Molecular Targets of TRAIL-Sensitizing Agents in Colorectal Cancer



Abstract
:1. Introduction
2. TRAIL Signaling Pathway and Mechanisms of TRAIL Resistance
3. TRAIL-Sensitizing Agents in CRC
3.1. Chemotherapeutics–DNA Damage Agents
3.2. Natural Products
3.2.1. Polyphenols and Related Compounds
3.2.2. Terpenoids
3.2.3. Other Natural Products
3.3. Other TRAIL-Sensitizing Agents
3.3.1. Proteasome Inhibitors
3.3.2. ER Stress Inducers
3.3.3. Molecules Controlling p53, AMP-Activated Protein Kinase, Protein Kinase C and the Molecular Chaperone Hsp90
4. Conclusions
Acknowledgements
- Conflict of InterestThe authors declare no conflict of interest.
References
- Johnstone, R.W.; Ruefli, A.A.; Lowe, S.W. Apoptosis: A link between cancer genetics and chemotherapy. Cell 2002, 108, 153–164. [Google Scholar]
- Wiley, S.R.; Schooley, K.; Smolak, P.J.; Din, W.S.; Huang, C.P.; Nicholl, J.K.; Sutherland, G.R.; Smith, T.D.; Rauch, C.; Smith, C.A.; et al. Identification and characterization of a new member of the tnf family that induces apoptosis. Immunity 1995, 3, 673–682. [Google Scholar]
- Almasan, A.; Ashkenazi, A. Apo2l/trail: Apoptosis signaling, biology, and potential for cancer therapy. Cytokine Growth Factor Rev 2003, 14, 337–348. [Google Scholar]
- Ashkenazi, A.; Pai, R.C.; Fong, S.; Leung, S.; Lawrence, D.A.; Marsters, S.A.; Blackie, C.; Chang, L.; McMurtrey, A.E.; Hebert, A.; et al. Safety and antitumor activity of recombinant soluble apo2 ligand. J. Clin. Invest 1999, 104, 155–162. [Google Scholar]
- Walczak, H.; Miller, R.E.; Ariail, K.; Gliniak, B.; Griffith, T.S.; Kubin, M.; Chin, W.; Jones, J.; Woodward, A.; Le, T.; et al. Tumoricidal activity of tumor necrosis factor-related apoptosis-inducing ligand in vivo. Nat. Med 1999, 5, 157–163. [Google Scholar]
- Hopkins-Donaldson, S.; Bodmer, J.L.; Bourloud, K.B.; Brognara, C.B.; Tschopp, J.; Gross, N. Loss of caspase-8 expression in highly malignant human neuroblastoma cells correlates with resistance to tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis. Cancer Res 2000, 60, 4315–4319. [Google Scholar]
- Lee, S.H.; Shin, M.S.; Kim, H.S.; Lee, H.K.; Park, W.S.; Kim, S.Y.; Lee, J.H.; Han, S.Y.; Park, J.Y.; Oh, R.R.; et al. Somatic mutations of trail-receptor 1 and trail-receptor 2 genes in non-hodgkin’s lymphoma. Oncogene 2001, 20, 399–403. [Google Scholar]
- Lee, S.H.; Shin, M.S.; Kim, H.S.; Lee, H.K.; Park, W.S.; Kim, S.Y.; Lee, J.H.; Han, S.Y.; Park, J.Y.; Oh, R.R.; et al. Alterations of the dr5/trail receptor 2 gene in non-small cell lung cancers. Cancer Res 1999, 59, 5683–5686. [Google Scholar]
- Fisher, M.J.; Virmani, A.K.; Wu, L.; Aplenc, R.; Harper, J.C.; Powell, S.M.; Rebbeck, T.R.; Sidransky, D.; Gazdar, A.F.; El-Deiry, W.S. Nucleotide substitution in the ectodomain of trail receptor dr4 is associated with lung cancer and head and neck cancer. Clin. Cancer Res 2001, 7, 1688–1697. [Google Scholar]
- Strater, J.; Hinz, U.; Walczak, H.; Mechtersheimer, G.; Koretz, K.; Herfarth, C.; Moller, P.; Lehnert, T. Expression of trail and trail receptors in colon carcinoma: Trail-r1 is an independent prognostic parameter. Clin. Cancer Res 2002, 8, 3734–3740. [Google Scholar]
