expression
2.6. Expression of Akt/Protein Kinase B
2.7. Glucose Transporter 2 Expression
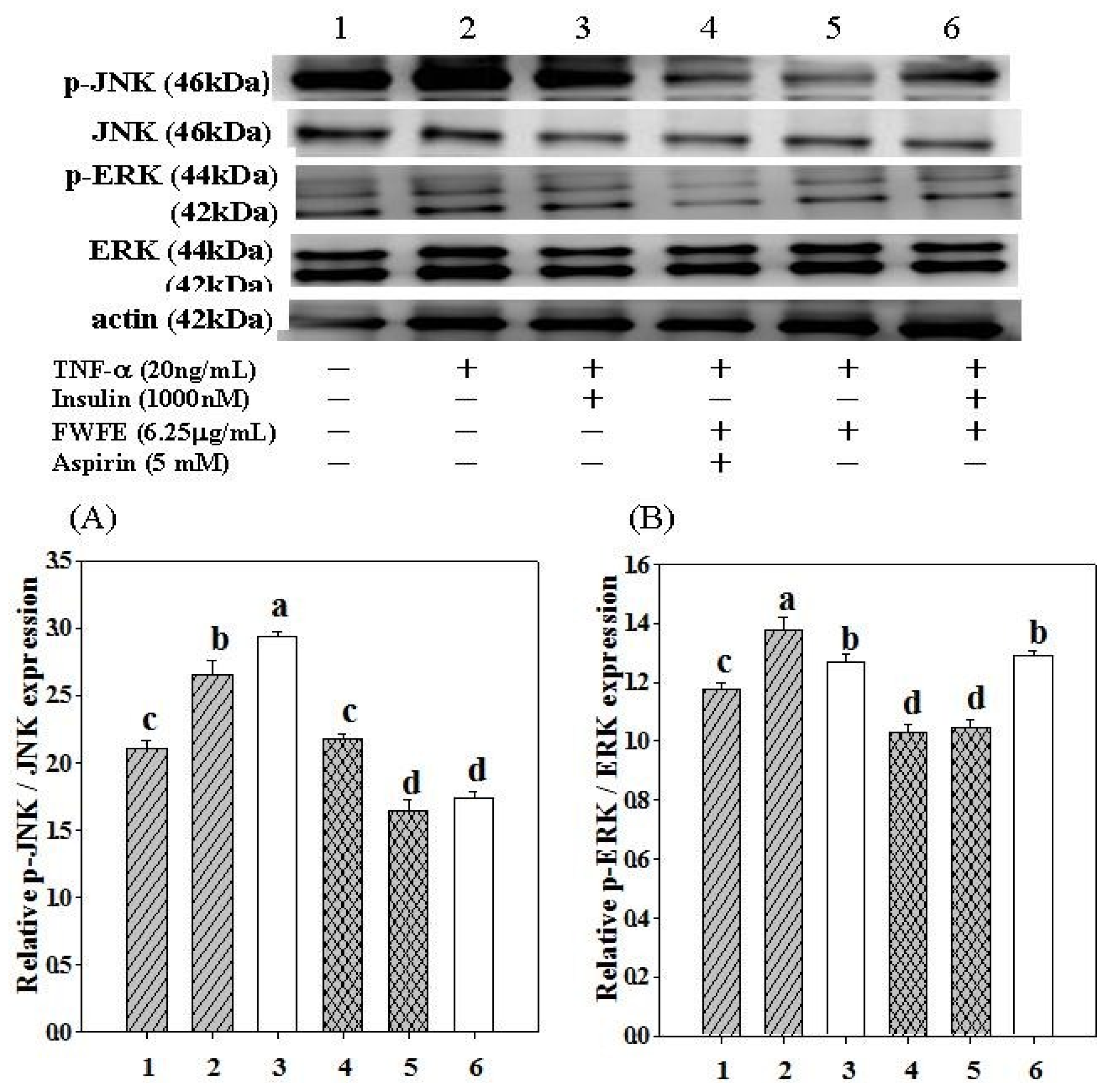
2.8. Expression of c-Jun N-terminal Kinases
2.9. Expression of Extracellular Signal-Regulated Kinases
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Wax Apple Fruits
3.3. Preparation of Extract
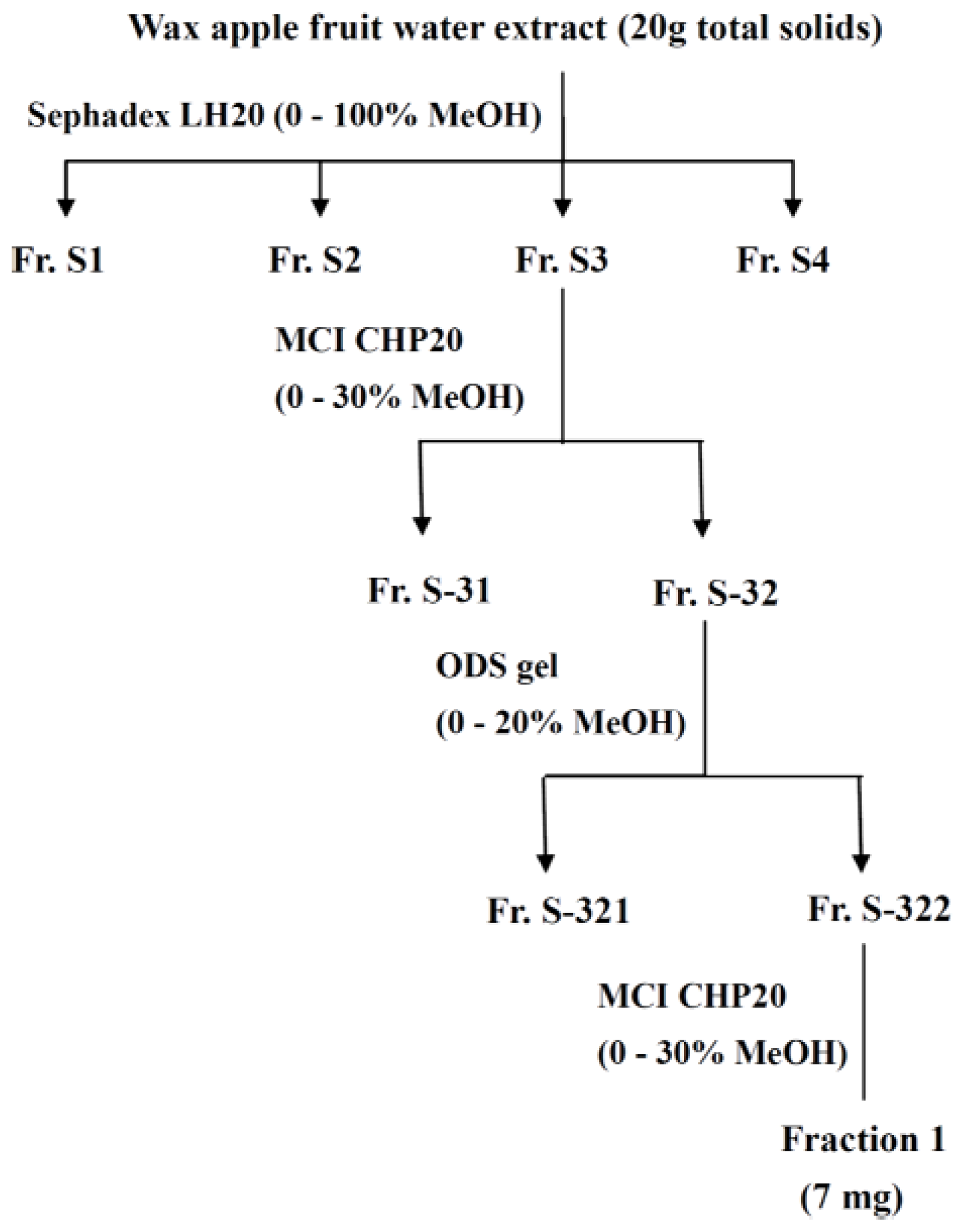
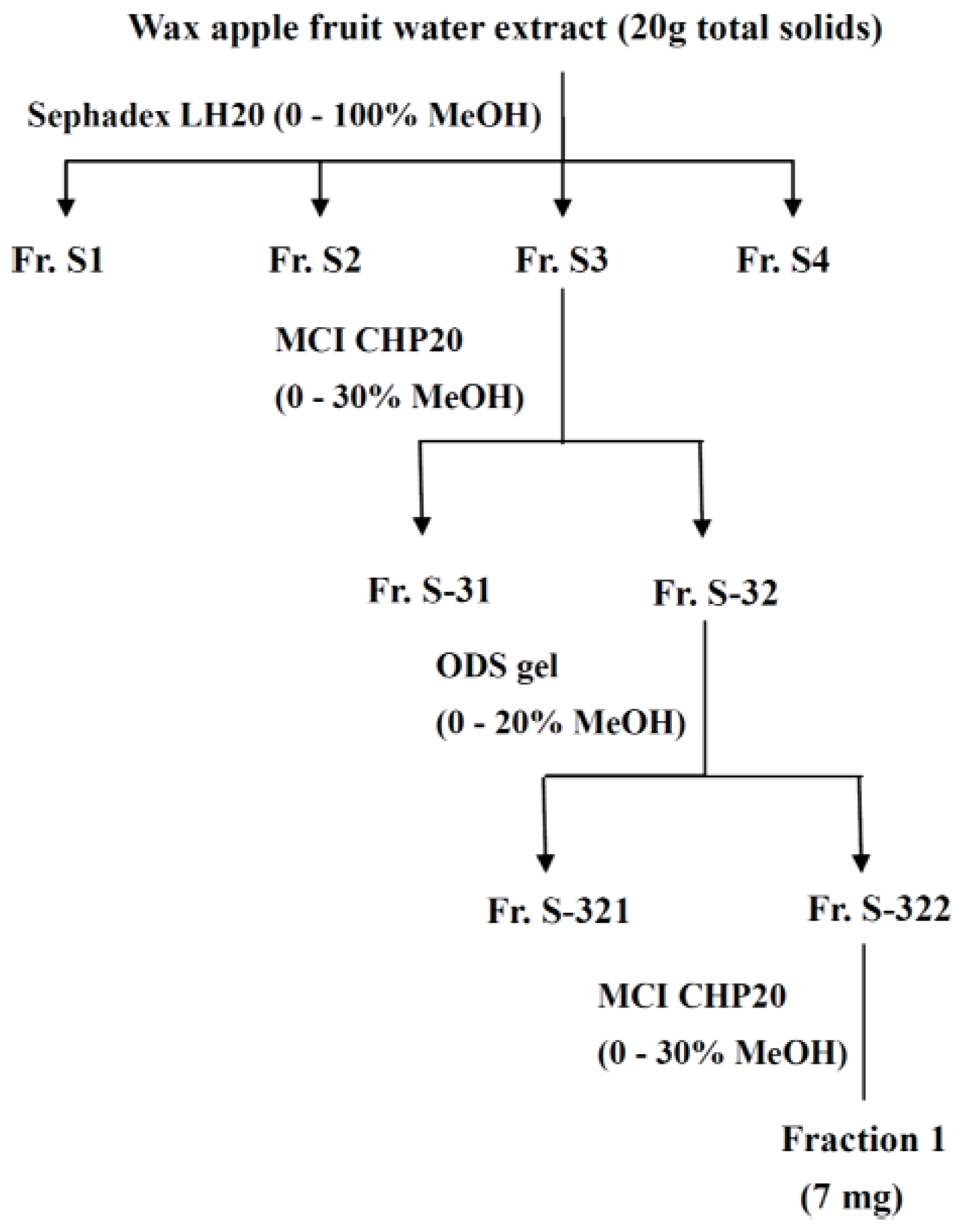
3.4. Fractionation of the Extract
3.5. Determination of Phenolics and Flavonoids Content
3.6. Cell Culture
3.7. Induction of Insulin Resistance Using TNF-α
3.8. Cell Preparation
3.9. Uptake of Fluorescent 2-NBDG in Mouse FL83B Hepatocytes
3.10. Protein Extraction from Cells
3.11. Determination of Protein Concentration
3.12. Western Blot Analysis
3.13. Statistical Analysis
4. Conclusions
Acknowledgments
Abbreviations
| Akt/PKB | protein kinase B |
| BSA | bovine serum albumin |
| DM | diabetes mellitus |
| ERK | extracellular signal-regulated kinases |
| FBS | fetal bovine serum |
| F12K | F12 Ham Kaighn’s modification |
| FWFE | fraction from wax apple fruit extract |
| GLUT2 | glucose transporter 2 |
| HEPES | 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid |
| IGF-IR | insulin-like growth factor-I receptor |
| IR | insulin receptor |
| IRS-1 | insulin receptor substrate-1 |
| IR-tyr | tyrosyl-phosphorylated insulin receptor |
| JNK | c-Jun N-terminal kinases |
| MAPK | Mitogen-activated protein kinase |
| 2-NBDG | 2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl) amino]-2-deoxy-D-glucose |
| p-Akt/p-PKB | phosphorylated protein kinase B |
| PBS | phosphate buffered saline |
| p-ERK | phosphorylated ERK |
| PI3K | phosphatidylinositol-3 kinase |
| p-JNK | phosphorylated JNK |
| SDS-PAGE | dodecyl sulfate polyacrylamide gel electrophoresis |
