Role of Microglia in Oxidative Toxicity Associated with Encephalomycarditis Virus Infection in the Central Nervous System


{kind=link}
Abstract
:1. Introduction
2. Encephalomycarditis Virus
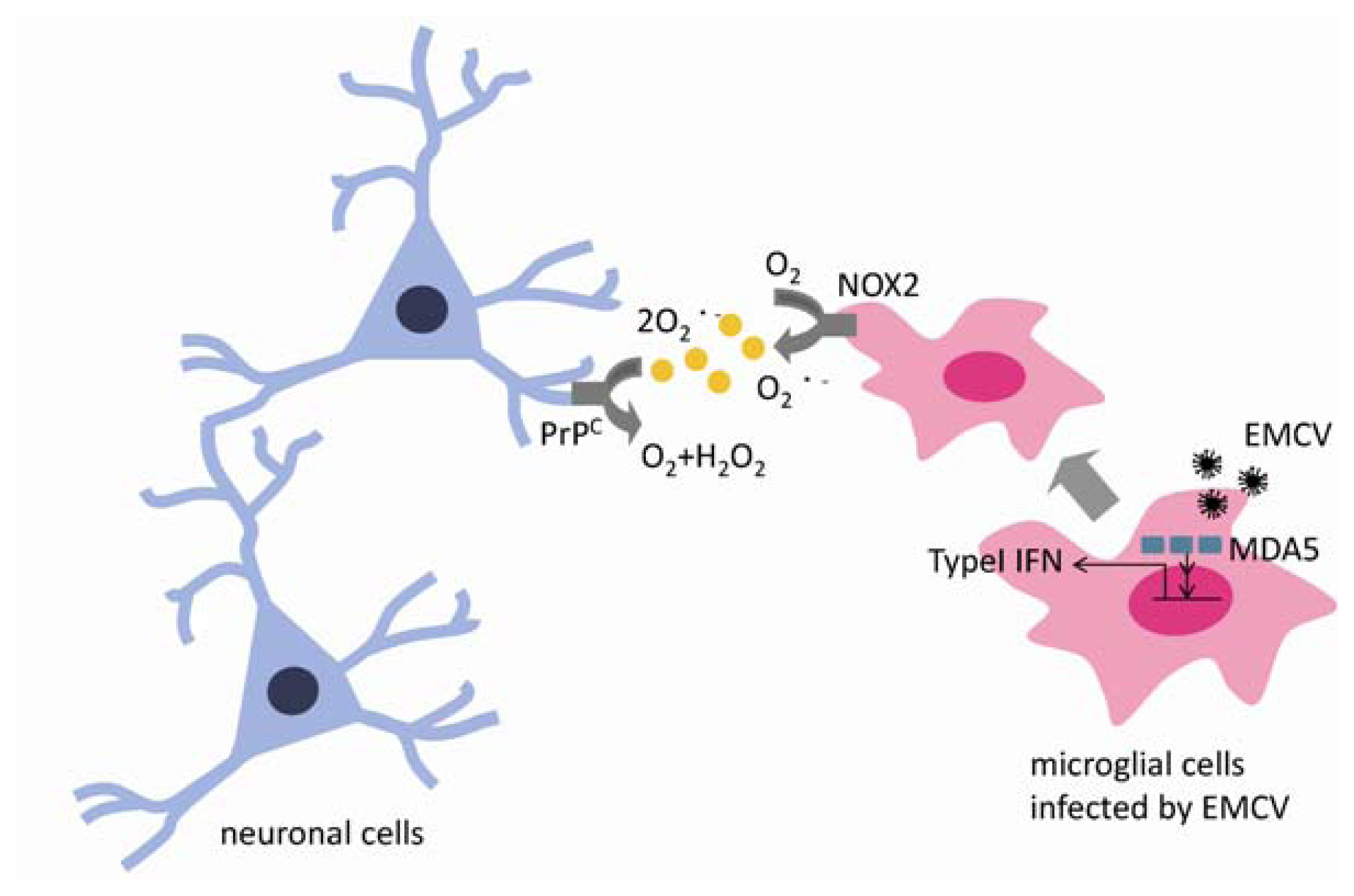
3. Innate Immunity in EMCV Virus Infection
4. Microglia Activation by Virus Infection in the CNS
5. Cellular Prion Proteins Have a Neuroprotective Effect against Oxidative Injury Associated with EMCV Infection in the CNS
6. Conclusions
References
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol 2004, 4, 181–189. [Google Scholar]
- Geiszt, M.; Leto, T.L. The Nox family of NAD(P)H oxidases: Host defense and beyond. J. Biol. Chem 2004, 279, 51715–51718. [Google Scholar]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem 2006, 97, 1634–1658. [Google Scholar]
- Sumimoto, H.; Miyano, K.; Takeya, R. Molecular composition and regulation of the Nox family NAD(P)H oxidases. Biochem. Biophys. Res. Commun 2005, 338, 677–686. [Google Scholar]
- Matthews, R.E.F. The classification and nomenclature of viruses. Intervirology 1979, 11, 133–135. [Google Scholar]
- Doi, K. Experimental encephalomyocarditis virus infection in small laboratory rodents. J. Comp. Path 2011, 144, 25–40. [Google Scholar]
- Ano, Y.; Sakudo, A.; Kimata, T.; Uraki, R.; Sugiura, K.; Onodera, T. Oxidative damage to neurons caused by the induction of microglial NADPH oxidase in encephalomycarditis virus infection. Neurosci. Lett 2010, 469, 39–43. [Google Scholar]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev 2007, 87, 245–313. [Google Scholar]
- Block, M.L.; Hong, J.S. Microglia and inflammation-mediated neurodegeneration: Multiple triggers with a common mechanism. Prog. Neurobiol 2005, 76, 77–98. [Google Scholar]
- Block, M.L.; Hong, J.S. Chronic microglial activation and progressive dopaminergic neurotoxicity. Biochem. Soc. Trans 2007, 35, 1127–1132. [Google Scholar]
- Helwig, F.C.; Schmidt, E.C.H. A filter-passing agentproducing interstitial myocarditis in anthropoid apes and small animals. Science 1945, 102, 31–33. [Google Scholar]
- Hill, B.D.; Ketterer, P.J.; Rodwell, B.J.; Eaves, F.W.; Webster, W.R. Encephalomyocarditis virus infection and pig disease in Queensland. Aust. Vet. J 1985, 62, 433–434. [Google Scholar]
- Simpson, C.F.; Lewis, A.L.; Gaskin, L.M. Encephalomyocarditis virus infection of captive elephants. J. Am. Vet. Med. Assoc 1977, 171, 902–905. [Google Scholar]
- Hubbard, G.B.; Soike, K.F.; Butler, T.M.; Carey, K.D.; Davis, H.; Buthcer, W.I.; Gauntt, C.J. An encephalomyocarditis virus epizootic in a baboon colony. Lab. Anim. Sci 1992, 42, 233–239. [Google Scholar]