- Zhang, L.; Fang, B. Mechanisms of resistance to trail-induced apoptosis in cancer. Cancer Gene Ther 2005, 12, 228–237. [Google Scholar]
- LeBlanc, H.N.; Ashkenazi, A. Apo2l/trail and its death and decoy receptors. Cell Death Differ 2003, 10, 66–75. [Google Scholar]
- Carlo-Stella, C.; Lavazza, C.; Locatelli, A.; Vigano, L.; Gianni, A.M.; Gianni, L. Targeting trail agonistic receptors for cancer therapy. Clin. Cancer Res 2007, 13, 2313–2317. [Google Scholar]
- Sheridan, J.P.; Marsters, S.A.; Pitti, R.M.; Gurney, A.; Skubatch, M.; Baldwin, D.; Ramakrishnan, L.; Gray, C.L.; Baker, K.; Wood, W.I.; et al. Control of trail-induced apoptosis by a family of signaling and decoy receptors. Science 1997, 277, 818–821. [Google Scholar]
- Pei, G.T.; Wu, C.W.; Lin, W.W. Hypoxia-induced decoy receptor 2 gene expression is regulated via a hypoxia-inducible factor 1alpha-mediated mechanism. Biochem. Biophys. Res. Commun 2010, 391, 1274–1279. [Google Scholar]
- Meng, R.D.; McDonald, E.R., 3rd; Sheikh, M.S.; Fornace, A.J., Jr.; El-Deiry, W.S. The trail decoy receptor trundd (dcr2, trail-r4) is induced by adenovirus-p53 overexpression and can delay trail-, p53-, and killer/dr5-dependent colon cancer apoptosis. Mol. Ther 2000, 1, 130–144. [Google Scholar]
- De Toni, E.N.; Thieme, S.E.; Herbst, A.; Behrens, A.; Stieber, P.; Jung, A.; Blum, H.; Goke, B.; Kolligs, F.T. Opg is regulated by beta-catenin and mediates resistance to trail-induced apoptosis in colon cancer. Clin. Cancer Res 2008, 14, 4713–4718. [Google Scholar]
- Scaffidi, C.; Fulda, S.; Srinivasan, A.; Friesen, C.; Li, F.; Tomaselli, K.J.; Debatin, K.M.; Krammer, P.H.; Peter, M.E. Two cd95 (apo-1/fas) signaling pathways. EMBO J 1998, 17, 1675–1687. [Google Scholar]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar]
- Zou, H.; Henzel, W.J.; Liu, X.; Lutschg, A.; Wang, X. Apaf-1, a human protein homologous to c. Elegans CED-4, participates in cytochrome c-dependent activation of caspase-3. Cell 1997, 90, 405–413. [Google Scholar]
- Verhagen, A.M.; Ekert, P.G.; Pakusch, M.; Silke, J.; Connolly, L.M.; Reid, G.E.; Moritz, R.L.; Simpson, R.J.; Vaux, D.L. Identification of diablo, a mammalian protein that promotes apoptosis by binding to and antagonizing iap proteins. Cell 2000, 102, 43–53. [Google Scholar]
- Elrod, H.A.; Sun, S.Y. Modulation of death receptors by cancer therapeutic agents. Cancer Biol. Ther 2008, 7, 163–173. [Google Scholar]
- Jin, Z.; McDonald, E.R., 3rd; Dicker, D.T.; El-Deiry, W.S. Deficient tumor necrosis factor-related apoptosis-inducing ligand (trail) death receptor transport to the cell surface in human colon cancer cells selected for resistance to trail-induced apoptosis. J. Biol. Chem 2004, 279, 35829–35839. [Google Scholar]
- Muppidi, J.R.; Tschopp, J.; Siegel, R.M. Life and death decisions: Secondary complexes and lipid rafts in tnf receptor family signal transduction. Immunity 2004, 21, 461–465. [Google Scholar]
- Martin, S.; Phillips, D.C.; Szekely-Szucs, K.; Elghazi, L.; Desmots, F.; Houghton, J.A. Cyclooxygenase-2 inhibition sensitizes human colon carcinoma cells to trail-induced apoptosis through clustering of dr5 and concentrating death-inducing signaling complex components into ceramide-enriched caveolae. Cancer Res 2005, 65, 11447–11458. [Google Scholar]
