An Advanced Electron Spin Resonance (ESR) Spin-Trapping and LC/(ESR)/MS Technique for the Study of Lipid Peroxidation

Abstract
:1. Introduction
2. History of the ESR Spin-Trapping Technique
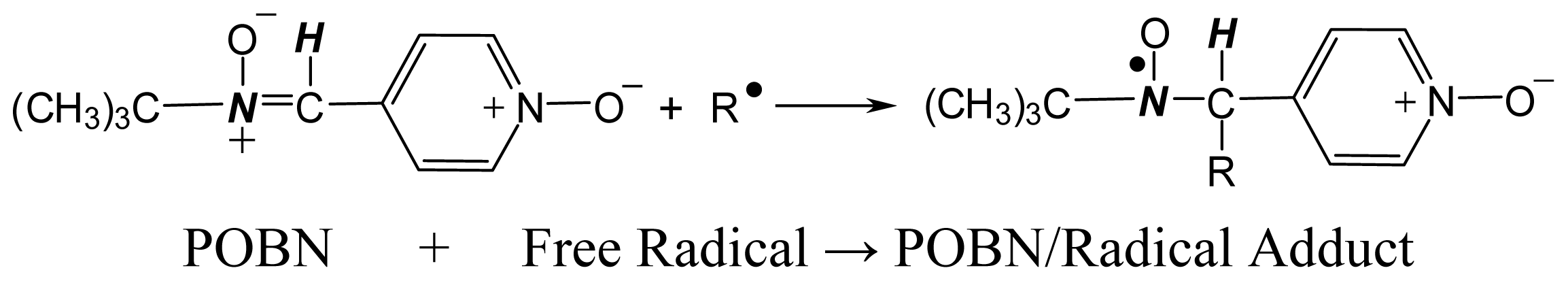
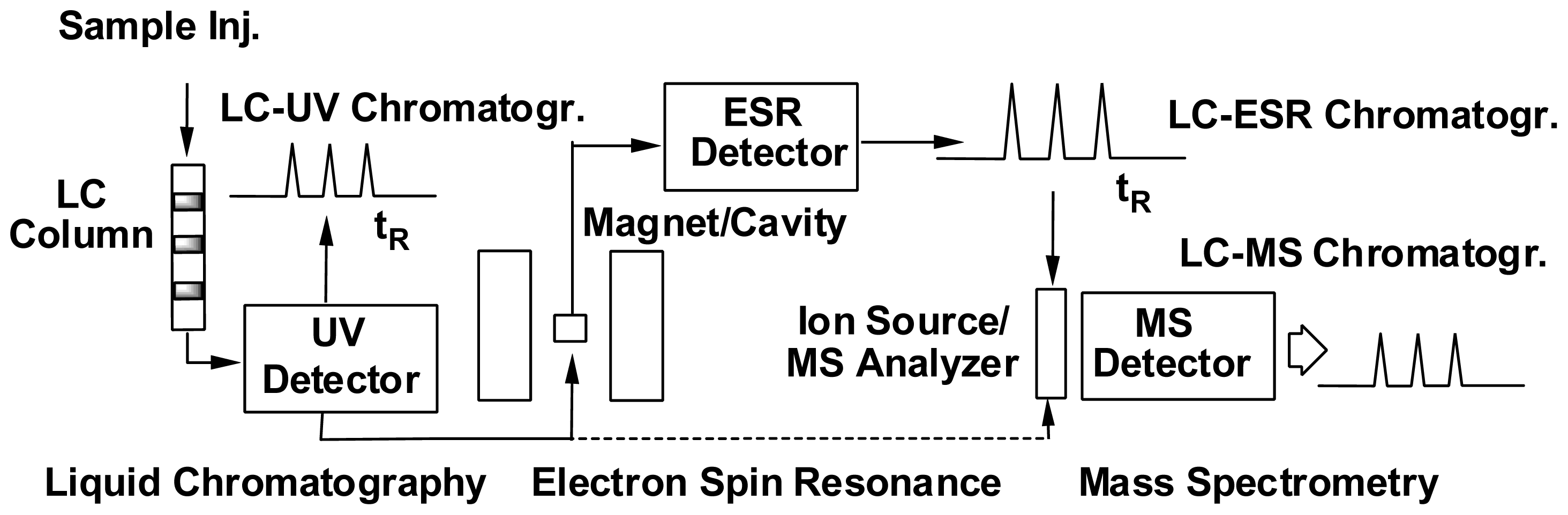
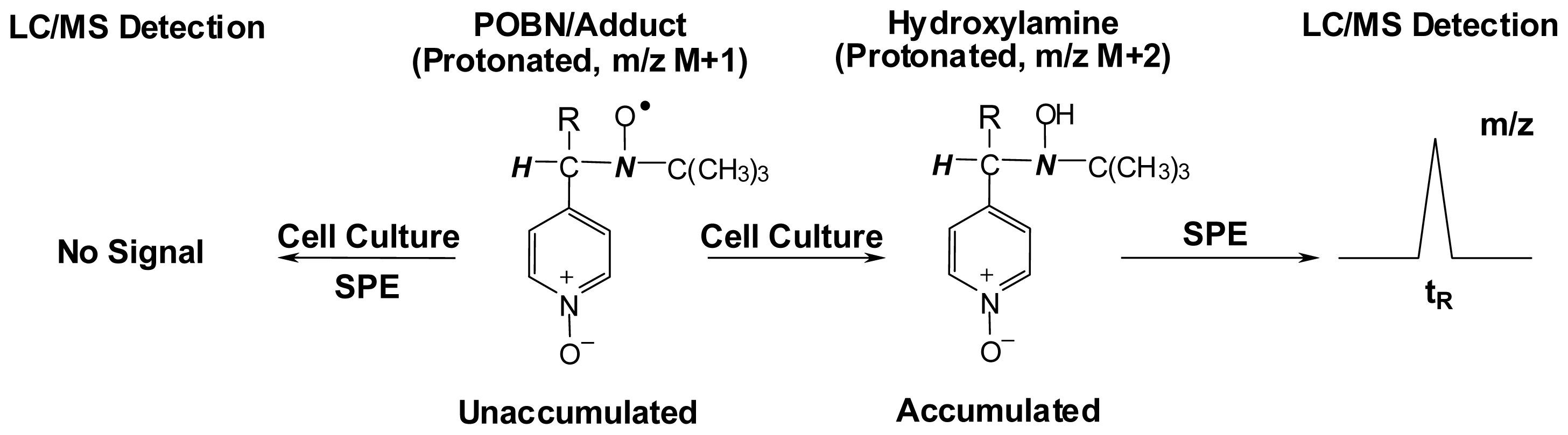
3. Development of the Combined ESR Spin-Trapping and LC/ESR/MS Technique