| TLC | thin layer chromatography |
| TNF-α | tumor necrosis factor-alpha |
References
- World Health Organization. 2012. Available online: http://www.who.int/diabetes/en/index.html accessed on 17 May 2012.
- Ferrannini, E. Insulin resistance versus insulin deficiency in non-insulin-dependent diabetes mellitus: Problems and prospects. Endocr. Rev 1998, 19, 477–490. [Google Scholar]
- Saltiel, A.R.; Kahn, C.R. Insulin signaling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar]
- Coletta, D.K.; Mandarino, L.J. Mitochondrial dysfunction and insulin resistance from the outside in: Extracellular matrix, the cytoskeleton, and mitochondria. Am. J. Physiol. Endocrinol. Metab 2011, 301, E749–E755. [Google Scholar]
- Hotamisligil, G.S. Inflammatory pathways and insulin action. Int. J. Obes. Relat. Metab. Disord 2003, 27, S53–S55. [Google Scholar]
- Nilsson-Öhman, J.; Fredrikson, G.N.; Nilsson-Berglund, L.M.; Gustavsson, C.; Bengtsson, E.; Smith, M.; Agardh, C.; Agardh, E.; Jovinge, S.; Gomez, M.F.; et al. Tumor necrosis factor-α does not mediate diabetes-induced vascular inflammation in mice. Arterioscler Thromb. Vasc. Biol 2009, 29, 1465–1470. [Google Scholar]
- Gurib-Fakim, A. Phytochemical screening of 38 mauritian medicinal plants. Revue Agricole et Sucrière de l’lle Maurice 1991, 69, 42–50. [Google Scholar]
- Neergheen, V.; Soobrattee, M.; Bahorun, T.; Aruoma, O. Characterization of the phenolic constituents in Mauritian endemic plants as determinants of their antioxidant activities in vitro. J. Plant Physiol 2006, 163, 787–799. [Google Scholar]
- Shen, S.C.; Cheng, F.C.; Wu, N.J. Effect of guava (Psidium guajava Linn.) leaf soluble solids on glucose metabolism in type 2 diabetic rats. Phytother. Res 2008, 22, 1458–1464. [Google Scholar]
