Environmental Signals and Regulatory Pathways That Influence Exopolysaccharide Production in Rhizobia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
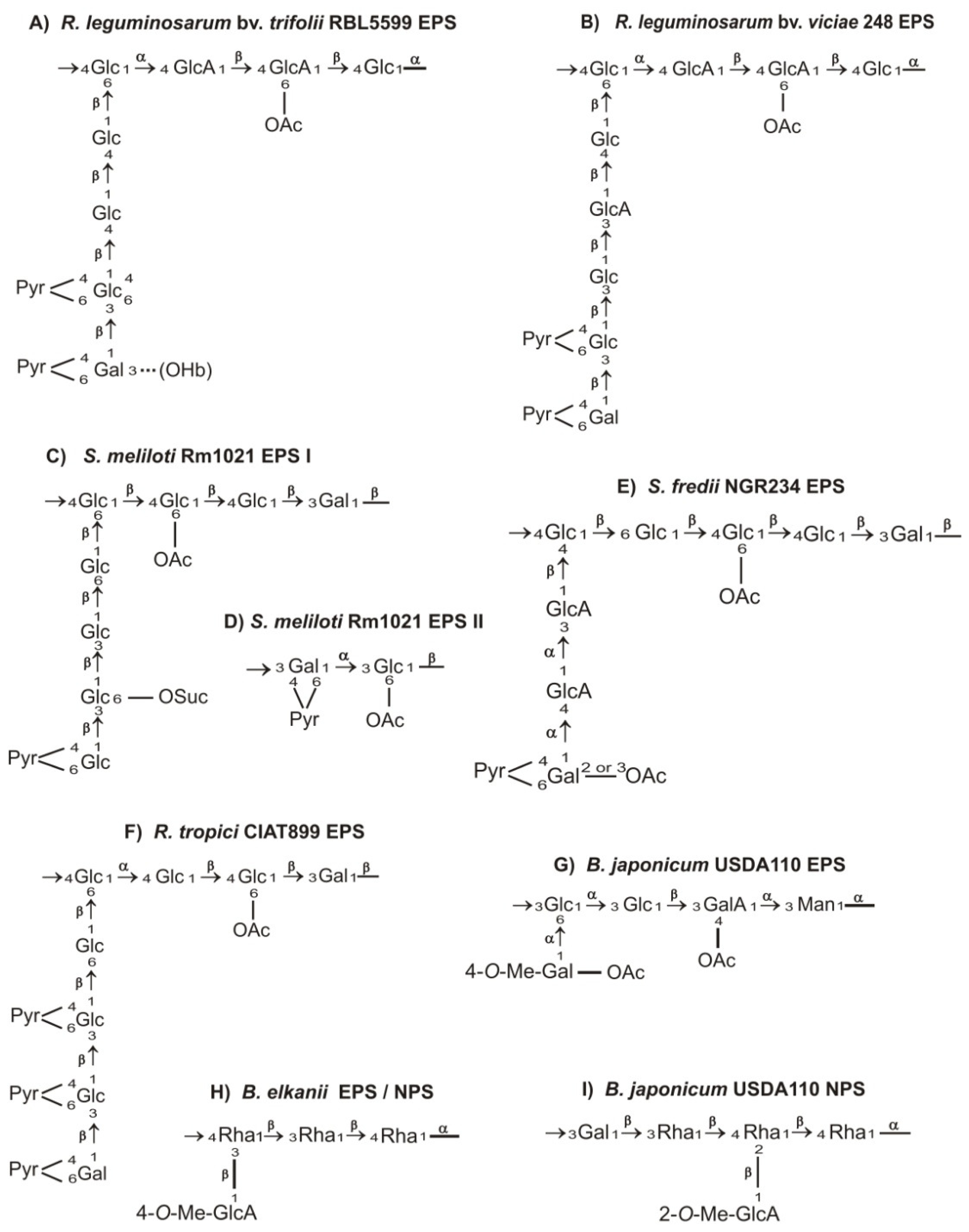
2. Chemical Structure of Rhizobial Exopolysaccharides
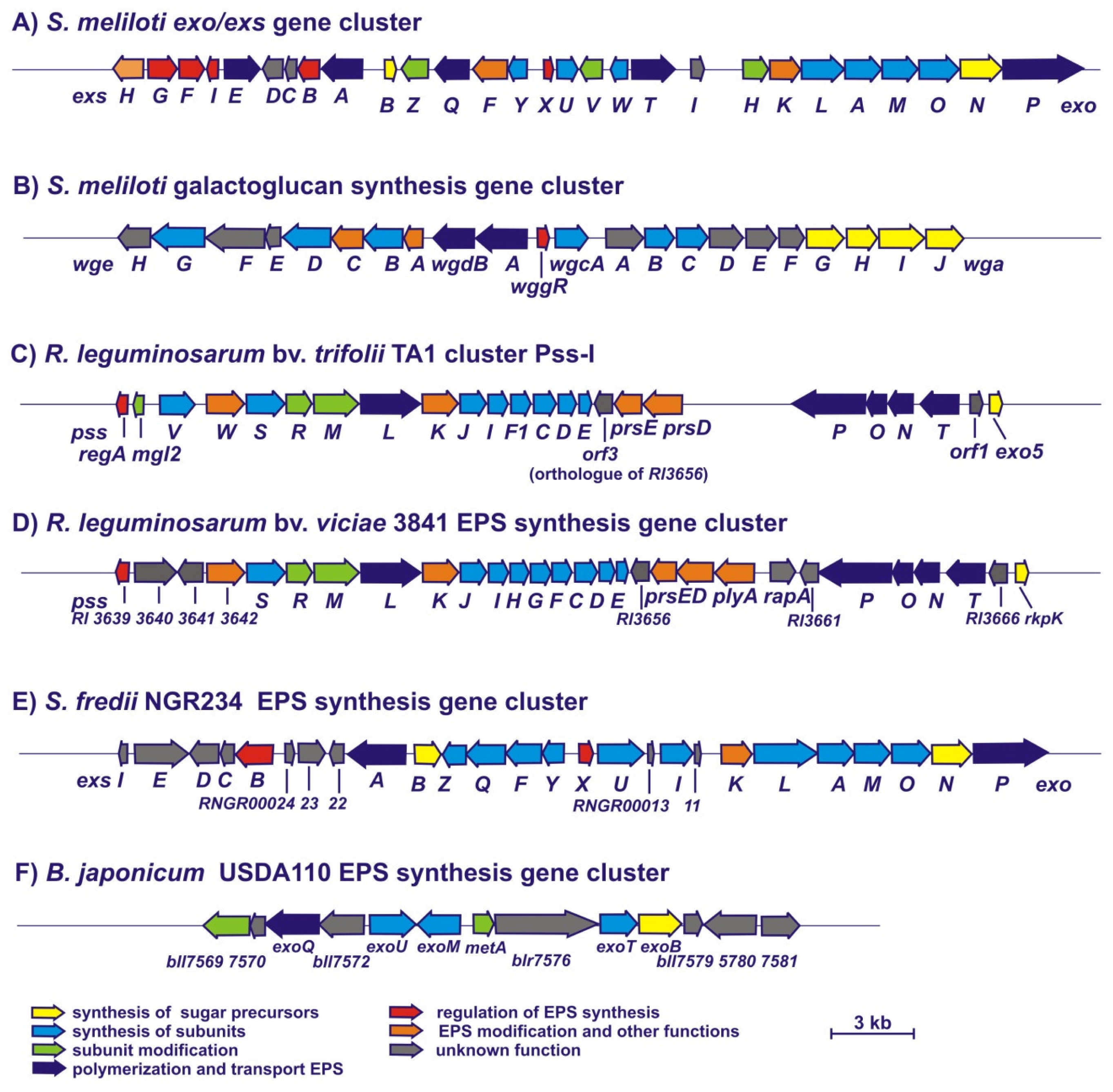
3. Genetic Control of EPS Synthesis in Rhizobia
3.1. Genes Involved in the Synthesis of S. meliloti EPS I and EPS II
3.2. Genes Involved in the Synthesis of EPS in R. leguminosarum
3.3. Genes Involved in the Synthesis of EPS in Other Rhizobia
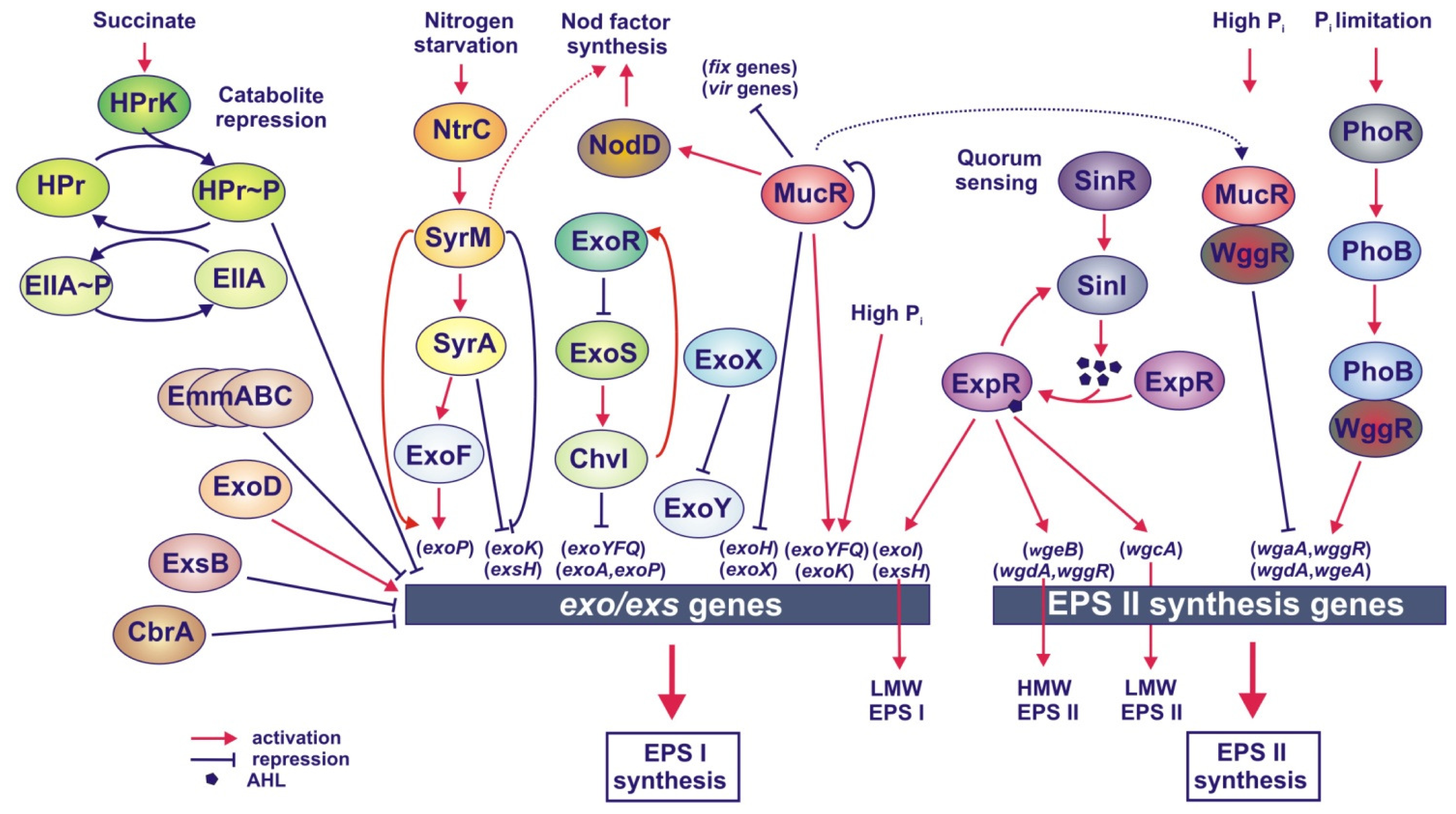
4. Regulation of EPS Biosynthesis
4.1. Regulation of EPS Synthesis in S. meliloti
4.1.1. Regulation of EPS I Synthesis
4.1.2. Regulation of EPS I Synthesis by Succinate-Mediated Catabolite Repression
4.1.3. Regulation of EPS II Synthesis
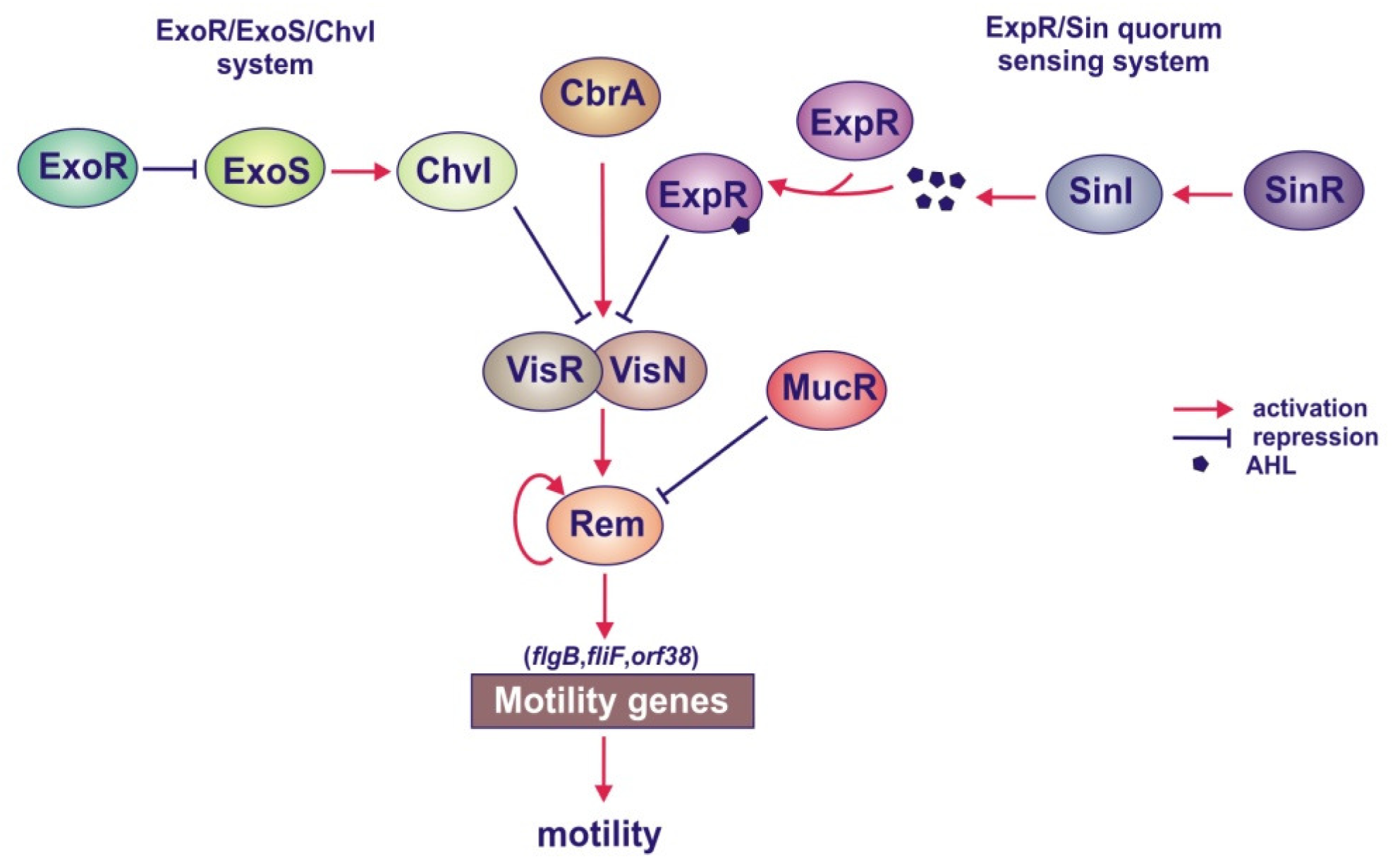
4.1.4. Role of Quorum Sensing in the Regulation of EPS I and EPS II Synthesis
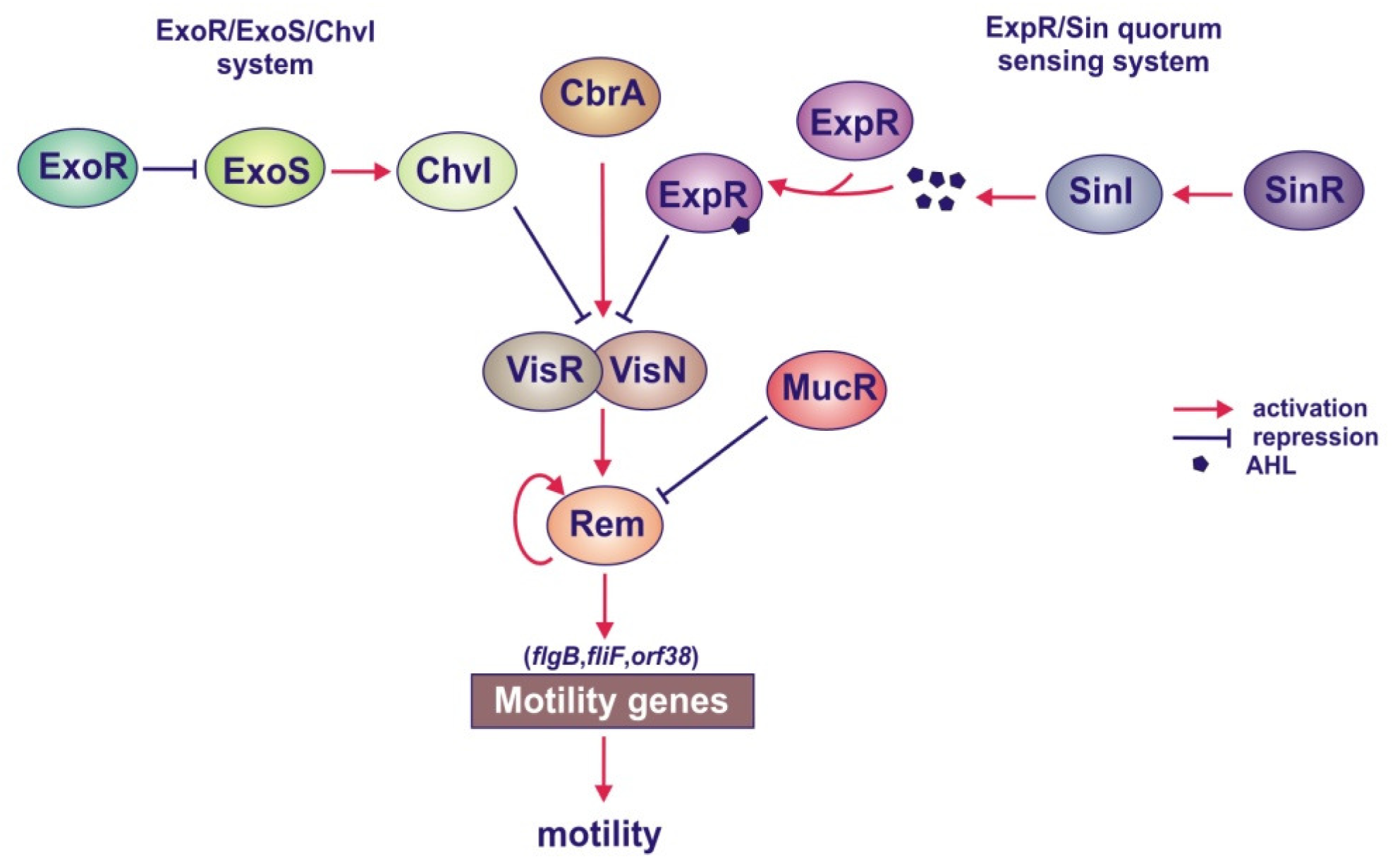
4.1.5. Roles of the MucR, ExoR/ExoS/ChvI and ExpR/Sin Systems in the Regulation of Motility Genes
4.2. Regulation of EPS Synthesis in R. leguminosarum
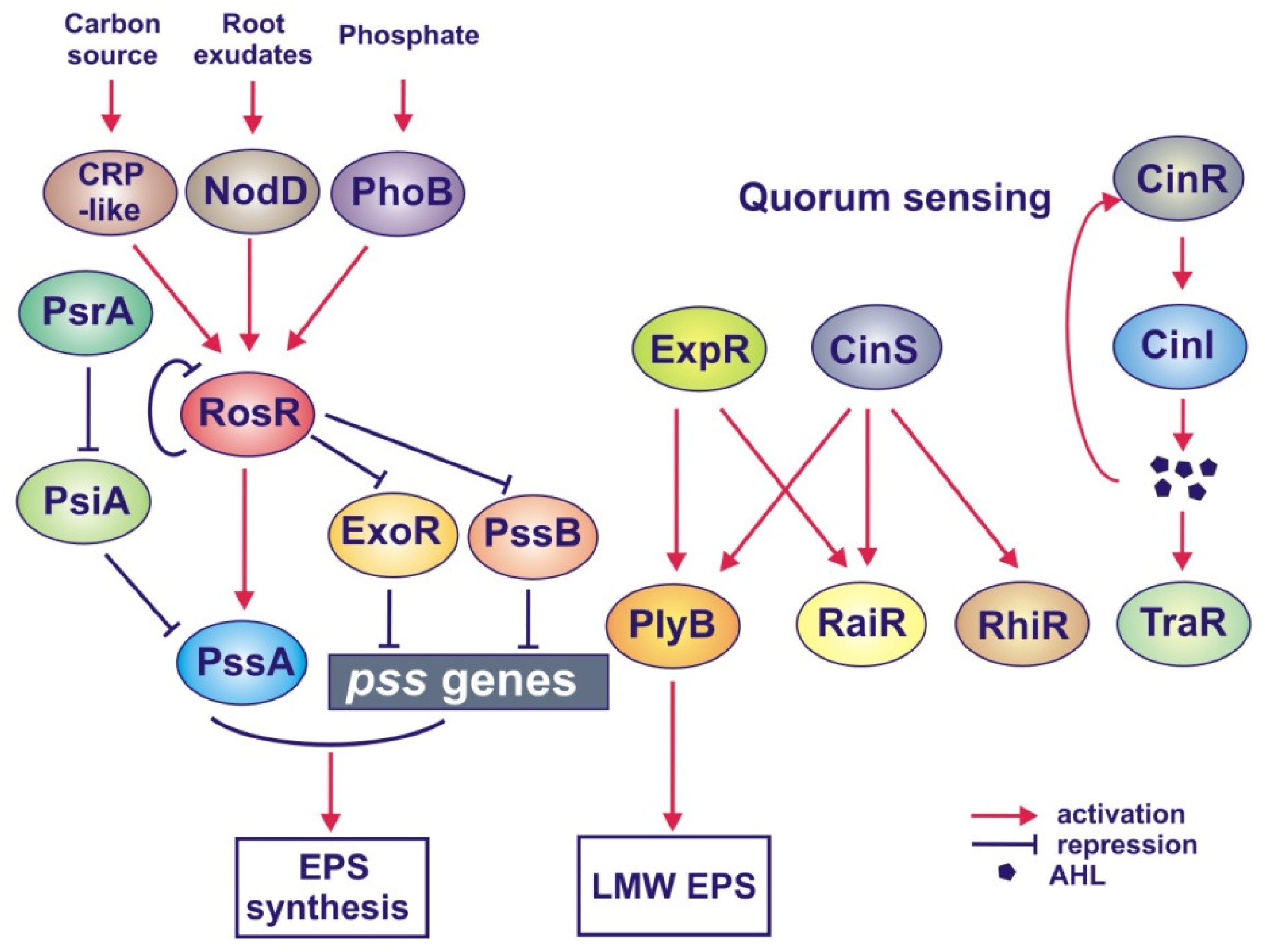
4.2.1. Role of RosR in EPS Synthesis and Symbiosis
4.2.2. Role of Quorum Sensing in the Regulation of EPS Synthesis
4.3. Role of EPS Synthesis Genes in Biofilm Formation in Rhizobia
5. Conclusions
Acknowledgements
Abbreviations
| Glc | glucose |
| GlcA | glucuronic acid |
| Gal | galactose |
| Man | mannose |
| Rha | rhamnose |
| OAc | acetyl |
| Pyr | ketal pyruvate |
| Suc | succinyl |
| OHb | hydroxybutanoyl |
| Me | methyl groups |
References
- Dresler-Nurmi, A.; Fewer, D.P.; Räsänen, L.A.; Lindstrőm, K. The diversity and evolution of rhizobia. Microbiol. Monogr 2009, 8, 3–41. [Google Scholar]
- Gibson, K.E.; Kobayashi, H.; Walker, G.C. Molecular determinants of a symbiotic chronic infection. Annu. Rev. Genet 2008, 42, 413–441. [Google Scholar]
- Skorupska, A.; Janczarek, M.; Marczak, M.; Mazur, A.; Król, J. Rhizobial exopolysaccharides: Genetic control and symbiotic functions. Microb. Cell Fact 2006, 5. [Google Scholar] [CrossRef]
- Downie, J.A. The roles of extracellular proteins, polysaccharides and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev 2010, 34, 150–170. [Google Scholar]
- Fraysse, N.; Couderc, F.; Poinsot, V. Surface polysaccharide involvement in establishing the rhizobium-legume symbiosis. Eur. J. Biochem 2003, 270, 1365–1380. [Google Scholar]
- Laus, M.C.; Logman, T.J.; Lamers, G.E.; van Brussel, A.A.; Carlson, R.W.; Kijne, J.W. A novel polar surface polysaccharide from Rhizobium leguminosarum binds host plant lectin. Mol. Microbiol 2006, 59, 1704–1713. [Google Scholar]
- Becker, A.; Fraysse, N.; Sharypova, L. Recent advances in studies on structure and symbiosis-related function of rhizobial K-antigens and lipopolysaccharides. Mol. Plant Microbe Interact 2005, 18, 899–905. [Google Scholar]
- Breedveld, M.; Miller, K.J. Cyclic β-glucans of members of the family Rhizobiaceae. Microbiol. Rev 1994, 58, 145–161. [Google Scholar]
- Crespo-Rivas, J.C.; Margaret, I.; Hidalgo, A.; Buendía-Clavería, A.M.; Ollero, F.J.; López-Baena, F.J.; del Socorro Murdoch, P.; Rodríguez-Carvajal, M.A.; Soria-Díaz, M.E.; Reguera, M.; et al. Sinorhizobium fredii HH103 cgs mutants are unable to nodulate determinate-and indeterminate nodule—forming legumes and overproduce an altered EPS. Mol. Plant Microbe Interact 2009, 22, 575–588. [Google Scholar]
- Rolfe, B.G.; Carlson, R.W.; Ridge, R.W.; Dazzo, R.W.; Mateos, F.B.; Pankhurst, C.E. Defective infection and nodulation of clovers by exopolysaccharide mutants of Rhizobium leguminosarum bv trifolii. Aust. J. Plant Physiol 1996, 23, 285–303. [Google Scholar]
- Cheng, H.-P.; Walker, G.C. Succinoglycan is required for initiation and elongation of infection threads during nodulation of alfalfa by Rhizobium meliloti. J. Bacteriol 1998, 180, 5183–5191. [Google Scholar]
- van Workum, W.A.; Canter Cremers, H.C.J.; Wijfjes, A.H.M.; van der Kolk, C.; Wijffelman, C.A.; Kijne, J.W. Cloning and characterization of four genes of Rhizobium leguminosarum bv. trifolii involved in exopolysaccharide production and nodulation. Mol. Plant Microbe Interact 1997, 10, 290–301. [Google Scholar]
- Williams, M.N.; Hollingsworth, R.I.; Klein, S.; Signer, E.R. The symbiotic defect of Rhizobium meliloti exopolysaccharide mutants is suppressed by lpsZ+, a gene involved in lipopolysaccharide biosynthesis. J. Bacteriol 1990, 172, 2622–2632. [Google Scholar]
- Reuhs, B.L.; Williams, M.N.; Kim, J.S.; Carlson, R.W.; Cote, F. Suppression of the FixZ phenotype of Rhizobium meliloti exoB mutants by lpsZ is correlated to a modified expression of the K polysaccharide. J. Bacteriol 1995, 177, 4289–4296. [Google Scholar]
- Margaret-Oliver, I.; Lei, W.; Parada, M.; Rodríguez-Carvajal, M.A.; Crespo-Rivas, J.C.; Hidalgo, A.; Gil-Serrano, A.; Moreno, J.; Rodríguez-Navarro, D.N.; Buendía-Clavería, A.; et al. Sinorhizobium fredii HH103 does not strictly require KPS and/or EPS to nodulate Glycyrrhiza uralensis, an indeterminate nodule-forming legume. Arch. Microbiol 2011. [Google Scholar] [CrossRef]
- Quelas, J.I.; Mongiardini, E.J.; Casabuono, A.; López-García, S.L.; Althabegoiti, M.J.; Covelli, J.M.; Pérez-Giménez, J.; Couto, A.; Lodeiro, A.R. Lack of galactose or galacturonic acid in Bradyrhizobium japonicum USDA 110 exopolysaccharide leads to different symbiotic responses in soybean. Mol. Plant Microbe Interact 2010, 23, 1592–1604. [Google Scholar]
- Rinaudi, L.V.; Giordano, W. An integrated view of biofilm formation in rhizobia. FEMS Microbiol. Lett 2010, 304, 1–11. [Google Scholar]
