Behavioral Impairment and Oxidative Damage Induced by Chronic Application of Nonylphenol

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of NP on the Behavior of Mice
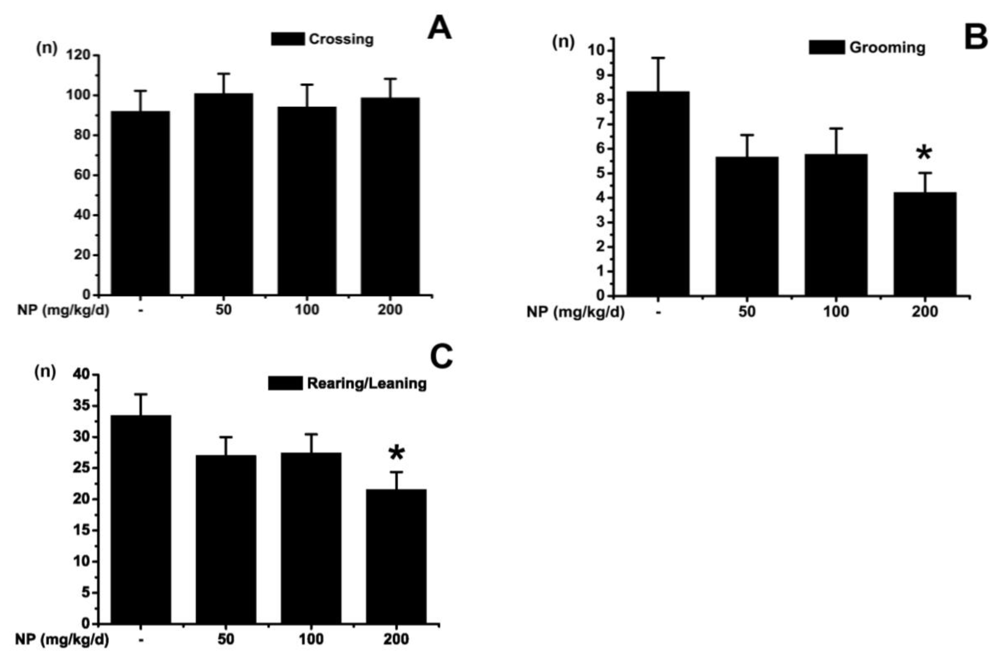
2.1.1. Open Field Test
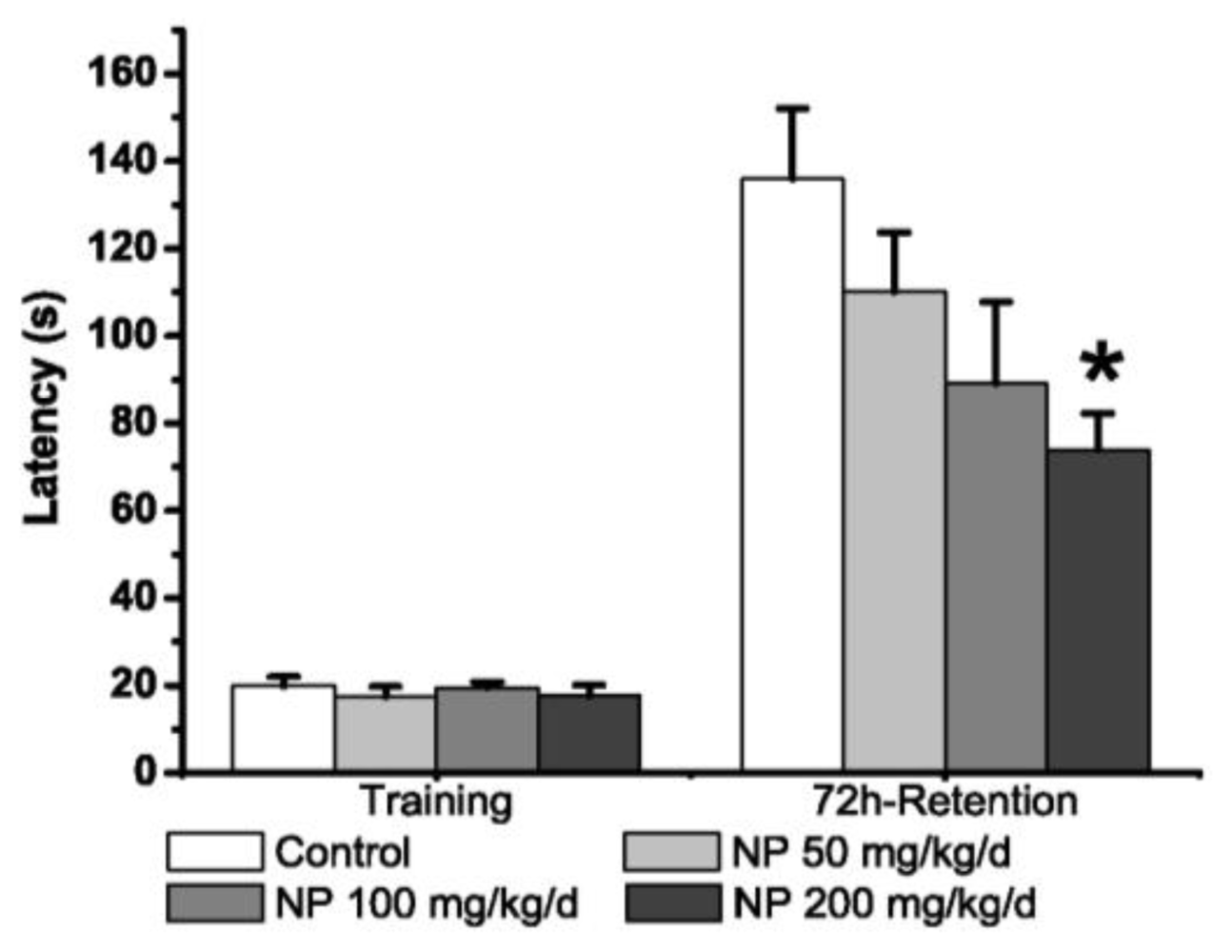
2.1.2. Step-Through Test
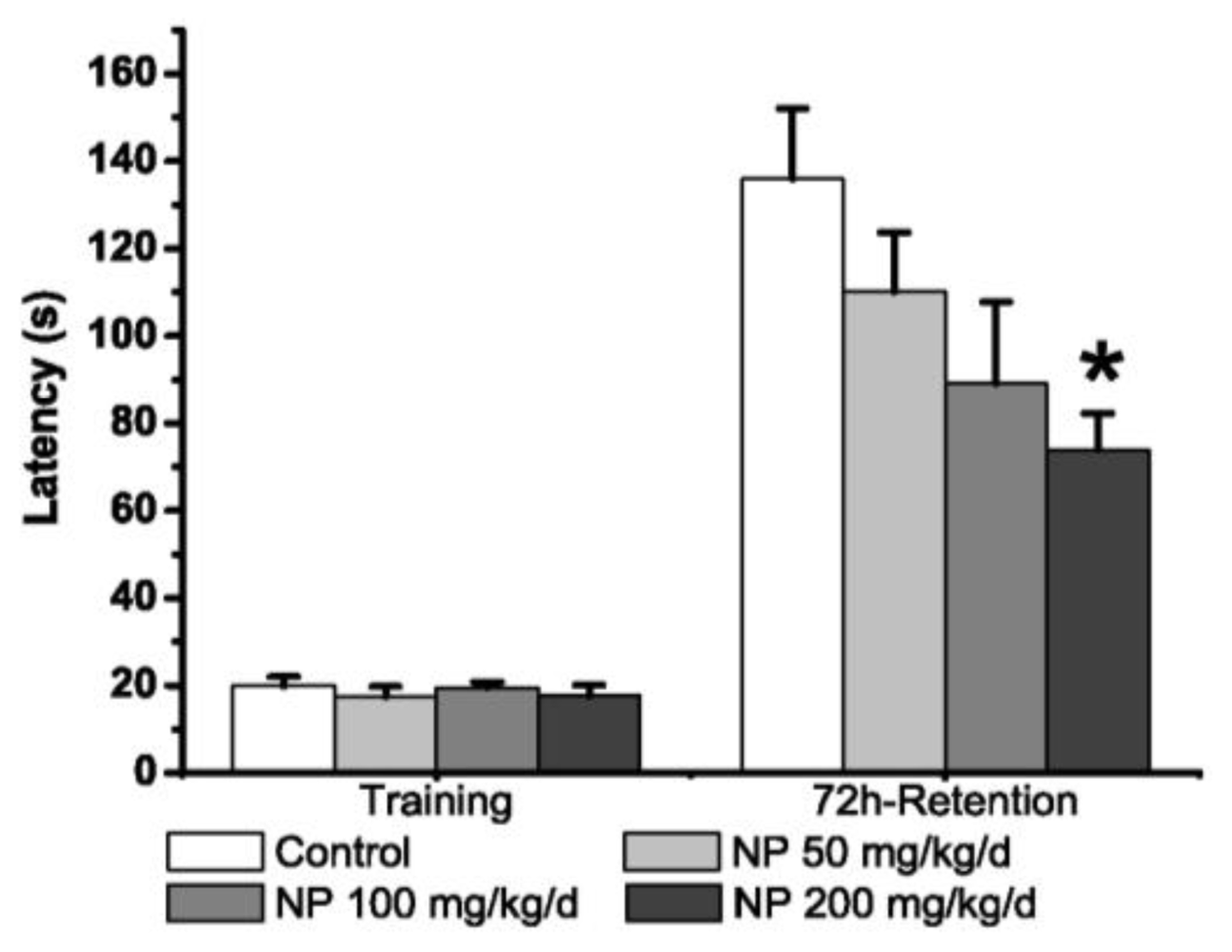
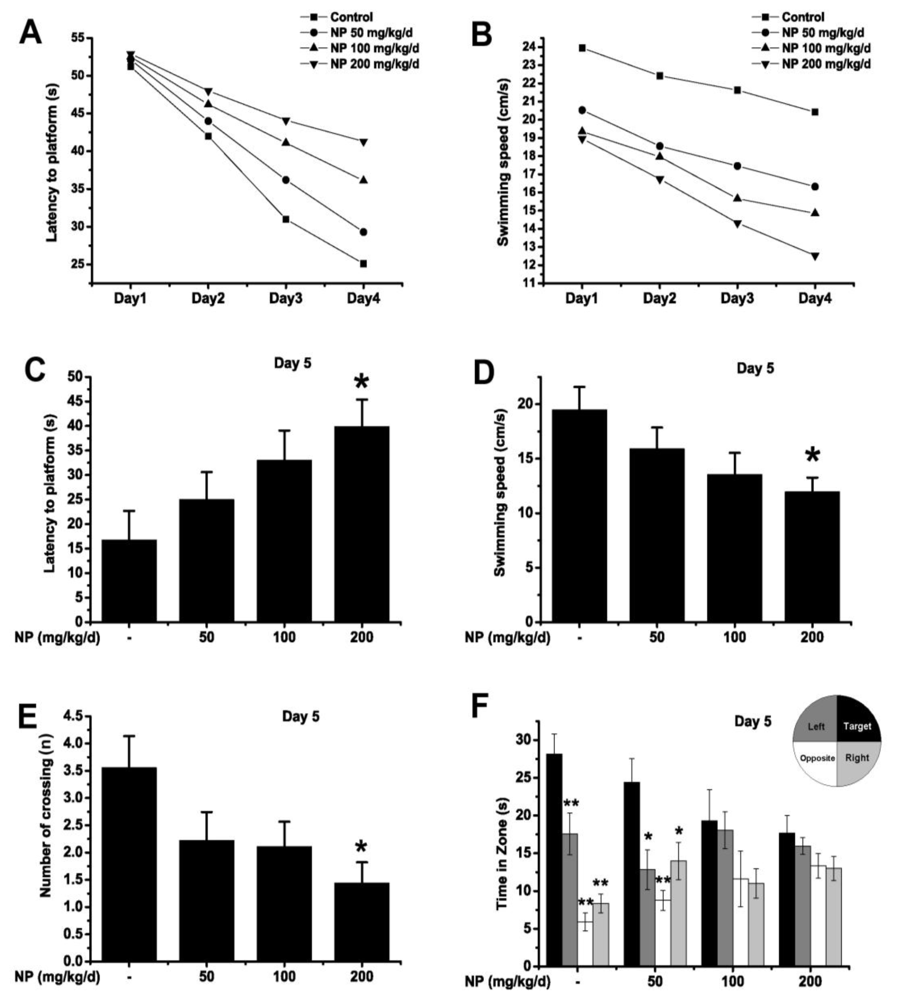
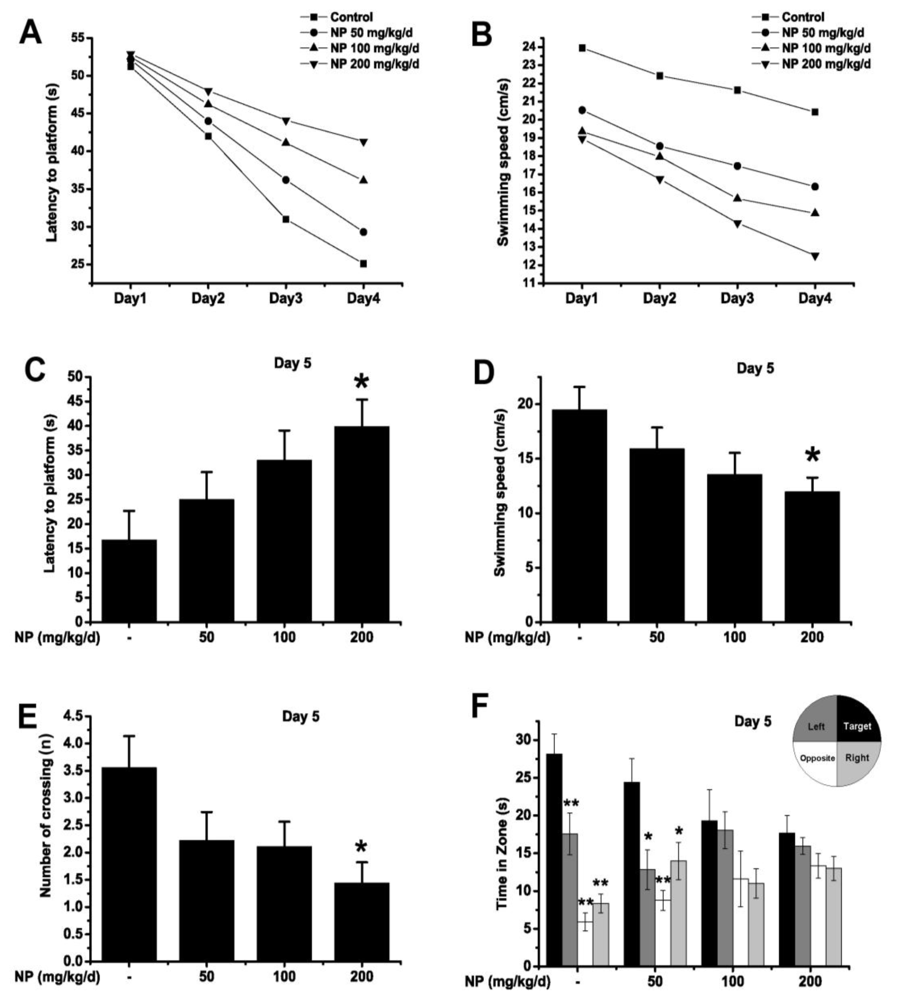
2.1.3. Morris Water Maze Test
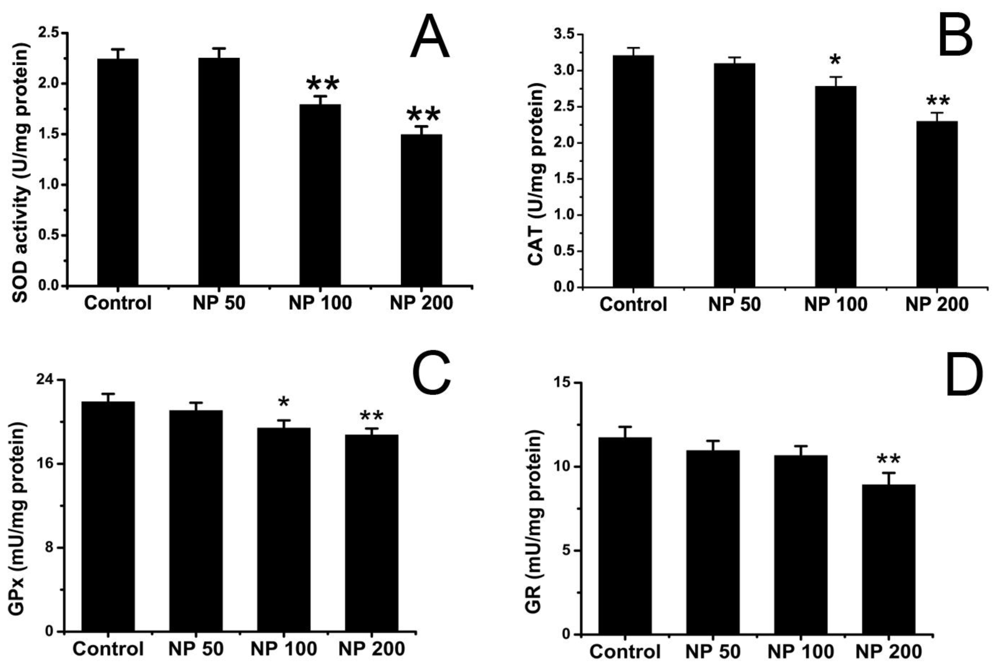
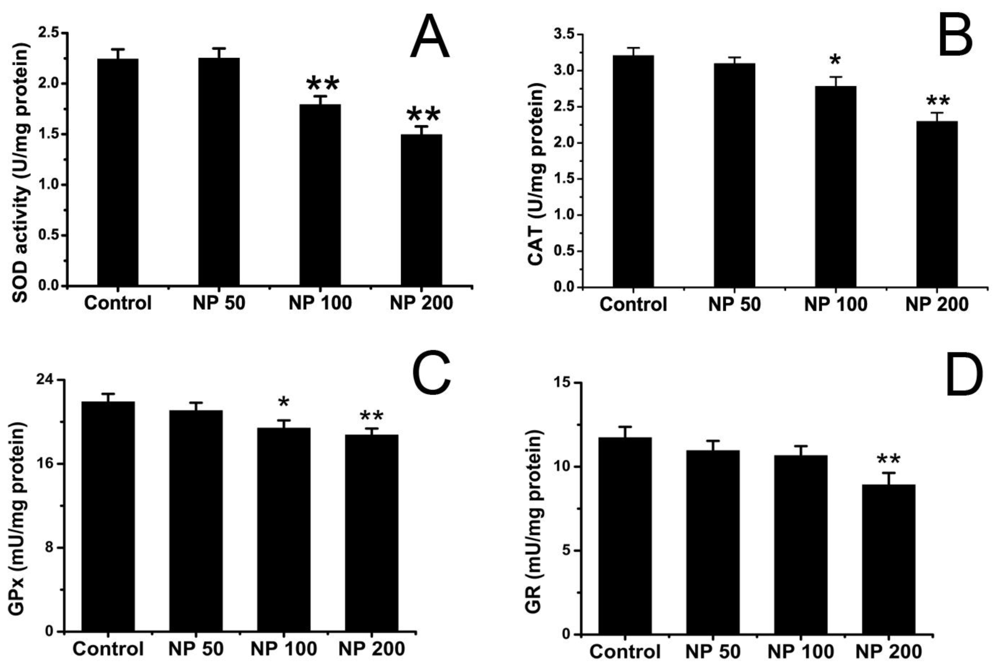
2.2. Effects of NP on Antioxidative Status of Mouse Brain
