Isoflurane Protects Against Human Endothelial Cell Apoptosis by Inducing Sphingosine Kinase-1 via ERK MAPK

Abstract
:1. Introduction
2. Results and Discussion
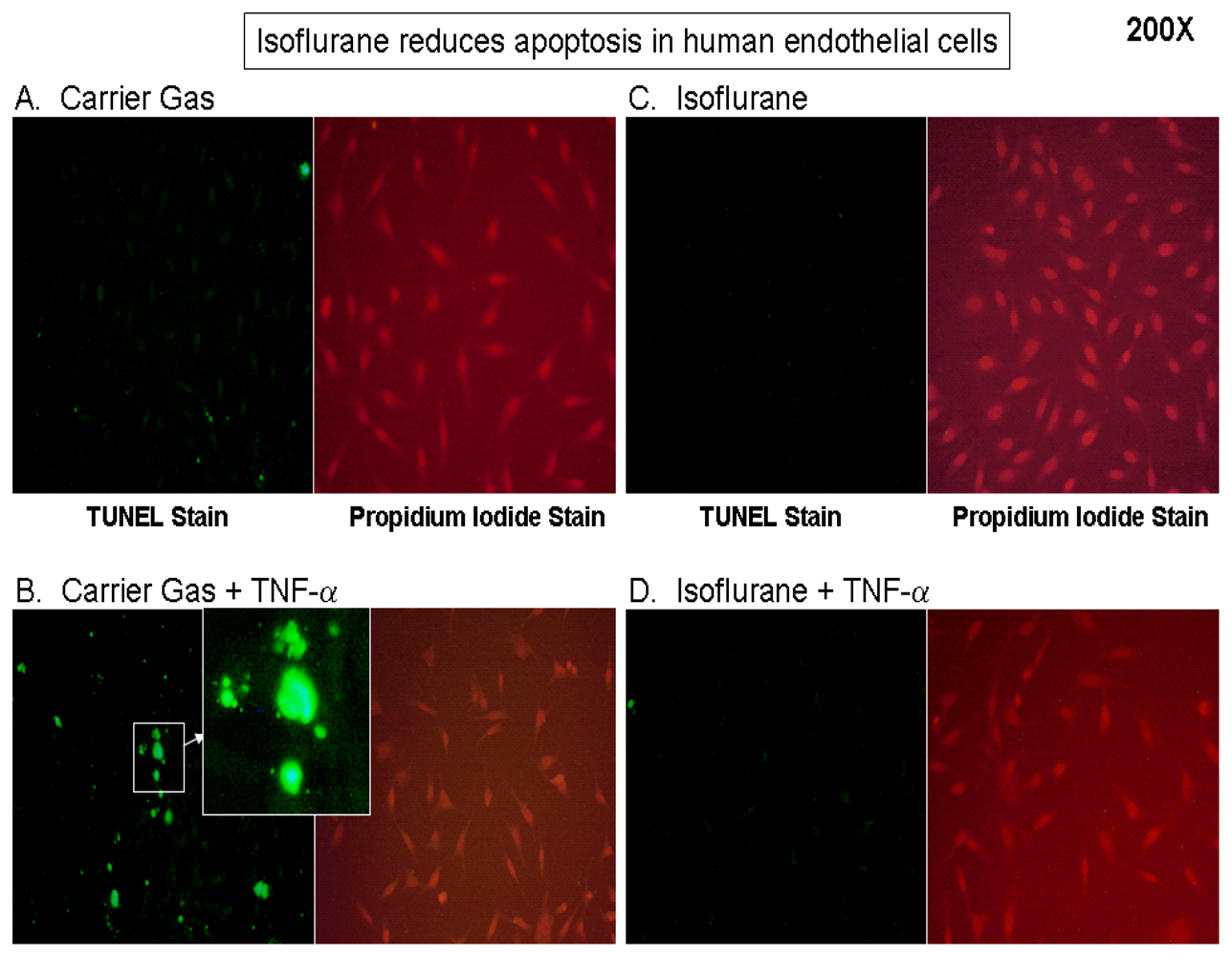
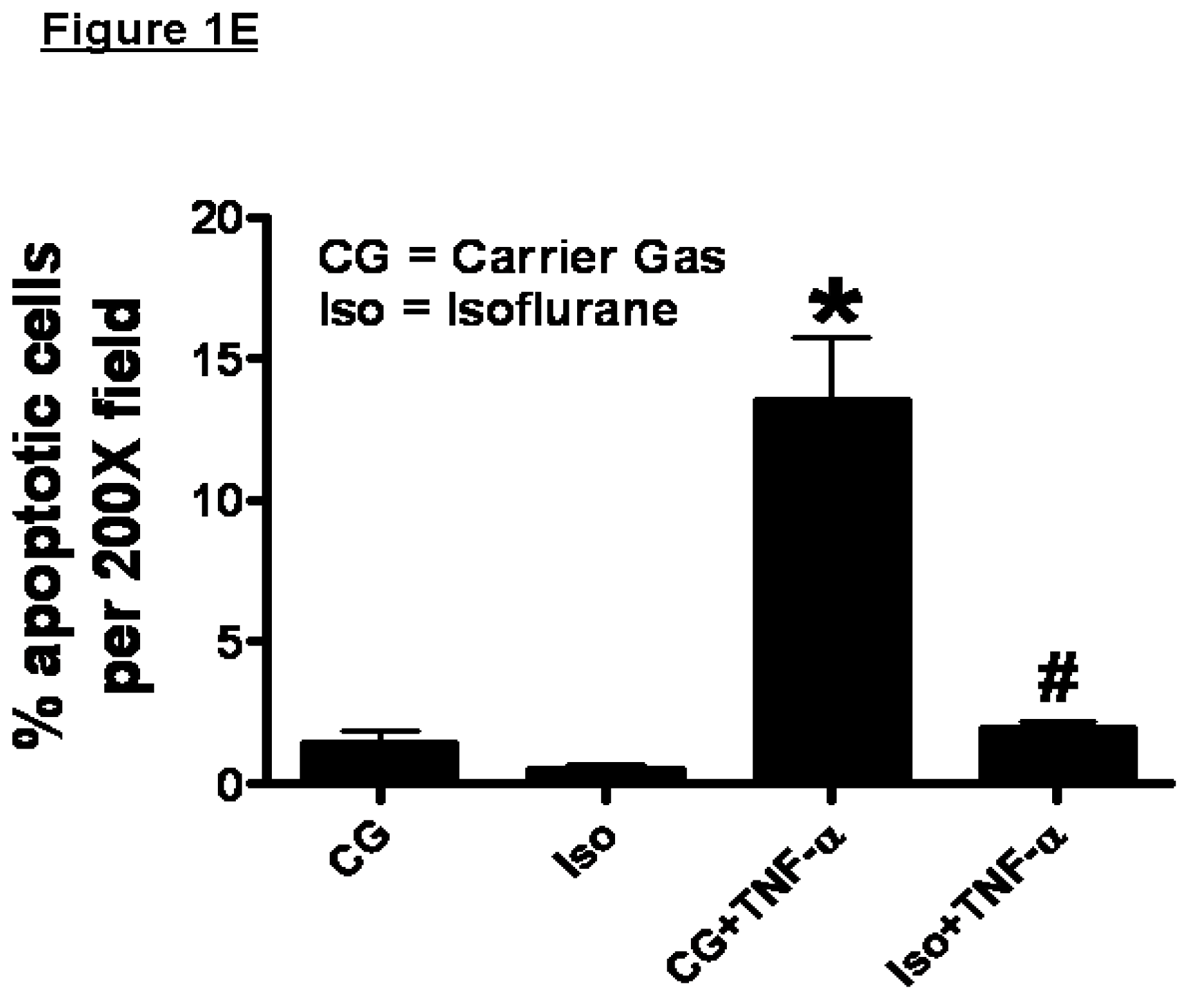
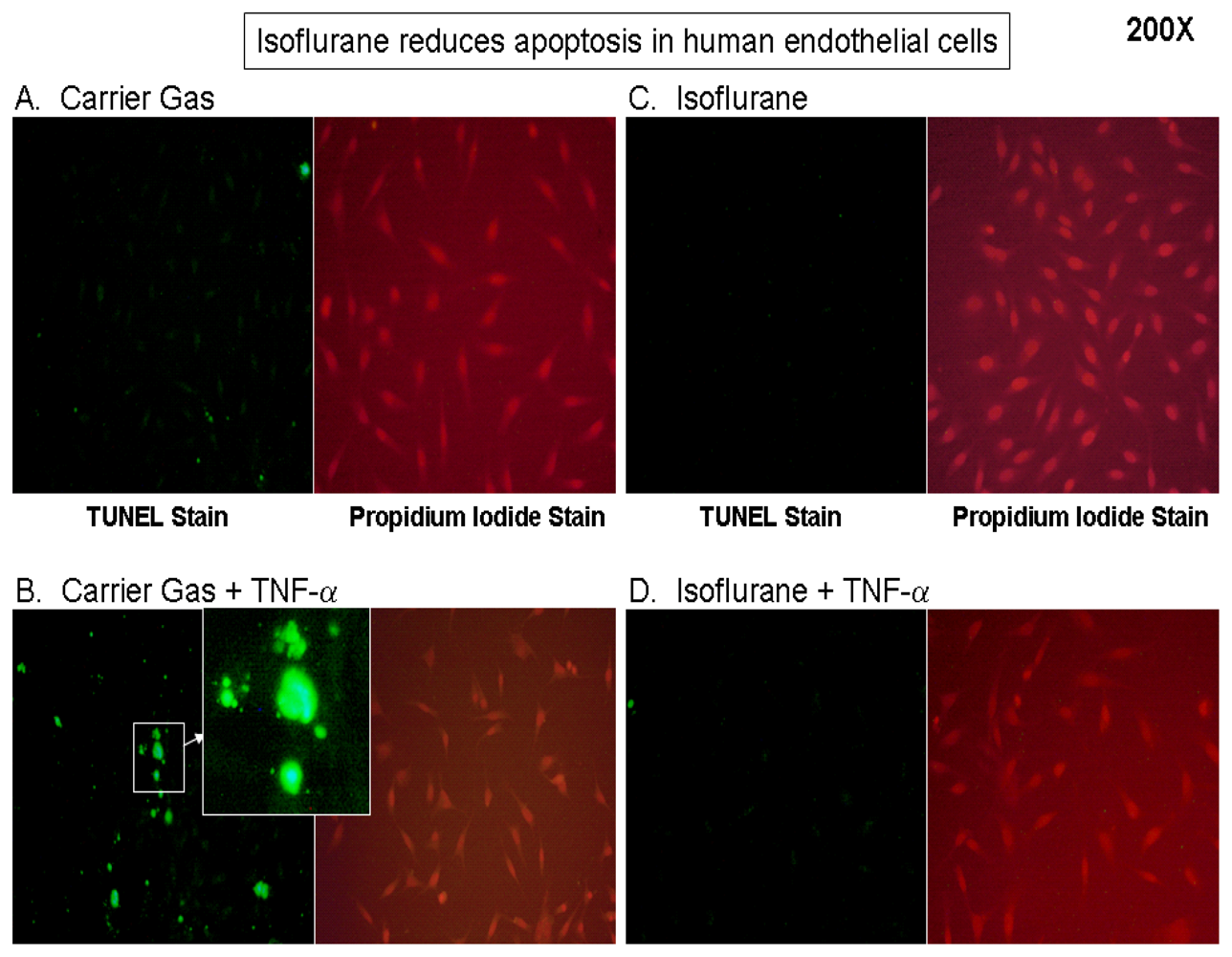
