Role of Rho Kinase in Microvascular Damage Following Cerebral Ischemia Reperfusion in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and MCAO Model
2.2. Ischemic Core and Penumbra Dissections
2.3. Determination of BBB Permeability
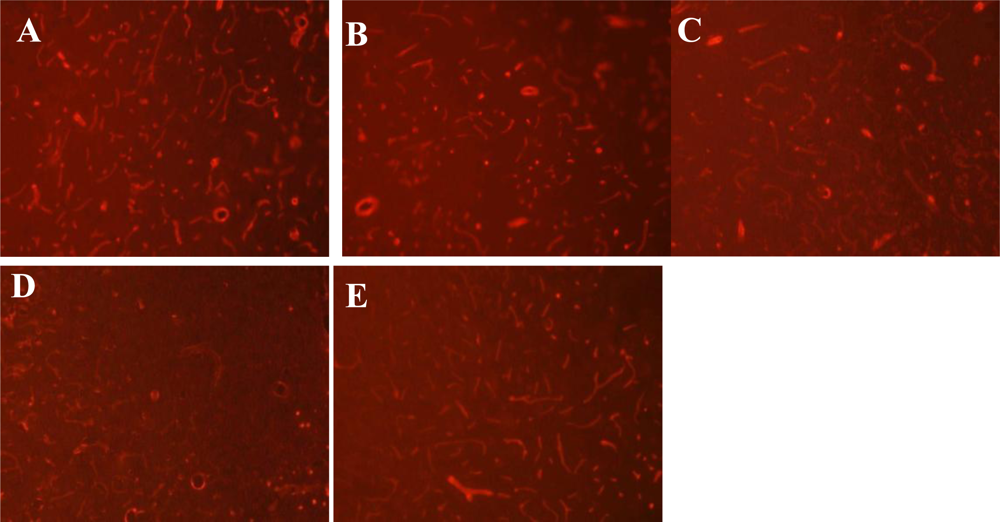
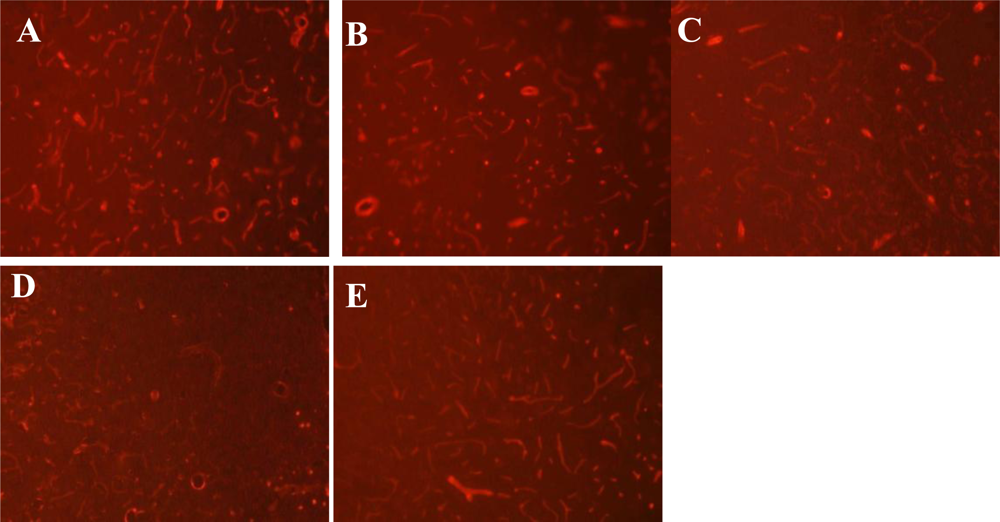
2.4. Immunofluorescence Staining for Laminin
2.5. RNA Isolation and Real-Time Quantitative PCR
2.6. Statistical Analyses
3. Results
3.1. Expression of ROCK at Different Time Points after MCAO
3.2. BBB Permeability at Different Time Points after MCAO
3.3. Expression of Laminin at Different Time Points after MCAO
3.4. Correlation among ROCK mRNA Level, Brain EB Content and Laminin Expression after MCAO
3.5. ROCK Inhibitor Decreases BBB Permeability after MCAO
3.6. ROCK Inhibitor Rescues Laminin Expression after MCAO
3.7. ROCK Inhibitor Inhibits MMP9 Expression after MCAO
4. Discussion
Acknowledgments
References
- Doyle, KP; Simon, RP; Stenzel-Poore, MP. Mechanisms of ischemic brain damage. Neuropharmacology 2008, 55, 310–318. [Google Scholar]
- Weiss, N; Miller, F; Cazaubon, S; Couraud, PO. The blood-brain barrier in brain homeostasis and neurological diseases. Biochim. Biophys. Acta 2009, 1788, 842–857. [Google Scholar]
- Montaner, J; Alvarez-Sabin, J; Molina, CA; Angles, A; Abilleira, S; Arenillas, J; Monasterio, J. Matrix metalloproteinase expression is related to hemorrhagic transformation after cardioembolic stroke. Stroke 2001, 32, 2762–2767. [Google Scholar]
- Asahi, M; Wang, X; Mori, T; Sumii, T; Jung, JC; Moskowitz, MA; Fini, ME; Lo, EH. Effects of matrix metalloproteinase-9 gene knock-out on the proteolysis of blood-brain barrier and white matter components after cerebral ischemia. J. Neurosci 2001, 21, 7724–7732. [Google Scholar]
- Rikitake, Y; Kim, HH; Huang, Z; Seto, M; Yano, K; Asano, T; Moskowitz, MA; Liao, JK. Inhibition of Rho kinase (ROCK) leads to increased cerebral blood flow and stroke protection. Stroke 2005, 36, 2251–2257. [Google Scholar]
- Longa, EZ; Weinstein, PR; Carlson, S; Cummins, R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 1989, 20, 84–91. [Google Scholar]
- Ashwal, S; Tone, B; Tian, HR; Cole, DJ; Pearce, WJ. Core and penumbral nitric oxide synthase activity during cerebral ischemia and reperfusion. Stroke 1998, 29, 1037–1046. [Google Scholar]
- Zhang, W; Zhao, WQ; Lu, HD. Change of neurovascular unit after ischemia-reperfusion damage in rat brain. Chin. J. Geriatr. Heart Brain Vessel Dis 2008, 10, 620–622. [Google Scholar]
- Arita, R; Hata, Y; Nakao, S; Kita, T; Miura, M; Kawahara, S; Zandi, S; Almulki, L; Tayyari, F; Shimokawa, H; et al. Rho kinase inhibition by fasudil ameliorates diabetes-induced microvascular damage. Diabetes 2009, 58, 215–226. [Google Scholar]
- Gasche, Y; Soccal, PM; Kanemitsu, M; Copin, JC. Matrix mettalloproteinases and diseases of the central nervous system with a special emphasis on ischemic brain. Front Biosci 2006, 11, 1289–1301. [Google Scholar]