- Craighead, J.E. Pathogenicity of M and E variants of encephalomyocarditis (EMC) virus. I. Myocardiotropic and neurotropic properties. Am. J. Path 1966, 48, 333–345. [Google Scholar]
- Yoon, J.W.; McClintock, P.R.; Onodera, T.; Notkins, A.L. Virus-induced diabetes mellitus. XVIII. Inhibition by a non-diabetogenic variant of encephalomyocarditis virus. J. Exp. Med 1980, 152, 872–892. [Google Scholar]
- Kilham, L.; Mason, P.; Davies, J.N.P. Pathogenesis of fatal encephalomyocarditis (EMC) virus infections in Albino rats. Proc. Soc. Exp. Biol. Med 1955, 90, 383–387. [Google Scholar]
- Matsuzaki, H.; Doi, K.; Doi, C.; Onodera, T.; Mitsuoka, T. Susceptibility of four species of small rodents to encephalomyocarditis (EMC) virus infection. Exp. Anim 1989, 38, 357–361. [Google Scholar]
- Sugawara, Y.; Hirasawa, K.; Takeda, M.; Han, J.S.; Doi, K. Acute infection of encephalomyocarditis (EMC) virus in Syrian hamsters. J. Vet. Med. Sci 1991, 53, 463–468. [Google Scholar]
- Shafi, R.; Certis, D.R.; Giron, D.J. Pathogenesis of the B variant of encephalomyocarditis virus. J. Med. Virol 1993, 40, 193–199. [Google Scholar]
- Doi, K.; Onodera, T.; Tsuda, T.; Matsuzaki, H.; Mitsuoka, T. Histopathology of BALB/c mice infected with the D variant of encephalomyocarditis virus. Br. J. Exp. Pathol 1988, 69, 395–401. [Google Scholar]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol 2004, 5, 987–995. [Google Scholar]
- Beutler, B. Inferences, questions and possibilities in Toll-like receptor signalling. Nature 2004, 430, 257–263. [Google Scholar]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar]
- Yoneyama, M.; Kikuci, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol 2004, 5, 730–737. [Google Scholar]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takenouchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat. Immunol 2005, 6, 981–988. [Google Scholar]
- Honda, K.; Yanai, H.; Negishi, H.; Asagiri, M.; Sato, M.; Mizutani, T.; Shimada, N.; Ohba, Y.; Takaoka, A.; Yoshida, N.; et al. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Nature 2005, 434, 772–777. [Google Scholar]
- Yoneyama, M.; Kikuchi, M.; Matsumoto, K.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Foy, E.; Loo, Y.M.; Gale, M., Jr; Akira, S.; et al. Shared and unique functions of the DExD/H-box helicases RIG-I, MDA5 and LGP2 in antiviral innate immunity. J. Immunol 2005, 175, 2851–2858. [Google Scholar]
- Kato, H.; Takeuhi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar]