- Amemura, A.; Harada, T.; Abe, M.; Higashi, S. Structural studies of the acidic polysaccharide from Rhizobium trifolii 4S. Carbohydr. Res 1983, 115, 165–174. [Google Scholar]
- Djordjevic, S.P.; Chen, H.; Batley, M.; Redmond, J.W.; Rolfe, B.G. Nitrogen fixation ability of exopolysaccharide synthesis mutants of Rhizobium sp. strain NGR234 and Rhizobium trifolii is restored by addition of homologous exopolysaccharides. J. Bacteriol 1987, 169, 53–60. [Google Scholar]
- Gonzalez, J.E.; Semino, C.E.; Wang, L.X.; Castellano-Torres, L. Biosynthetic control of molecular weight in the polymerization of the octasaccharide subunits of succinoglycan, a symbiotically important exopolysaccharide of Rhizobium meliloti. Proc. Natl. Acad. Sci. USA 1998, 95, 13477–13482. [Google Scholar]
- Robertson, B.K.; Aman, P.; Darvill, A.G.; McNeil, M.; Albersheim, P. Host-symbiont interactions. The structure of acidic extracellular polysaccharides secreted by Rhizobium leguminosarum and Rhizobium trifolii. Plant Physiol 1981, 67, 389–400. [Google Scholar]
- O’Neill, M.A.; Darvill, A.G.; Albersheim, P. The degree of esterification and points of substitution by O-acetyl and O-(3-hydroxybutanoyl) groups in the acidic extracellular polysaccharides secreted by Rhizobium leguminosarum biovars viciae, trifolii, and phaseoli are not related to host range. J. Biol. Chem 1991, 266, 9549–9555. [Google Scholar]
- Breedveld, M.W.; Canter Cremers, H.C.J.; Batley, M.; Posthumus, M.A.; Zevenhuizen, L.P.T.M.; Wijfelman, C.A.; Zehnder, A.J.B. Polysaccharide synthesis in relation to nodulation behaviour of Rhizobium leguminosarum. J. Bacteriol 1993, 175, 750–757. [Google Scholar]
- McNeil, M.; Darvill, J.; Darvill, A.; Albersheim, P.; van Veen, R.; Hooykaas, P.; Schilperoort, R.; Dell, A. The discernible structural features of the acidic exopolysaccharides secreted by different Rhizobium species are the same. Carbohydr. Res 1986, 146, 307–326. [Google Scholar]
- Philip-Hollingsworth, S.; Hollingsworth, R.I.; Dazzo, F.B. Host-range related structural features of the acidic extracellular polysaccharides of Rhizobium trifolii and Rhizobium leguminosarum. J. Biol. Chem 1989, 264, 1461–1466. [Google Scholar]
- Canter Cremers, H.C.J.; Stevens, K.; Lugtenberg, B.J.J.; Wijffelman, C.A.; Batley, M.; Redmond, J.W.; Breedveld, M.; Zevenhuizen, L.P.T.M. Unusual structure of the exopolysaccharide of Rhizobium leguminosarum biovar viciae strain 248. Carbohydr. Res 1991, 218, 185–200. [Google Scholar]
- Zhan, H; Lee, C.C.; Leigh, J.A. Induction of the second exopolysaccharide (EPSb) in Rhizobium meliloti SU47 by low phosphate concentrations. J. Bacteriol 1991, 173, 7391–7394. [Google Scholar]
- Reinhold, B.B.; Chan, S.Y.; Reuber, T.L.; Marra, A.; Walker, G.C.; Reinhold, V.N. Detailed structural characterization of succinoglycan, the major exopolysaccharide of Rhizobium meliloti Rm 1021. J. Bacteriol 1994, 176, 1997–2002. [Google Scholar]
- Zevenhuizen, L.P.T.M. Succinoglycan and galactoglucan. Carbohydr. Polym 1997, 33, 139–144. [Google Scholar]
- Reuber, T.L.; Walker, G.C. Biosynthesis of succinoglycan, a symbiotically important exopolysaccharide of Rhizobium meliloti. Cell 1993, 74, 269–280. [Google Scholar]
- Her, G.R.; Glazebrook, J.; Walker, G.C.; Reinhold, V.N. Structural studies of a novel exopolysaccharide produced by a mutant of Rhizobium meliloti strain Rm 1021. Carbohydr. Res 1990, 198, 305–312. [Google Scholar]
- Djordjevic, S.P.; Batley, M.; Redmond, J.W.; Rolfe, B.G. The structure of the exopolysaccharide from Rhizobium sp. strain ANU280 (NGR234). Carbohydr. Res 1986, 148, 87–99. [Google Scholar]
- Gil-Serrano, A.; Del Junco, S.A.; Tejero-Mateo, P. Structure of extracellular polysaccharide secreted by Rhizobium leguminosarum bv. phaseoli CIAT899. Carbohydr. Res 1990, 204, 103–107. [Google Scholar]
- Minamisawa, K. Comparison of extracellular polysaccharide composition, rhizobiotoxine production, and hydrogenase phenotype among various strains of Bradyrhizobium japonicum. Plant Cell Physiol 1989, 30, 877–884. [Google Scholar]
- Poveda, A.; Santamaria, M.; Bernabe, M.; Prieto, A.; Briux, M.; Corzo, J.; Jimenez-Barbero, J. Studies on the structure and the solution conformation of an acidic extracellular polysaccharide isolated from Bradyrhizobium. Carbohydr. Res 1997, 304, 209–217. [Google Scholar]
- An, J.; Carlson, R.W.; Glushka, J.; Streeter, J.G. The structure of a novel polysaccharide produced by Bradyrhizobium species within soybean nodules. Carbohydr. Res 1995, 269, 303–317. [Google Scholar]
- Streeter, J.G.; Salminen, S.O.; Whitmoyer, R.F.; Carlson, R.W. Formation of novel polysaccharide by Bradyrhizobium japonicum bacteroids in soybean nodules. Appl. Environ. Microbiol 1992, 58, 607–613. [Google Scholar]
- Streeter, J.G. Failure of inoculant rhizobia to overcome the dominance of indigenous strains for nodule formation. Can. J. Microbiol 1994, 40, 513–522. [Google Scholar]
- D’Haeze, W.; Glushka, J.; De Rycke, R.; Holsters, M.; Carlson, R.W. Structural characterization of extracellular polysaccharides of Azorhizobium caulinodans and importance for nodule initiation on Sesbania rostrata. Mol. Microbiol 2004, 52, 485–500. [Google Scholar]
- Finan, T.M.; Weidner, S.; Womg, K.; Buhrmester, J.; Chain, P.; Vorhölter, F.J.; Hernandez-Lucas, I.; Becker, A.; Cowie, A.; Gouzy, J.; et al. The complete sequence of the 1683-kb pSymB megaplasmid from the N2-fixing endosymbiont Sinorhizobium meliloti. Proc. Natl. Acad. Sci. USA 2001, 98, 9889–9894. [Google Scholar]