- Delmas, D.; Rebe, C.; Micheau, O.; Athias, A.; Gambert, P.; Grazide, S.; Laurent, G.; Latruffe, N.; Solary, E. Redistribution of cd95, dr4 and dr5 in rafts accounts for the synergistic toxicity of resveratrol and death receptor ligands in colon carcinoma cells. Oncogene 2004, 23, 8979–8986. [Google Scholar]
- Pai, S.I.; Wu, G.S.; Ozoren, N.; Wu, L.; Jen, J.; Sidransky, D.; El-Deiry, W.S. Rare loss-of-function mutation of a death receptor gene in head and neck cancer. Cancer Res 1998, 58, 3513–3518. [Google Scholar]
- Van Geelen, C.M.; Pennarun, B.; Ek, W.B.; Le, P.T.; Spierings, D.C.; de Vries, E.G.; de Jong, S. Downregulation of active caspase 8 as a mechanism of acquired trail resistance in mismatch repair-proficient colon carcinoma cell lines. Int. J. Oncol 2010, 37, 1031–1041. [Google Scholar]
- Micheau, O.; Thome, M.; Schneider, P.; Holler, N.; Tschopp, J.; Nicholson, D.W.; Briand, C.; Grutter, M.G. The long form of flip is an activator of caspase-8 at the fas death-inducing signaling complex. J. Biol. Chem 2002, 277, 45162–45171. [Google Scholar]
- Chang, D.W.; Xing, Z.; Pan, Y.; Algeciras-Schimnich, A.; Barnhart, B.C.; Yaish-Ohad, S.; Peter, M.E.; Yang, X. C-flip(l) is a dual function regulator for caspase-8 activation and cd95-mediated apoptosis. EMBO J 2002, 21, 3704–3714. [Google Scholar]
- Van Geelen, C.M.; de Vries, E.G.; de Jong, S. Lessons from trail-resistance mechanisms in colorectal cancer cells: Paving the road to patient-tailored therapy. Drug Resist. Updat 2004, 7, 345–358. [Google Scholar]
- Deveraux, Q.L.; Reed, J.C. Iap family proteins—Suppressors of apoptosis. Genes Dev 1999, 13, 239–252. [Google Scholar]
- Kim, P.J.; Plescia, J.; Clevers, H.; Fearon, E.R.; Altieri, D.C. Survivin and molecular pathogenesis of colorectal cancer. Lancet 2003, 362, 205–209. [Google Scholar]
- Takeuchi, H.; Kim, J.; Fujimoto, A.; Umetani, N.; Mori, T.; Bilchik, A.; Turner, R.; Tran, A.; Kuo, C.; Hoon, D.S. X-linked inhibitor of apoptosis protein expression level in colorectal cancer is regulated by hepatocyte growth factor/c-met pathway via akt signaling. Clin. Cancer Res 2005, 11, 7621–7628. [Google Scholar]
- Lacour, S.; Hammann, A.; Wotawa, A.; Corcos, L.; Solary, E.; Dimanche-Boitrel, M.T. Anticancer agents sensitize tumor cells to tumor necrosis factor-related apoptosis-inducing ligand-mediated caspase-8 activation and apoptosis. Cancer Res 2001, 61, 1645–1651. [Google Scholar]
- Lacour, S.; Micheau, O.; Hammann, A.; Drouineaud, V.; Tschopp, J.; Solary, E.; Dimanche-Boitrel, M.T. Chemotherapy enhances tnf-related apoptosis-inducing ligand disc assembly in ht29 human colon cancer cells. Oncogene 2003, 22, 1807–1816. [Google Scholar]
- Galligan, L.; Longley, D.B.; McEwan, M.; Wilson, T.R.; McLaughlin, K.; Johnston, P.G. Chemotherapy and trail-mediated colon cancer cell death: The roles of p53, trail receptors, and c-flip. Mol. Cancer Ther 2005, 4, 2026–2036. [Google Scholar]
- Henry, R.E.; Andrysik, Z.; Paris, R.; Galbraith, M.D.; Espinosa, J.M. A dr4:Tbid axis drives the p53 apoptotic response by promoting oligomerization of poised bax. EMBO J 2012, 31, 1266–1278. [Google Scholar]