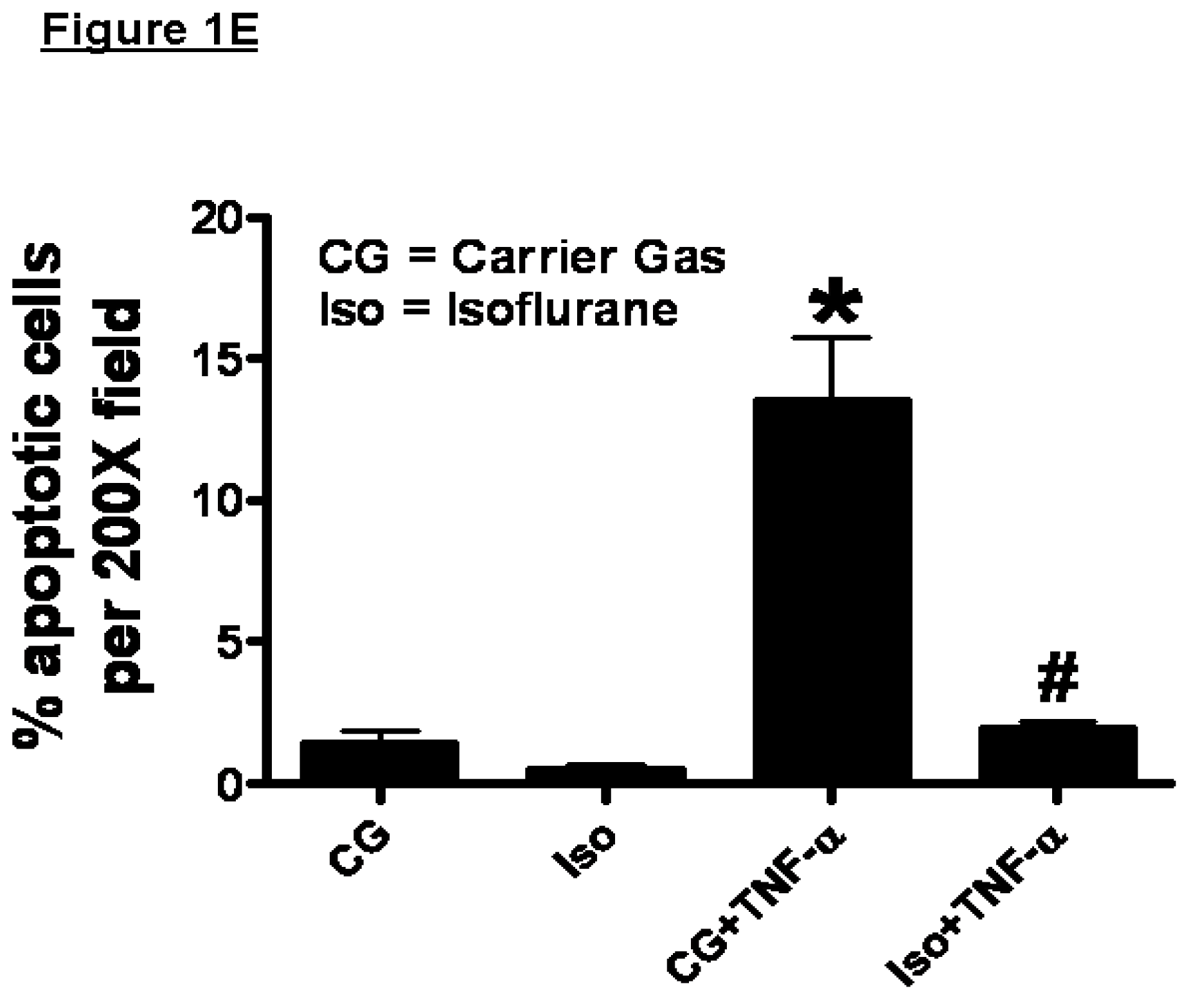
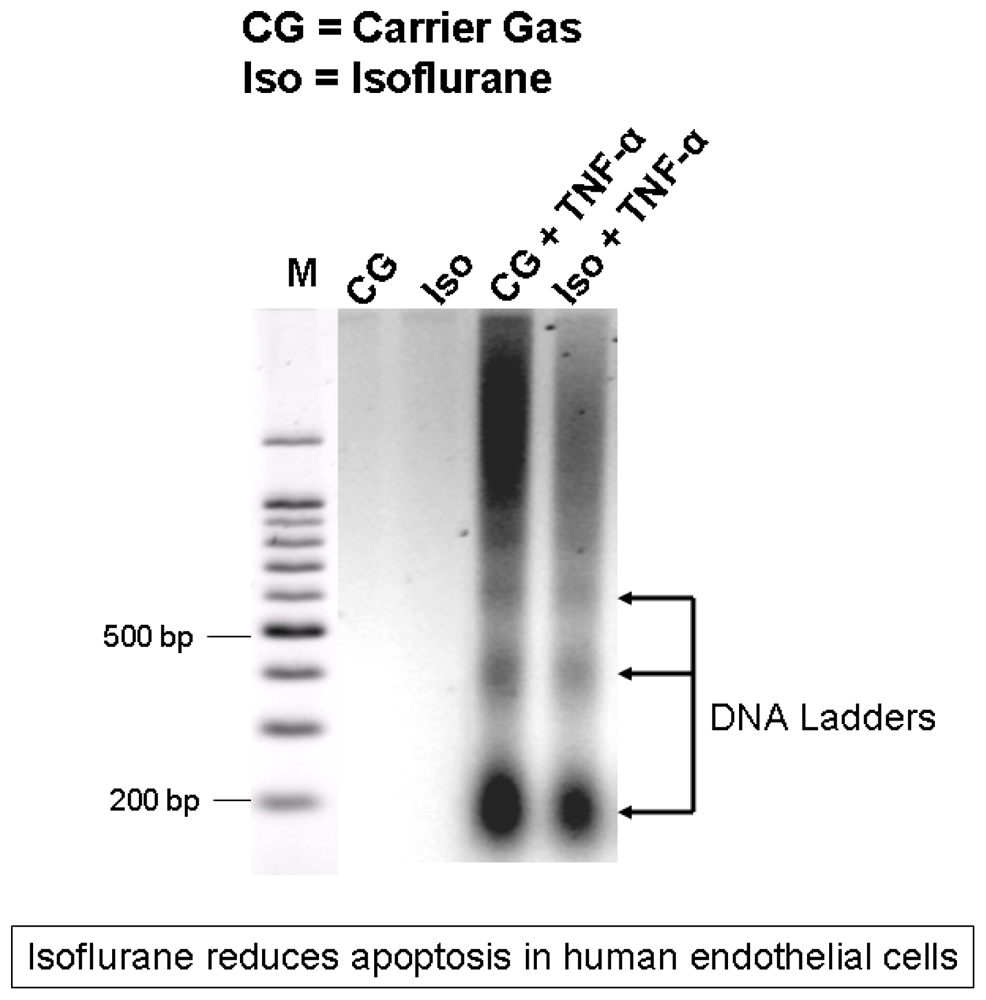
2.1. Isoflurane Pretreatment Reduces Apoptosis in EA.hy926 Cells Exposed to TNF-α
2.2. Isoflurane Does not Protect Against H2O2-Induced Necrosis in EA.hy926 Cells
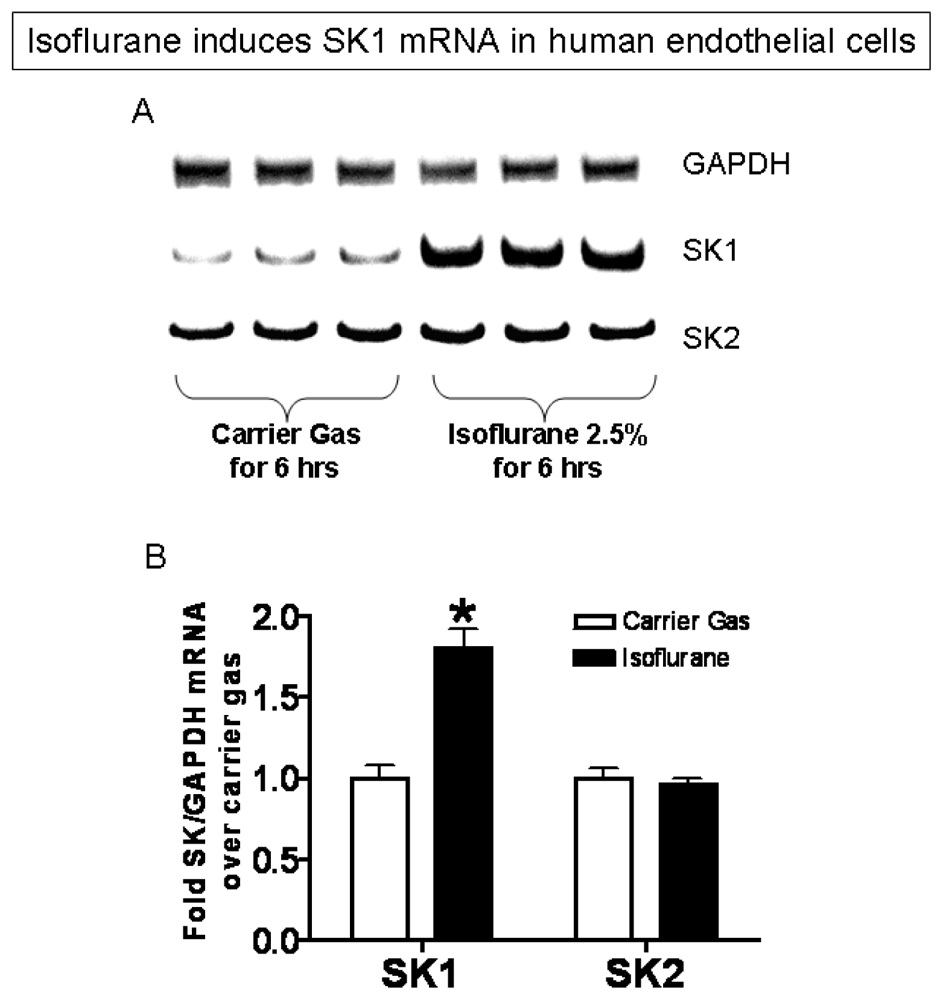
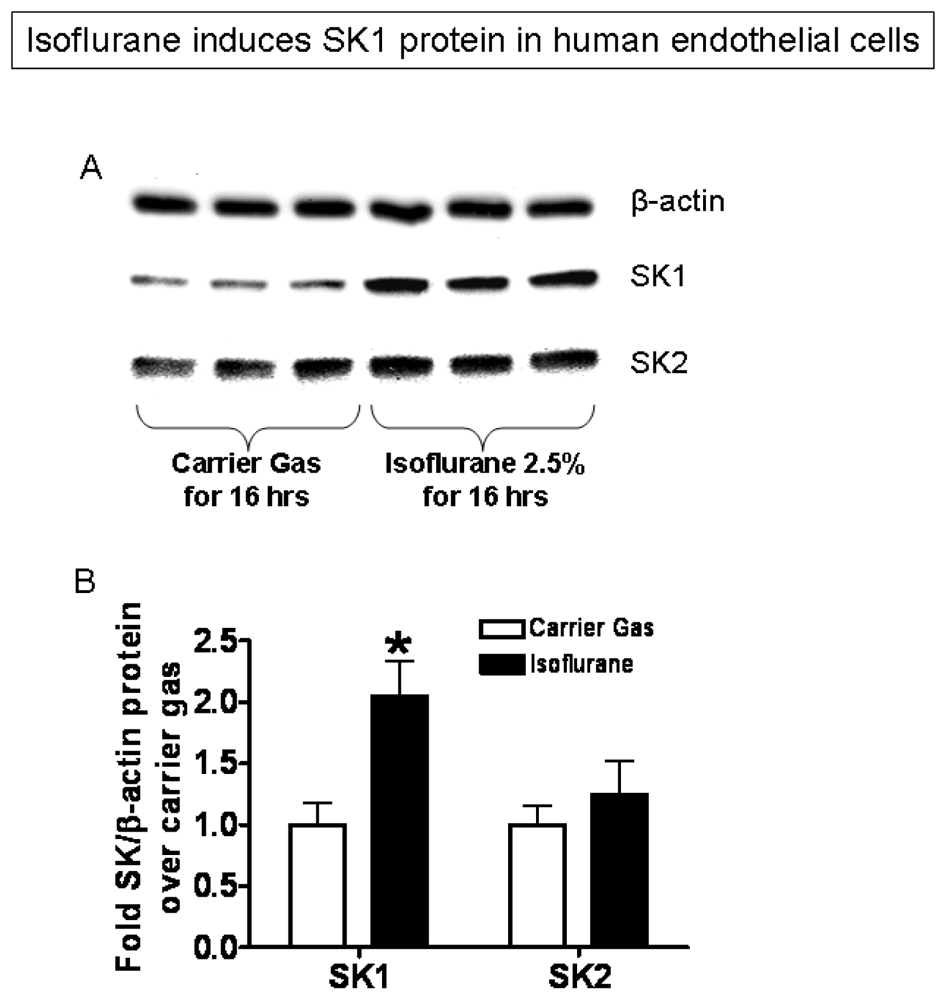
