Effects of New Dietary Fiber from Japanese Apricot (Prunus mume Sieb. et Zucc.) on Gut Function and Intestinal Microflora in Adult Mice

Abstract
:1. Introduction
2. Results and Discussion
2.1. General Observations
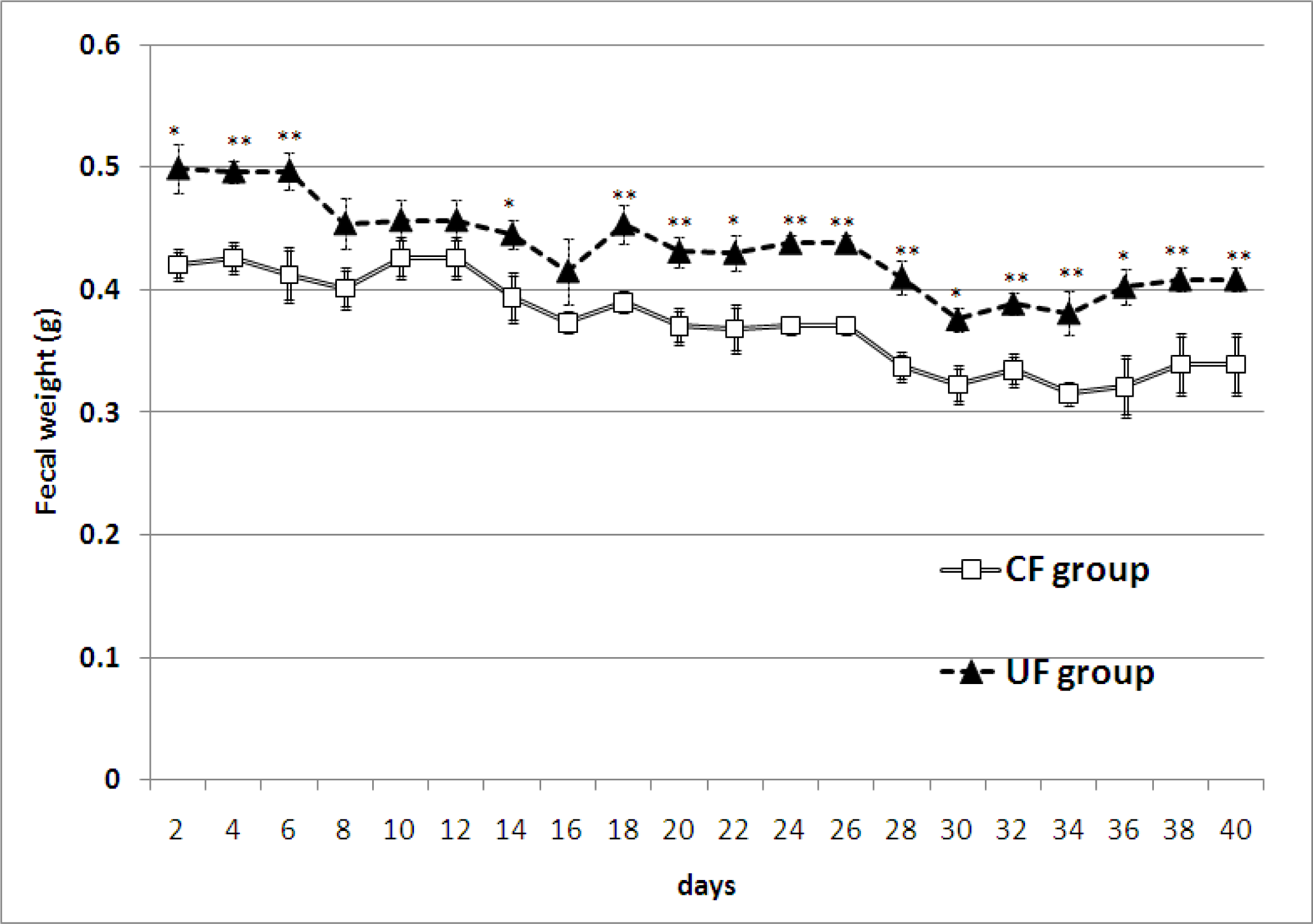
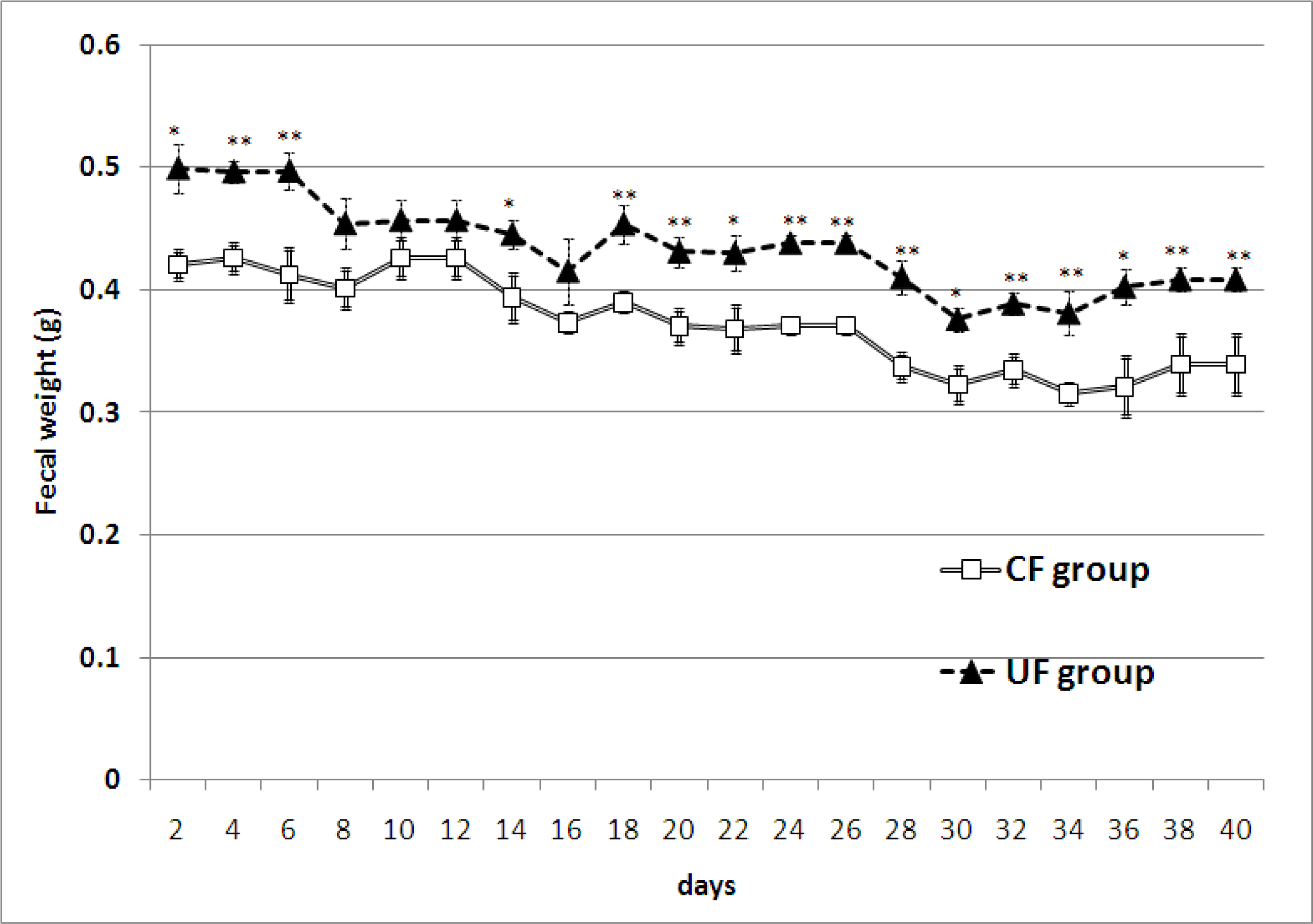
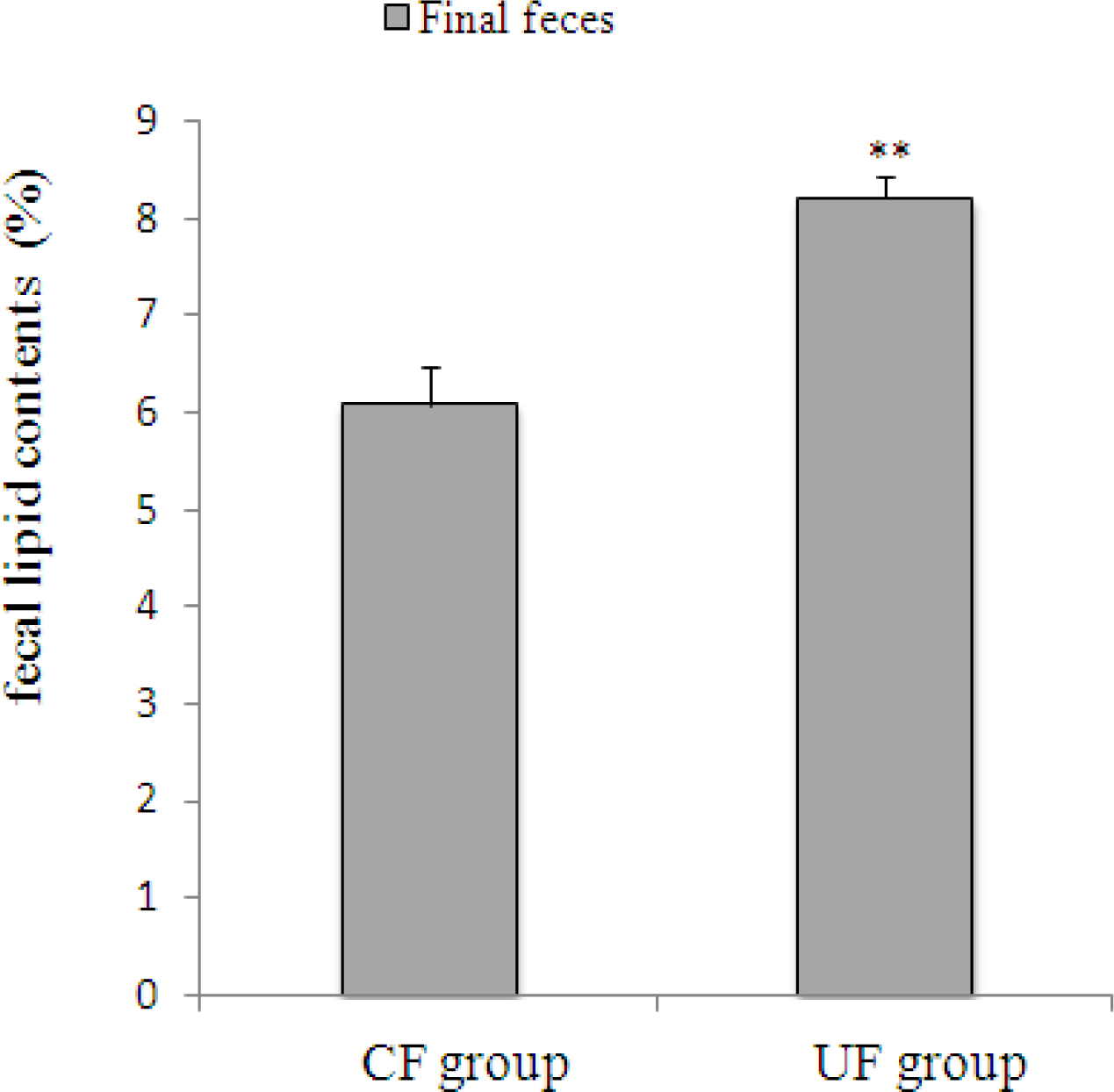
2.2. Amount of Feces and Fecal Lipid Contents
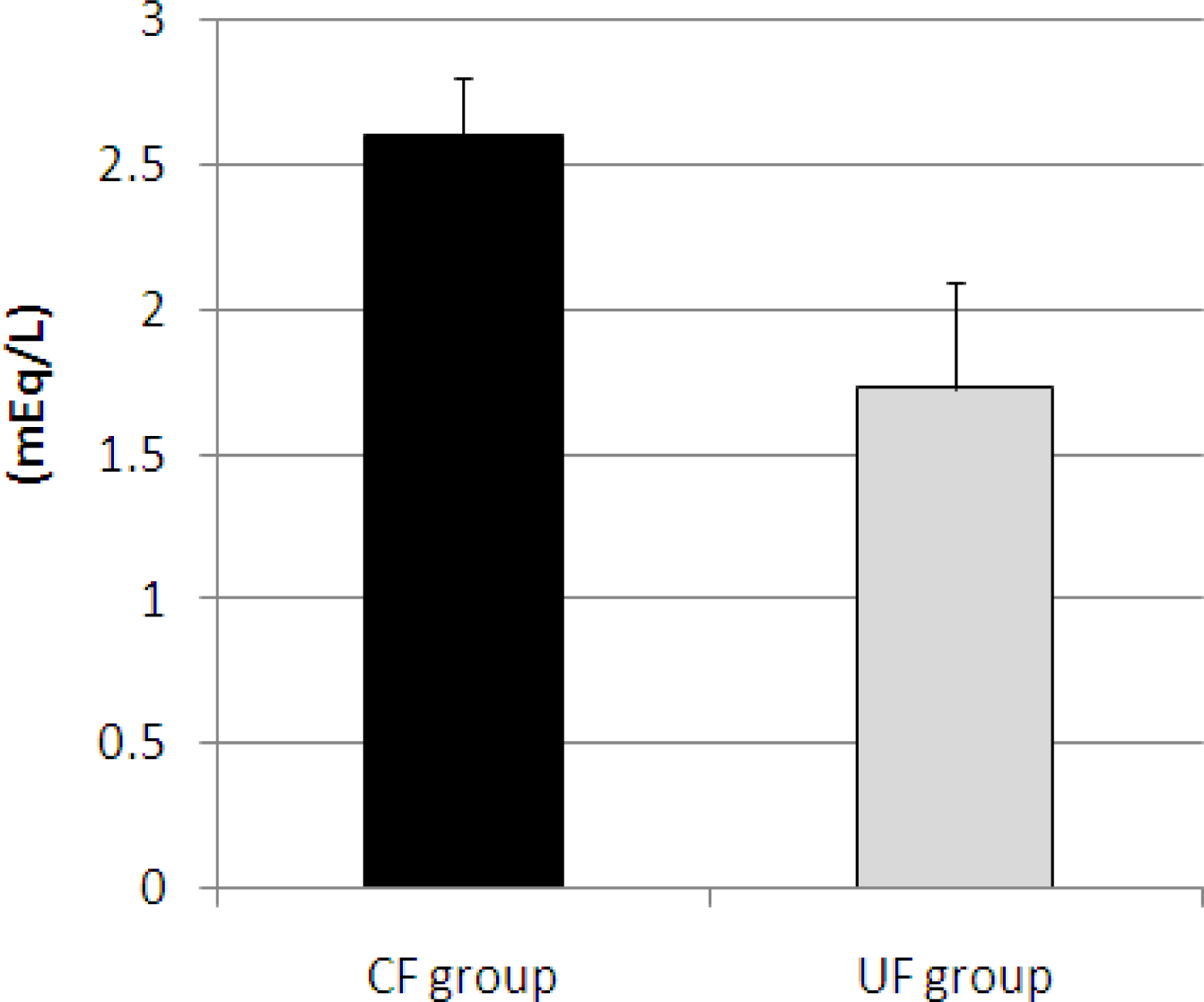
2.3. Plasma Total Cholesterol, Triglyceride, Phospholipids, NEFA and Plasma Glucose
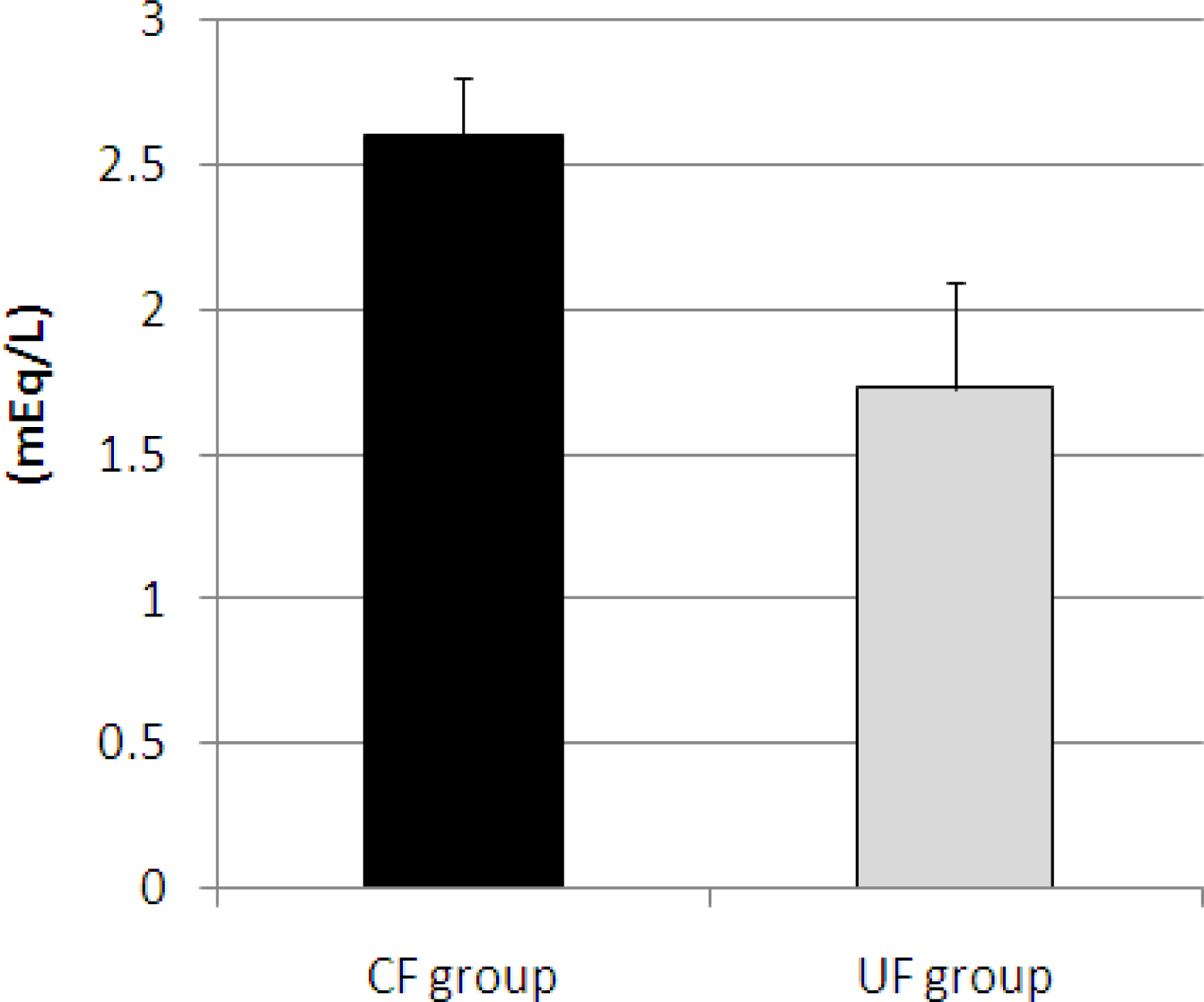
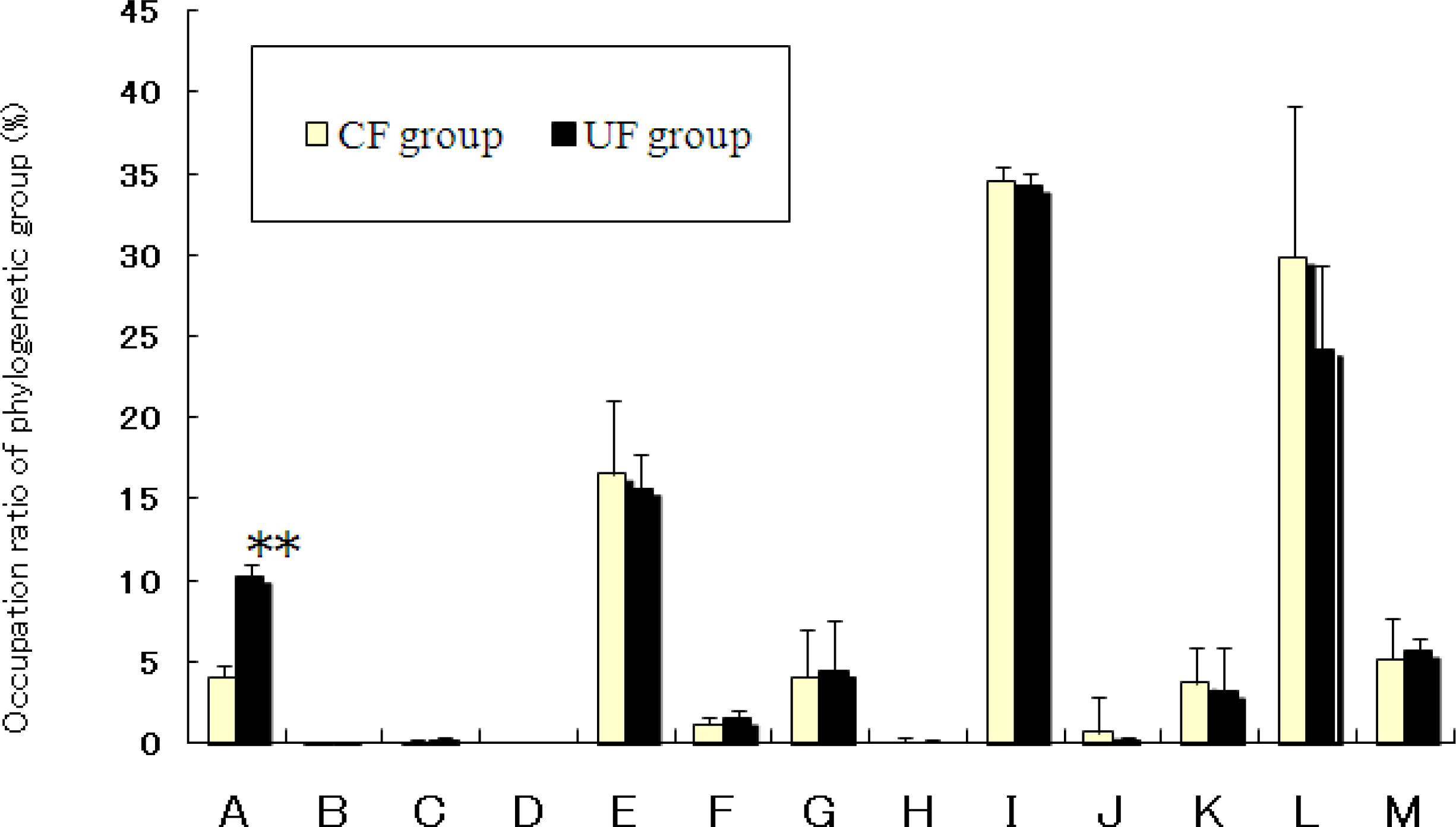
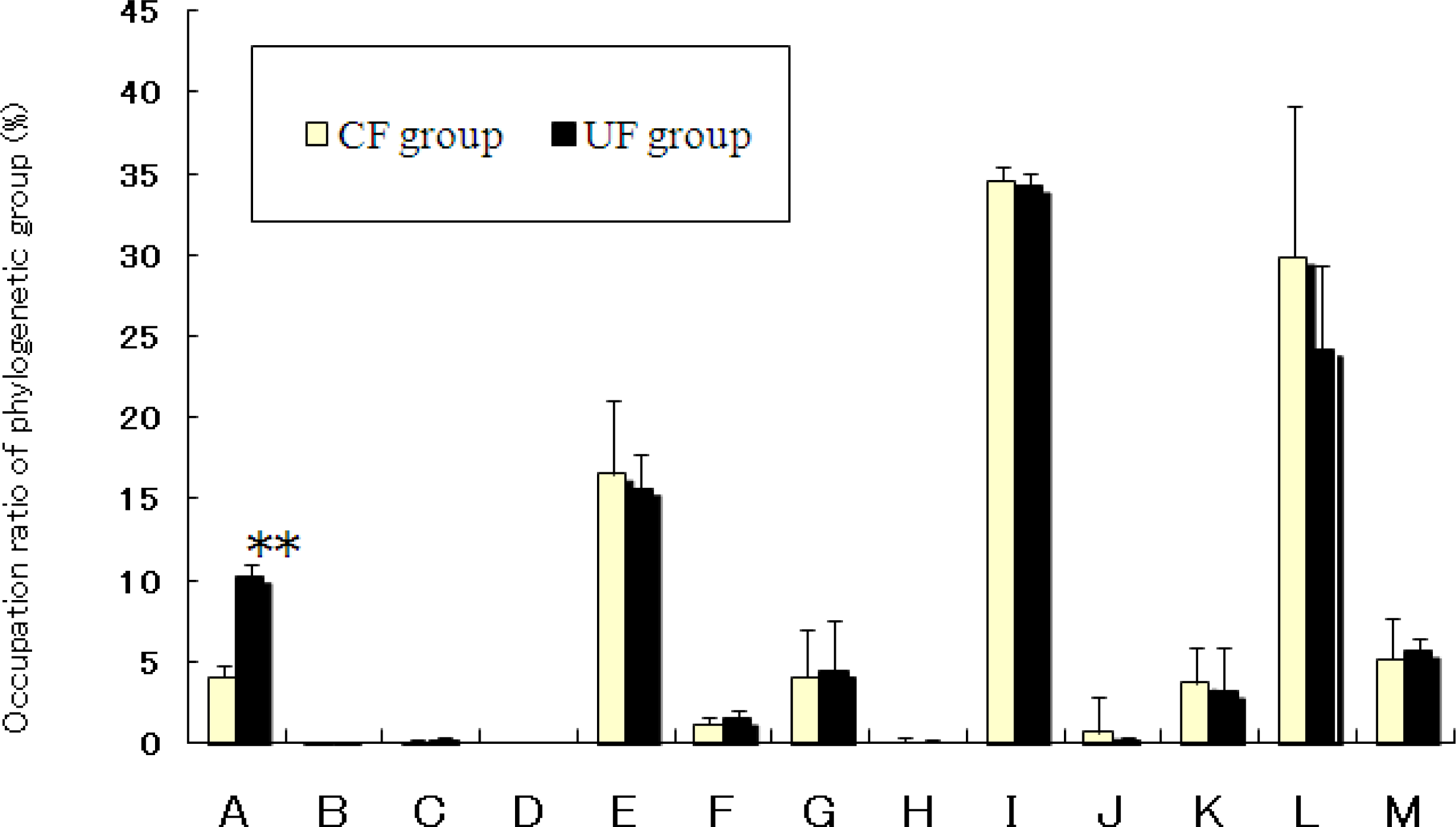
2.4. Effects of Diet on Cecal Flora of Mice
3. Experimental Section
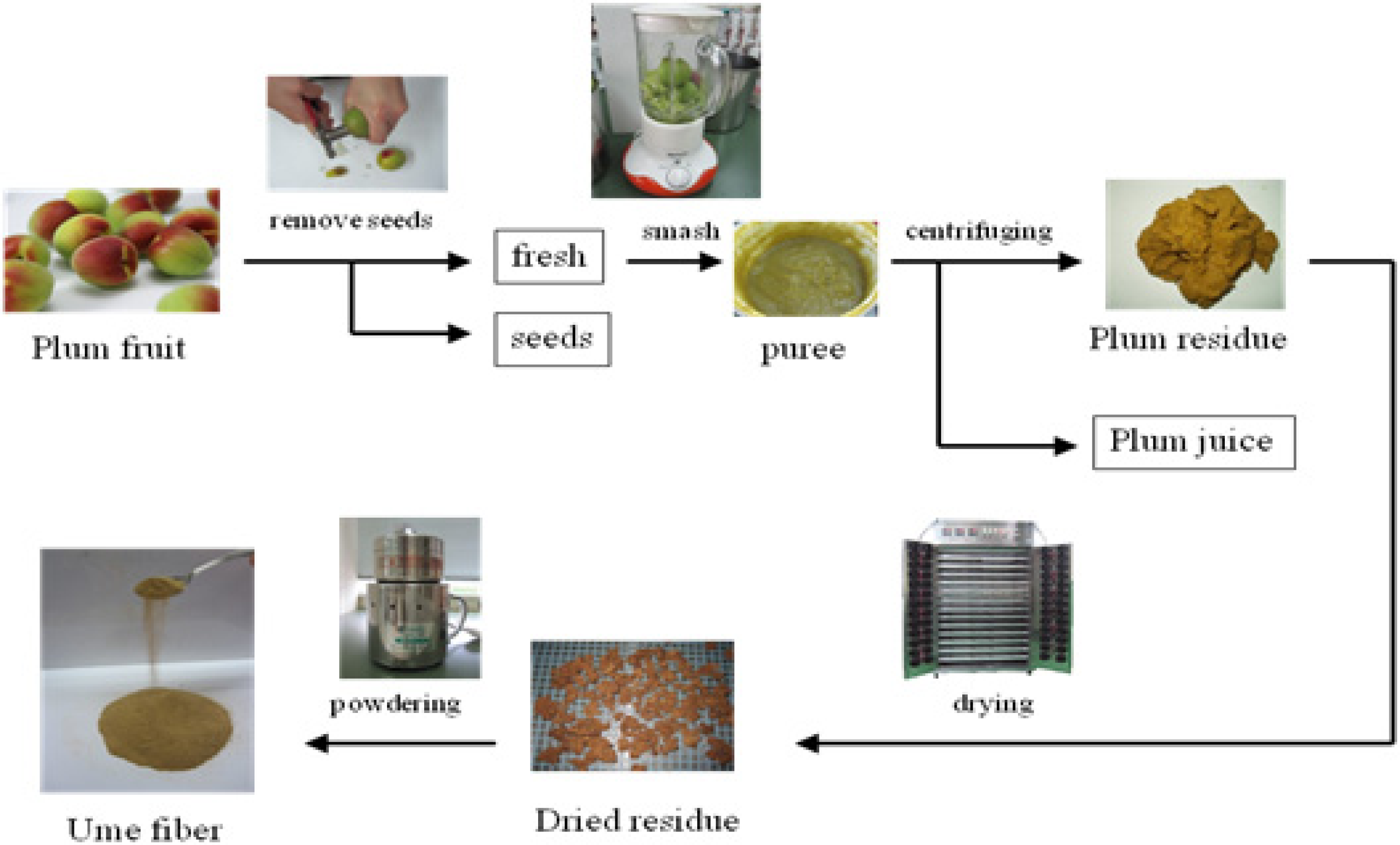
3.1. Production of Fiber from Fruit of Prunus mume
3.2. Treatment of Animals
3.3. Analysis of ume Fiber, Mice Feeding Conditions and Sampling
3.4. Measurement of Plasma Cholesterol, Triglyceride, Phospholipids, NEFA and Plasma Glucose