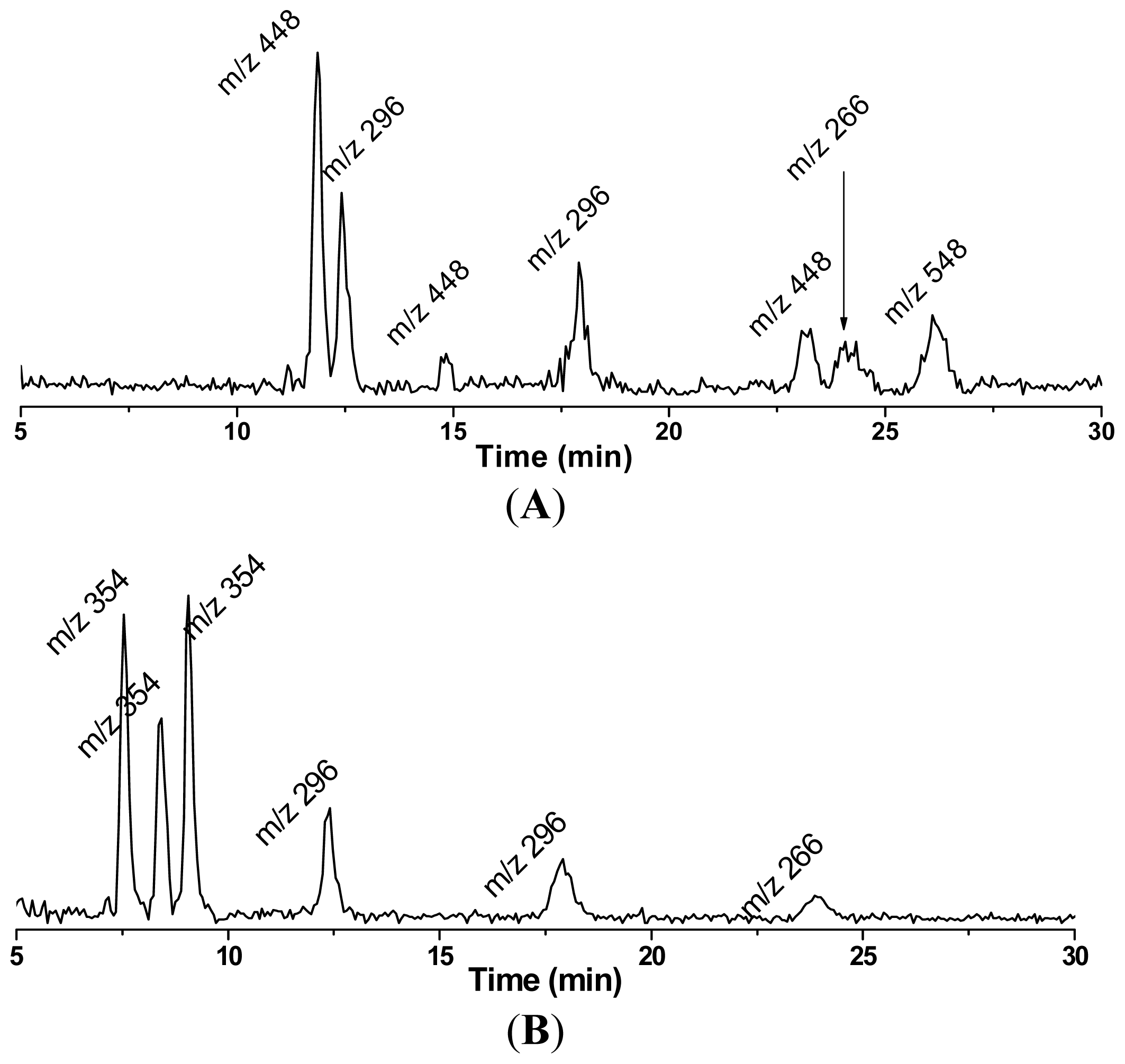
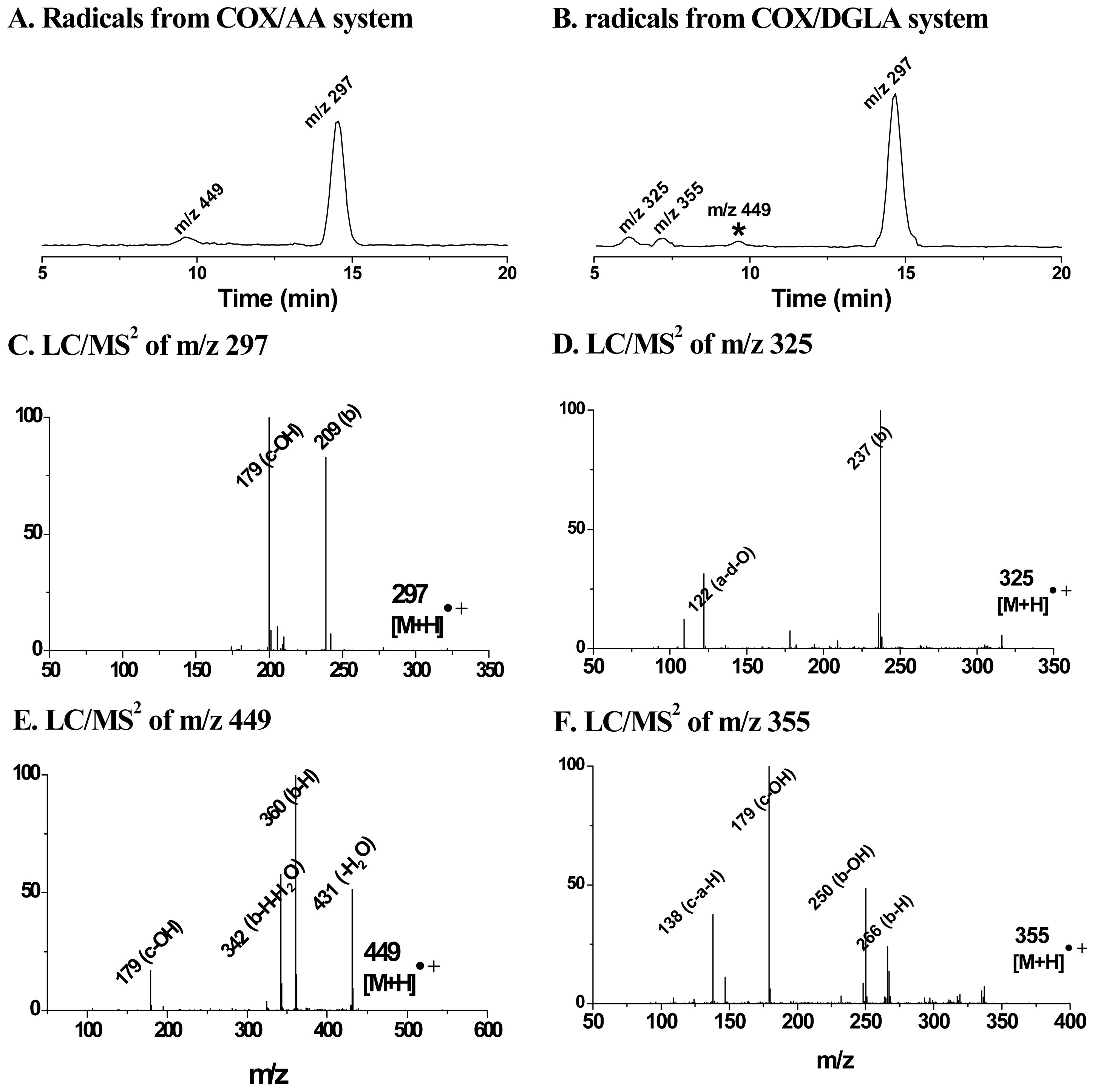
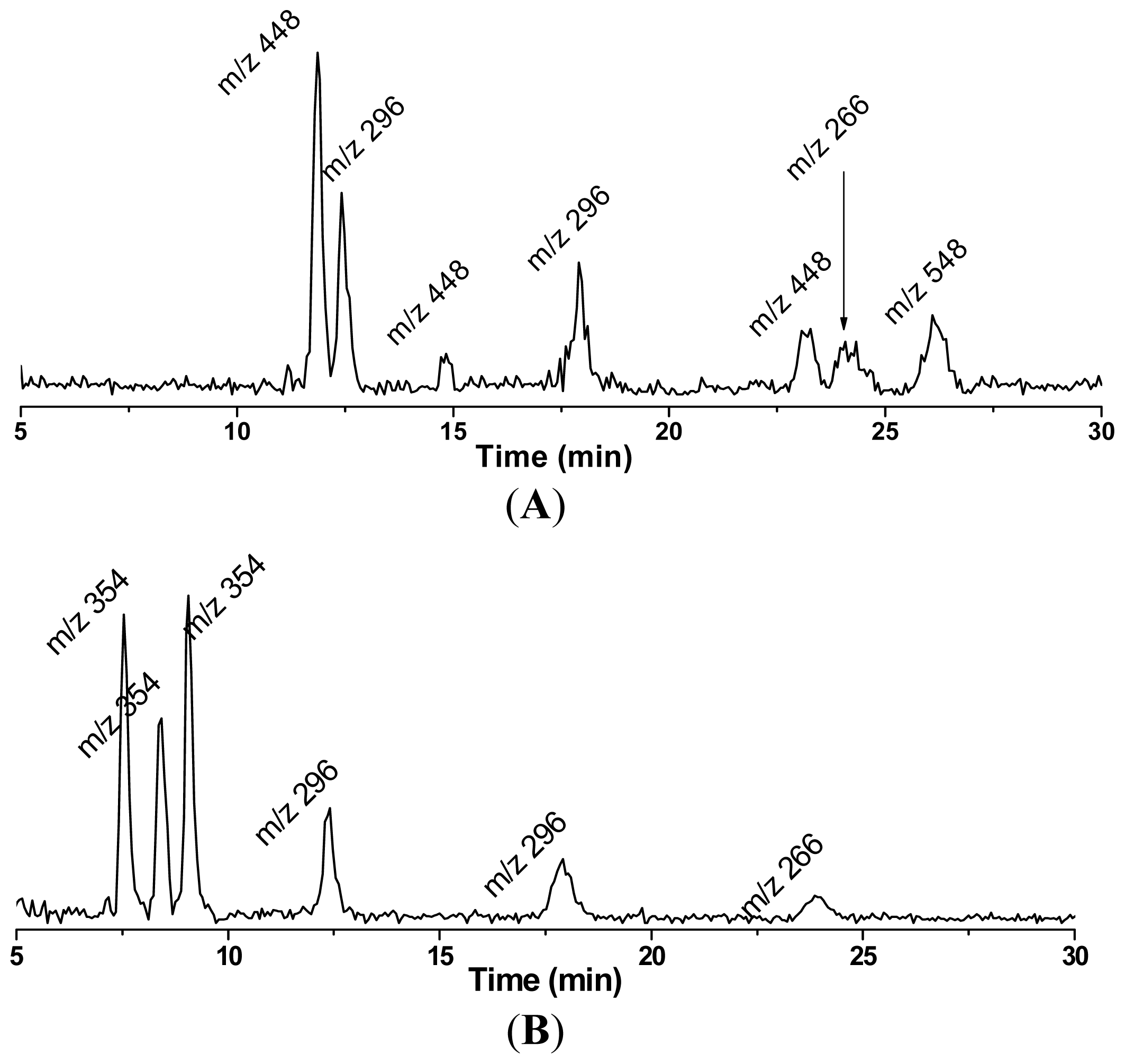
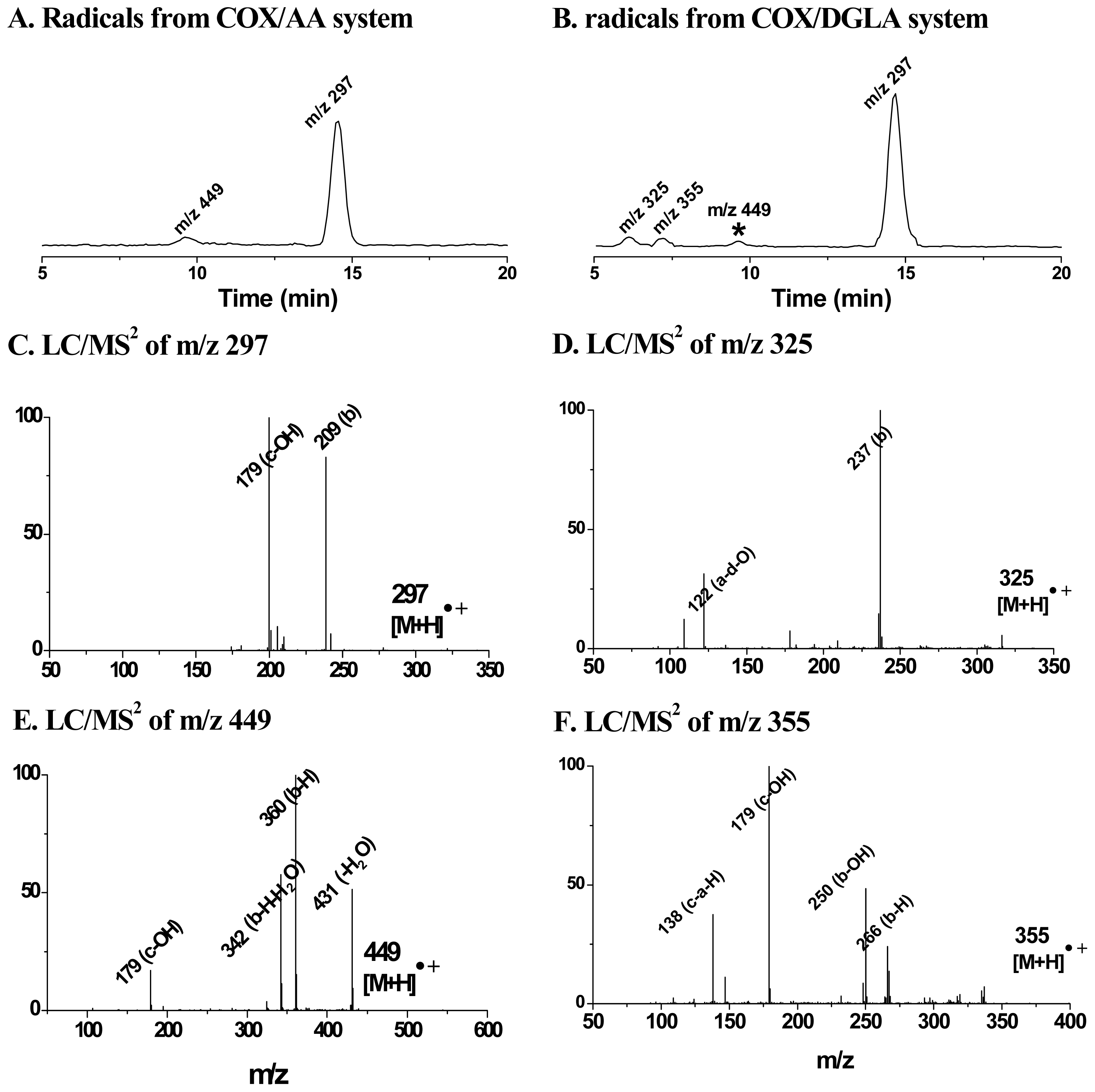
4. Refined Combination of ESR Spin-Trapping and LC/MS Technique in Radical Characterization in in vitro (cell-free) COX-Catalyzed AA and DGLA Peroxidation
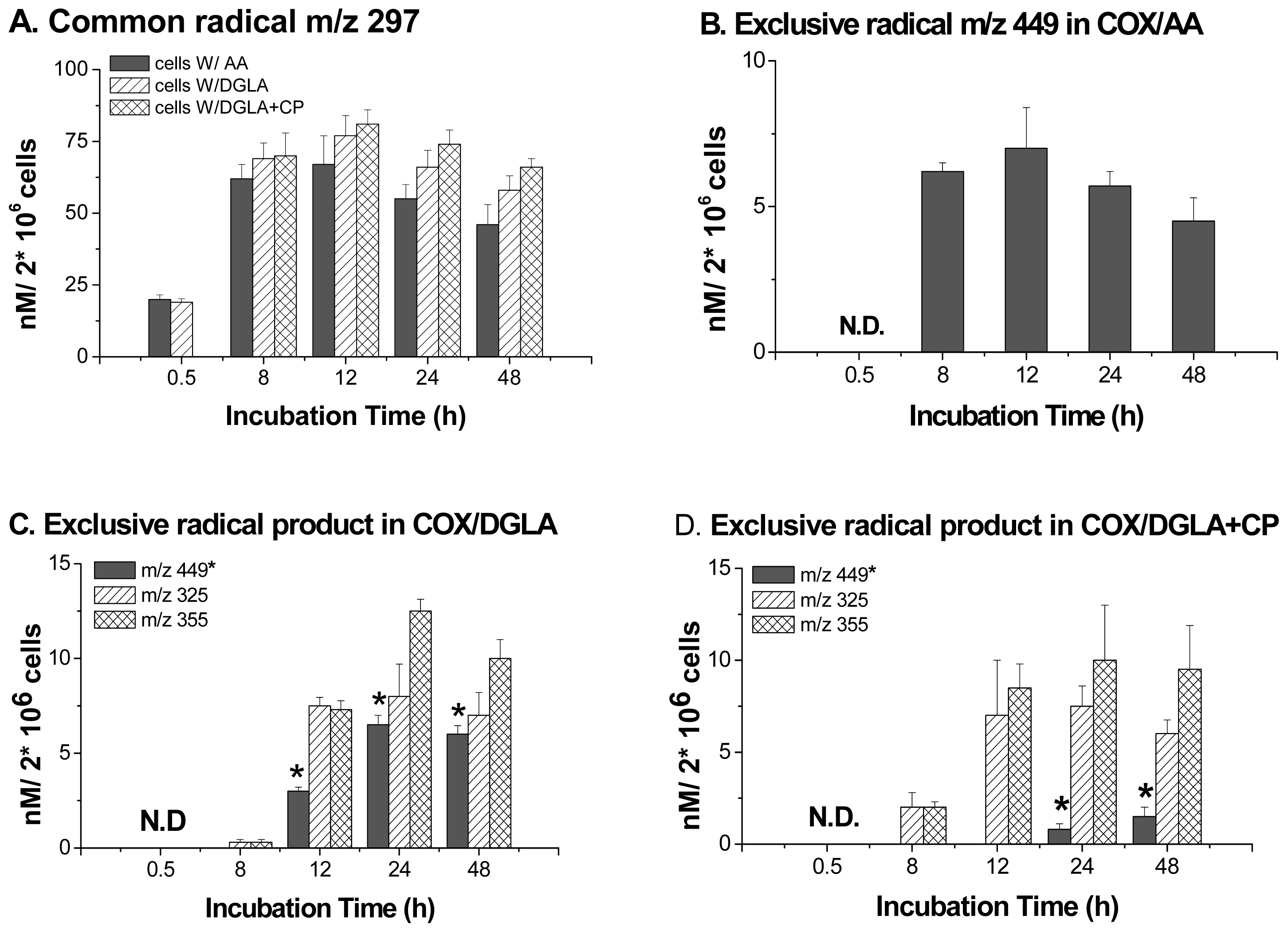