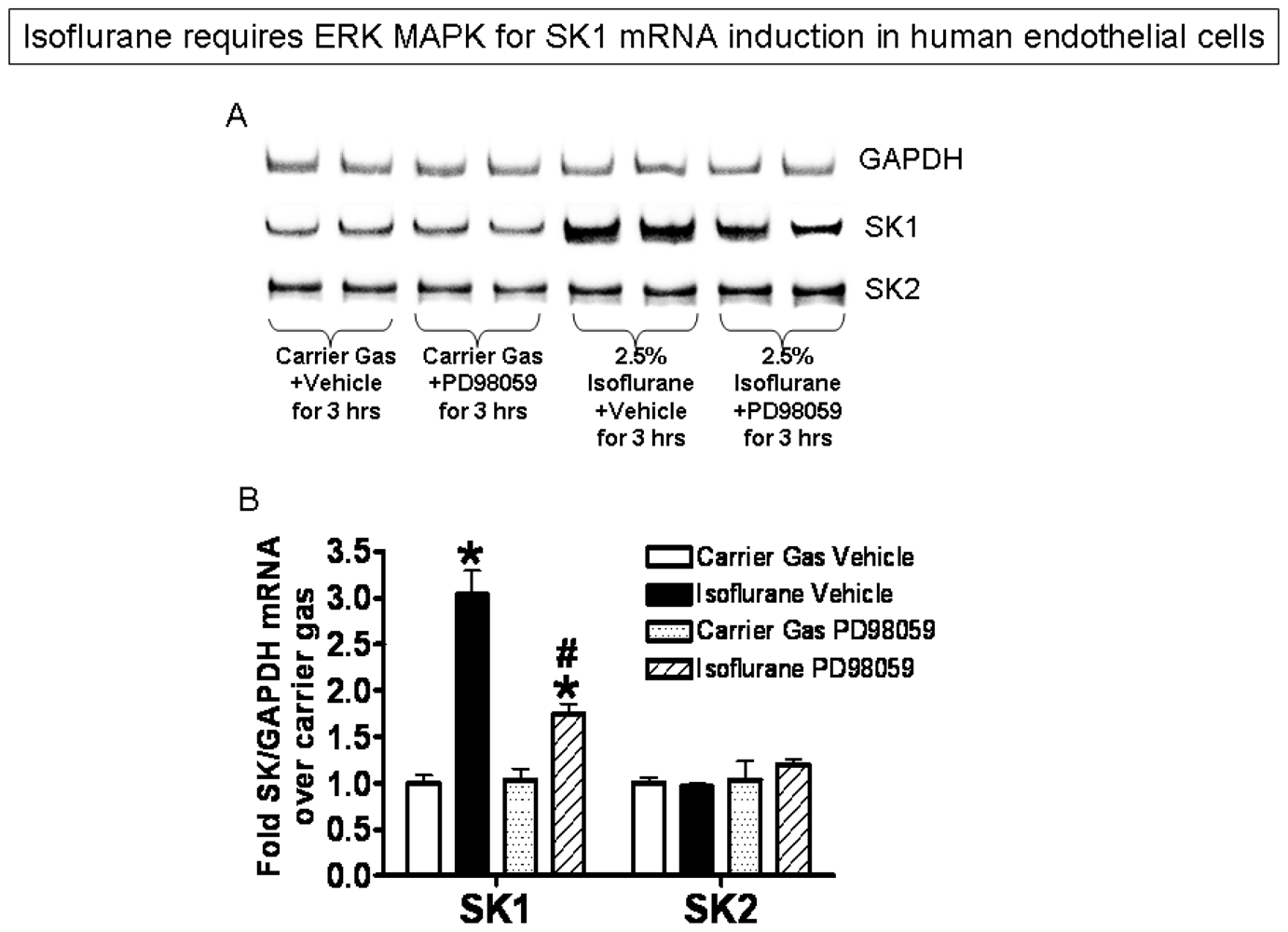
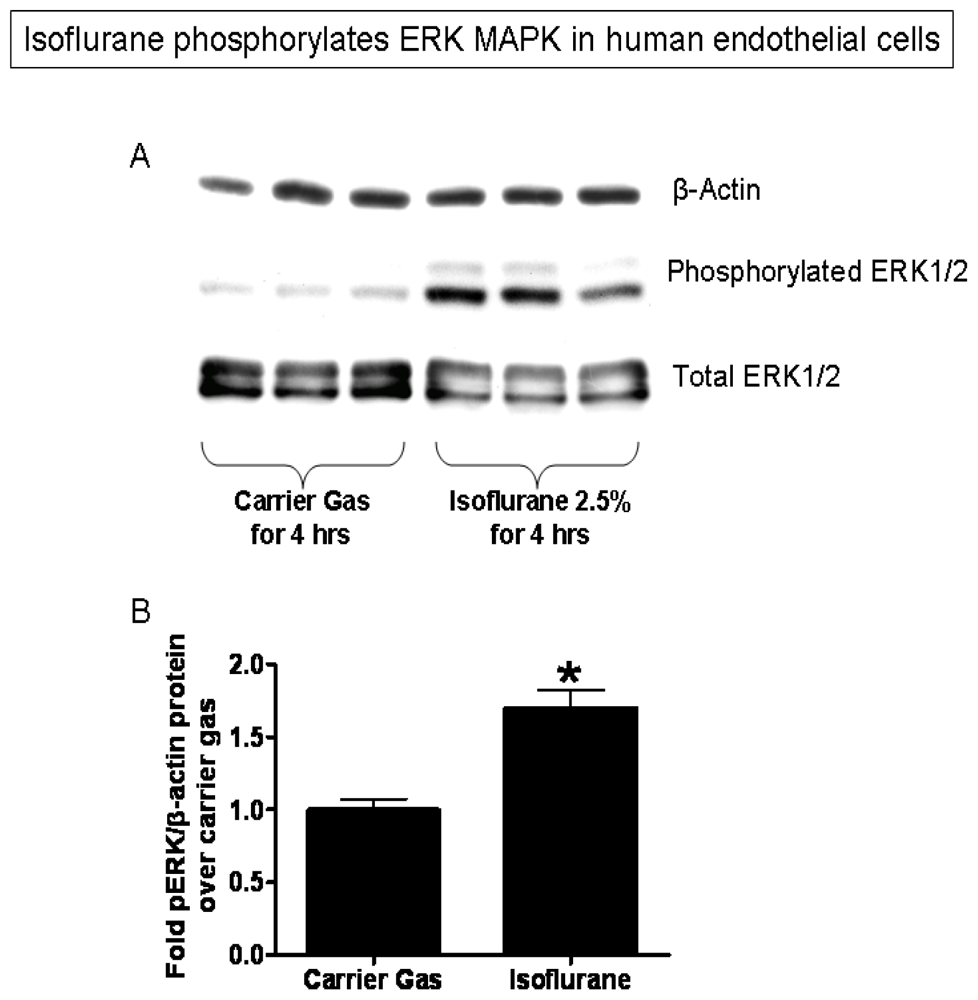
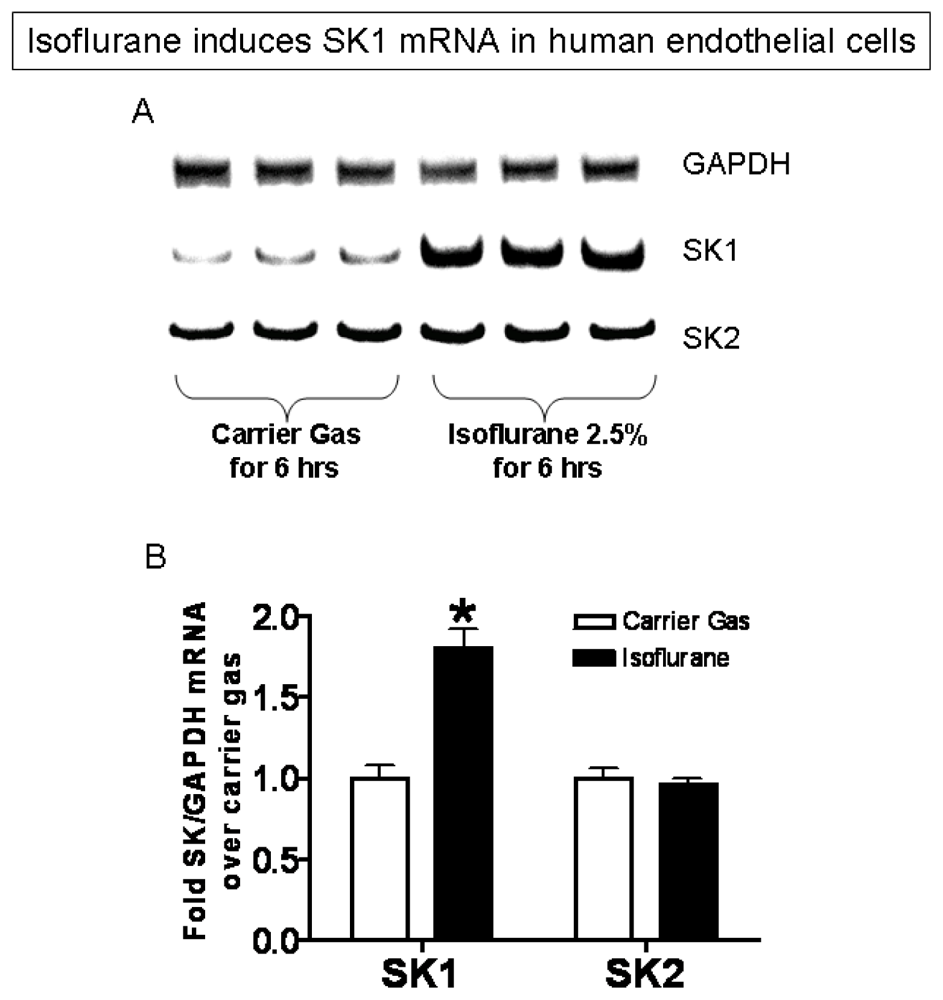
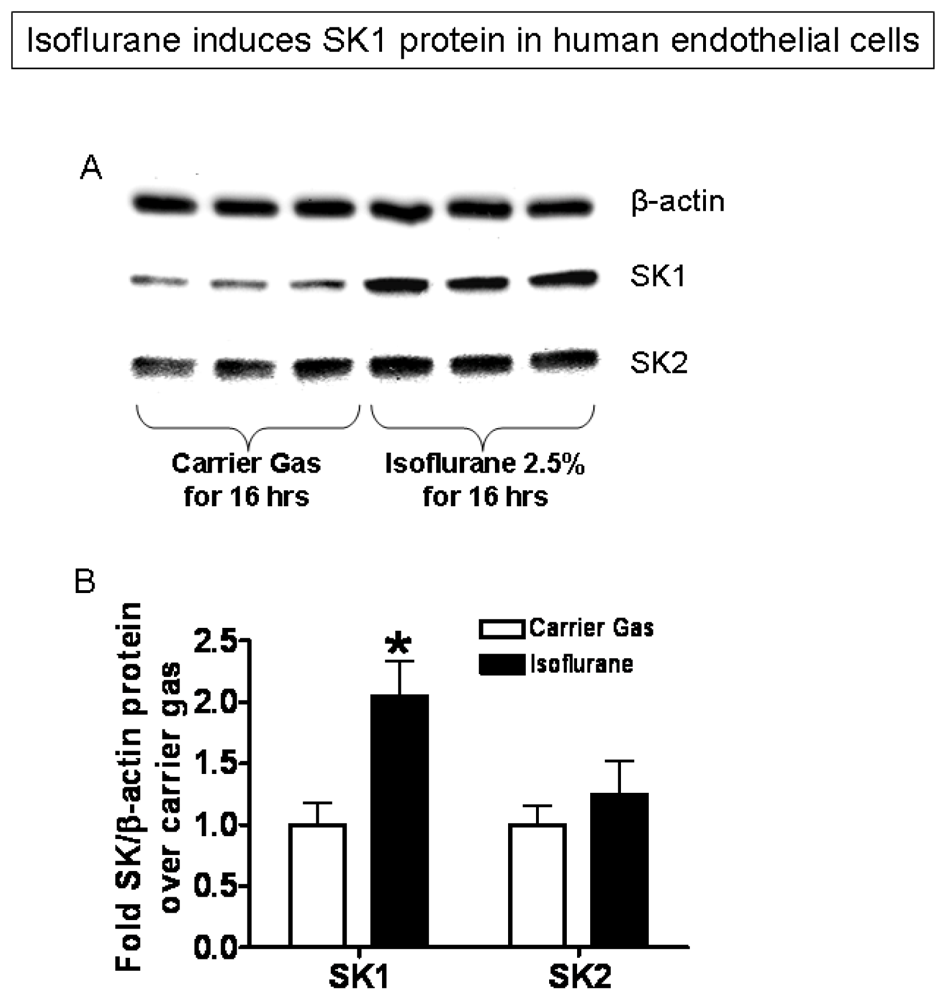
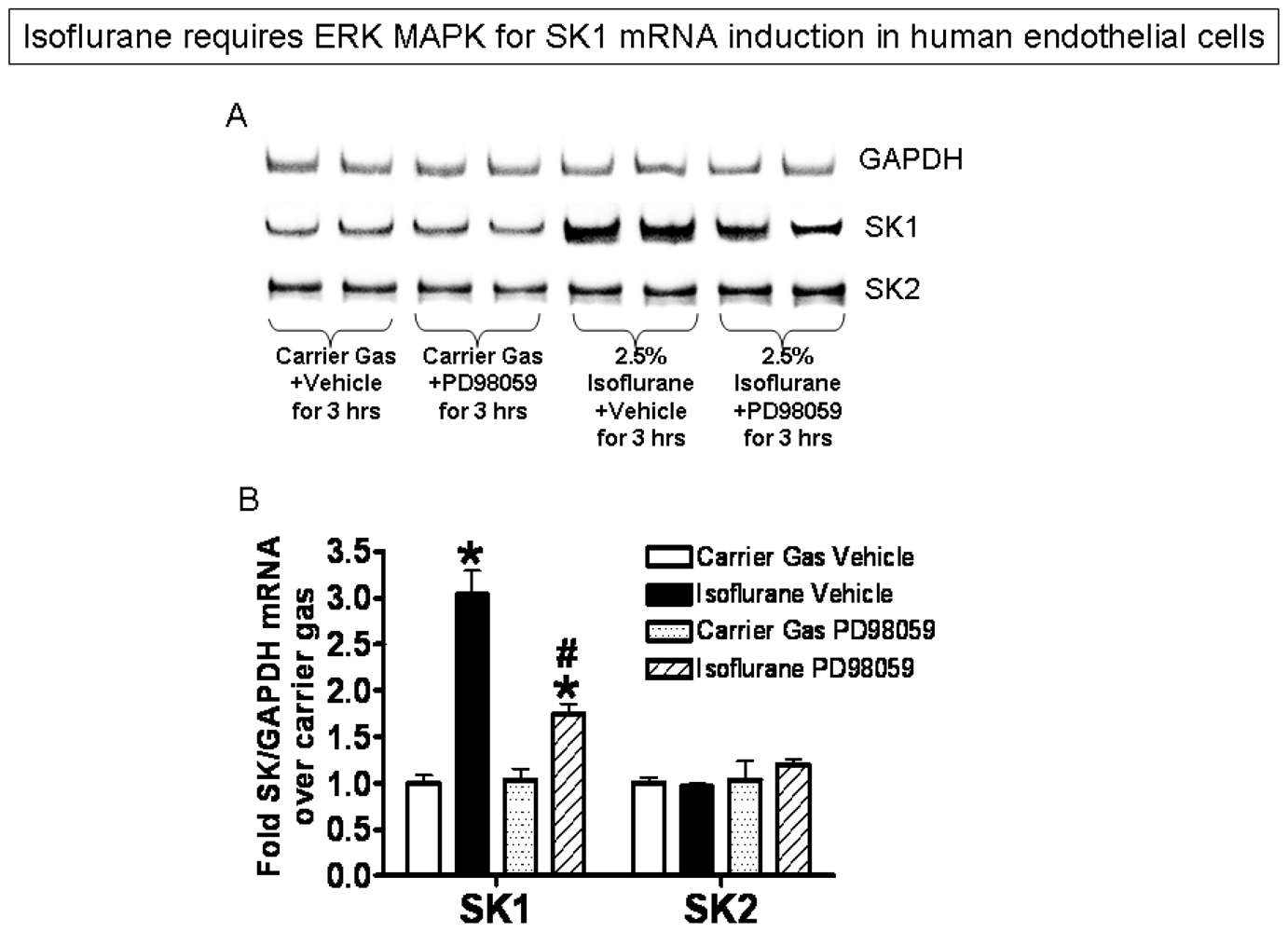
2.3. Isoflurane Treatment Phosphorylates ERK MAPK to Induce of SK1
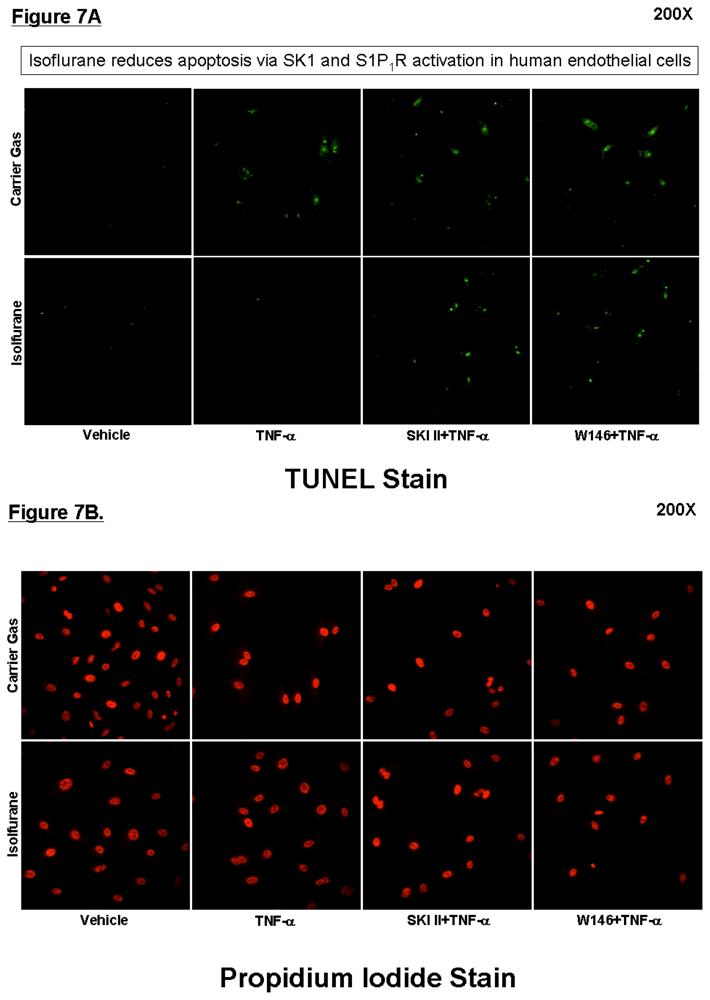
2.4. SK or S1P1 Receptor Inhibition Prevents Isoflurane-Mediated Reduction in Apoptosis
2.5. Isoflurane Increases SK Activity and S1P Formation in EA.hy926 Cells
2.6. Discussion
3. Experimental Section
3.1. Cell Culture
3.2. Exposure of EA.hy926 Cells to Isoflurane
3.3. Induction of EA.hy926 Cell Necrosis with Hydrogen Peroxide (H2O2)
3.4. Induction of EA.hy926 Cell Apoptosis Using Tumor Necrosis Factor-α (TNF-α)
3.5. Detection of Apoptosis
3.6. Detection of necrosis with LDH Measurement
3.7. Immunoblot Analysis
3.8. Semi-Quantitative Reverse Transcription–Polymerase Chain Reaction (RTPCR)
3.9. SK Activity Assay
3.10. High-Pressure Liquid Chromatography (HPLC) Detection of S1P
3.11. Statistical Analysis
3.12. Reagents and Protein Determination
4. Conclusion
Acknowledgements
- Conflict of Interest StatementWe declare that no financial conflict of interest exists for each author.