3.5. Fecal Lipid Extraction
3.6. DNA Extraction from Cecal Contents
3.7. PCR Conditions and Restriction Enzyme Digestion
3.8. T-RFLP Analysis
3.9. Statistics
4. Conclusions
Acknowledgments
References
- Chuda, Y; Ono, H; Ohnishi-Kameyama, M; Matsumoto, K; Nagata, T; Kikuchi, Y. Mumefural, citric acid derivative improving blood fluidity from fruit-juice concentrate of Japanese apricot (Prunus mume Sieb. et Zucc). J. Agr. Food. Chem 1999, 47, 828–831. [Google Scholar]
- Weickert, MO; Pfeiffer, AF. Metabolic effects of dietary fiber consumption and prevention of diabetes. J. Nutr 2008, 138, 439–442. [Google Scholar]
- Hillman, L; Peters, S; Fisher, A; Pomare, EW. Differing effects of pectin, cellulose and lignin on stool pH, transit time and weight. Br. J. Nutr 1983, 50, 189–195. [Google Scholar]
- Johnson, IT; Gee, JM; Brown, JC. Plasma enteroglucagon and small bowel cytokinetics in rats fed soluble nonstarch polysaccharides. Am. J. Clin. Nutr 1988, 47, 1004–1009. [Google Scholar]