- Cuadrado, E; Rosell, A; Penalba, A; Slevin, M; Alvarez-Sabín, J; Ortega-Aznar, A; Montaner, J. Vascular MMP-9/TIMP-2 and neuronal MMP-10 up-regulation in human brain after stroke: A combined laser microdissection and protein array study. J. Proteome Res 2009, 8, 3191–3197. [Google Scholar]
- Lescot, T; Fulla-Oller, L; Palmier, B; Po, C; Beziaud, T; Puybasset, L; Plotkine, M; Gillet, B; Meric, P; Marchand-Leroux, C. Effect of acute Poly(ADP-Ribose) polymerase inhibition by 3-AB on blood-brain barrier permeability and edema formation after focal traumatic brain injury in rats. J. Neurotrauma 2010, 27, 1069–1079. [Google Scholar]
- Maddahi, A; Chen, Q; Edvinsson, L. Enhanced cerebrovascular expression of matrix metalloproteinase-9 and tissue inhibitor of metalloproteinase-1 via the MEK/ERK pathway during cerebral ischemia in the rat. BMC Neurosci 2009, 4, 56. [Google Scholar]
- Barr, TL; Latour, LL; Lee, KY; Schaewe, TJ; Luby, M; Chang, GS; El-Zammar, Z; Alam, S; Hallenbeck, JM; Kidwell, CS; et al. Blood-brain barrier disruption in humans is independently associated with increased matrix metalloproteinase-9. Stroke 2010, 41, 123–128. [Google Scholar]
- Wakisaka, Y; Chu, Y; Miller, JD; Rosenberg, GA; Heistad, DD. Spontaneous intracerebral hemorrhage during acute and chronic hypertension in mice. J. Cereb. Blood Flow Metab 2010, 30, 56–69. [Google Scholar]
- Guo, Z; Sun, X; He, Z; Jiang, Y; Zhang, X. Role of matrix metalloproteinase-9 in apoptosis of hippocampal neurons in rats during early brain injury after subarachnoid hemorrhage. Neurol. Sci 2010, 31, 143–149. [Google Scholar]


{kind=link}
{kind=link}
| Group | n | ROCK mRNA | EB content (μg/g) | Laminin expression |
|---|---|---|---|---|
| sham | 5 | 0.466 ± 0. 030 | 0.202 ±0.019 | 168.91 ± 6.73 |
| ischemia reperfusion | ||||
| reperfusion 6 h | 5 | 0.617 ± 0.017 * | 1.372 ± 0.123 * | 149.45 ± 4.38 * |
| reperfusion 24 h | 5 | 0.662 ± 0.016 * | 3.764 ± 0.160 * | 115.56 ± 8.29 * |
| reperfusion 48 h | 5 | 0.838 ± 0.036 * | 4.650 ± 0.286 * | 82.95 ± 12.33 * |
| reperfusion 72 h | 5 | 0.713 ± 0.018 * | 2.982 ± 0.163 * | 124.34 ± 5.29 * |
| Group | EB content (mg/g) (n = 5) | Laminin expression (n = 5) | MMP9 mRNA (n = 10) |
|---|---|---|---|
| sham | 0.210 ± 0.039 | 167.89 ± 8.97 | 0.514 ± 0.055 |
| MCAO | 3.376 ± 0. 134 * | 109.72 ± 12.01 * | 0.762 ± 0.021 * |
| Fasudil | 1.930 ± 0.079 *,† | 142.02 ± 4.82 *,† | 0.659 ± 0.028 *,† |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, K.; Li, Z.; Wu, T.; Ding, S. Role of Rho Kinase in Microvascular Damage Following Cerebral Ischemia Reperfusion in Rats. Int. J. Mol. Sci. 2011, 12, 1222-1231. https://doi.org/10.3390/ijms12021222
Liu K, Li Z, Wu T, Ding S. Role of Rho Kinase in Microvascular Damage Following Cerebral Ischemia Reperfusion in Rats. International Journal of Molecular Sciences. 2011; 12(2):1222-1231. https://doi.org/10.3390/ijms12021222
Chicago/Turabian StyleLiu, Kang, Zhen Li, Tao Wu, and Suju Ding. 2011. "Role of Rho Kinase in Microvascular Damage Following Cerebral Ischemia Reperfusion in Rats" International Journal of Molecular Sciences 12, no. 2: 1222-1231. https://doi.org/10.3390/ijms12021222