- Young, J.P.W.; Crossman, L.C.; Johnston, A.W.B.; Thomson, N.R.; Ghazoui, Z.F.; Hull, K.H.; Wexler, M.; Curson, A.R.; Todd, J.D.; Poole, P.S.; et al. The genome of Rhizobium leguminosarum has recognizable core and accessory components. Genome Biol 2006, 7. [Google Scholar] [CrossRef]
- Król, J.E.; Mazur, A.; Marczak, M.; Skorupska, A. Syntenic arrangements of the surface polysaccharide biosynthesis genes in Rhizobium leguminosarum. Genomics 2007, 89, 237–247. [Google Scholar]
- González, V.; Santamaría, R.I.; Bustos, P.; Hernández-González, I.; Medrano-Soto, A.; Moreno-Hagelsieb, G.; Janga, S.C.; Ramírez, M.A.; Jiménez-Jacinto, V.; Collado-Vides, J.; et al. The partitioned Rhizobium etli genome: Genetic and metabolic redundancy in seven interacting replicons. Proc. Natl. Acad. Sci. USA 2006, 103, 3834–3839. [Google Scholar]
- Reeve, W.; O’Hara, G.; Chain, P.; Ardley, J.; Bräu, L.; Nandesena, K.; Tiwari, R.; Copeland, A.; Nolan, M.; Han, C.; et al. Complete genome sequence of Rhizobium leguminosarum bv. trifolii strain WSM1325, an effective microsymbiont of annual Mediterranean clovers. Stand. Genomic Sci 2010, 2, 347–356. [Google Scholar]
- Reeve, W.; O’Hara, G.; Chain, P.; Ardley, J.; Bräu, L.; Nandesena, K.; Tiwari, R.; Malfatti, S.; Kiss, H.; Lapidus, A.; et al. Complete genome sequence of Rhizobium leguminosarum bv. trifolii strain WSM2304, an effective microsymbiont of the South American clover Trifolium polymorphum. Stand. Genomic Sci 2010, 2, 66–76. [Google Scholar]
- Kaneko, T.; Nakamura, Y.; Sato, S.; Asamizu, E.; Kato, T.; Sasamoto, S.; Watanabe, A.; Idesawa, K.; Ishikawa, A.; Kawashima, K.; et al. Complete genome structure of the nitrogen-fixing symbiotic bacterium Mesorhizobium loti. DNA Res 2000, 7, 331–338. [Google Scholar]
- Kaneko, T.; Nakamura, Y.; Sato, S.; Minamisawa, K.; Uchiumi, T.; Sasamoto, S.; Watanabe, A.; Idesawa, K.; Iriguchi, M.; Kawashima, K.; et al. Complete genomic sequence of nitrogen-fixing symbiotic bacterium Bradyrhizobium japonicum USDA110. DNA Res 2002, 9, 225–256. [Google Scholar]
- Whitfield, C.; Paiment, A. Biosynthesis and assembly of Group I capsular polysaccharides in Escherichia coli and related extracellular polysaccharides in other bacteria. Carbohydr. Res 2003, 338, 2491–2502. [Google Scholar]
- Liu, D.; Cole, R.A.; Reeves, P.R. An O-antigen processing function for Wzx(RfbX): A promising candidate for O-unit flippase. J. Bacteriol 1996, 178, 2102–2107. [Google Scholar]
- Paulsen, I.T.; Beness, A.M.; Saier, M.H. Computer-based analyses of the protein constituents of transport systems catalyzing export of complex carbohydrates in bacteria. Microbiology 1997, 143, 2685–2699. [Google Scholar]
- Glucksmann, M.A.; Reuber, T.L.; Walker, G.C. Genes needed for the modification, polimerization, export and processing of succinoglycan by Rhizobium meliloti: A model for succinoglycan biosynthesis. J. Bacteriol 1993, 175, 7045–7055. [Google Scholar]
- Glucksmann, M.A.; Reuber, T.L.; Walker, G.C. Family of glycosyl transferases needed for the synthesis of succinoglycan by Rhizobium meliloti. J. Bacteriol 1993, 175, 7033–7044. [Google Scholar]
- Becker, A.; Kleickmann, A.; Arnold, W.; Pühler, A. Analysis of the Rhizobium meliloti exoH/exoK/exoL fragment: ExoK shows homology to excreted endo-β-1,3-1,4-glucanases and ExoH resembles membrane proteins. Mol. Gen. Genet 1993, 238, 145–154. [Google Scholar]
- Becker, A.; Kleickmann, A.; Keller, M.; Arnold, W.; Pühler, A. Identification and analysis of the Rhizobium meliloti exoAMONP genes involved in exopolysaccharide biosynthesis and mapping of promoters located on the exoHKLAMONP fragment. Mol. Gen. Genet 1993, 241, 367–379. [Google Scholar]
- Becker, A.; Kleickmann, A.; Küster, H.; Keller, M.; Arnold, W.; Pühler, A. Analysis of the Rhizobium meliloti genes exoU, exoV, exoW, exoT and exoI involved in exopolysaccharide biosynthesis and nodule invasion: exoU and exoW probably encode glucosyltransferases. Mol. Plant Microbe Interact 1993, 6, 735–744. [Google Scholar]
- Müller, P.; Keller, M.; Weng, W.M.; Quandt, J.; Arnold, W.; Pühler, A. Genetic analysis of the Rhizobium meliloti exoYFQ operon: ExoY is homologous to sugar transferases and ExoQ represents a transmembrane protein. Mol. Plant Microbe Interact 1993, 6, 55–65. [Google Scholar]
- Becker, A.; Küster, H.; Niehaus, K.; Pühler, A. Extension of the Rhizobium meliloti succinoglycan biosynthesis gene cluster: Identification of the exsA gene encoding an ABC transporter protein, and the exsB gene which probably codes for a regulator of succinoglycan biosynthesis. Mol. Gen. Genet 1995, 249, 487–497. [Google Scholar]
- Becker, A.; Niehaus, K.; Pühler, A. Low-molecular-weight succinoglycan is predominantly produced by Rhizobium meliloti strains carrying a mutated ExoP protein characterized by a periplasmic N-terminal domain and a missing C-terminal domain. Mol. Microbiol 1995, 16, 191–203. [Google Scholar]
- Jofre, E.; Becker, A. Production of succinoglycan polymer in Sinorhizobium meliloti is affected by SMb21506 and requires the N-terminal domain of ExoP. Mol. Plant Microbe Interact 2009, 22, 1656–1668. [Google Scholar]
- Keller, M.; Roxlau, A.; Wenig, W.M.; Schmidt, M.; Quandt, J.; Niehaus, K.; Jording, D.; Arnold, W.; Pühler, A. Molecular analysis of the Rhizobium meliloti mucR gene regulating the biosynthesis of the exopolysaccharides succinoglycan and galactoglucan. Mol. Plant Microbe Interact 1995, 8, 267–277. [Google Scholar]
- Doherty, D.; Leigh, J.A.; Glazebrook, J.; Walker, G.C. Rhizobium meliloti mutants that overproduce the R. meliloti acidic calcofluor-binding exopolysaccharide. J. Bacteriol 1988, 170, 4249–4256. [Google Scholar]