- Morizot, A.; Merino, D.; Lalaoui, N.; Jacquemin, G.; Granci, V.; Iessi, E.; Lanneau, D.; Bouyer, F.; Solary, E.; Chauffert, B.; et al. Chemotherapy overcomes trail-r4-mediated trail resistance at the disc level. Cell Death Differ 2011, 18, 700–711. [Google Scholar]
- Xiang, H.; Fox, J.A.; Totpal, K.; Aikawa, M.; Dupree, K.; Sinicropi, D.; Lowe, J.; Escandon, E. Enhanced tumor killing by apo2l/trail and cpt-11 co-treatment is associated with p21 cleavage and differential regulation of apo2l/trail ligand and its receptors. Oncogene 2002, 21, 3611–3619. [Google Scholar]
- Kouhara, J.; Yoshida, T.; Nakata, S.; Horinaka, M.; Wakada, M.; Ueda, Y.; Yamagishi, H.; Sakai, T. Fenretinide up-regulates dr5/trail-r2 expression via the induction of the transcription factor chop and combined treatment with fenretinide and trail induces synergistic apoptosis in colon cancer cell lines. Int. J. Oncol 2007, 30, 679–687. [Google Scholar]
- Dolloff, N.G.; Mayes, P.A.; Hart, L.S.; Dicker, D.T.; Humphreys, R.; El-Deiry, W.S. Off-target lapatinib activity sensitizes colon cancer cells through trail death receptor up-regulation. Sci. Transl. Med 2011, 3. [Google Scholar] [CrossRef]
- Ding, W.; Cai, T.; Zhu, H.; Wu, R.; Tu, C.; Yang, L.; Lu, W.; He, Q.; Yang, B. Synergistic antitumor effect of trail in combination with sunitinib in vitro and in vivo. Cancer Lett 2010, 293, 158–166. [Google Scholar]
- El Fajoui, Z.; Toscano, F.; Jacquemin, G.; Abello, J.; Scoazec, J.Y.; Micheau, O.; Saurin, J.C. Oxaliplatin sensitizes human colon cancer cells to trail through jnk-dependent phosphorylation of bcl-xl. Gastroenterology 2011, 141, 663–673. [Google Scholar]
- Horinaka, M.; Yoshida, T.; Shiraishi, T.; Nakata, S.; Wakada, M.; Sakai, T. The dietary flavonoid apigenin sensitizes malignant tumor cells to tumor necrosis factor-related apoptosis-inducing ligand. Mol. Cancer Ther 2006, 5, 945–951. [Google Scholar]
- Taniguchi, H.; Yoshida, T.; Horinaka, M.; Yasuda, T.; Goda, A.E.; Konishi, M.; Wakada, M.; Kataoka, K.; Yoshikawa, T.; Sakai, T. Baicalein overcomes tumor necrosis factor-related apoptosis-inducing ligand resistance via two different cell-specific pathways in cancer cells but not in normal cells. Cancer Res 2008, 68, 8918–8927. [Google Scholar]
- Yoshida, T.; Horinaka, M.; Takara, M.; Tsuchihashi, M.; Mukai, N.; Wakada, M.; Sakai, T. Combination of isoliquiritigenin and tumor necrosis factor-related apoptosis-inducing ligand induces apoptosis in colon cancer ht29 cells. Environ. Health Prev. Med 2008, 13, 281–287. [Google Scholar]
- Yoshida, T.; Konishi, M.; Horinaka, M.; Yasuda, T.; Goda, A.E.; Taniguchi, H.; Yano, K.; Wakada, M.; Sakai, T. Kaempferol sensitizes colon cancer cells to trail-induced apoptosis. Biochem. Biophys. Res. Commun 2008, 375, 129–133. [Google Scholar]
- Psahoulia, F.H.; Drosopoulos, K.G.; Doubravska, L.; Andera, L.; Pintzas, A. Quercetin enhances trail-mediated apoptosis in colon cancer cells by inducing the accumulation of death receptors in lipid rafts. Mol. Cancer Ther 2007, 6, 2591–2599. [Google Scholar]