References
- Ait-Oufella, H.; Maury, E.; Lehoux, S.; Guidet, B.; Offenstadt, G. The endothelium: Physiological functions and role in microcirculatory failure during severe sepsis. Intensive Care Med 2010, 36, 1286–1298. [Google Scholar]
- Carden, D.L.; Granger, D.N. Pathophysiology of ischaemia-reperfusion injury. J. Pathol 2000, 190, 255–266. [Google Scholar]
- Stefanec, T. Endothelial apoptosis: Could it have a role in the pathogenesis and treatment of disease? Chest 2000, 117, 841–854. [Google Scholar]
- Preckel, B.; Bolten, J. Pharmacology of modern volatile anaesthetics. Best Pract. Res. Clin. Anaesthesiol 2005, 19, 331–348. [Google Scholar]
- Lee, H.T.; Kim, M.; Jan, M.; Emala, C.W. Anti-inflammatory and antinecrotic effects of the volatile anesthetic sevoflurane in kidney proximal tubule cells. Am. J. Physiol. Ren. Physiol 2006, 291, F67–F78. [Google Scholar]
- Lochhead, K.M.; Zager, R.A. Fluorinated anesthetic exposure “activates” the renal cortical sphingomyelinase cascade. Kidney Int 1998, 54, 373–381. [Google Scholar]
- Hait, N.C.; Oskeritzian, C.A.; Paugh, S.W.; Milstien, S.; Spiegel, S. Sphingosine kinases, sphingosine 1-phosphate, apoptosis and diseases. Biochim. Biophys. Acta 2006, 1758, 2016–2026. [Google Scholar]
- de Klaver, M.J.; Manning, L.; Palmer, L.A.; Rich, G.F. Isoflurane pretreatment inhibits cytokine-induced cell death in cultured rat smooth muscle cells and human endothelial cells. Anesthesiology 2002, 97, 24–32. [Google Scholar]
- Johnson, M.E.; Sill, J.C.; Uhl, C.B.; Halsey, T.J.; Gores, G.J. Effect of volatile anesthetics on hydrogen peroxide-induced injury in aortic and pulmonary arterial endothelial cells. Anesthesiology 1996, 84, 103–116. [Google Scholar]
- Zhou, Y.; Lekic, T.; Fathali, N.; Ostrowski, R.P.; Martin, R.D.; Tang, J.; Zhang, J.H. Isoflurane posttreatment reduces neonatal hypoxic-ischemic brain injury in rats by the sphingosine-1- phosphate/phosphatidylinositol-3-kinase/Akt pathway. Stroke 2010, 41, 1521–1527. [Google Scholar]
- Cason, B.A.; Gamperl, A.K.; Slocum, R.E.; Hickey, R.F. Anesthetic-induced preconditioning: Previous administration of isoflurane decreases myocardial infarct size in rabbits. Anesthesiology 1997, 87, 1182–1190. [Google Scholar]
- Kim, M.; Kim, M.; Kim, N.; D’Agati, V.D.; Emala, C.W.; Lee, H.T. Isoflurane mediates protection from renal ischemia-reperfusion injury via sphingosine kinase and sphingosine-1- phosphate-dependent pathways. Am. J. Physiol. Ren. Physiol 2007, 293, F1827–F1835. [Google Scholar]
- Kim, M.; Kim, M.; Park, S.W.; Pitson, S.M.; Lee, H.T. Isoflurane protects human kidney proximal tubule cells against necrosis via sphingosine kinase and sphingosine-1-phosphate generation. Am. J. Nephrol 2010, 31, 353–362. [Google Scholar]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol 2007, 35, 495–516. [Google Scholar]
- Weber, N.C.; Kandler, J.; Schlack, W.; Grueber, Y.; Fradorf, J.; Preckel, B. Intermitted pharmacologic pretreatment by xenon, isoflurane, nitrous oxide, and the opioid morphine prevents tumor necrosis factor alpha induced adhesion molecule expression in human umbilical vein endothelial cells. Anesthesiology 2008, 108, 199–207. [Google Scholar]
- de Klaver, M.J.; Buckingham, M.G.; Rich, G.F. Isoflurane pretreatment has immediate and delayed protective effects against cytokine-induced injury in endothelial and vascular smooth muscle cells. Anesthesiology 2003, 99, 896–903. [Google Scholar]
- Lee, H.T.; Emala, C.W. Adenosine attenuates oxidant injury in human kidney proximal tubular cells via A1 and A2a adenosine receptor activation. Am. J. Physiol. Ren. Physiol 2002, 282, F844–F852. [Google Scholar]
- Kim, M.; Park, S.W.; Kim, M.; D’Agati, V.D.; Lee, H.T. Isoflurane protects against intestinal ischemia-reperfusion injury and multi-organ dysfunction via transforming growth factor-beta1. Ann. Surg 2011, in press. [Google Scholar]
- Venkataraman, K.; Lee, Y.M.; Michaud, J.; Thangada, S.; Ai, Y.; Bonkovsky, H.L.; Parikh, N.S.; Habrukowich, C.; Hla, T. Vascular endothelium as a contributor of plasma sphingosine 1-phosphate. Circ. Res 2008, 102, 669–676. [Google Scholar]
- Jo, S.K.; Bajwa, A.; Awad, A.S.; Lynch, K.R.; Okusa, M.D. Sphingosine-1-phosphate receptors: Biology and therapeutic potential in kidney disease. Kidney Int 2008, 73, 1220–1230. [Google Scholar]
- Liu, H.; Chakravarty, D.; Maceyka, M.; Milstien, S.; Spiegel, S. Sphingosine kinases: A novel family of lipid kinases. Prog. Nucleic Acid Res. Mol. Biol 2002, 71, 493–511. [Google Scholar]
- Olivera, A.; Kohama, T.; Edsall, L.; Nava, V.; Cuvillier, O.; Poulton, S.; Spiegel, S. Sphingosine kinase expression increases intracellular sphingosine-1-phosphate and promotes cell growth and survival. J. Cell Biol 1999, 147, 545–558. [Google Scholar]
- Moriue, T.; Igarashi, J.; Yoneda, K.; Nakai, K.; Kosaka, H.; Kubota, Y. Sphingosine 1-phosphate attenuates H2O2-induced apoptosis in endothelial cells. Biochem. Biophys. Res. Commun 2008, 368, 852–857. [Google Scholar]
- Kimura, T.; Sato, K.; Kuwabara, A.; Tomura, H.; Ishiwara, M.; Kobayashi, I.; Ui, M.; Okajima, F. Sphingosine 1-phosphate may be a major component of plasma lipoproteins responsible for the cytoprotective actions in human umbilical vein endothelial cells. J. Biol. Chem 2001, 276, 31780–31785. [Google Scholar]
- Vessey, D.A.; Kelley, M.; Li, L.; Huang, Y.; Zhou, H.Z.; Zhu, B.Q.; Karliner, J.S. Role of sphingosine kinase activity in protection of heart against ischemia reperfusion injury. Med. Sci. Monit 2006, 12, BR318–324. [Google Scholar]
- Kon, S.; Imai, M.; Inaba, H. Isoflurane attenuates early neutrophil independent hypoxia-reoxygenation injuries in the reperfused liver in fasted rats. Anesthesiology 1997, 86, 128–136. [Google Scholar]