5. Most Recent Update of ESR Spin-Trapping and LC/MS Protocol in Characterization of Free Radicals Formed from Cellular COX-Catalyzed AA and DGLA Peroxidation
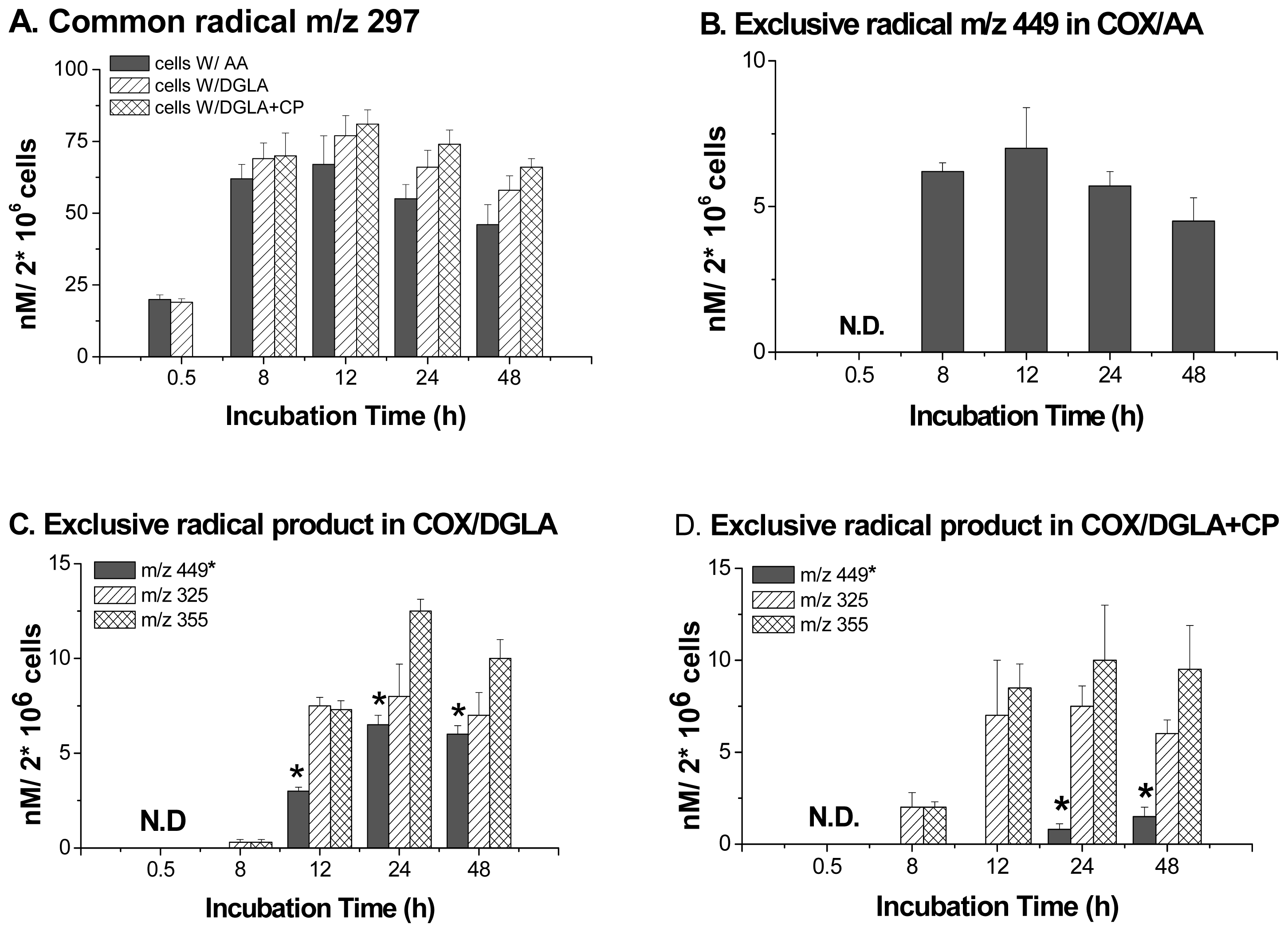
6. Direct Assessment of the Association between Radical Production and Cell Growth Response
7. Summary
Acknowledgements
- Conflict of InterestThe authors declare no conflict of interest.