- Ding, J.; Polier, G.; Kohler, R.; Giaisi, M.; Krammer, P.H.; Li-Weber, M. Wogonin and related natural flavones overcome tumor necrosis factor-related apoptosis-inducing ligand (trail) protein resistance of tumors by down-regulation of c-flip protein and up-regulation of trail receptor 2 expression. J. Biol. Chem 2012, 287, 641–649. [Google Scholar]
- Kong, C.K.; Lam, W.S.; Chiu, L.C.; Ooi, V.E.; Sun, S.S.; Wong, Y.S. A rice bran polyphenol, cycloartenyl ferulate, elicits apoptosis in human colorectal adenocarcinoma sw480 and sensitizes metastatic sw620 cells to trail-induced apoptosis. Biochem. Pharmacol 2009, 77, 1487–1496. [Google Scholar]
- Kauntz, H.; Bousserouel, S.; Gosse, F.; Raul, F. Silibinin triggers apoptotic signaling pathways and autophagic survival response in human colon adenocarcinoma cells and their derived metastatic cells. Apoptosis 2011, 16, 1042–1053. [Google Scholar]
- Sung, B.; Ravindran, J.; Prasad, S.; Pandey, M.K.; Aggarwal, B.B. Gossypol induces death receptor-5 through activation of the ros-erk-chop pathway and sensitizes colon cancer cells to trail. J. Biol. Chem 2010, 285, 35418–35427. [Google Scholar]
- Yadav, V.R.; Prasad, S.; Aggarwal, B.B. Cardamonin sensitizes tumour cells to trail through ros- and chop-mediated up-regulation of death receptors and down-regulation of survival proteins. Br. J. Pharmacol 2012, 165, 741–753. [Google Scholar]
- Yodkeeree, S.; Sung, B.; Limtrakul, P.; Aggarwal, B.B. Zerumbone enhances trail-induced apoptosis through the induction of death receptors in human colon cancer cells: Evidence for an essential role of reactive oxygen species. Cancer Res 2009, 69, 6581–6589. [Google Scholar]
- Gupta, S.C.; Reuter, S.; Phromnoi, K.; Park, B.; Hema, P.S.; Nair, M.; Aggarwal, B.B. Nimbolide sensitizes human colon cancer cells to trail through reactive oxygen species- and erk-dependent up-regulation of death receptors, p53, and bax. J. Biol. Chem 2011, 286, 1134–1146. [Google Scholar]
- Yoshida, T.; Maoka, T.; Das, S.K.; Kanazawa, K.; Horinaka, M.; Wakada, M.; Satomi, Y.; Nishino, H.; Sakai, T. Halocynthiaxanthin and peridinin sensitize colon cancer cell lines to tumor necrosis factor-related apoptosis-inducing ligand. Mol. Cancer Res 2007, 5, 615–625. [Google Scholar]
- Kannappan, R.; Ravindran, J.; Prasad, S.; Sung, B.; Yadav, V.R.; Reuter, S.; Chaturvedi, M.M.; Aggarwal, B.B. Gamma-tocotrienol promotes trail-induced apoptosis through reactive oxygen species/extracellular signal-regulated kinase/p53-mediated upregulation of death receptors. Mol. Cancer Ther 2010, 9, 2196–2207. [Google Scholar]
- Prasad, S.; Ravindran, J.; Sung, B.; Pandey, M.K.; Aggarwal, B.B. Garcinol potentiates trail-induced apoptosis through modulation of death receptors and antiapoptotic proteins. Mol. Cancer Ther 2010, 9, 856–868. [Google Scholar]
- Zhang, C.; Wu, R.; Zhu, H.; Hu, Y.Z.; Jiang, H.; Lin, N.M.; He, Q.J.; Yang, B. Enhanced anti-tumor activity by the combination of trail/apo-2l and combretastatin a-4 against human colon cancer cells via induction of apoptosis in vitro and in vivo. Cancer Lett 2011, 302, 11–19. [Google Scholar]