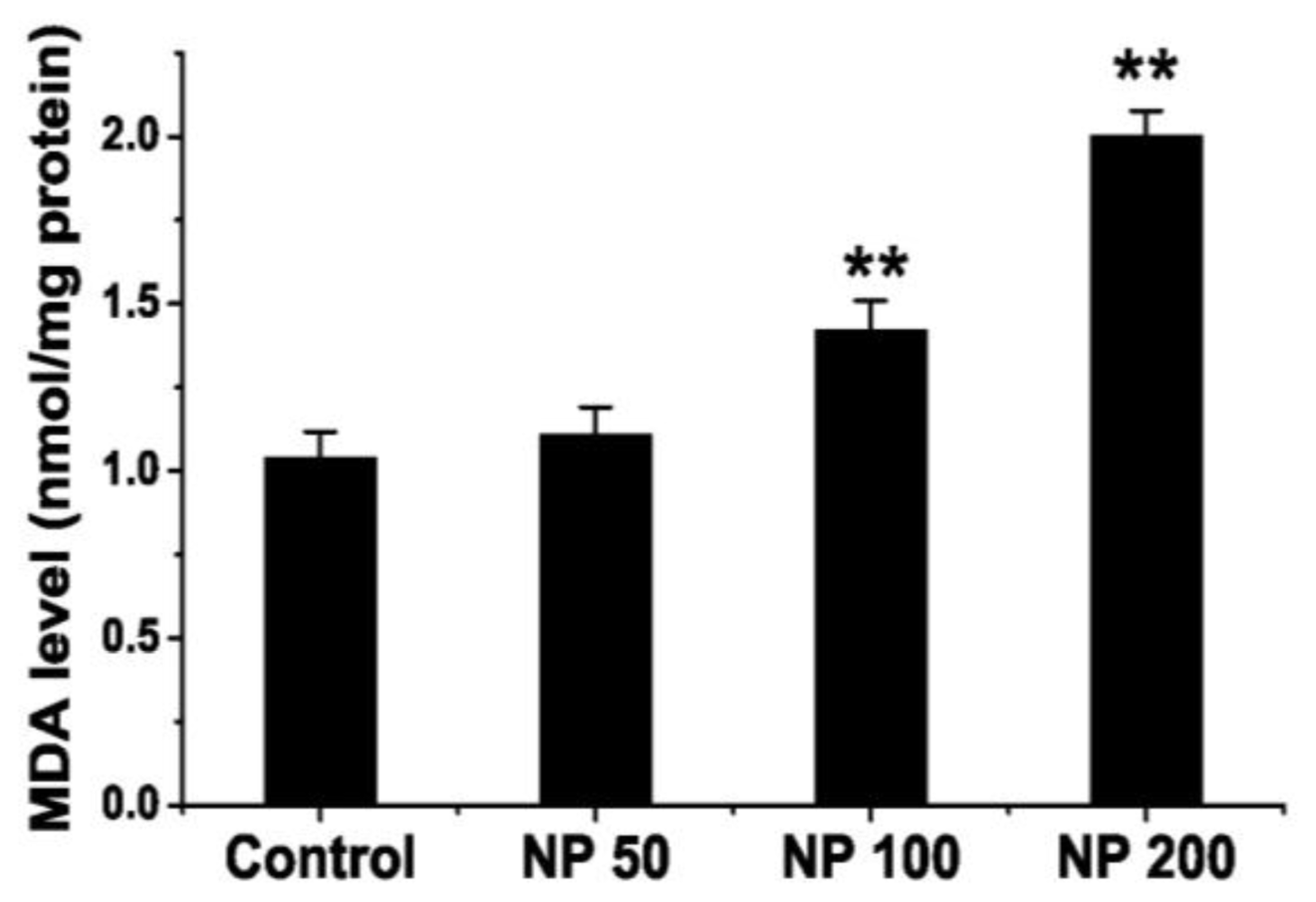
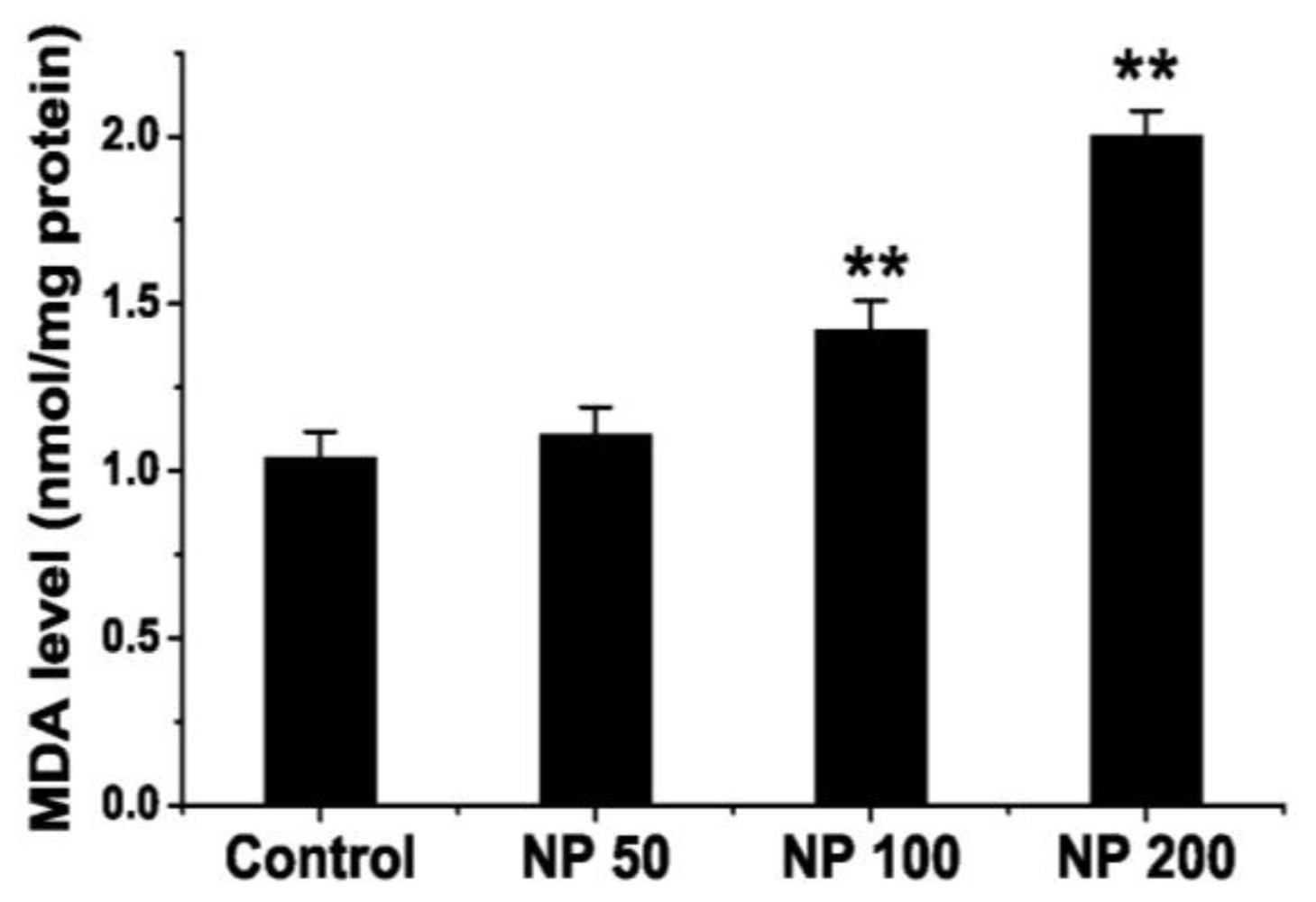
2.3. Effects of NP on Lipid Peroxidation Level of Mouse Brain
2.4. Discussion
3. Experimental Section
3.1. Animals and Treatments
3.2. Behavioral Tests
3.2.1. Open Field Test
3.2.2. Step-Through Test
3.2.3. Morris Water Maze Test
3.3. Biochemical Studies
3.3.1. Tissue Homogenate
3.3.2. Assay of SOD Activity
3.3.3. Analysis of CAT Activity
3.3.4. Assay of GPx Activity
3.3.5. Analysis of GR Activity
3.3.6. Measurement of Malondialdehyde (MDA) Levels