- Reed, J.W.; Glazebrook, J.; Walker, G.C. The exoR gene of Rhizobium meliloti affect RNA levels of other exo genes but lacks homology to known transcriptional regulators. J. Bacteriol 1991, 173, 3789–3794. [Google Scholar]
- Reed, J.W.; Walker, G.C. The exoD gene of Rhizobium meliloti encodes a novel function needed for alfalfa nodule invasion. J. Bacteriol 1991, 173, 664–677. [Google Scholar]
- Uttaro, A.D.; Cangelosi, G.A.; Geremia, R.A.; Nester, E.W.; Ugalde, R.A. Biochemical characterization of avirulent exoC mutants of Agrobacterium tumefaciens. J. Bacteriol 1990, 172, 1640–1646. [Google Scholar]
- Buendia, A.M.; Enenkel, B.; Koplin, R.; Niehaus, K.; Arnold, W.; Pühler, A. The Rhizobium meliloti exoZ/exoB fragment of megaplasmid 2: ExoB functions as an UDP-glucose 4-epimerase and ExoZ shows homology to NodX of Rhizobium leguminosarum biovar viciae strain TOM. Mol. Microbiol 1991, 5, 1519–1530. [Google Scholar]
- York, G.M.; Walker, G.C. The succinyl and acetyl modifications of succinoglycan influence susceptibility of succinoglycan to cleavage by the Rhizobium meliloti glycanases ExoK and ExsH. J. Bacteriol 1998, 180, 4184–4191. [Google Scholar]
- Becker, A.; Rüberg, S.; Küster, H.; Roxlau, A.A.; Keller, M.; Ivashina, T.; Cheng, H.; Walker, G.C.; Pühler, A. The 32-kilobase exp gene cluster of Rhizobium meliloti directing the biosynthesis of galactoglucan: Genetic organization and properties of the encoded gene products. J. Bacteriol 1997, 179, 1375–1384. [Google Scholar]
- Moreira, L.M.; Becker, J.D.; Pühler, A.; Becker, A. The Sinorhizobium meliloti ExpE1 protein secreted by a type I secretion system involving ExpD1 and ExpD2 is required for biosynthesis or secretion of the exopolysaccharide galactoglucan. Microbiology 2000, 136, 2237–2248. [Google Scholar]
- Bahlawane, C.; Baumgarth, B.; Serrania, J.; Rüberg, S.; Becker, A. Fine-tuning of galactoglucan biosynthesis in Sinorhizobium meliloti by differential WggR (ExpG)-, PhoB-, and MucR-dependent regulation of two promoters. J. Bacteriol 2008, 190, 3456–3466. [Google Scholar]
- Canter Cremers, H.C.J.; Batley, M.; Redmond, J.W.; Eysdems, L.; Breedveld, M.W.; Zevenhuizen, L.P.T.M.; Pees, E.; Wijffelman, C.A.; Lugtenberg, B.J.J. Rhizobium leguminosarum exoB mutants are deficient in the synthesis of UDP-glucose 4′-epimerase. J. Biol. Chem 1990, 265, 21122–21127. [Google Scholar]
- Sánchez-Andújar, B.; Coronado, C.; Philip-Hollingsworth, S.; Dazzo, F.B.; Palomares, A.J. Structure and role in symbiosis of the exoB gene of Rhizobium leguminosarum bv trifolii. Mol. Gen. Genet 1997, 255, 131–140. [Google Scholar]
- Laus, M.C.; Logman, T.J.; van Brussel, A.A.N.; Carlson, R.W.; Azadi, P.; Gao, M.; Kijne, J.W. Involvement of exo5 in production of surface polysaccharides in Rhizobium leguminosarum and its role in nodulation of Vicia sativa subsp nigra. J. Bacteriol 2004, 186, 6617–6625. [Google Scholar]
- Muszyński, A.; Laus, M.; Kijne, J.W.; Carlson, R.W. The structures of the lipopolysaccharides from Rhizobium leguminosarum RBL5523 and its UDP-glucose dehydrogenase mutant (exo5). Glycobiology 2011, 21, 55–68. [Google Scholar]
- Borthakur, D.; Barker, R.F.; Latchford, J.W.; Rossen, L.; Johnston, A.W.B. Analysis of pss genes of Rhizobium leguminosarum required for exopolysaccharide synthesis and nodulation of peas: Their primary structure and their interaction with psi and other nodulation genes. Mol. Gen. Genet 1988, 213, 155–162. [Google Scholar]
- Pollock, T.J.; Workum, W.A.; Thorne, L.; Mikolajczak, M.J.; Yamazaki, M.; Kijne, J.W.; Armentrout, R.W. Assignment of biochemical functions to glycosyl transferase genes which are essential for biosynthesis of exopolysaccharides in Sphingomonas strain S88 and Rhizobium leguminosarum. J. Bacteriol 1998, 180, 586–593. [Google Scholar]
- Ivashina, T.V.; Khmelnitsky, M.I.; Shlyapnikov, M.G.; Kanapin, A.A.; Ksenzenko, V.N. The pss4 gene from Rhizobium leguminosarum biovar viciae VF39: Cloning, sequence and the possible role in polysaccharide production and nodule formation. Gene 1994, 50, 111–116. [Google Scholar]
- Janczarek, M.; Skorupska, A. Exopolysaccharide synthesis in R. leguminosarum bv. trifolii is related to various metabolic pathway. Res. Microbiol 2003, 154, 433–442. [Google Scholar]
- Janczarek, M.; Kalita, M.; Skorupska, A. New taxonomic markers for identification of Rhizobium leguminosarum and discrimination between closely related species. Arch. Microbiol 2009, 191, 207–219. [Google Scholar]
- Janczarek, M.; Jaroszuk-Œciseł, J.; Skorupska, A. Multiple copies of rosR and pssA genes enhance exopolysaccharide production, symbiotic competitiveness and clover nodulation in Rhizobium leguminosarum bv trifolii. Antonie Van Leeuwenhoek 2009, 96, 471–486. [Google Scholar]
- Janczarek, M.; Skorupska, A. Regulation of pssA and pssB gene expression in R. leguminosarum bv. trifoliiin response to environmental factors. Antonie Van Leeuwenhoek 2004, 85, 217–227. [Google Scholar]
- Latchford, J.W.; Borthakur, D.; Johnston, A.W.B. The products of Rhizobium genes, psi and pss, which affect exopolysaccharide production, are associated with the bacterial cell surface. Mol. Microbiol 1991, 5, 2107–2114. [Google Scholar]
- Mimmack, M.L.; Borthakur, D.; Jones, M.A.; Downie, J.A.; Johnston, A.W.B. The psi operon of Rhizobium leguminosarum biovar phaseoli: Identification of two genes whose products are located at the bacterial cell surface. Microbiology 1994, 140, 1223–1229. [Google Scholar]
- Mimmack, M.L.; Hong, G.F.; Johnston, A.W.B. Sequence and regulation of psrA, a gene on the Sym plasmid of Rhizobium leguminosarum biovar phaseoli which inhibits transcription of the psi genes. Microbiology 1994, 140, 455–461. [Google Scholar]