Abbreviations
| AA | arachidonic acid |
| COX | cyclooxygenase |
| DGLA | dihomo-γ-linolenic acid |
| DHA | docosahexaenoic acid |
| DMPO | 5,5-dimethyl-1-pyrroline N-oxide |
| DPA | docosapentaenoic acid |
| EIC | extracted ion current |
| EPA | eicosapentaenoic acid |
| ESR | electron spin resonance |
| HOAc | acetic acid |
| HPLC (LC) | high performance liquid chromatography |
| LA | linoleic acid |
| LNA | linolenic acid |
| LOX | lipoxygenase |
| MS | mass spectrometry |
| MNP | 2-methyl-2-nitrosopropane |
| NMR | nuclear magnetic resonance |
| PBN | σ-phenyl-tert-butylnitrone |
| POBN | σ-[4-pyridyl-1-oxide]-N-tert-butylnitrone |
| PG | prostaglandin |
| PUFAs | polyunsaturated fatty acids |
| THF | tetrahydrofuran |
| TMPO | 3,3,5,5-tetramethylpyrroline-N-oxide |
| tR | retention time |
References
- Das, U.N. Essential fatty acids: Biochemistry, physiology and pathology. Biotechnol. J 2006, 1, 420–439. [Google Scholar]
- Das, U.N. Essential fatty acids, lipid peroxidation and apoptosis. Prostaglandins Leukot. Essent. Fatty Acids 1999, 61, 157–163. [Google Scholar]
- Sardesai, V.M. Nutritional role of polyunsaturated fatty acids. J. Nutr. Biochem 1992, 3, 154–166. [Google Scholar]
- Gardner, H.W. Oxygen radical chemistry of polyunsaturated fatty acids. Free Radic. Biol. Med 1989, 7, 65–86. [Google Scholar]
- Wagner, B.A.; Buettner, G.R.; Burns, C.P. Free radical-mediated lipid peroxidation in cells: Oxidizability is a function of cell lipid bis-allylic hydrogen content. Biochemistry 1994, 33, 4449–4453. [Google Scholar]
- Girotti, A.W. Mechanisms of lipid peroxidation. Free Radic. Biol. Med 1985, 1, 87–95. [Google Scholar]
- Qian, S.Y.; Wang, H.P.; Schafer, F.Q.; Buettner, G.R. EPR detection of lipid-derived radicals from PUFA, LDL, and cell oxidations. Free Radic. Biol. Med 2000, 29, 568–579. [Google Scholar]
- Porter, N.A. Mechanisms for the autoxidation of polyunsaturated lipids. Accounts Chem. Res 1986, 19, 262–268. [Google Scholar]
- North, J.A.; Spector, A.A.; Buettner, G.R. Detection of lipid radicals by electron paramagnetic resonance spin trapping using intact cells enriched with polyunsaturated fatty acid. J. Biol. Chem 1992, 267, 5743–5746. [Google Scholar]
- Smith, W.L.; DeWitt, D.L.; Garavito, R.M. Cyclooxygenases: Structural, cellular, and molecular biology. Annu. Rev. Biochem 2000, 69, 145–182. [Google Scholar]
- Marnett, L.J.; Rowlinson, S.W.; Goodwin, D.C.; Kalgutkar, A.S.; Lanzo, C.A. Arachidonic acid oxygenation by COX-1 and COX-2. J. Biol. Chem 1999, 274, 22903–22906. [Google Scholar]
- Yamamoto, S. Mammalian lipoxygenases: Molecular structures and functions. Biochim. Biophys. Acta 1992, 1128, 117–131. [Google Scholar]
- Sano, H.; Kawahito, Y.; Wilder, R.L.; Hashiramoto, A.; Mukai, S.; Asai, K.; Kimura, S.; Kato, H.; Kondo, M.; Hla, T. Expression of cyclooxygenase-1 and -2 in human colorectal cancer. Cancer Res 1995, 55, 3785–3789. [Google Scholar]
- Wolff, H.; Saukkonen, K.; Anttila, S.; Antti, K.; Harri, V.; Ristimaki, A. Expression of cyclooxygenase-1 in human lung carcinoma. Cancer Res 1998, 58, 4997–5001. [Google Scholar]
- Buskens, C.J.; van Rees, B.P.; Sivula, A.; Reitsma, J.B.; Haglund, C.; Bosma, P.J.; Offerhaus, G.J.A.; van Lanschot, J.J.B.; Ristimaki, A. Prognostic significance of elevated cyclooxygenase 2 expression in patients with adenocarcinoma of the esophagus. Gastroenterology 2002, 122, 1800–1807. [Google Scholar]
- Sales, K.D.; Katz, A.A.; Davis, M.; Hinz, S.; Soeters, R.P.; Hofmeyr, M.D.; Millar, R.P.; Jabbour, H.N. Cyclooxygenase-2 expression and prostaglandin E2 synthesis are upregulated in carcinomas of the cervix: A possible autocrine/paracrine regulation of neoplastic cell function via EP2/EP4 receptor. J. Clin. Endocrinol. Metab 2001, 86, 2243–2249. [Google Scholar]
- Lee, L.M.; Pan, C.C.; Cheng, C.J.; Chi, C.W.; Liu, T.Y. Expression of cyclooxygenase-2 in prostate adenocarcinoma and benign prostatic hyperplasia. Anticancer Res 2001, 21, 1291–1294. [Google Scholar]