3.4. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Mathieu, A; Bonvallot, N; Belzunces, LP; Beausoleil, C; Guerbet, M; Parent-Massin, D; Rousselle, C. Elaboration of a toxicological reference value (TRV) for the reprotoxic effects of nonylphenol. Toxicol. Lett 2009, 189, S242. [Google Scholar]
- Wu, JJ; Wang, KL; Wang, SW; Hwang, GS; Mao, IF; Chen, ML; Wang, PS. Differential effects of nonylphenol on testosterone secretion in rat Leydig cells. Toxicology 2010, 268, 1–7. [Google Scholar]
- El-Dakdoky, MH; Helal, MAM. Reproductive toxicity of male mice after exposure to nonylphenol. Bulletin Environ. Contam. Toxicol 2007, 79, 188–191. [Google Scholar]
- Jie, X; Yang, W; Jie, Y; Fan, QY; Liu, XY; Yan, L; Huang, HJ. Immune Effects of Nonylphenol on Offspring of Rats Exposed During Pregnancy. Hum. Ecol. Risk Assess 2010, 16, 444–452. [Google Scholar]
- Hebert, N; Gagne, F; Cyr, D; Pellerin, J; Blaise, C; Fournier, M. Effects of 4-Nonylphenol on the Immune System of Rainbow Trout (Oncorhynchus mykiss). Fresenius Environ Bulletin 2009, 18, 757–761. [Google Scholar]
- Korkmaz, A; Ahbab, MA; Kolankaya, D; Barlas, N. Influence of vitamin C on bisphenol