- Awad, A.S.; Ye, H.; Huang, L.; Li, L.; Foss, F.W.; Macdonald, T.L.; Lynch, K.R.; Okusa, M.D. Selective sphingosine 1-phosphate 1 receptor activation reduces ischemia-reperfusion injury in mouse kidney. Am. J. Physiol. Ren. Physiol 2006, 290, F1516–F1524. [Google Scholar]
- Kolch, W. Meaningful relationships: The regulation of the Ras/Raf/MEK/ERK pathway by protein interactions. Biochem. J 2000, 351, 289–305. [Google Scholar]
- Wu, J.; Spiegel, S.; Sturgill, T.W. Sphingosine 1-phosphate rapidly activates the mitogen-activated protein kinase pathway by a G protein-dependent mechanism. J. Biol. Chem 1995, 270, 11484–11488. [Google Scholar]
- Kersten, J.R.; Schmeling, T.J.; Pagel, P.S.; Gross, G.J.; Warltier, D.C. Isoflurane mimics ischemic preconditioning via activation of K(ATP) channels: Reduction of myocardial infarct size with an acute memory phase. Anesthesiology 1997, 87, 361–370. [Google Scholar]
- Shayevitz, J.R.; Rodriguez, J.L.; Gilligan, L.; Johnson, K.J.; Tait, A.R. Volatile anesthetic modulation of lung injury and outcome in a murine model of multiple organ dysfunction syndrome. Shock 1995, 4, 61–67. [Google Scholar]
- Stratmann, G.; Sall, J.W.; May, L.D.; Bell, J.S.; Magnusson, K.R.; Rau, V.; Visrodia, K.H.; Alvi, R.S.; Ku, B.; Lee, M.T.; et al. Isoflurane differentially affects neurogenesis and long-term neurocognitive function in 60-day-old and 7-day-old rats. Anesthesiology 2009, 110, 834–848. [Google Scholar]
- Head, B.P.; Patel, H.H.; Niesman, I.R.; Drummond, J.C.; Roth, D.M.; Patel, P.M. Inhibition of p75 neurotrophin receptor attenuates isoflurane-mediated neuronal apoptosis in the neonatal central nervous system. Anesthesiology 2009, 110, 813–825. [Google Scholar]
- Lemkuil, B.P.; Head, B.P.; Pearn, M.L.; Patel, H.H.; Drummond, J.C.; Patel, P.M. Isoflurane neurotoxicity is mediated by p75NTR-RhoA activation and actin depolymerization. Anesthesiology 2011, 114, 49–57. [Google Scholar]
- Jevtovic-Todorovic, V.; Hartman, R.E.; Izumi, Y.; Benshoff, N.D.; Dikranian, K.; Zorumski, C.F.; Olney, J.F.; Wozniak, D.F. Early exposure to common anesthetic agents causes widespread neurodegeneration in the developing rat brain and persistent learning deficits. J. Neurosci 2003, 23, 876–882. [Google Scholar]
- Levin, E.D.; Uemura, E.; Bowman, R.E. Neurobehavioral toxicology of halothane in rats. Neurotoxicol. Teratol 1991, 13, 461–470. [Google Scholar]
- Chi, O.Z.; Hunter, C.; Liu, X.; Weiss, H.R. The effects of isoflurane pretreatment on cerebral blood flow, capillary permeability, and oxygen consumption in focal cerebral ischemia in rats. Anesth. Analg 2010, 110, 1412–1418. [Google Scholar]
- Li, L.; Zuo, Z. Isoflurane preconditioning improves short-term and long-term neurological outcome after focal brain ischemia in adult rats. Neuroscience 2009, 164, 497–506. [Google Scholar]
- Statler, K.D.; Alexander, H.; Vagni, V.; Holubkov, R.; Dixon, C.E.; Clark, R.S.; Jenkins, L.; Kochanek, P.M. Isoflurane exerts neuroprotective actions at or near the time of severe traumatic brain injury. Brain Res 2006, 1076, 216–224. [Google Scholar]
- Statler, K.D.; Alexander, H.; Vagni, V.; Dixon, C.E.; Clark, R.S.; Jenkins, L.; Kochanek, P.M. Comparison of seven anesthetic agents on outcome after experimental traumatic brain injury in adult, male rats. J. Neurotrauma 2006, 23, 97–108. [Google Scholar]
- Edgell, C.J.; McDonald, C.C.; Graham, J.B. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc. Natl. Acad. Sci. USA 1983, 80, 3734–3737. [Google Scholar]
- Herrmann, M.; Lorenz, H.M.; Voll, R.; Grunke, M.; Woith, W.; Kalden, J.R. A rapid and simple method for the isolation of apoptotic DNA fragments. Nucleic Acids Res 1994, 22, 5506–5507. [Google Scholar]
- Lee, H.T.; Ota-Setlik, A.; Fu, Y.; Nasr, S.H.; Emala, C.W. Differential protective effects of volatile anesthetics against renal ischemia-reperfusion injury in vivo. Anesthesiology 2004, 101, 1313–1324. [Google Scholar]
- Vessey, D.A.; Kelley, M.; Karliner, J.S. A rapid radioassay for sphingosine kinase. Anal. Biochem 2005, 337, 136–142. [Google Scholar]
- Min, J.K.; Yoo, H.S.; Lee, E.Y.; Lee, W.J.; Lee, Y.M. Simultaneous quantitative analysis of sphingoid base 1-phosphates in biological samples by ophthalaldehyde precolumn derivatization after dephosphorylation with alkaline phosphatase. Anal. Biochem 2002, 303, 167–175. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Accession Number | Sequence (Sense/Antisense | Product Size (bp) | Cycle Number | Annealing Temp. (°C) |
|---|---|---|---|---|---|
| Human SK1 | NM_021972 | 5′-ATCTCCTTCACGCTGATGC-3′ 5′-GTGCAGAGACAGCAGGTTCA-3′ | 330 | 26 | 66 |
| Human SK2 | NM_020126 | 5′-GGAGGAAGCTGTGAAGATGC-3′ 5′-GCAGGTCAGACACAGAACGA-3′ | 482 | 22 | 66 |
| Human GAPDH | NM_002046 | 5′-ACCACAGTCCATGCCATCAC-3′ 5′-CACCACCCTGTTGCTGTAGCC-3′ | 450 | 15 | 65 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bakar, A.M.; Park, S.W.; Kim, M.; Lee, H.T. Isoflurane Protects Against Human Endothelial Cell Apoptosis by Inducing Sphingosine Kinase-1 via ERK MAPK. Int. J. Mol. Sci. 2012, 13, 977-993. https://doi.org/10.3390/ijms13010977
Bakar AM, Park SW, Kim M, Lee HT. Isoflurane Protects Against Human Endothelial Cell Apoptosis by Inducing Sphingosine Kinase-1 via ERK MAPK. International Journal of Molecular Sciences. 2012; 13(1):977-993. https://doi.org/10.3390/ijms13010977
Chicago/Turabian StyleBakar, Adnan M., Sang Won Park, Mihwa Kim, and H. Thomas Lee. 2012. "Isoflurane Protects Against Human Endothelial Cell Apoptosis by Inducing Sphingosine Kinase-1 via ERK MAPK" International Journal of Molecular Sciences 13, no. 1: 977-993. https://doi.org/10.3390/ijms13010977