- Chauhan, V.S.; Furr, S.R.; Sterka, D.G., Jr; Nelson, D.A.; Moerdyk-Schauwecker, M.; Marriott, I.; Grdzelishvili, V.Z. Vesicular stomatitis virus infects resident cells of the central nervous system and induces replication-dependent inflammatory responses. Virology 2010, 400, 187–196. [Google Scholar]
- Furr, S.R.; Chauhan, V.S.; Sterka, D., Jr; Grdzelishvili, V.; Marriott, I. Characterization of retinoic acid-inducible gene-I expression in primary murine glia following exposure to vesicular stomatitis virus. J. Neurovirol 2008, 14, 503–513. [Google Scholar]
- Soucy-Faulkner, A.; Mukawera, E.; Fink, K.; Martel, A.; Jouan, L.; Nzengue, Y.; Lamarre, D.; Vande Velde, C.; Grandvaux, N. Requirement of NOX2 and reactive oxygen species for efficient RIG-I-mediated antiviral response through regulation of MAVS expression. PLoS Pathog 2010, 6. [Google Scholar] [CrossRef]
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol 2004, 4, 181–189. [Google Scholar]
- Geiszt, M.; Leto, T.L. The Nox family of NAD(P)H oxidases: Host defense and beyond. J. Biol. Chem 2004, 279, 51715–51718. [Google Scholar]
- Sumimoto, H. Structure, regulation and evolution of Nox-family NADPH oxidases that produce reactive oxygen species. FEBS J 2008, 275, 3249–3277. [Google Scholar]
- Nauseef, W.M. Assembly of the phagocyte NADPH oxidase. Histochem. Cell Biol 2004, 122, 277–291. [Google Scholar]
- Lam, G.Y.; Huang, J.; Brumell, J.H. The many roles of NOX2 NADPH oxidase-derived ROS in immunity. Semin Immunopathol 2010, 32, 415–430. [Google Scholar]
- Savina, A.; Jancic, C.; Hugues, S.; Guermonprez, P.; Vargas, P.; Moura, I.C.; Lennon-Dumenil, A.M.; Seabra, M.C.; Raposo, G.; Amigorena, S. NOX2 controls phagosomal pH to regulate antigen processing during crosspresentation by dendritic cells. Cell 2006, 126, 205–218. [Google Scholar]
- Tacke, R.S.; Lee, H.C.; Goh, C.; Courtney, J.; Polyak, S.J.; Rosen, H.R.; Hahn, Y.S. Myeloid suppressor cells induced by hepatitis C virus suppress T-cell responses through the production of reactive oxygen species. Hepatology 2012, 55, 343–353. [Google Scholar]
- Park, H.S.; Jung, H.Y.; Park, E.Y.; Kim, J.; Lee, W.J.; Bae, Y.S. Cutting edge: Direct interaction of TLR4 with NAD(P)H oxidase 4 isozyme is essential for lipopolysaccharide-induced production of reactive oxygen species and activation of NF-κB. J. Immunol 2004, 173, 3589–3593. [Google Scholar]
- Vlahos, R.; Stambas, J.; Bozinovski, S.; Broughton, B.R.; Drummond, G.R.; Selemidis, S. Inhibition of Nox2 oxidase activity ameliorates influenza A virus-induced lung inflammation. PLoS Pathog 2011, 7. [Google Scholar] [CrossRef]