A, nonylphenol and octylphenol induced oxidative damages in liver of male rats. Food Chem. Toxicol 2010, 48, 2865–2871. [Google Scholar]
- Mao, Z; Zheng, YL; Zhang, YQ; Han, BP; Chen, LT; Li, J; Li, F; Shan, Q. Chronic application of nonylphenol-induced apoptosis via suppression of bcl-2 transcription and up-regulation of active caspase-3 in mouse brain. Neurosci. Lett 2008, 439, 147–152. [Google Scholar]
- Zhang, YQ; Mao, Z; Zheng, YL; Han, BP; Chen, LT; Li, J; Li, F. Elevation of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Expression in the Mouse Brain after Chronic Nonylphenol Exposure. Int. J. Mol. Sci 2008, 9, 1977–1988. [Google Scholar]
- Flynn, KM; Newbold, RR; Ferguson, SA. Multigenerational exposure to dietary nonylphenol has no severe effects on spatial learning in female rats. Neurotoxicology 2002, 23, 87–94. [Google Scholar]
- Negishi, T; Kawasaki, K; Suzaki, S; Maeda, H; Ishii, Y; Kyuwa, S; Kuroda, Y; Yoshikawa, Y. Behavioral alterations in response to fear-provoking stimuli and tranylcypromine induced by perinatal exposure to bisphenol a and nonylphenol in male rats. Environ. Health Perspect 2004, 112, 1159–1164. [Google Scholar]