- Cheng, F.C.; Shen, S.C.; Wu, J.S.B. Effect of guava (Psidium guajava L.) leaf extract on glucose uptake in rat hepatocytes. J. Food Sci 2009, 74, H132–H138. [Google Scholar]
- Nonaka, G.; Aiko, Y.; Aritake, K.; Nishioka, I. Tannins and related compounds. CXIX. Samarangenins a and b, novel proanthocyanidins with doubly bonded structures, from Syzygium samarangens and S. aqueum. Chem. Pharm. Bull 1992, 40, 2671–2673. [Google Scholar]
- Kuo, Y.C.; Yang, L.M.; Lin, L.C. Isolation and inmunomodulatory effects of flavonoids from Syzygium samarangense. Planta Med 2004, 70, 1237–1239. [Google Scholar]
- Nair, A.; Krishnan, S.; Ravikrishna, C.; Madhusudanan, K. New and rare flavonol glycosides from leaves of Syzygium samarangense. Fitoterapia 1999, 70, 148–151. [Google Scholar]
- Srivastava, R.; Shaw, A.; Kulshreshtha, D. Triterpenoids and chalcone from Syzygium samarangense. Phytochemistry 1995, 38, 687–689. [Google Scholar]
- Resurreccion-Magno, M.; Villasenor, I.; Harada, N.; Monde, K. Antihyperglycaemic flavonoids from Syzygium samarangense (Blume) Merr. and Perry. Phytother. Res 2005, 19, 246–251. [Google Scholar]
- Wong, K.; Lai, F. Volatile constituents from the fruits of four Syzygium species grown in Malaysia. Flavour Fragr. J 1996, 11, 61–66. [Google Scholar]
- Tsao, R.; Deng, Z. Separation procedures for naturally occurring antioxidant phytochemicals. J. Chromatogr. B 2004, 812, 85–99. [Google Scholar]
- Peraldi, P.; Xu, M.; Spiegelman, B.M. Thiazolidinediones block tumor necrosis factor-α-induced inhibition of insulin singaling. J. Clin. Invest 1996, 100, 1863–1869. [Google Scholar]
- Moller, D.E. Potential role of TNF-R in the pathogenesis of insulin resistance and type 2 diabetes. Trends Endocrinol. Metab 2000, 6, 212–217. [Google Scholar]