- Demigné, C; Levrat, MA; Rémésy, C. Effects of feeding fermentable carbohydrates on the cecal concentrations of minerals and their fluxes between the cecum and blood plasma in the rat. J. Nutr 1989, 119, 1625–1630. [Google Scholar]
- Mao, Y; Kasravi, B; Nobaek, S; Wang, LQ; Adawi, D; Roos, G; Stenram, U; Molin, G; Bengmark, S; Jeppsson, B. Pectin-supplemented enteral diet reduces the severity of methotrexate induced enterocolitis in rats. Scand. J. Gastroenterol 1996, 31, 558–567. [Google Scholar]
- Armstrong, EF; Eastwood, MA; Brydon, WG. The influence of wheat bran and pectin on the distribution of water in rat caecal contents and faeces. Br. J. Nutr 1993, 69, 913–920. [Google Scholar]
- Wijnands, MV; Appel, MJ; Hollanders, VM; Woutersen, RA. A comparison of the effects of dietary cellulose and fermentable galacto-oligosaccharide, in a rat model of colorectal carcinogenesis: Fermentable fibre confers greater protection than non-fermentable fibre in both high and low fat backgrounds. Carcinogenesis 1999, 20, 651–656. [Google Scholar]
- Rao, CV; Hirose, Y; Indranie, C; Reddy, BS. Modulation of experimental colon tumorigenesis by types and amounts of dietary fatty acids. Cancer Res 2001, 61, 1927–1933. [Google Scholar]
- Zoran, DL; Turner, ND; Taddeo, SS; Chapkin, RS; Lupton, JR. Wheat bran diet reduces tumor incidence in a rat model of colon cancer independent of effects on distal luminal butyrate concentrations. J. Nutr 1997, 127, 2217–2225. [Google Scholar]
- Pool-Zobel, BL; Sauer, J. Overview of experimental data on reduction of colorectal cancer risk by inulin-type fructans. J. Nutr 2007, 137, 2580S–2584S. [Google Scholar]
- Okazaki, H; Nishimune, T; Matsuzaki, H; Miura, T; Morita, S; Yanagimoto, Y; Yamagishi, H; Yamada, K; Ikegami, S. Increased incidence rate of colorectal tumors due to the intake of a soluble dietary fiber in rat chemical carcinogenesis can be suppressed by substituting partially an insoluble dietary fiber for the soluble one. Int. J. Cancer 2002, 100, 388–394. [Google Scholar]