- Sadykov, M.R.; Ivashina, T.V.; Kanapin, A.A.; Shlyapnikov, M.G.; Ksenzenko, V.N. Structural and functional organization of the exopolysaccharide biosynthesis genes in Rhizobium leguminosarum bv. viciae VF39. Mol. Biol 1998, 32, 665–671. [Google Scholar]
- Guerreiro, N.; Ksenzenko, V.; Djordjevic, M.A.; Ivashina, T.V.; Rolfe, B. Elevated levels of synthesis of over 20 proteins results after mutation of the Rhizobium leguminosarum exopolysaccharide synthesis gene pssA. J. Bacteriol 2000, 182, 4521–4532. [Google Scholar]
- Janczarek, M.; Król, J.; Kutkowska, J.; Mazur, A.; Wielbo, J.; Borucki, W.; Kopcińska, J.; Łotocka, B.; Urbanik-Sypniewska, T.; Skorupska, A. Mutation in the pssB-pssA intergenic region of Rhizobium leguminosarum bv. trifolii affects the surface polysaccharide synthesis and nitrogen fixation ability. J. Plant Physiol 2001, 158, 1565–1574. [Google Scholar]
- Kutkowska, J.; Turska-Szewczuk, A.; Janczarek, M.; Paduch, R.; Kaminska, T.; Urbanik-Sypniewska, T. Biological activity of (lipo)polysaccharides of the exopolysaccharide-deficient mutant Rt120 derived from Rhizobium leguminosarum bv. trifolii strain TA1. Biochemistry (Mosc. ) 2011, 76, 840–850. [Google Scholar]
- Ivashina, T.V.; Fedorova, E.E.; Ashina, N.P.; Kalinchuk, N.A.; Druzhinina, T.N.; Shashkov, A.S.; Shibaev, V.N.; Ksenzenko, V.N. Mutation in the pssM gene encoding ketal pyruvate transferase leads to disruption of Rhizobium leguminosarum bv. viciae-Pisum sativum symbiosis. J. Appl. Microbiol 2010, 109, 731–742. [Google Scholar]
- Mazur, A.; Król, J.E.; Wielbo, J.; Urbanik-Sypniewska, T.; Skorupska, A. Rhizobium leguminosarum bv. trifolii PssP protein is required for exopolysaccharide biosynthesis and polymerization. Mol. Plant Microbe Interact 2002, 15, 388–397. [Google Scholar]
- Mazur, A.; Król, J.E.; Marczak, M.; Skorupska, A. Membrane topology of PssT, the transmembrane protein component of type I exopolysaccharide transport system in the Rhizobium leguminosarum bv. trifolii strain TA1. J. Bacteriol 2003, 185, 2503–2511. [Google Scholar]
- Marczak, M.; Mazur, A.; Król, J.E.; Gruszecki, W.I.; Skorupska, A. Lipoprotein PssN of Rhizobium leguminosarum bv. trifolii: Subcellular localization and possible involvment in exopolysaccharide export. J. Bacteriol 2006, 188, 6943–6952. [Google Scholar]
- Mazur, A.; Marczak, M.; Król, J.; Skorupska, A. Topological and transcrptional analysis of pssL gene product: A putative Wzx-like exopolysaccharide translocase in Rhizobium leguminosarum bv. trifolii TA1. Arch. Microbiol 2005, 184, 1–10. [Google Scholar]
- Marczak, M.; Mazur, A.; Gruszecki, W.I.; Skorupska, A. PssO, a unique extracellular protein important for exopolysaccharide synthesis in Rhizobium leguminosarum bv trifolii. Biochimie 2008, 90, 1781–1790. [Google Scholar]
- Finnie, C.; Hartley, N.M.; Findlay, K.C.; Downie, J.A. The Rhizobium leguminosarum prsDE genes are required for secretion of several proteins, some of which influence nodulation, symbiotic nitrogen fixation and exopolysaccharide modification. Mol. Microbiol 1997, 25, 135–146. [Google Scholar]
- Finnie, C.; Zorreguieta, A.; Hartley, N.M.; Downie, J.A. Characterization of Rhizobium leguminosarum exopolysaccharide glycanases that are secreted via a type I exporter and have a novel heptapeptide repeat motif. J. Bacteriol 1998, 180, 1691–1699. [Google Scholar]
- Ausmees, N.; Jacobsson, K.; Lindberg, M. A unipolarly located, cell-surface-associated agglutinin, RapA, belongs to a family of Rhizobium-adhering proteins (Rap) in Rhizobium leguminosarum bv trifolii. Microbiology 2001, 147, 549–559. [Google Scholar]
- Krehenbrink, M.; Downie, J.A. Identification of protein secretion systems and novel secreted proteins in Rhizobium leguminosarum bv viciae. BMC Genomics 2008, 9. [Google Scholar] [CrossRef]
- Zorreguieta, A.; Finnie, C.; Downie, J.A. Extracellular glycanases of Rhizobium leguminosarum are activated on the cell surface by an exopolysaccharide-related component. J. Bacteriol 2000, 182, 1304–1312. [Google Scholar]
- Gray, J.X.; Djordjevic, M.A.; Rolfe, B.G. Two genes that regulate exopolysaccharide production in Rhizobium sp. strain NGR234: DNA sequences and resultant phenotypes. J. Bacteriol 1990, 172, 193–203. [Google Scholar]
- Zhan, H.; Leigh, J.A. Two genes that regulate exopolysaccharide production in Rhizobium meliloti. J. Bacteriol 1990, 172, 5254–5259. [Google Scholar]
- Zhan, H.J.; Gray, J.X.; Levery, S.B.; Rolfe, B.G.; Leigh, J.A. Functional and evolutionary relatedness of genes for exopolysaccharide synthesis in Rhizobium meliloti and Rhizobium sp. strain NGR234. J. Bacteriol 1990, 172, 5245–5253. [Google Scholar]
- Staehelin, C.; Forsberg, L.S.; D’Haeze, W.; Gao, M.Y.; Carlson, R.W.; Xie, Z.P.; Pellock, B.J.; Jones, K.M.; Walker, G.C.; Streit, W.R.; et al. Exo-oligosaccharides of Rhizobium sp. strain NGR234 are required for symbiosis with various legumes. J. Bacteriol 2006, 188, 6168–6178. [Google Scholar]
- Schmeisser, C.; Liesegang, H.; Krysciak, D.; Bakkou, N.; Le Quéré, A.; Wollherr, A.; Heinemeyer, I.; Morgenstern, B.; Pommerening-Röser, A.; Flores, M.; et al. Rhizobium sp. strain NGR234 possesses a remarkable number of secretion systems. Appl. Environ. Microbiol 2009, 75, 4035–4045. [Google Scholar]
- Becker, B.; Kosch, K.; Parniske, M.; Müller, P. Exopolysaccharide synthesis in Bradyrhizobium japonicum: Sequence, operon structure and mutational analysis of an exo gene cluster. Mol. Gen. Genet 1998, 259, 161–171. [Google Scholar]
- Wang, P.; Zhong, Z.; Zhou, J.; Cai, T.; Zhu, J. Exopolysaccharide biosynthesis is important for Mesorhizobium tianshanense-plant host interaction. Arch. Microbiol 2008, 189, 525–530. [Google Scholar]