- Cheung, A.T.; Wang, J.; Ree, D.; Kolls, J.K.; Bryer-Ash, M. Tumor necrosis factor-R induces hepatic insulin resistance in obese zucker (fa/fa) rats via interaction of leukocyte antigen-related tyrosine phosphatase with focal adhesion kinase. Diabetes 2000, 49, 810–819. [Google Scholar]
- Jung, E.H.; Kim, S.R.; Hwang, I.K.; Ha, T.Y. Hypoglycemic effects of a phenolic acid fraction of rice bran and ferulic acid in C57BL/KsJ-db/db mice. J. Agric. Food Chem 2007, 55, 9800–9804. [Google Scholar]
- Czinner, E.; Hagymasi, K.; Blazovics, A.; Kery, A.; Szoke, E.; Lemberkovics, E. In vitro antioxidant properties of Helichrysum arenarium (L.) Moench. J. Ethnopharmacol 2000, 73, 437–443. [Google Scholar]
- Bjornholm, M.; Zierath, J.R. Insulin signal transduction in human skeletal muscle: Identifying the defects in type II diabetes. Biochem. Soc. Trans 2005, 33, 354–357. [Google Scholar]
- Zick, Y. Insulin resistance: A phosphorylation-based uncoupling of insulin signaling. Trends Cell Biol 2001, 11, 437–441. [Google Scholar]
- White, M.F. Insulin signaling in health and disease. Science 2003, 302, 1710–1711. [Google Scholar]
- Shahid, G.; Hussain, T. GRK2 negatively regulates glycogen synthesis in mouse liver FL83B cells. J. Biol. Chem 2007, 282, 20612–20620. [Google Scholar]
- Rui, L.; Aguirre, V.; Kim, J.K.; Shulman, G.I.; Lee, A.; Corbould, A.; Dunaif, A.; White, M.F. Insulin/IGF-1 and TNF-α stimulate phosphorylation of IRS-1 at inhibitory Ser307 via distinct pathways. J. Clin. Invest 2001, 107, 181–189. [Google Scholar]
- Dearth, R.K.; Cui, X.; Kim, H.J.; Hadsell, D.L.; Lee, A.V. Oncogenic transformation by the signaling adaptor proteins insulin receptor substrate (IRS)-1 and IRS-2. Cell Cycle 2007, 6, 705–713. [Google Scholar]
- Yenush, L.; White, M.F. The IRS-signalling system during insulin and cytokine action. Bioessays 1997, 19, 491–500. [Google Scholar]