- Raviv, Z.; Zilberberg, A.; Cohen, S.; Reischer-Pelech, D.; Horrix, C.; Berger, M.R.; Rosin-Arbesfeld, R.; Flescher, E. Methyl jasmonate down-regulates survivin expression and sensitizes colon carcinoma cells towards trail-induced cytotoxicity. Br. J. Pharmacol 2011, 164, 1433–1444. [Google Scholar]
- Vaculova, A.; Hofmanova, J.; Andera, L.; Kozubik, A. Trail and docosahexaenoic acid cooperate to induce ht-29 colon cancer cell death. Cancer Lett 2005, 229, 43–48. [Google Scholar]
- Lepage, C.; Leger, D.Y.; Bertrand, J.; Martin, F.; Beneytout, J.L.; Liagre, B. Diosgenin induces death receptor-5 through activation of p38 pathway and promotes trail-induced apoptosis in colon cancer cells. Cancer Lett 2011, 301, 193–202. [Google Scholar]
- Zhang, L.; Ren, X.; Alt, E.; Bai, X.; Huang, S.; Xu, Z.; Lynch, P.M.; Moyer, M.P.; Wen, X.F.; Wu, X. Chemoprevention of colorectal cancer by targeting apc-deficient cells for apoptosis. Nature 2010, 464, 1058–1061. [Google Scholar]
- He, Q.; Huang, Y.; Sheikh, M.S. Proteasome inhibitor mg132 upregulates death receptor 5 and cooperates with apo2l/trail to induce apoptosis in bax-proficient and -deficient cells. Oncogene 2004, 23, 2554–2558. [Google Scholar]
- Johnson, T.R.; Stone, K.; Nikrad, M.; Yeh, T.; Zong, W.X.; Thompson, C.B.; Nesterov, A.; Kraft, A.S. The proteasome inhibitor ps-341 overcomes trail resistance in bax and caspase 9-negative or bcl-xl overexpressing cells. Oncogene 2003, 22, 4953–4963. [Google Scholar]
- Su, R.Y.; Chi, K.H.; Huang, D.Y.; Tai, M.H.; Lin, W.W. 15-deoxy-delta12,14-prostaglandin j2 up-regulates death receptor 5 gene expression in hct116 cells: Involvement of reactive oxygen species and c/ebp homologous transcription factor gene transcription. Mol. Cancer Ther 2008, 7, 3429–3440. [Google Scholar]
- Prasad, S.; Yadav, V.R.; Ravindran, J.; Aggarwal, B.B. Ros and chop are critical for dibenzylideneacetone to sensitize tumor cells to trail through induction of death receptors and downregulation of cell survival proteins. Cancer Res 2011, 71, 538–549. [Google Scholar]
- Stolfi, C.; Caruso, R.; Franze, E.; Rizzo, A.; Rotondi, A.; Monteleone, I.; Fantini, M.C.; Pallone, F.; Monteleone, G. 2-methoxy-5-amino-n-hydroxybenzamide sensitizes colon cancer cells to trail-induced apoptosis by regulating death receptor 5 and survivin expression. Mol. Cancer Ther 2011, 10, 1969–1981. [Google Scholar]
- Goda, A.E.; Yoshida, T.; Horinaka, M.; Yasuda, T.; Shiraishi, T.; Wakada, M.; Sakai, T. Mechanisms of enhancement of trail tumoricidal activity against human cancer cells of different origin by dipyridamole. Oncogene 2008, 27, 3435–3445. [Google Scholar]
- Hori, T.; Kondo, T.; Kanamori, M.; Tabuchi, Y.; Ogawa, R.; Zhao, Q.L.; Ahmed, K.; Yasuda, T.; Seki, S.; Suzuki, K.; et al. Nutlin-3 enhances tumor necrosis factor-related apoptosis-inducing ligand (trail)-induced apoptosis through up-regulation of death receptor 5 (DR5) in human sarcoma hos cells and human colon cancer hct116 cells. Cancer Lett 2010, 287, 98–108. [Google Scholar]
- Su, R.Y.; Chao, Y.; Chen, T.Y.; Huang, D.Y.; Lin, W.W. 5-aminoimidazole-4-carboxamide riboside sensitizes trail- and tnf{alpha}-induced cytotoxicity in colon cancer cells through amp-activated protein kinase signaling. Mol. Cancer Ther 2007, 6, 1562–1571. [Google Scholar]