- Bardin, S.D.; Finan, T.M. Regulation of phosphate assimilation in Rhizobium (Sinorhizobium) meliloti. Genetics 1998, 148, 1689–1700. [Google Scholar]
- Mendrygal, K.E.; González, J.E. Environmental regulation of exopolysaccharide production in Sinorhizobium meliloti. J. Bacteriol 2000, 182, 599–606. [Google Scholar]
- Janczarek, M.; Skorupska, A. Modulation of rosR expression and exopolysaccharide production in Rhizobium leguminosarum bv. trifolii by phosphate and clover root exudates. Int. J. Mol. Sci 2011, 12, 4132–4155. [Google Scholar]
- Zevenhuizen, L.P.T.M. Selective synthesis of polysaccharides by Rhizobium trifolii. FEMS Microbiol. Lett 1986, 35, 43–47. [Google Scholar]
- Breedveld, M.W.; Zevenhuizen, L.P.; Canter Cremers, H.C.; Zehnder, A.J. Influence of growth conditions on production of capsular and extracellular polysaccharides by Rhizobium leguminosarum. Antonie Van Leeuwenhoek 1993, 64, 1–8. [Google Scholar]
- Quelas, J.I.; López-García, S.L.; Casabuono, A.; Althabegoiti, M.J.; Mongiardini, E.J.; Pérez-Giménez, J.; Couto, A.; Lodeiro, A.R. Effects of N-starvation and C-source on Bradyrhizobium japonicum exopolysaccharide production and composition, and bacterial infectivity to soybean roots. Arch. Microbiol 2006, 186, 119–128. [Google Scholar]
- Yao, S.Y.; Luo, L.; Har, K.J.; Becker, A.; Rüberg, S.; Yu, G.Q.; Zhu, J.B.; Cheng, H.P. Sinorhizobium meliloti ExoR and ExoS proteins regulate both succinoglycan and flagellum production. J. Bacteriol 2004, 186, 6042–6049. [Google Scholar]
- Rüberg, S.; Pühler, A.; Becker, A. Biosynthesis of the exopolysaccharide galactoglucan in Sinorhizobium meliloti is subject to a complex control by the phosphate-dependent regulator PhoB and the proteins ExpG and MucR. Microbiology 1999, 145, 603–611. [Google Scholar]
- Bertram-Drogatz, P.A.; Quester, I.; Becker, A.; Pühler, A. The Sinorhizobium meliloti MucR protein, which is essential for the production of high-molecular-weight succinoglycan exopolysaccharide, binds to short DNA regions upstream of exoH and exoY. Mol. Gen. Genet 1998, 257, 433–441. [Google Scholar]
- Bertram-Drogatz, P.A.; Rüberg, S.; Becker, A. The regulatory protein MucR binds to a short DNA region located upstream of the mucR coding region in Rhizobium meliloti. Mol. Gen. Genet 1997, 254, 529–538. [Google Scholar]
- Bahlawane, C.; McIntosh, M.; Krol, E.; Becker, A. Sinorhizobium meliloti regulator MucR couples exopolysaccharide synthesis and motility. Mol. Plant Microbe Interact 2008, 21, 1498–1509. [Google Scholar]
- Breedveld, M.W.; Zevenhuizen, L.P.T.M.; Zehnder, A.J.B. Osmotically induced oligo- and polysaccharide synthesis by Rhizobium meliloti SU-47. J. Gen. Microbiol 1990, 136, 2511–2519. [Google Scholar]
- Dusha, I.; Olah, B.; Szegletes, Z.; Erdei, L.; Kondorosi, A. syrM is involved in the determination of the amount and ratio of the two forms of the acidic exopolysaccharide EPS I in Rhizobium meliloti. Mol. Plant Microbe Interact 1999, 12, 755–765. [Google Scholar]
- Reed, J.W.; Capage, M.; Walker, G.C. Rhizobium meliloti exoG and exoJ mutations affect the ExoX-ExoY system for modulation of exopolysaccharide production. J. Bacteriol 1991, 173, 3776–3788. [Google Scholar]
- Osterås, M.; Stanley, J.; Finan, T.M. Identification of Rhizobium-specific intergenic mosaic elements within an essential two-component regulatory system of Rhizobium species. J. Bacteriol 1995, 177, 5485–5494. [Google Scholar]
- Cheng, H.P.; Walker, G.C. Succinoglycan production by Rhizobium meliloti is regulated through the ExoS-ChvI two-component regulatory system. J. Bacteriol 1998, 180, 20–26. [Google Scholar]
- Wells, D.H.; Chen, E.J.; Fisher, R.F.; Long, S.R. ExoR is genetically coupled to the ExoS-ChvI two-component system and located in the periplasm of Sinorhizobium meliloti. Mol. Microbiol 2007, 64, 647–664. [Google Scholar]
- Chen, E.J.; Sabio, E.A.; Long, S.R. The periplasmic regulator ExoR inhibits ExoS/ChvI two-component signalling in Sinorhizobium meliloti. Mol. Microbiol 2008, 69, 1290–1303. [Google Scholar]
- Lu, H.Y.; Cheng, H.P. Autoregulation of Sinorhizobium meliloti exoR gene expression. Microbiology 2010, 156, 2092–2101. [Google Scholar]
- Morris, J.; González, J.E. The novel genes emmABC are associated with exopolysaccharide production, motility, stress adaptation, and symbiosis in Sinorhizobium meliloti. J. Bacteriol 2009, 191, 5890–5900. [Google Scholar]
- Bélanger, L.; Dimmick, K.A.; Fleming, J.S.; Charles, T.C. Null mutations in Sinorhizobium meliloti exoS and chvI demonstrate the importance of this two-component regulatory system for symbiosis. Mol. Microbiol 2009, 74, 1223–1237. [Google Scholar]
- Wang, C.; Kemp, J.; Da Fonseca, I.O.; Equi, R.C.; Sheng, X.; Charles, T.C.; Sobral, B.W.S. Sinorhizobium meliloti 1021 loss-of-function deletion mutation in chvI and its phenotypic characteristics. Mo. Plant Microbe Interact 2010, 23, 153–160. [Google Scholar]
- Barnett, M.J.; Long, S.R. Identification and characterization of a gene on Rhizobium meliloti pSymA, syrB, that negatively affects syrM expression. Mol. Plant Microbe Interact 1997, 10, 550–559. [Google Scholar]
- Barnett, M.J.; Long, S.R. DNA sequence and translational product of a new nodulation-regulatory locus: syrM has sequence similarity to NodD proteins. J. Bacteriol 1990, 172, 3695–3700. [Google Scholar]
- Gibson, K.E.; Barnett, M.J.; Toman, C.J.; Long, S.R.; Walker, G.C. The symbiosis regulator CbrA modulates a complex regulatory network affecting the flagellar apparatus and cell envelope proteins. J. Bacteriol 2007, 189, 3591–3602. [Google Scholar]