- Moeschel, K.; Beck, A.; Weigert, C.; Lammers, R.; Kalbacher, H.; Voelter, W.; Schleicher, E.D.; Häring, H.; Lehmann, R. Protein kinase C-δ-induced phosphorylation of Ser318 in insulin receptor substrate-1 (IRS-1) attenuates the interaction with the insulin receptor and the tyrosine phosphorylation of IRS-1. J. Biol. Chem 2004, 279, 25157–25163. [Google Scholar]
- Benito, M. Tissue specificity on insulin action and resistance: Past to recent mechanism. Acta Physiol 2011, 201, 297–312. [Google Scholar]
- Hotamisligil, G.S.; Peraldi, P.; Budvari, A.; Ellis, R.W.; White, M.F. Spiegelman BM: IRS-1 mediated inhibition of insulin receptor tyrosine kinase activity in TNF-α and obesity-induced insulin resistance. Science 1996, 271, 665–668. [Google Scholar]
- Czech, M.P.; Corvera, S. Signaling mechanisms that regulate glucose transport. J. Biol. Chem 1999, 74, 1865–1868. [Google Scholar]
- Nikoulina, S.E.; Ciaraldi, T.P.; Carter, L.; Mudaliar, S.; Park, K.S.; Henry, R.R. Impaired muscle glycogen synthase in type 2 diabetes is associated with diminished phosphatidylinositol 3-kinase activation. J. Clin. Endocrinol. Metab 2001, 86, 4307–4314. [Google Scholar]
- Cichy, S.B.; Uddin, S.; Danilkovich, A.; Guo, S.; Klippel, A.; Unterman, T.G. Protein kinase B/Akt mediates effect of insulin on hepatic insulin-like growth factor-binding protein-1 gene expression through a conserved insulin response sequence. J. Biol. Chem 1998, 273, 6482–6487. [Google Scholar]
- Maleppillil, V.V.; Sandeep, S.; Rishi, R.C. The hypoglycaemic activity of fenugreek seed extract is mediated through the stimulation of an insulin signaling pathway. Br. J. Pharm 2005, 146, 41–48. [Google Scholar]