- Tillman, D.M.; Izeradjene, K.; Szucs, K.S.; Douglas, L.; Houghton, J.A. Rottlerin sensitizes colon carcinoma cells to tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis via uncoupling of the mitochondria independent of protein kinase c. Cancer Res 2003, 63, 5118–5125. [Google Scholar]
- Vasilevskaya, I.A.; O’Dwyer, P.J. 17-allylamino-17-demethoxygeldanamycin overcomes trail resistance in colon cancer cell lines. Biochem. Pharmacol 2005, 70, 580–589. [Google Scholar]
- Toscano, F.; Fajoui, Z.E.; Gay, F.; Lalaoui, N.; Parmentier, B.; Chayvialle, J.A.; Scoazec, J.Y.; Micheau, O.; Abello, J.; Saurin, J.C. P53-mediated upregulation of dcr1 impairs oxaliplatin/trail-induced synergistic anti-tumour potential in colon cancer cells. Oncogene 2008, 27, 4161–4171. [Google Scholar]
- Wu, X.X.; Ogawa, O.; Kakehi, Y. Trail and chemotherapeutic drugs in cancer therapy. Vitam. Horm 2004, 67, 365–383. [Google Scholar]
- Siu, D. Natural products and their role in cancer therapy. Med. Oncol 2011, 28, 888–900. [Google Scholar]
- Oliver, C.L.; Bauer, J.A.; Wolter, K.G.; Ubell, M.L.; Narayan, A.; O’Connell, K.M.; Fisher, S.G.; Wang, S.; Wu, X.; Ji, M.; et al. In vitro effects of the bh3 mimetic, (−)-gossypol, on head and neck squamous cell carcinoma cells. Clin. Cancer Res 2004, 10, 7757–7763. [Google Scholar]
- Rabi, T.; Bishayee, A. Terpenoids and breast cancer chemoprevention. Breast Cancer Res. Treat 2009, 115, 223–239. [Google Scholar]
- Thoppil, R.J.; Bishayee, A. Terpenoids as potential chemopreventive and therapeutic agents in liver cancer. World J. Hepatol 2011, 3, 228–249. [Google Scholar]
- Kirana, C.; McIntosh, G.H.; Record, I.R.; Jones, G.P. Antitumor activity of extract of zingiber aromaticum and its bioactive sesquiterpenoid zerumbone. Nutr. Cancer 2003, 45, 218–225. [Google Scholar]
- Sterz, J.; von Metzler, I.; Hahne, J.C.; Lamottke, B.; Rademacher, J.; Heider, U.; Terpos, E.; Sezer, O. The potential of proteasome inhibitors in cancer therapy. Expert Opin. Investig. Drugs 2008, 17, 879–895. [Google Scholar]
- Schroder, M.; Kaufman, R.J. Er stress and the unfolded protein response. Mutat. Res 2005, 569, 29–63. [Google Scholar]
- Martin-Perez, R.; Niwa, M.; Lopez-Rivas, A. Er stress sensitizes cells to trail through down-regulation of flip and mcl-1 and perk-dependent up-regulation of trail-r2. Apoptosis 2012, 17, 349–363. [Google Scholar]
- Stolfi, C.; Sarra, M.; Caruso, R.; Fantini, M.C.; Fina, D.; Pellegrini, R.; Palmieri, G.; Macdonald, T.T.; Pallone, F.; Monteleone, G. Inhibition of colon carcinogenesis by 2-methoxy-5-amino-n-hydroxybenzamide, a novel derivative of mesalamine. Gastroenterology 2010, 138, 221–230. [Google Scholar]
- Zhao, J.; Lu, Y.; Shen, H.M. Targeting p53 as a therapeutic strategy in sensitizing trail-induced apoptosis in cancer cells. Cancer Lett 2012, 314, 8–23. [Google Scholar]
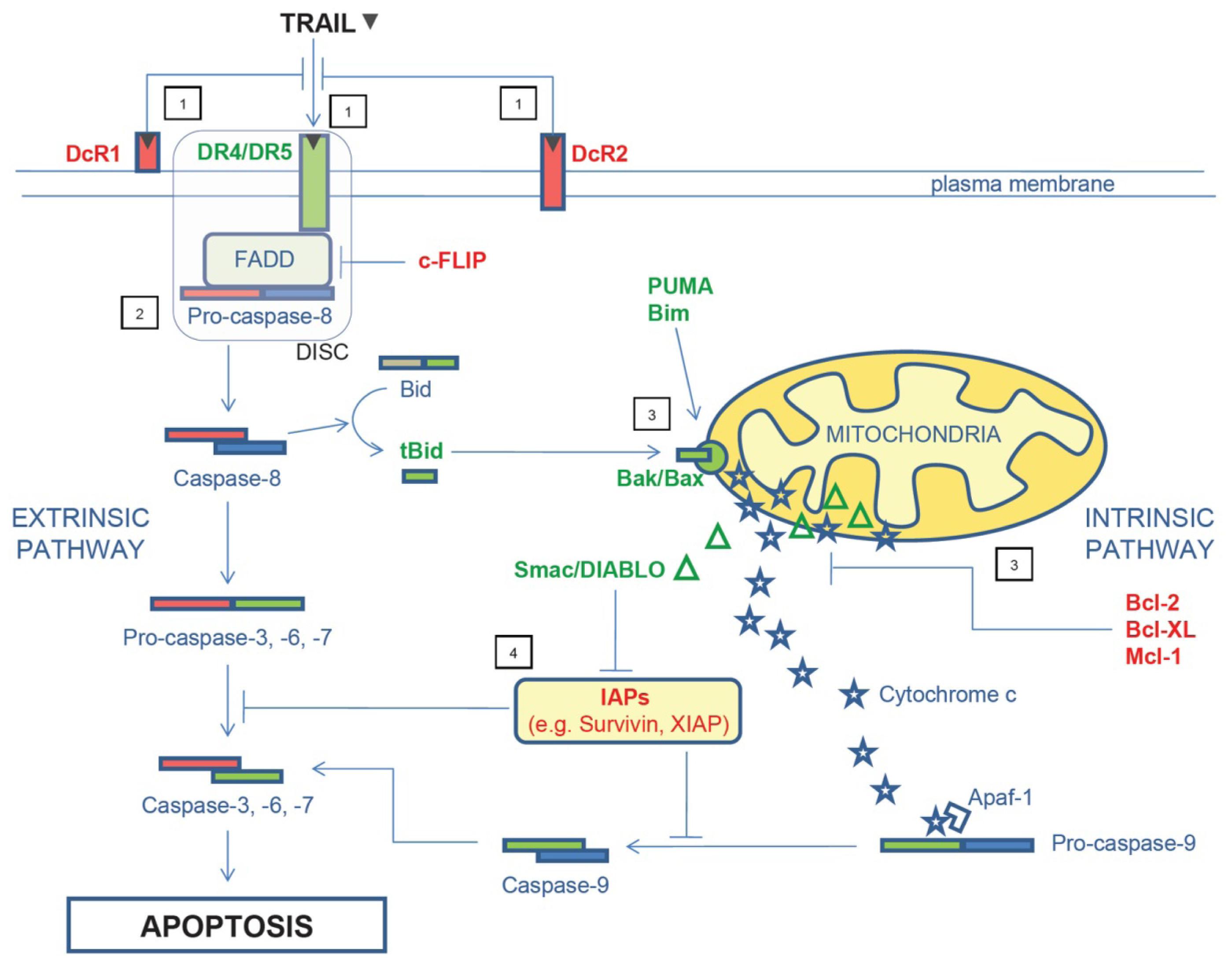

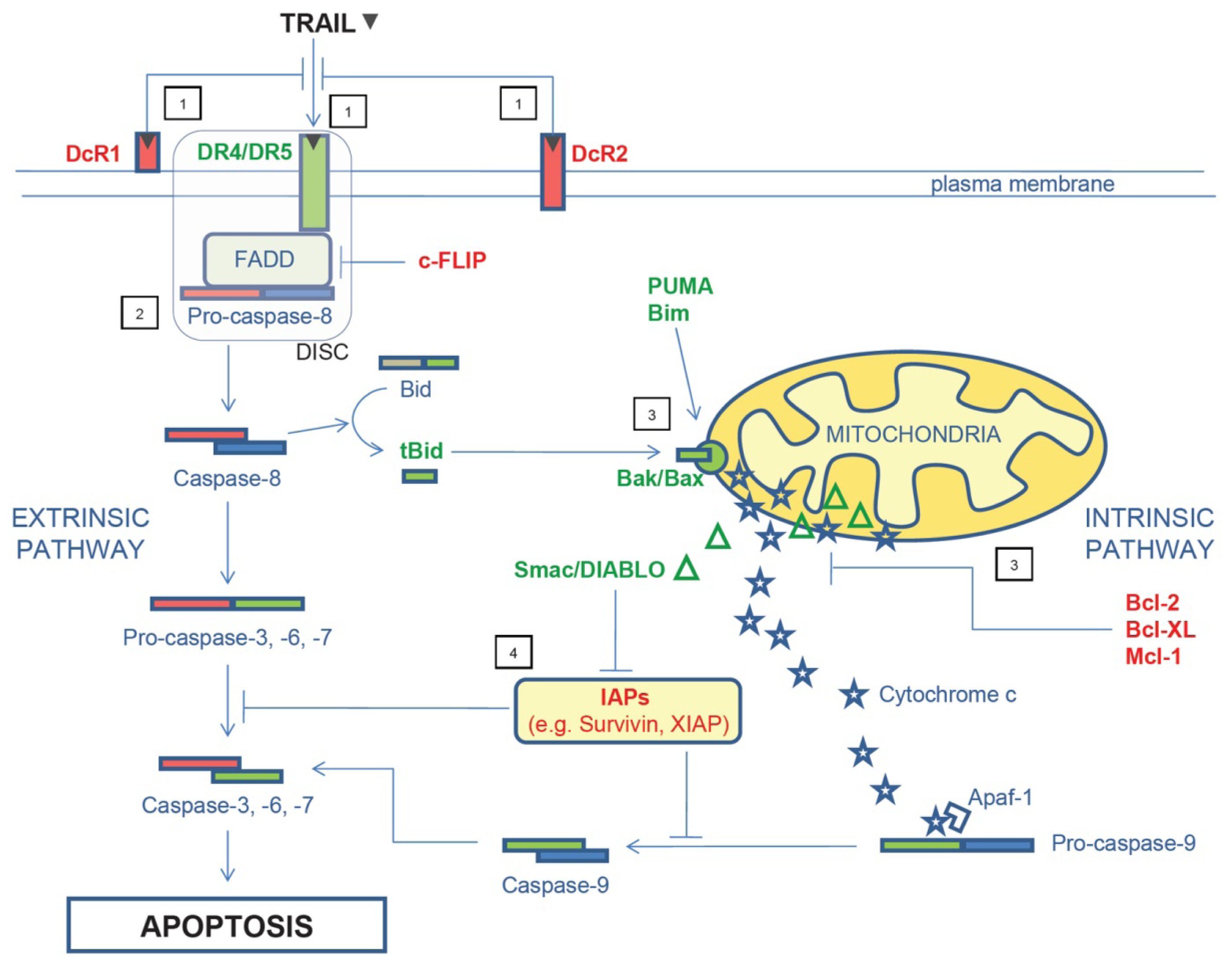{kind=link}
| Therapeutics | Cells | Mechanism(s) | Reference |
|---|---|---|---|
| Chemotherapeutics | |||