- Pinedo, C.A.; Bringhurst, R.M.; Gage, D.J. Sinorhizobium meliloti mutants lacking phosphotransferase system enzyme HPr or EIIA are altered in diverse processes, including carbon metabolism, cobalt requirements and succinoglycan production. J. Bacteriol 2008, 190, 2947–2956. [Google Scholar]
- Pinedo, C.A.; Gage, D.J. HPrK regulates succinate-mediated catabolite repression in the gram-negative symbiont Sinorhizobium meliloti. J. Bacteriol 2009, 191, 298–309. [Google Scholar]
- Rinaudi, L.V.; González, J.E. The low-molecular-weight fraction of the exopolysaccharide II from Sinorhizobium meliloti is a crucial determinant of biofilm formation. J. Bacteriol 2009, 191, 7216–7224. [Google Scholar]
- Summers, M.L.; Elkins, J.G.; Elliot, B.A.; McDermott, T.R. Expression and regulation of phosphate stress inducible genes in Sinorhizobium meliloti. Mol. Plant Microbe Interact 1999, 11, 1094–1101. [Google Scholar]
- Krol, E.; Becker, A. Global transcriptional analysis of the phosphate starvation response in Sinorhizobium meliloti strains 1021 and 2011. Mol. Gen. Genomics 2004, 272, 1–17. [Google Scholar]
- Yuan, Z.; Zaheer, R.; Morton, R.; Finan, T.M. Genome prediction of PhoB regulated promoters in Sinorhizobium meliloti and twelve proteobacteria. Nucleic Acids Res 2006, 34, 2686–2697. [Google Scholar]
- Quester, I.; Becker, A. Four promoters subject to regulation by ExoR and PhoB direct transcription of the Sinorhizobium meliloti exoYFQ operon involved in the biosynthesis of succinoglycan. J. Mol. Microbiol. Biotechnol 2004, 7, 115–132. [Google Scholar]
- Baumgarth, B.; Bartels, F.W.; Anselmetti, D.; Becker, A.; Ros, R. Detailed studies of the binding mechanism of the Sinorhizobium meliloti transcriptional activator ExpG to DNA. Microbiology 2005, 151, 1–10. [Google Scholar]
- Marketon, M.M.; Glenn, S.A.; Eberhard, A.; González, J.E. Quorum sensing controls exopolysaccharide production in Sinorhizobium meliloti. J. Bacteriol 2003, 185, 325–331. [Google Scholar]
- González, J.E.; Marketon, M.M. Quorum sensing in nitrogen-fixing rhizobia. Microbiol. Mol. Biol. Rev 2003, 67, 574–592. [Google Scholar]
- Hoang, H.H.; Becker, A.; González, J.E. The LuxR homolog ExpR, in combination with the Sin quorum sensing system, plays a central role in Sinorhizobium meliloti gene expression. J. Bacteriol 2004, 186, 5460–5472. [Google Scholar]
- McIntosh, M.; Krol, E.; Becker, A. Competitive and cooperative effects in quorum-sensing-regulated galactoglucan biosynthesis in Sinorhizobium meliloti. J. Bacteriol 2008, 190, 5308–5317. [Google Scholar]
- Gurich, N.; González, J.E. Role of quorum sensing in Sinorhizobium meliloti-alfalfa symbiosis. J. Bacteriol 2009, 191, 4372–4382. [Google Scholar]
- Mueller, K.; González, J.E. Complex regulation of symbiotic functions is coordinated by MucR and quorum sensing in Sinorhizobium meliloti. J. Bacteriol 2011, 193, 485–496. [Google Scholar]
- Pellock, B.J.; Teplicki, M.; Boinay, R.P.; Bauer, W.D.; Walker, G.C. A LuxR homolog controls production of symbiotically active extracellular polysaccharide II by Sinorhizobium meliloti. J. Bacteriol 2002, 184, 5067–5076. [Google Scholar]
- Glenn, S.A.; Gurich, N.; Feeney, M.A.; González, J.E. The ExpR/Sin quorum-sensing system controls succinoglycan production in Sinorhizobium meliloti. J. Bacteriol 2007, 189, 7077–7088. [Google Scholar]
- Hoang, H.H.; Gurich, N.; González, J.E. Regulation of motility by the ExpR/Sin quorum-sensing system in Sinorhizobium meliloti. J. Bacteriol 2008, 190, 861–871. [Google Scholar]
- Sourjik, V.; Muschler, P.; Scharf, B.; Schmitt, R. VisN and VisR are global regulators of chemotaxis, flagellar, and motility genes in Sinorhizobium (Rhizobium) meliloti. J. Bacteriol 2000, 182, 782–788. [Google Scholar]
- Galibert, F.; Finan, T.M.; Long, S.R.; Pühler, A.; Abola, P.; Ampe, F.; Barloy-Hubler, F.; Barnett, M.J.; Becker, A.; Boistard, P.; et al. The composite genome of the legume symbiont Sinorhizobium meliloti. Science 2001, 293, 668–672. [Google Scholar]
- Tambalo, D.D.; Del Bel, K.L.; Bustard, D.E.; Greenwood, P.R.; Steedman, A.E.; Hynes, M.F. Regulation of flagellar, motility and chemotaxis genes in Rhizobium leguminosarum by the VisN/R-Rem cascade. Microbiology 2010, 156, 1673–1685. [Google Scholar]
- Tambalo, D.D.; Yost, C.K.; Hynes, M.F. Characterization of swarming motility in Rhizobium leguminosarum bv viciae. FEMS Microbiol. Lett 2010, 307, 165–174. [Google Scholar]
- Borthakur, D.; Johnston, A.W.B. Sequence of psi, a gene of the symbiotic plasmid of Rhizobium phaseoli which inhibits exopolysaccharide synthesis and nodulation and demonstration that its transcription is inhibited by psr, another gene on the symbiotic plasmid. Mol. Gen. Genet 1987, 207, 149–154. [Google Scholar]
- Reeve, W.G.; Dilworth, M.J.; Tiwari, R.P.; Glenn, A.R. Regulation of exopolysaccharide production in Rhizobium leguminosarum biovar viciae WSM710 involves exoR. Microbiology 1997, 143, 1951–1958. [Google Scholar]
- Janczarek, M.; Król, J.; Skorupska, A. The pssB gene product of Rhizobium leguminosarum bv. trifolii is homologous to a family of inositol monophosphatases. FEMS Microbiol. Lett 1999, 173, 319–325. [Google Scholar]
- Janczarek, M.; Skorupska, A. The Rhizobium leguminosarum bv. trifolii RosR: Transcriptional regulator involved in exopolysaccharide production. Mol. Plant Microbe Interact 2007, 20, 867–881. [Google Scholar]
- Edwards, A.; Frederix, M.; Wisniewski-Dyé, F.; Jones, J.; Zorreguieta, A.; Downie, J.A. The cin and rai quorum-sensing regulatory systems in Rhizobium leguminosarum are coordinated by ExpR and CinS, a small regulatory protein coexpressed with CinI. J. Bacteriol 2009, 191, 3059–3067. [Google Scholar]