- Zhou, L.; Sell, H.; Eckardt, K.; Yang, Z.; Eckel, J. Conditioned medium obtained from in vitro differentiated adipocytes and resistin induce insulin resistance in human hepatocytes. FEBS Lett 2007, 581, 4303–4308. [Google Scholar]
- Guitton, C.; Cottereau, A.; Gérard, N.; Quillard, T.; Chauveau, A.; Devallière, J.; Tonnerre, P.; Charreau, B. Protective crosstalk between activated protein C and TNF signaling in vascular endothelial cells: Implication of EPCR, non canonical NFκB and ERK1/2 MAPKinases. J. Physiol. Cell Physiol 2011, 300, C833–C842. [Google Scholar]
- Joost, H.G.; Bell, G.I.; Best, J.D.; Birnbaum, M.J.; Charron, M.J.; Chen, Y.T.; Doege, H.; James, D.E.; Lodish, H.F.; Moley, K.H. Nomenclature of the GLUT/SLC2A family of sugar/polyol transport facilitators. Am. J. Physiol 2002, 282, E974–E976. [Google Scholar]
- Thorens, B.; Cheng, Z.Q.; Brown, D.; Lodish, H.F. Liver glucose transporter: A basolateral protein in hepatocytes and intestine and kidney cells. Am. J. Physiol 1990, 259, C279–C285. [Google Scholar]
- James, D.E. The mammalian facilitative glucose transporter family. Int. Union Physiol. Sci 1995, 10, 67–71. [Google Scholar]
- Luca, C.D.; Olefsky, J.M. Inflammation and insulin resistance. FEBS Lett 2008, 582, 97–105. [Google Scholar]
- Chang, L.; Karin, M. Mammalian MAP kinase signaling cascades. Nature 2001, 410, 37–40. [Google Scholar]
- Barreiro, G.C.; Prattali, R.R.; Caliseo, C.T.; Fugiwara, F.Y.; Ueno, M.; Prada, P.O.; Velloso, L.A.; Saad, M.J.A.; Carvalheira, J.B.C. Aspirin inhibits serine phosphorylation of IRS-1 in muscle and adipose tissue of septic rats. Biochem. Biophys. Res. Commun 2004, 320, 992–997. [Google Scholar]
- Li, R.W.; Theriault, A.G.; Au, K.; Douglas, T.D.; Casaschi, A.; Kurowska, E.M.; Mukherjee, R. Citrus polymethoxylated flavones improve lipid and glucose homeostasis and modulate adipocytokines in fructose-induced insulin resistant hamsters. Life Sci 2006, 79, 365–373. [Google Scholar]