- Rouzer, C.A.; Marnett, L.J. Structural and functional differences between cyclooxygenases: Fatty acid oxygenases with a critical role in cell signaling. Biochem. Biophys. Res. Commun 2005, 338, 34–44. [Google Scholar]
- Pidgeon, G.P.; Lysaght, J.; Krishnamoorthy, S.; Reynolds, J.V.; O’Byrne, K.; Nie, D.; Honn, K.V. Lipoxygenase metabolism: Roles in tumor progression and survival. Cancer Metastasis Rev 2007, 26, 503–524. [Google Scholar]
- Vang, K.; Ziboh, V.A. 15-lipoxygenase metabolites of γ-linolenic acid/eicosapentaenoic acid suppress growth and arachidonic acid metabolism in human prostatic adenocarcinoma cells: Possible implications of dietary fatty acids. Prostaglandins Leukot. Essent. Fatty Acids 2005, 72, 363–372. [Google Scholar]
- Das, U.N. Nutrients, essential fatty acids and prostaglandins interact to augment immune responses and prevent genetic damage and cancer. Nutrition 1989, 5, 106–110. [Google Scholar]
- Porter, N.A.; Funk, M.O. Peroxy radical cyclization as a model for prostaglandin biosynthesis. J. Org. Chem 1975, 40, 3614–3615. [Google Scholar]
- Fan, Y.Y.; Ramos, K.S.; Chapkin, R.S. Cell cycle related inhibition of mouse vascular smooth muscle cell proliferation by prostaglandin E1: Relationship between prostaglandin E1 and intracellular cAMP levels. Prostaglandins Leukot. Essent. Fatty Acids 1996, 54, 101–107. [Google Scholar]
- Gianetti, J.; de Caterina, M.; de Cristofaro, T.; Ungaro, B.; del Guercio, R.; de Caterina, R. Intravenous prostaglandin E1 reduces soluble vascular cell adhesion molecule-1 in peripheral arterial obstructive disease. Am. Heart J 2001, 142, 733–739. [Google Scholar]
- Pham, H.; Vang, K.; Ziboh, V.A. Dietary γ-linolenate attenuates tumor growth in a rodent model of prostatic adenocarcinoma via suppression of elevated generation of PGE2 and 5S-HETE. Prostaglandins Leukot. Essent. Fatty Acids 2006, 74, 271–282. [Google Scholar]
- James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary polyunsaturated fatty acids and inflammatory mediator production. Am. J. Clin. Nutr 2000, 71, 343S–348S. [Google Scholar]
- Fan, Y.Y.; Chapkin, R.S. Importance of dietary γ-linolenic acid in human health and nutrition. J. Nutr 1998, 128, 1411–1414. [Google Scholar]
- Takai, S.; Jin, D.; Kawashima, H.; Kimura, M.; Shiraishi-Tateishi, A.; Tanaka, T.; Kakutani, S.; Tanaka, K.; Kiso, Y.; Miyazaki, M. Anti-atherosclerotic effects of dihomo-γ-linolenic acid in ApoE-deficient mice. J. Atheroscler. Thromb 2009, 16, 480–489. [Google Scholar]
- Menendez, J.A.; del Mar Barbacid, M.; Montero, S.; Sevilla, E.; Escrich, E.; Solanas, M.; Cortes-Funes, H.; Colomer, R. Effects of γ-linolenic acid and oleic acid on paclitaxel cytotoxicity in human breast cancer cells. Eur. J. Cancer 2001, 37, 402–413. [Google Scholar]
- Henniga, B.; Lei, W.; Arzuaga, X.; Ghosha, D.D.; Saraswathia, V.; Toborekc, M. Linoleic acid induces proinflammatory events in vascular endothelial cells via activation of PI3K/Akt and ERK1/2 signaling. J. Nutr. Biochem 2006, 17, 766–772. [Google Scholar]
- Rose, D.P. Dietary fat, fatty acids and breast cancer. Breast Cancer 1997, 4, 7–16. [Google Scholar]
- Tsuyoshi, T.; Igarashi, M.; Miyazawa, T. Conjugated eicosapentaenoic acid (EPA) inhibits transplanted tumor growth via membrane lipid peroxidation in nude mice. J. Nutr 2004, 134, 1162–1166. [Google Scholar]
- Wen, B.; Deutsch, E.; Opolon, P.; Auperin, A.; Frascognal, V.; Connault, E.; Bourhis, J. n-3 Polyunsaturated fatty acids decrease mucosal/epidermal reactions and enhance antitumour effect of ionising radiation with inhibition of tumour angiogenesis. Br. J. Cancer 2003, 89, 1102–1107. [Google Scholar]