- Fink, K.; Duval, A.; Martel, A.; Soucy-Faulkner, A.; Grandvaux, N. Dual role of NOX2 in respiratory syncytial virus- and sendai virus-induced activation of NF-κB in airway epithelial cells. J. Immunol 2008, 180, 6911–6922. [Google Scholar]
- Gruhne, B.; Sompallae, R.; Marescotti, D.; Kamranvar, S.A.; Gastaldello, S.; Masucci, M.G. The Epstein-Barr virus nuclear antigen-1 promotes genomic instability via induction of reactive oxygen species. Proc. Natl. Acad. Sci. USA 2009, 106, 2313–2318. [Google Scholar]
- Nasu-Nishimura, Y.; Taniuchi, Y.; Nishimura, T.; Sakudo, A.; Nakajima, K.; Ano, Y.; Sugiura, K.; Sakaguchi, S.; Itohara, S.; Onodera, T. Cellular prion protein prevents brain damage after encephalomyocarditis virus infection in mice. Arch. Virol 2008, 153, 1007–1012. [Google Scholar]
- Onodera, T.; Sakudo, A.; Wu, G.; Saeki, K. Bovine spongiform encephalopathy in Japan: History and recent studies on oxidative stress in prion diseases. Microbiol. Immunol 2006, 50, 565–578. [Google Scholar]
- Kuwahara, C.; Takeuchi, A.M.; Nishimura, T.; Haraguchi, K.; Kubosaki, A.; Matsumoto, T.; Saeki, K.; Matsumoto, T.; Yokoyama, T.; Itohara, S.; et al. Prions prevent neuronal cell-line death. Nature 1999, 400, 225–226. [Google Scholar]
- Sakudo, A.; Lee, D.C.; Nishimura, T.; Li, S.; Tsuji, S.; Nakamura, T.; Matsumoto, Y.; Saeki, K.; Itohara, S.; Ikuta, K.; et al. Octapeptide repeat region and N-terminal half of hydrophobic region of prion protein (PrP) mediate PrP-dependent activation of superoxide dismutase. Biochem. Biophys. Res. Commun 2005, 326, 600–606. [Google Scholar]
- Kishimoto, C.; Tomioka, N.; Nakayama, Y.; Miyamoto, M. Anti-oxidant effects of coenzyme Q10 on experimental viral myocarditis in mice. J. Cardiovasc. Pharmacol 2003, 42, 588–592. [Google Scholar]
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ano, Y.; Sakudo, A.; Onodera, T. Role of Microglia in Oxidative Toxicity Associated with Encephalomycarditis Virus Infection in the Central Nervous System. Int. J. Mol. Sci. 2012, 13, 7365-7374. https://doi.org/10.3390/ijms13067365
Ano Y, Sakudo A, Onodera T. Role of Microglia in Oxidative Toxicity Associated with Encephalomycarditis Virus Infection in the Central Nervous System. International Journal of Molecular Sciences. 2012; 13(6):7365-7374. https://doi.org/10.3390/ijms13067365
Chicago/Turabian StyleAno, Yasuhisa, Akikazu Sakudo, and Takashi Onodera. 2012. "Role of Microglia in Oxidative Toxicity Associated with Encephalomycarditis Virus Infection in the Central Nervous System" International Journal of Molecular Sciences 13, no. 6: 7365-7374. https://doi.org/10.3390/ijms13067365