- Ksenzenko, V.N.; Ivashina, T.V.; Dubeĭkovskaia, Z.A.; Ivanov, S.G.; Nanazashvili, M.B.; Druzhinina, T.N.; Kalinchuk, N.A.; Shibaev, V.N. The pssA gene encodes UDP-glucose: Polyprenyl phosphate-glucosyl phosphotransferase initiating biosynthesis of Rhizobium leguminosarum exopolysaccharide. Russ. J. Bioorg. Chem 2007, 33, 150–155. [Google Scholar]
- Ivashina, T.V.; Sadykov, M.R.; Chatuev, B.M.; Kanapin, A.A.; Shliapnikov, M.G.; Ksenzenko, V.N. The Rhizobium leguminosarum bv. viciae VF39 pssB gene product participates in symbiotic fixation of molecular nitrogen. Dokl. Akad. Nauk 1996, 350, 712–715. [Google Scholar]
- Kutkowska, J.; Janczarek, M.; Kopcińska, J.; Urbanik-Sypniewska, T.; Skorupska, A. Effects of pssB mutation on surface polysaccharides and symbiotic phenotype of Rhizobium leguminosarum bv trifolii. Acta Biol. Crac. Ser. Bot 2007, 49, 81–89. [Google Scholar]
- Janczarek, M.; Skorupska, A. The Rhizobium leguminosarum bv. trifolii pssB gene product is an inositol monophosphatase that influences exopolysaccharide synthesis. Arch. Microbiol 2001, 175, 143–151. [Google Scholar]
- Bittinger, M.A.; Milner, J.L.; Saville, B.J.; Handelsman, J. rosR, a determinant of nodulation competitiveness in Rhizobium etli. Mol. Plant Microbe Interact 1997, 10, 180–186. [Google Scholar]
- Chou, A.Y.; Archdeacon, J.; Kado, C.I. Agrobacterium transcriptional regulator Ros is a prokaryotic zinc finger protein that regulates the plant oncogene ipt. Proc. Natl. Acad. Sci. USA 1998, 95, 5293–5298. [Google Scholar]
- Hussain, H.; Johnston, A.W. Iron-dependent transcription of the regulatory gene ros of Agrobacterium radiobacter. Mol. Plant Microbe Interact 1997, 10, 1087–1093. [Google Scholar]
- Bittinger, M.A.; Handelsman, J. Identification of genes in the RosR regulon of Rhizobium etli. J. Bacteriol 2000, 182, 1706–1713. [Google Scholar]
- Janczarek, M.; Kutkowska, J.; Piersiak, T.; Skorupska, A. Rhizobium leguminosarum bv. trifolii rosR is required for interaction with clover, biofilm formation and adaptation to the environment. BMC Microbiol 2010, 10. [Google Scholar] [CrossRef]
- Janczarek, M.; Skorupska, A. Rhizobium leguminosarum bv. trifolii rosR gene expression is regulated by catabolic repression. FEMS Microbiol. Lett 2009, 291, 112–119. [Google Scholar]
- Wielbo, J.; Skorupska, A. Influence of phosphate and ammonia on the growth, exopolysaccharide production and symbiosis of Rhizobium leguminosarum bv. trifolii TA1 with clover (Trifolium pratense). Acta Biol. Hung 2008, 59, 115–127. [Google Scholar]
- Janczarek, M. Personal communication, University of M. Curie-Skłodowska: Lublin, Poland, 2011.
- Wielbo, J.; Mazur, A.; Krol, J.E.; Marczak, M.; Skorupska, A. Environmental modulation of the pssTNOP gene expression in Rhizobium leguminosarum bv trifolii. Can. J. Microbiol 2004, 50, 1–11. [Google Scholar]
- Vinardell, J.M.; Ollero, F.J.; Hidalgo, A.; López-Baena, F.J.; Medina, C.; Ivanov-Vangelov, K.; Parada, M.; Madinabeitia, N.; Espuny Mdel, R.; Bellogín, R.A.; et al. NolR regulates diverse symbiotic signals of Sinorhizobium fredii HH103. Mol. Plant Microbe Interact 2004, 17, 676–685. [Google Scholar]
- Chen, H.; Gao, K.; Kondorosi, E.; Kondorosi, A.; Rolfe, B.G. Functional genomic analysis of global regulator NolR in Sinorhizobium meliloti. Mol. Plant Microbe Interact 2005, 18, 1340–1352. [Google Scholar]
- Wisniewski-Dyé, F.; Downie, J.A. Quorum sensing in Rhizobium. Antonie Van Leeuwenhoek 2002, 81, 397–407. [Google Scholar]
- Frederix, M.; Edwards, A.; McAnulla, C.; Downie, J.A. Co-ordination of quorum-sensing regulation in Rhizobium leguminosarum by induction of an anti-repressor. Mol. Microbiol 2011, 81, 994–1007. [Google Scholar]
- Russo, D.M.; Williams, A.; Edwards, A.; Posadas, D.M.; Finnie, C.; Dankert, M.; Downie, J.A.; Zorreguieta, A. Proteins exported via the PrsD-PrsE type I secretion system and the acidic exopolysaccharide are involved in biofilm formation by Rhizobium leguminosarum. J. Bacteriol 2006, 188, 4474–4486. [Google Scholar]
- Fujishige, N.A.; Kapadia, N.N.; De Hoff, P.L.; Hirsch, A.M. Investigations of Rhizobium biofilm formation. FEMS Microbiol. Ecol 2006, 56, 195–206. [Google Scholar]
- Rinaudi, L.V.; Fujishige, N.A.; Hirsch, A.M.; Banchio, E.; Zorreguieta, A.; Giordano, W. Effects of nutritional and environmental conditions on Sinorhizobium meliloti biofilm formation. Res. Microbiol 2006, 157, 867–875. [Google Scholar]
- Fujishige, N.A.; Lum, M.R.; De Hoff, P.L.; Whitelegge, J.P.; Faull, K.F.; Hirsch, A.M. Rhizobium common nod genes are required for biofilm formation. Mol. Microbiol 2008, 67, 504–515. [Google Scholar]
- Williams, A.; Wilkinson, A.; Krehenbrink, M.; Russo, D.; Zorreguieta, A.; Downie, J.A. Glucomannan-mediated attachment of Rhizobium leguminosarum to pea root hairs is required for competitive nodule infection. J. Bacteriol 2008, 190, 4706–4715. [Google Scholar]

© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Janczarek, M. Environmental Signals and Regulatory Pathways That Influence Exopolysaccharide Production in Rhizobia. Int. J. Mol. Sci. 2011, 12, 7898-7933. https://doi.org/10.3390/ijms12117898
Janczarek M. Environmental Signals and Regulatory Pathways That Influence Exopolysaccharide Production in Rhizobia. International Journal of Molecular Sciences. 2011; 12(11):7898-7933. https://doi.org/10.3390/ijms12117898
Chicago/Turabian StyleJanczarek, Monika. 2011. "Environmental Signals and Regulatory Pathways That Influence Exopolysaccharide Production in Rhizobia" International Journal of Molecular Sciences 12, no. 11: 7898-7933. https://doi.org/10.3390/ijms12117898