- Kuriki, K.; Hirose, K.; Wakai, K.; Matsuo, K.; Ito, H.; Suzuki, T.; Hiraki, A.; Saito, T.; Iwata, H.; Tatematsu, M.; et al. Breast cancer risk and erythrocyte compositions of n-3 highly unsaturated fatty acids in Japanese. Int. J. Cancer 2007, 121, 377–385. [Google Scholar]
- Geelen, A.; Schouten, J.M.; Kamphuis, C.; Stam, B.E.; Burema, J.; Renkema, J.M.; Bakker, E.J.; van’t Veer, P.; Kampman, E. Fish consumption, n-3 fatty acids, and colorectal cancer: A meta-analysis of prospective cohort studies. Am. J. Epidemiol 2007, 166, 1116–1125. [Google Scholar]
- Kokura, S.; Nakagawa, S.; Hara, T.; Boku, Y.; Naito, Y.; Yoshida, N.; Yoshikawa, T. Enhancement of lipid peroxidation and of the antitumor effect of hyperthermia upon combination with oral eicosapentaenoic acid. Cancer Lett 2002, 185, 139–144. [Google Scholar]
- Janzen, E.G.; Blackburn, B.J. Detection and identification of short-lived free radicals by electron spin resonance trapping techniques (spin trapping). Photolysis of organolead, -tin, and -mercury compounds. J. Am. Chem. Soc 1969, 91, 4481–4490. [Google Scholar]
- Janzen, E.G.; Gerlock, J.L. Substituent effects on the photochemistry and nitroxide radical formation of nitro aromatic compounds as studied by electron spin resonance spin-trapping techniques. J. Am. Chem. Soc 1969, 91, 3108–3109. [Google Scholar]
- Janzen, E.G. Spin trapping. Accounts Chem. Res 1971, 4, 31–40. [Google Scholar]
- Lagercrantz, C. Spin trapping of some short-lived radicals by the nitroxide method. J. Phys. Chem 1971, 75, 3466–3475. [Google Scholar]
- Dikalova, A.E.; Kadiiska, M.B.; Mason, R.P. An in vivo ESR spin-trapping study: Free radical generation in rats from formate intoxication—Role of the Fenton reaction. Proc. Natl. Acad. Sci. USA 2001, 98, 13549–13553. [Google Scholar]
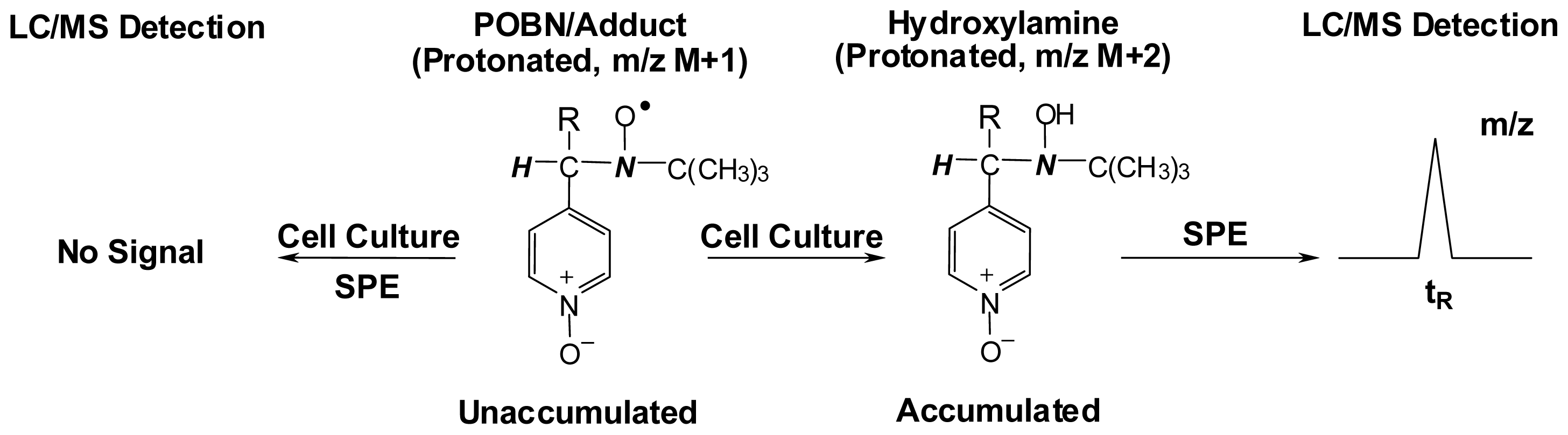
- Qian, S.Y.; Kadiiska, M.B.; Guo, Q.; Mason, R.P. A novel protocol to identify and quantify all spin trapped free radicals from in vitro/in vivo interaction of HO and DMSO: LC/ESR, LC/MS, and dual spin trapping combinations. Free Radic. Biol. Med 2005, 38, 125–135. [Google Scholar]
- Pazos, M.; Andersen, M.L.; Skibsted, L.H. Heme-mediated production of free radicals via preformed lipid hydroperoxide fragmentation. J. Agric. Food Chem 2008, 56, 11478–11484. [Google Scholar]
- Spulber, M.; Schlick, S. Using cyclodextrins to encapsulate oxygen-centered and carbon-centered radical adducts: The case of DMPO, PBN, and MNP spin traps. J. Phys. Chem. A 2010, 114, 6217–6225. [Google Scholar]
- Guo, Q.; Qian, S.Y.; Mason, R.P. Separation and identification of DMPO adducts of oxygen-centered radicals formed from organic hydroperoxides by HPLC-ESR, ESI-MS and MS/MS. J. Am. Soc. Mass Spectrom 2003, 14, 862–871. [Google Scholar]
- Chamulitrat, W.; Takahashis, N.; Mason, R.P. Peroxyl, alkoxyl, and carbon-centered radical formation from organic hydroperoxides by chloroperoxidase. J. Biol. Chem 1989, 264, 7889–7899. [Google Scholar]
- Bose-Basu, B.; Derose, E.F.; Chen, Y.R.; Mason, R.P.; London, R.E. Protein NMR spin trapping with [methyl-13C3]-MNP: Application to the tyrosyl radical of equine myoglobinm. Free Radic. Biol. Med 2001, 31, 383–390. [Google Scholar]
- Qian, S.Y.; Chen, Y.R.; Deterding, L.J.; Fann, Y.C.; Chignell, C.F.; Tomer, K.B.; Mason, R.P. Identification of protein-derived tyrosyl radical in the reaction of cytochrome c and hydrogen peroxide: Characterization by ESR spin-trapping, HPLC and MS. Biochem. J 2002, 363, 281–288. [Google Scholar]
- Gunther, M.R.; Tschirret-Guth, R.A.; Witkowska, H.E.; Fann, Y.C.; Barr, D.P.; Ortiz De Montellano, P.R.; Mason, R.P. Site-specific spin trapping of tyrosine radicals in the oxidation of metmyoglobin by hydrogen peroxide. Biochem. J 1998, 330, 1293–1299. [Google Scholar]
- Barr, D.P.; Gunther, M.R.; Deterding, L.J.; Tomer, K.B.; Mason, R.P. ESR Spin-trapping of a protein-derived tyrosyl radical from the reaction of cytochrome c with hydrogen peroxide. J. Biol. Chem 1996, 271, 15498–15503. [Google Scholar]