- Chang, W.C.; Shen, S.C. Effect of water extracts from edible Myrtaceae plants on uptake of 2-(N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)-2-deoxyglucose in TNF-α-treated FL83B mouse hepatocytes. Phytother. Res 2012. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin Ciocalteu agent. Methods Enzymol 1999, 299, 152–178. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid content in mulberry and their scavenging effects on superoxide radicals. Food Chem 1999, 64, 555–559. [Google Scholar]



© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shen, S.-C.; Chang, W.-C.; Chang, C.-L. Fraction from Wax Apple [Syzygium samarangense (Blume) Merrill and Perry] Fruit Extract Ameliorates Insulin Resistance via Modulating Insulin Signaling and Inflammation Pathway in Tumor Necrosis Factor α-Treated FL83B Mouse Hepatocytes. Int. J. Mol. Sci. 2012, 13, 8562-8577. https://doi.org/10.3390/ijms13078562
Shen S-C, Chang W-C, Chang C-L. Fraction from Wax Apple [Syzygium samarangense (Blume) Merrill and Perry] Fruit Extract Ameliorates Insulin Resistance via Modulating Insulin Signaling and Inflammation Pathway in Tumor Necrosis Factor α-Treated FL83B Mouse Hepatocytes. International Journal of Molecular Sciences. 2012; 13(7):8562-8577. https://doi.org/10.3390/ijms13078562
Chicago/Turabian StyleShen, Szu-Chuan, Wen-Chang Chang, and Chiao-Li Chang. 2012. "Fraction from Wax Apple [Syzygium samarangense (Blume) Merrill and Perry] Fruit Extract Ameliorates Insulin Resistance via Modulating Insulin Signaling and Inflammation Pathway in Tumor Necrosis Factor α-Treated FL83B Mouse Hepatocytes" International Journal of Molecular Sciences 13, no. 7: 8562-8577. https://doi.org/10.3390/ijms13078562