- Isken, F; Klaus, S; Osterhoff, M; Pfeiffer, AF; Weickert, MO. Effects of long-term soluble vs. insoluble dietary fiber intake on high-fat diet-induced obesity in C57BL/6J mice. J. Nutr. Biochem 2010, 21, 278–284. [Google Scholar]
- Nagashima, K; Hisada, T; Sato, M; Mochizuki, J. Application of new primer-enzyme combinations to terminal restriction fragment length polymorphism profiling of bacterial populations in human feces. Appl. Environ. Microbiol 2003, 69, 1251–1262. [Google Scholar]
- Nagashima, K; Mochizuki, J; Hisada, T; Suzuki, S; Shimomura, K. Phylogenetic analysis of 16S ribosomal RNA gene sequences from human fecal microbiota and improved utility of terminal restriction fragment length polymorphism profiling. Biosci. Microflora 2006, 25, 99–107. [Google Scholar]
- Licht, TR; Hansen, M; Bergström, A; Poulsen, M; Krath, BN; Markowski, J; Dragsted, LO; Wilcks, A. Effects of apples and specific apple components on the cecal environment of conventional rats: Role of apple pectin. BMC Microbiol 2010, 10, 13. [Google Scholar]
- Mozes, S; Bujnáková, D; Sefcíková, Z; Kmet, V. Intestinal microflora and obesity in rats. Folia. Microbiol. (Praha.) 2008, 53, 225–228. [Google Scholar]
- Turnbaugh, PJ; Ley, RE; Mahowald, MA; Magrini, V; Mardis, ER; Gordon, JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar]
- Allain, CC; Poon, LS; Chan, CS; Richmond, W; Fu, PC. Enzymatic determination of total serum cholesterol. Clin. Chem 1974, 20, 470–475. [Google Scholar]
- Spayd, RW; Bruschi, B; Burdick, BA; Dappen, GM; Eikenberry, JN; Esders, TW; Figueras, J; Goodhue, CT; LaRossa, DD; Nelson, RW; Rand, RN; Wu, TW. Multilayer film elements for clinical analysis: Applications to representative chemical determinations. Clin. Chem 1978, 24, 1343–1350. [Google Scholar]