- Lardinois, O.M.; Detweiler, C.D.; Tomer, K.B.; Mason, R.P.; Deterding, L.J. Identifying the site of spin-trapping in proteins by a combination of liquid chromatography, ELISA and off-line tandem mass spectrometry. Free Radic. Biol. Med 2008, 44, 893–906. [Google Scholar]
- Bhattacharjee, S.; Deterding, L.J.; Chatterjee, S.; Jiang, J.J.; Ehrenshaft, M.; Lardinois, O.; Ramirez, D.C.; Tomer, K.B.; Mason, R.P. Site-specific radical formation in DNA induced by Cu(II)-H2O2 oxidizing system, using ESR, immuno-spin trapping, LC-MS and MS/MS. Free Radic. Biol. Med 2011, 50, 1536–1545. [Google Scholar]
- Chatterjee, S.; Lardinois, O.; Bhattacharjee, S.; Tucker, J.; Corbett, J.; Deterding, L.; Ehrenshaft, M.; Bonini, M.G.; Mason, R.P. Oxidative stress induces protein and DNA radical formation in follicular dendritic cells of the germinal center and modulates its cell death patterns in late sepsis. Free Radic. Biol. Med 2011, 50, 988–999. [Google Scholar]
- Buettner, G.R.; Kelley, E.E.; Burns, C.P. Membrane lipid free radicals produced from L1210 murine leukemia cells by photofrin photosensitization: An electron paramagnetic resonance spin trapping study. Cancer Res 1993, 53, 3670–3673. [Google Scholar]
- Qian, S.Y.; Buettner, G.R. Iron and dioxygen chemistry is an important route to initiation of biological free radical oxidations: An electron paramagnetic resonance spin trapping study. Free Radic. Biol. Med 1999, 26, 1447–1456. [Google Scholar]
- Kadiiska, M.B.; Hanna, P.M.; Jordan, S.J.; Mason, R.P. Electron spin resonance evidence for free radical generation in copper-treated vitamin E- and selenium-deficient rats: In vivo spin-trapping investigation. Mol. Pharmacol 1993, 44, 222–227. [Google Scholar]
- Kadiiska, M.B.; Mason, R.P.; Dreher, K.L.; Costa, D.L.; Ghio, A.J. In vivo evidence of free radical formation in the rat lung after exposure to an emission source air pollution particle. Chem. Res. Toxicol 1997, 10, 1104–1108. [Google Scholar]
- Kadiiska, M.B.; Morrow, J.D.; Awad, J.A.; Roberts, L.J.; Mason, R.P. Identification of free radical formation and F2- isoprostanes in vivo by acute Cr(VI) poisoning. Chem. Res. Toxicol 1998, 11, 1516–1520. [Google Scholar]
- Ghio, A.J.; Kadiiska, M.B.; Xiang, Q.H.; Mason, R.P. In vivo evidence of free radical formation after asbestos instillation: An ESR spin trapping investigation. Free Radic. Biol. Med 1998, 24, 11–17. [Google Scholar]
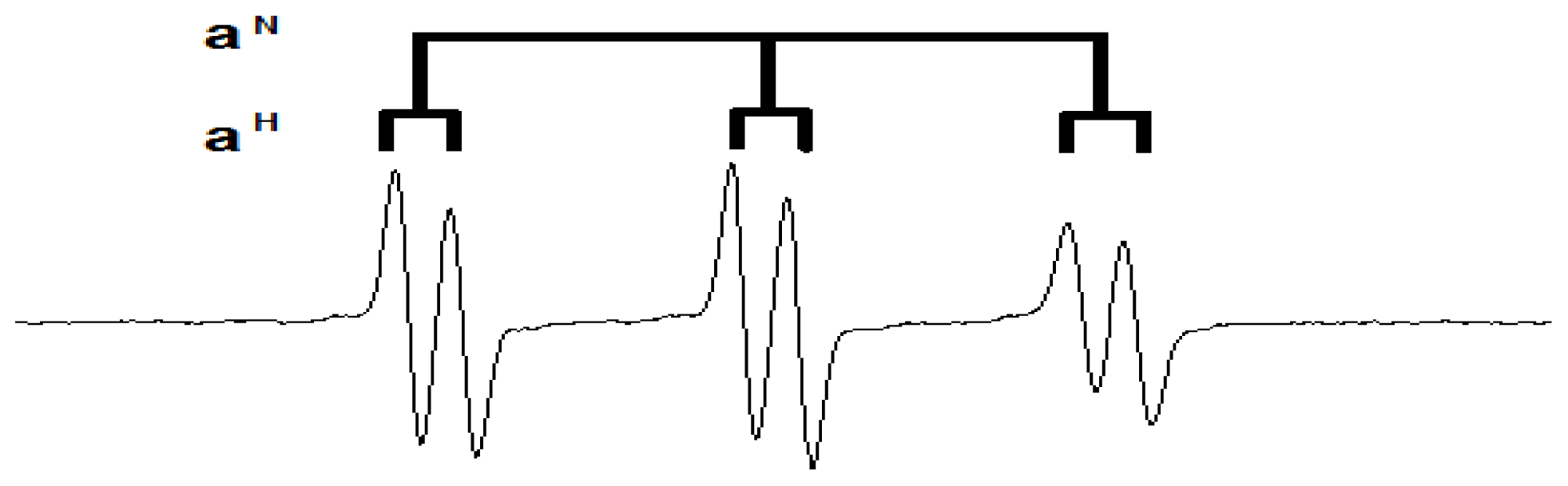
- Buettner, G.R. Spin trapping: EPR parameters of spin adducts. Free Radic. Biol. Med 1987, 3, 259–303. [Google Scholar]
- Iwahashi, H.; Parker, C.E.; Mason, R.P.; Tomer, K.B. Radical adducts of nitrosobenzene and 2-methyl-2-nitrosopropane with 12,13-epoxylinoleic acid radical, 12,13-epoxylinolenic acid radical and 14,15-epoxyarachidonic acid radical. Biochem. J 1991, 276, 447–453. [Google Scholar]
- Iwahashi, H.; Deterding, L.J.; Parker, C.E.; Mason, R.P.; Tomer, K.B. Identification of radical adducts formed in the reactions of unsaturated fatty acids with soybean lipoxygenase using continuous flow fast atom bombardment with tandem mass spectrometry. Free Radic. Res 1996, 25, 255–274. [Google Scholar]
- Qian, S.Y.; Tomer, K.B.; Yue, G.H.; Guo, Q.; Kadiiska, M.B.; Mason, R.P. Characterization of the initial carbon-centered pentadienyl radical and subsequent radicals in lipid peroxidation: Identification via on-line high performance liquid chromatography/electron spin resonance and mass spectrometry. Free Radic. Biol. Med 2002, 33, 998–1009. [Google Scholar]
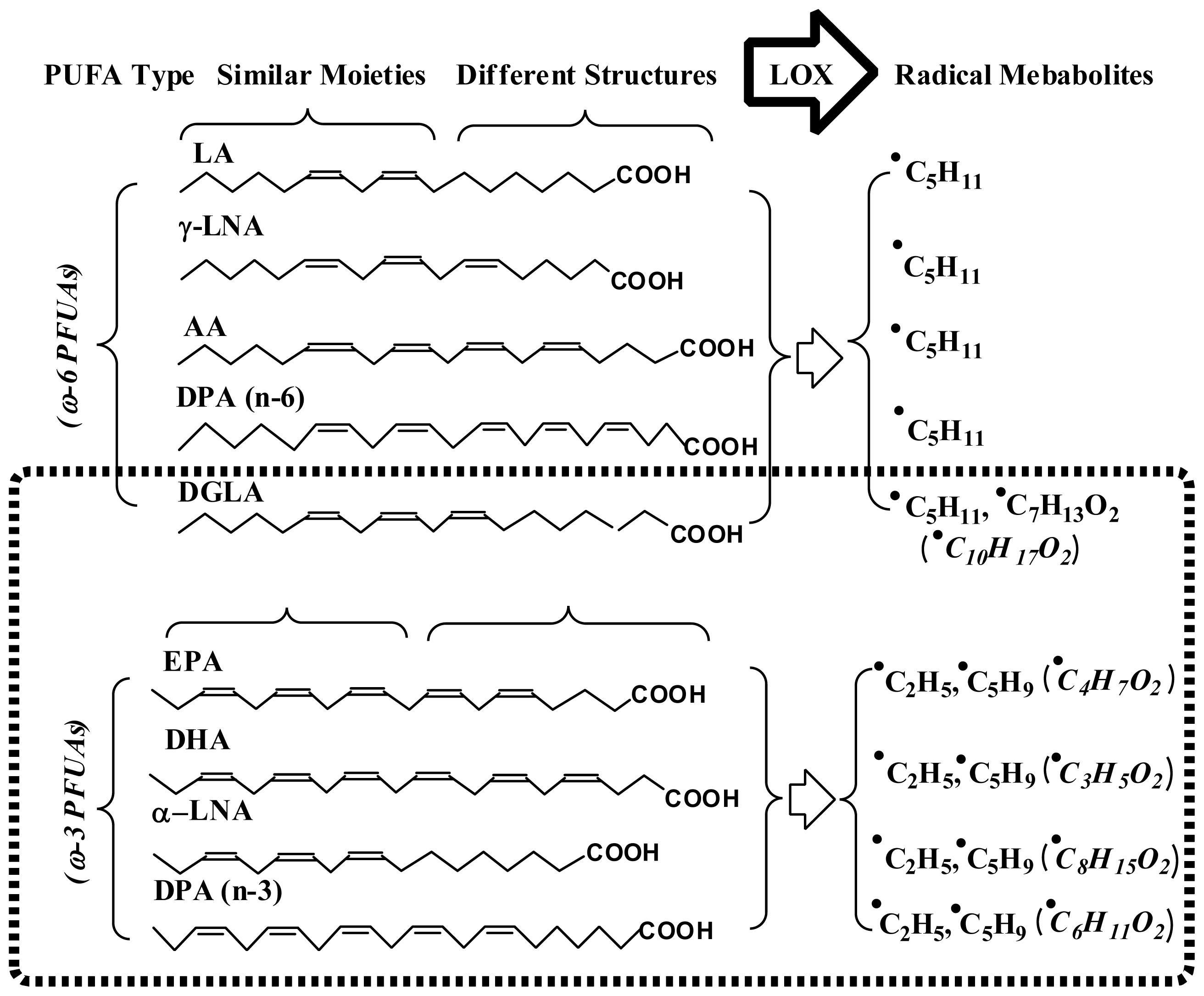
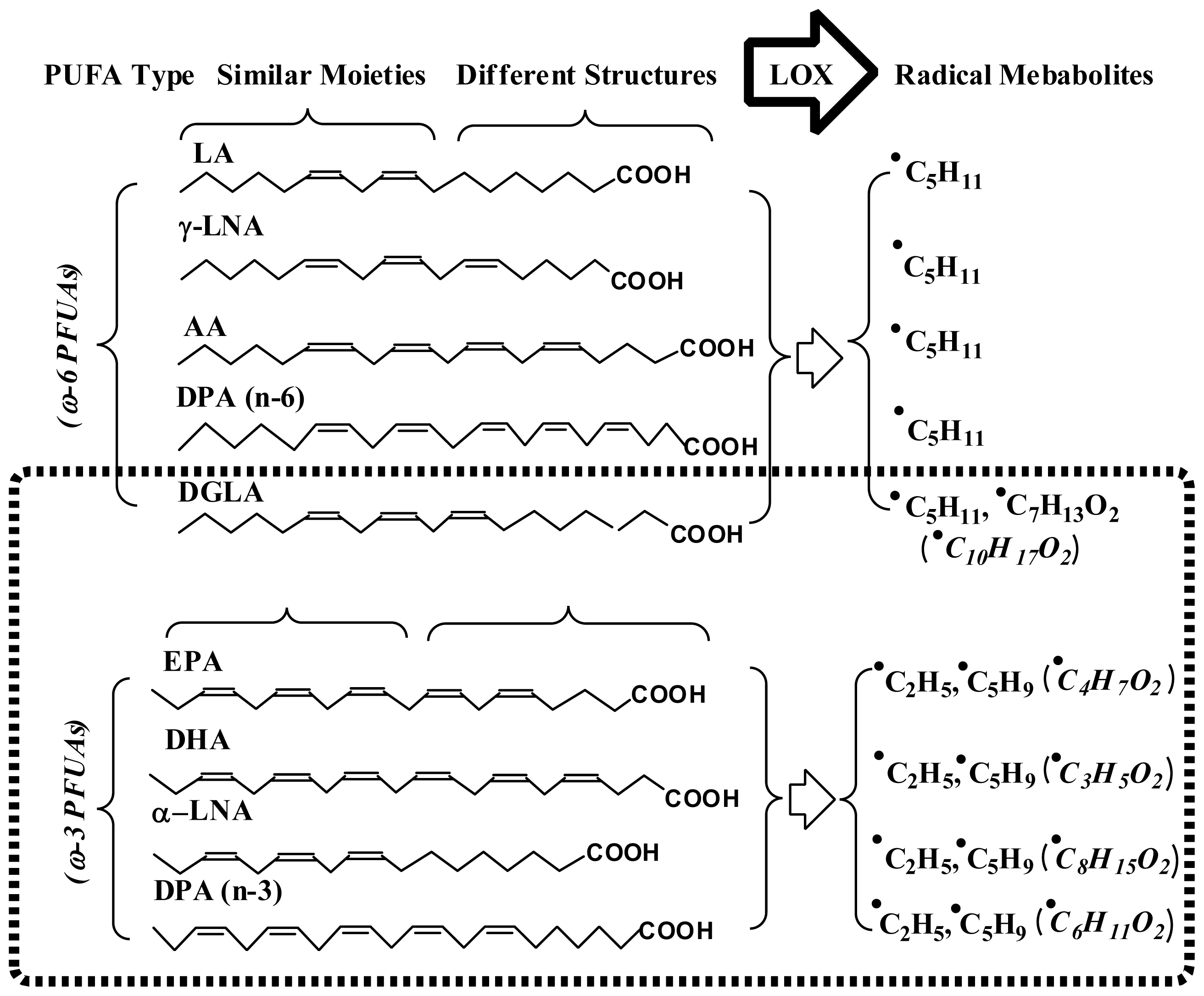
- Qian, S.Y.; Yue, G.H.; Tomer, K.B.; Mason, R.P. Identification of all classes of spin trapped carbon-centered radicals in soybean lipoxygenase-dependent lipid peroxidation of ω-6 polyunsaturated fatty acids via LC/ESR, LC/MS, and tandem MS. Free Radic. Biol. Med 2003, 34, 1017–1028. [Google Scholar]