| Cisplatin | HT-29, SW480 | Caspase-8 activation, Bid cleavage | [35,36] |
| 5-FU | HCT-116, HT-29 | Caspase-8 activation, ↓c-FLIP, Bid cleavage | [35–38] |
| VP16 | HCT-116, SW480 | Caspase-8 activation | [39] |
| Doxorubicin | HT-29, SW480 | Caspase-8 activation, Bid cleavage | [35,36] |
| DuP-697 | HT-29 | DR5 and DISC component redistribution in lipid rafts | [25] |
| CPT-11 | HCT-116 | ↑DR4, ↑DR5 | [40] |
| Fenretinide | HT-29, SW480 | ↑DR5, Bid cleavage | [41] |
| Lapatinib | HCT-116, SW480, SW620, DLD-1, HT-29 | ↑DR4, ↑DR5 | [42] |
| Sunitinib | SW620 | ↓c-FLIP, ↓Mcl-1, ↓XIAP | [43] |
| Oxaliplatin | HT-29, V9P | ↓Bcl-xL anti-apoptotic activity | [44] |
| Natural products | |||
| Apigenin | DLD1 | ↑DR5 | [45] |
| Baicalein | SW480 | ↑DR5 | [46] |
| Isoliquiritigenin | HT-29 | ↑DR5 | [47] |
| Kaempferol | SW480 | ↑DR5 | [48] |
| Quercetin | HT-29, SW620, CACO-2 | DR4 and DR5 redistribution in lipid rafts | [49] |
| Wogonin | HT-29 | ↑DR5, ↓c-FLIP | [50] |
| Cycloartenyl ferulate | SW480, SW620 | ↑DR4, ↑DR5, Bid cleavage, Bcl-2, Bax | [51] |
| Silibinin | SW480, SW620 | ↑DR5, ↓Mcl-1, ↓XIAP | [52] |
| Gossypol | HCT-116 | ↑DR5, ↓Bcl-2, ↓Bcl-xL, ↓survivin | [53] |
| Cardamonin | HCT-116 | ↑DR4, ↑DR5, ↓DcR1, ↑Bax, ↓Bcl-2, ↓c-IAP1 | [54] |
| Zerumbone | HCT-116 | ↑DR4, ↑DR5, ↓c-FLIP | [55] |
| Nimbolide | HCT-116, HT-29 | ↑DR4, ↑DR5, ↓c-FLIP, ↑Bax, ↓Bcl-2, ↓Bcl-xL, ↓c-IAP1, ↓c-IAP2, ↓survivin, ↓XIAP | [56] |
| Halocynthiaxanthin | DLD1, HT-29 | ↑DR5 | [57] |
| γ-T3 | HCT-116 | ↑DR4, ↑DR5 | [58] |
| Garcinol | HCT-116, HT-29 | ↑DR4, ↑DR5, Bid cleavage ↓c-FLIP, ↓Bcl-2, ↓XIAP, ↓survivin | [59] |
| Combretastatin A-4 | HCT-116, SW620 | ↓c-FLIP, ↓Mcl-1 | [60] |
| Methyl jasmonate | HCT-116, SW480 | Bid cleavage, ↓survivin | [61] |
| DHA | HT-29 | Caspase-8 activation, Bid cleavage | [62] |
| Diosgenin | HT-29 | ↑DR5 | [63] |
| RAc | APC-deleted NCM356 | ↑DR4, ↑DR5, ↓DcR1, ↓DcR2 | [64] |
| Other agents | |||
| MG-132 | HCT-116 | ↑DR5 | [65] |
| PS-341 | HCT-116, HC-4 | ↑DR4, ↑DR5, caspase-8 activation, Bid cleavage | [66] |
| 15dPGJ2 | HCT-116 | ↑DR5 | [67] |
| DBA | HCT-116 | ↑DR4, ↑DR5, ↓DcR2, ↓Bcl-2, ↓XIAP, ↓survivin | [68] |
| 2–14 | DLD1, HT-29 | ↑DR5, ↓survivin | [69] |
| Dipyridamole | SW480 | ↑DR5, ↓survivin | [70] |
| Nutlin-3 | HCT-116 | ↑DR5, ↓Bcl-2, ↓XIAP | [71] |
| AICAR | HCT-116 | Bid cleavage, ↓Bcl-2 | [72] |
| Rottlerin | HT-29, RKO | Cytochrome c and Smac/DIABLO release, ↓XIAP ↓c-IAP1 | [73] |
| 17-AAG | HT-29, RKO | Bid cleavage, ↓XIAP | [74] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stolfi, C.; Pallone, F.; Monteleone, G. Molecular Targets of TRAIL-Sensitizing Agents in Colorectal Cancer. Int. J. Mol. Sci. 2012, 13, 7886-7901. https://doi.org/10.3390/ijms13077886
Stolfi C, Pallone F, Monteleone G. Molecular Targets of TRAIL-Sensitizing Agents in Colorectal Cancer. International Journal of Molecular Sciences. 2012; 13(7):7886-7901. https://doi.org/10.3390/ijms13077886
Chicago/Turabian StyleStolfi, Carmine, Francesco Pallone, and Giovanni Monteleone. 2012. "Molecular Targets of TRAIL-Sensitizing Agents in Colorectal Cancer" International Journal of Molecular Sciences 13, no. 7: 7886-7901. https://doi.org/10.3390/ijms13077886