- Takayama, M; Itoh, S; Nagasaki, T; Tanimizu, I. A new enzymatic method for determination of serum choline-containing phospholipids. Clin. Chim. Acta 1977, 79, 93–98. [Google Scholar]
- Bligh, EG; Dyer, WJ. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol 1959, 37, 911–917. [Google Scholar]
- Matsuki, T. Procedure of DNA extraction from fecal sample for the analysis of intestinal microflora. J. Intestinal. Microbiol 2006, 20, 259–262. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/kg diet) | AIN-93M | UF diet | CF diet |
|---|---|---|---|
| Corn starch | 465.692 | 388.246 | 405.686 |
| Casein | 140 | 129.52 | 140 |
| α-Corn starch | 155 | 155 | 155 |
| Sucrose | 100 | 100 | 100 |
| Rice bran oil | - | 97.52 | 100 |
| Soy bean oil | 40 | - | - |
| Cellulose | 50 | - | 50 |
| Ume fiber | - | 80.4 | - |
| Mineral mix (AIN-93M-Mix) | 35 | 35 | 35 |
| Vitamin mix (AIN-93-Mix) | 10 | 10 | 10 |
| L-Cystine | 1.8 | 1.8 | 1.8 |
| Choline Bitartrate | 2.5 | 2.5 | 2.5 |
| Tert-butylhydroquinone | 0.008 | 0.014 | 0.014 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tamura, M.; Ohnishi, Y.; Kotani, T.; Gato, N. Effects of New Dietary Fiber from Japanese Apricot (Prunus mume Sieb. et Zucc.) on Gut Function and Intestinal Microflora in Adult Mice. Int. J. Mol. Sci. 2011, 12, 2088-2099. https://doi.org/10.3390/ijms12042088
Tamura M, Ohnishi Y, Kotani T, Gato N. Effects of New Dietary Fiber from Japanese Apricot (Prunus mume Sieb. et Zucc.) on Gut Function and Intestinal Microflora in Adult Mice. International Journal of Molecular Sciences. 2011; 12(4):2088-2099. https://doi.org/10.3390/ijms12042088
Chicago/Turabian StyleTamura, Motoi, Yuriko Ohnishi, Tatsuya Kotani, and Nobuki Gato. 2011. "Effects of New Dietary Fiber from Japanese Apricot (Prunus mume Sieb. et Zucc.) on Gut Function and Intestinal Microflora in Adult Mice" International Journal of Molecular Sciences 12, no. 4: 2088-2099. https://doi.org/10.3390/ijms12042088