- Qian, S.Y.; Guo, Q.; Mason, R.P. Identification of spin trapped carbon-centered radicals in soybean lipoxygenase-dependent peroxidation of ω-3 polyunsaturated fatty acids by LC/ESR, LC/MS, and tandem MS. Free Radic. Biol. Med 2003, 35, 33–44. [Google Scholar]
- Yu, Q.; Shan, Z.; Ni, K.; Qian, S.Y. LC/ESR/MS study of spin trapped carbon-centered radicals formed from in vitro lipoxygenase-catalyzed peroxidation of γ-linolenic acid. Free Radic. Res 2008, 42, 442–455. [Google Scholar]
- Shan, Z.; Yu, Q.; Purwaha, P.; Guo, B.; Qian, S.Y. A combination study of spin-trapping, LC/ESR and LC/MS on carbon-centred radicals formed from lipoxygenase-catalysed peroxidation of eicosapentaenoic acid. Free Radic. Res 2008, 43, 1–15. [Google Scholar]
- Purwaha, P.; Gu, Y.; Kelavkar, U.; Kang, J.X.; Law, B.; Wu, E.; Qian, S.Y. LC/ESR/MS study of pH-dependent radical generation from 15-LOX-catalyzed DPA peroxidation. Free Radic. Biol. Med 2011, 51, 1461–1470. [Google Scholar]
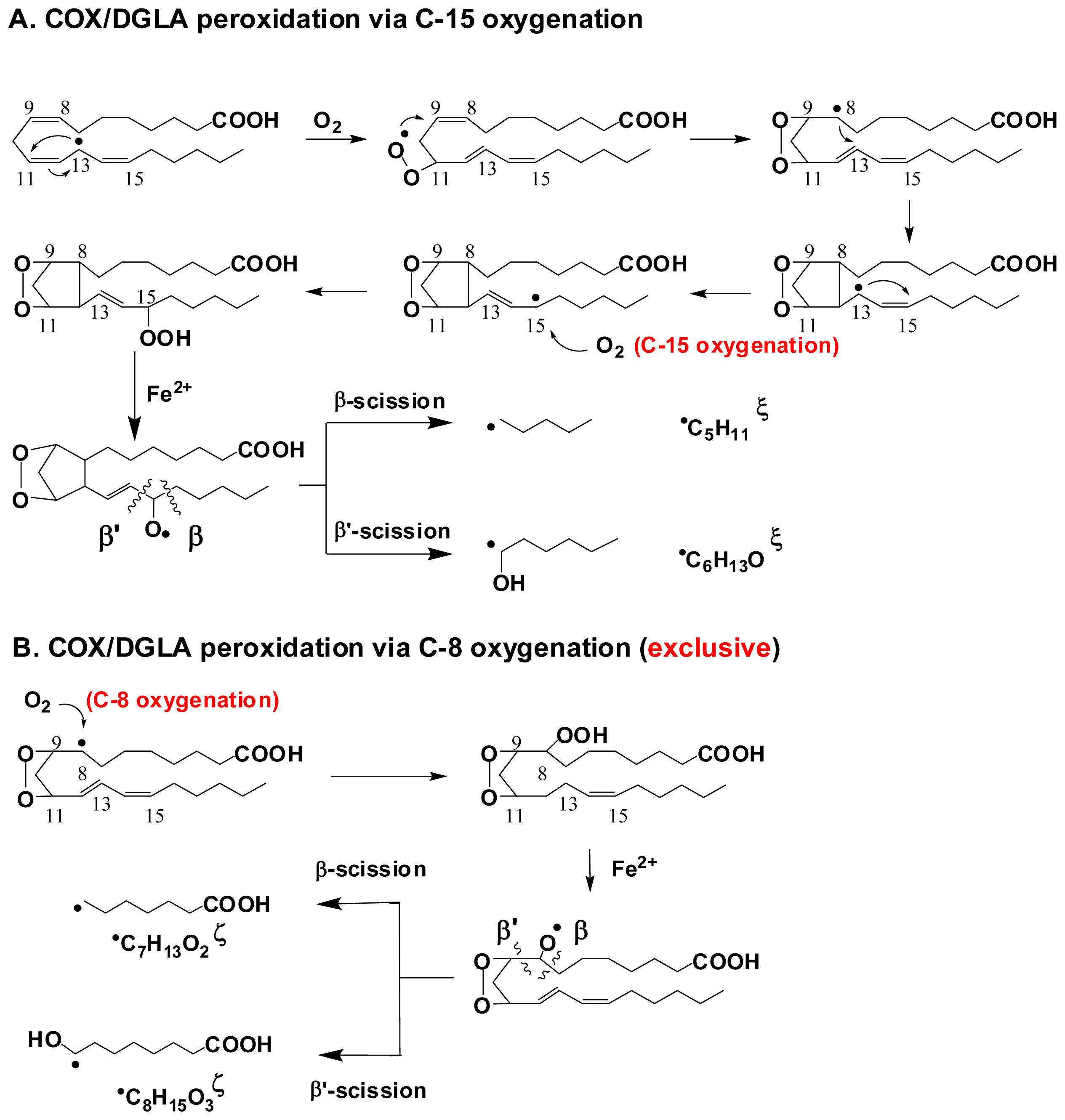
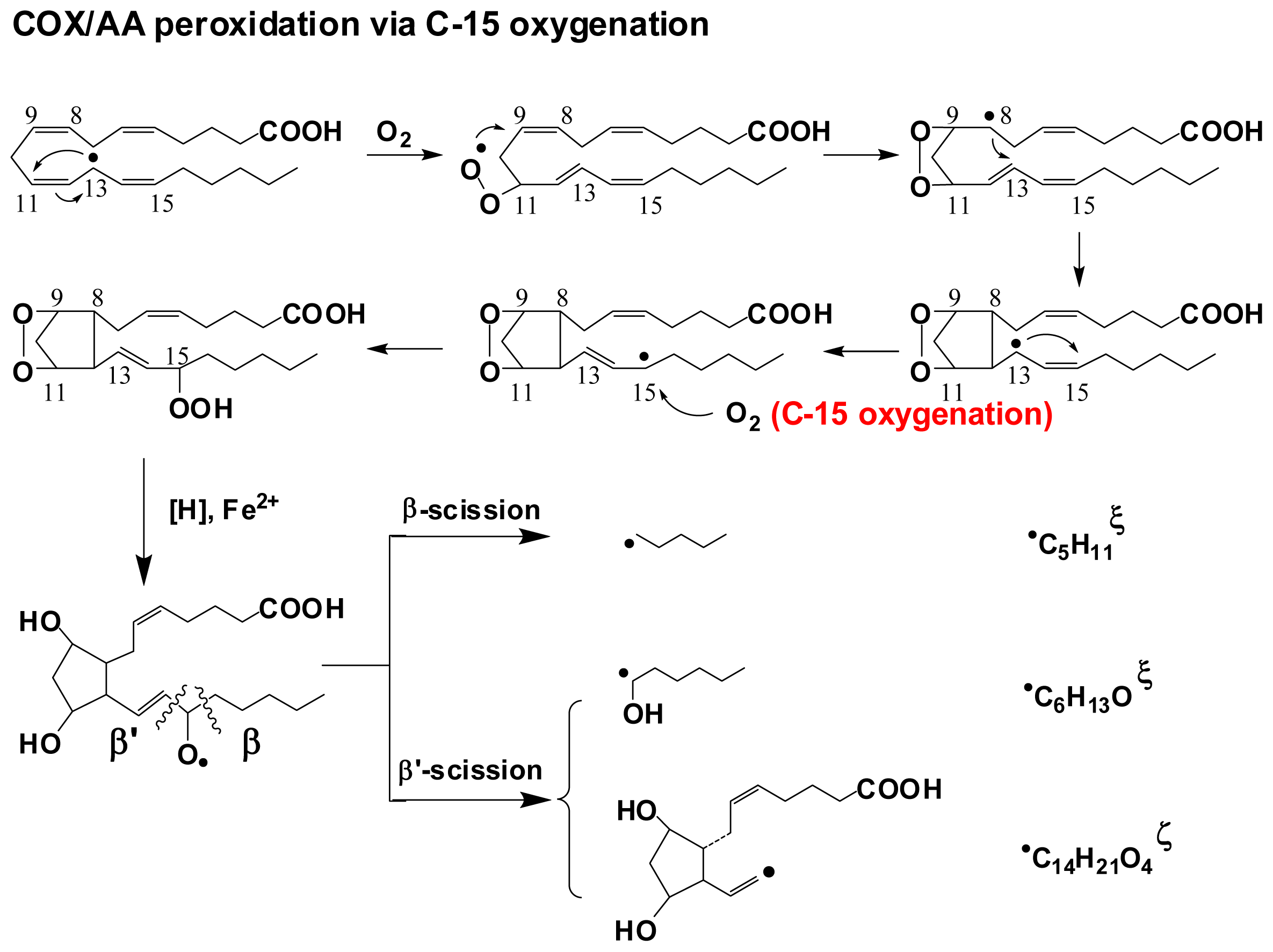
- Yu, Q.; Purwaha, P.; Ni, K.; Sun, C.; Mallik, S.; Qian, S.Y. Characterization of novel radicals from COX-catalyzed arachidonic acid peroxidation. Free Radic. Biol. Med 2009, 47, 568–576. [Google Scholar]
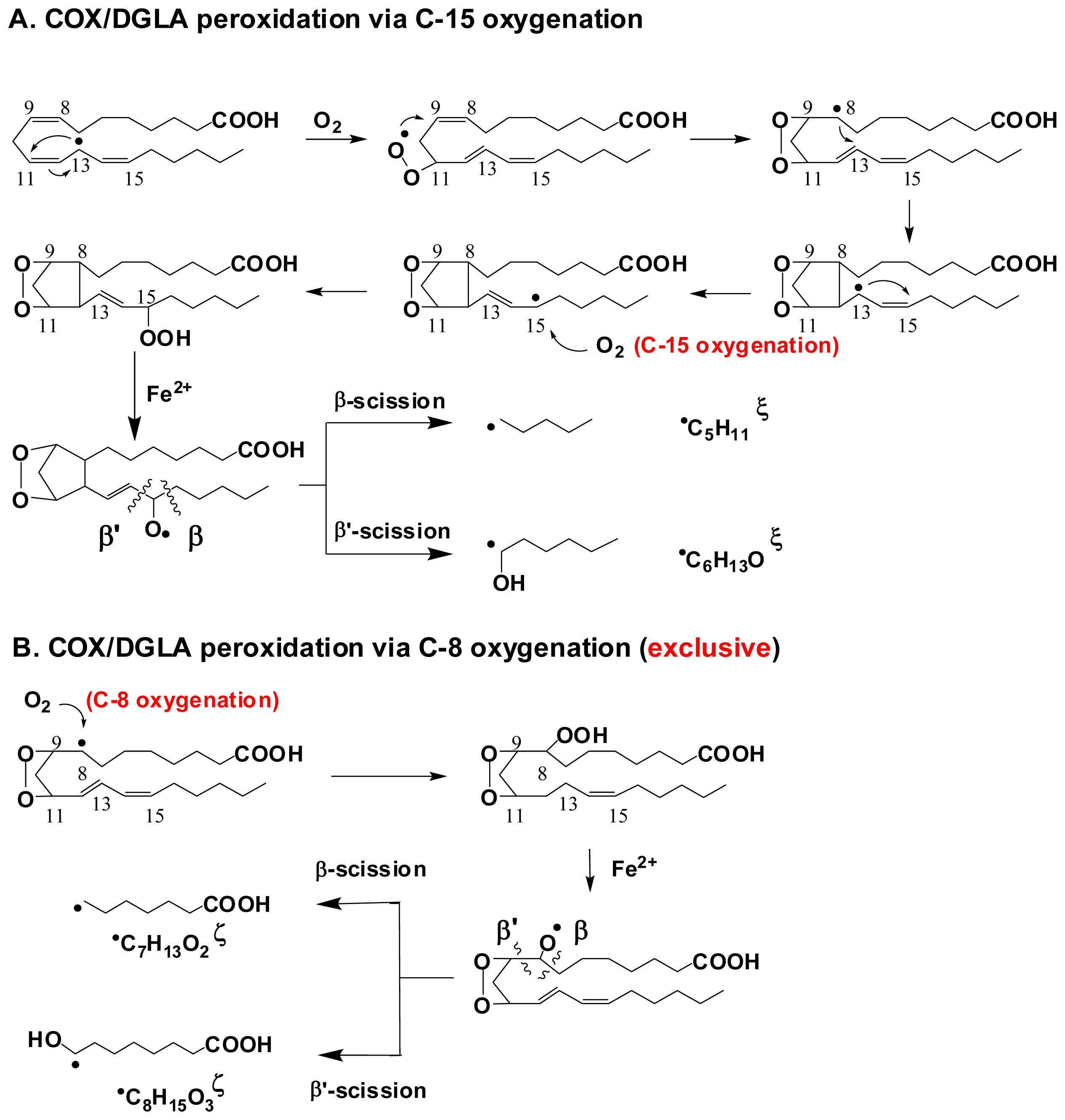
- Xiao, Y.; Gu, Y.; Purwaha, P.; Ni, K.; Law, B.; Mallik, S.; Qian, S.Y. Characterization of free radicals formed from COX-catalyzed DGLA peroxidation. Free Radic. Biol. Med 2011, 50, 1163–1170. [Google Scholar]
- Gu, Y.; Xu, Y.; Law, B.; Qian, S.Y. The first characterization of free radicals formed from cellular COX-catalyzed peroxidation. Free Radic. Biol. Med. 2012, in press. [Google Scholar]
- Miller, C.C.; McCreedy, C.A.; Jones, A.D.; Ziboh, V.A. Oxidative metabolism of dihomo-gammalinolenic acid by guinea pig epidermis: Evidence of generation of anti-inflammatory products. Prostaglandins 1988, 35, 917–938. [Google Scholar]
- Gago-Dominguez, M.; Yuan, J.M.; Sun, C.L.; Lee, H.P.; Yu, M.C. Opposing effects of dietary n-3 and n-6 fatty acids on mammary carcinogenesis: The Singapore Chinese health study. Br. J. Cancer 2003, 89, 1686–1692. [Google Scholar]
- Leitzmann, M.F.; Stampfer, M.J.; Michaud, D.S.; Augustsson, K.; Colditz, G.C.; Willett, W.C.; Giovannucci, E.L. Dietary intake of n-3 and n-6 fatty acids and the risk of prostate cancer. Am. J. Clin. Nutr 2004, 80, 204–216. [Google Scholar]
- Larsson, S.C.; Kumlin, M.; Ingelman-Sundberg, M.; Wolk, A. Dietary long-chain n-3 fatty acids for the prevention of cancer: A review of potential mechanisms. Am. J. Clin. Nutr 2004, 79, 935–945. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells (HCA-7 Colony 29) Cultured with | Cell G2 Arrest | Cell Proliferation | |
|---|---|---|---|
| % in G2 Phase | % Viability | ||
| 8 h | 24 h | 48 h | |
| Control a | 29.7 ± 2.3 | 28.6 ± 1.6 | 100 |
| AA(100 μM) | 27.9 ± 1.9 | 30.5 ± 2.1 | 112.1 ± 3.9 |
| DGLA(100 μM) | 41.3 b ± 2.5 | 31.4 ± 2.0 | 109.1 ± 5.7 |
| DGLA with 5.0 μM of CP | 42.4 b ± 1.8 | 35.3 b ± 0.9 | 98.3 c ± 5.0 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xu, Y.; Gu, Y.; Qian, S.Y. An Advanced Electron Spin Resonance (ESR) Spin-Trapping and LC/(ESR)/MS Technique for the Study of Lipid Peroxidation. Int. J. Mol. Sci. 2012, 13, 14648-14666. https://doi.org/10.3390/ijms131114648
Xu Y, Gu Y, Qian SY. An Advanced Electron Spin Resonance (ESR) Spin-Trapping and LC/(ESR)/MS Technique for the Study of Lipid Peroxidation. International Journal of Molecular Sciences. 2012; 13(11):14648-14666. https://doi.org/10.3390/ijms131114648
Chicago/Turabian StyleXu, Yi, Yan Gu, and Steven Y. Qian. 2012. "An Advanced Electron Spin Resonance (ESR) Spin-Trapping and LC/(ESR)/MS Technique for the Study of Lipid Peroxidation" International Journal of Molecular Sciences 13, no. 11: 14648-14666. https://doi.org/10.3390/ijms131114648