- Nagao, T; Wada, K; Marumo, H; Yoshimura, S; Ono, H. Reproductive effects of nonylphenol in rats after gavage administration: a two-generation study. Reprod. Toxicol 2001, 15, 293–315. [Google Scholar]
- Yu, YP; Ju, WP; Li, ZG; Wang, DZ; Wang, YC; Xie, AM. Acupuncture inhibits oxidative stress and rotational behavior in 6-hydroxydopamine lesioned rat. Brain Res 2010, 1336, 58–65. [Google Scholar]
- Qureshi, GA; Baig, S; Sarwar, M; Parvez, SH. Neurotoxicity, oxidative stress and cerebrovascular disorders. Neurotoxicology 2004, 25, 121–138. [Google Scholar]
- Sharma, DR; Sunkaria, A; Bal, A; Bhutia, YD; Vijayaraghavan, R; Flora, SJS; Gill, KD. Neurobehavioral impairments, generation of oxidative stress and release of proapoptotic factors after chronic exposure to sulphur mustard in mouse brain. Toxicol. Appl. Pharmacol 2009, 240, 208–218. [Google Scholar]
- Okai, Y; Sato, EF; Higashi-Okai, K; Inoue, M. Enhancing effect of the endocrine disruptor para-nonylphenol on the generation of reactive oxygen species in human blood neutrophils. Environ. Health Perspect 2004, 112, 553–556. [Google Scholar]
- Gong, Y; Han, XD. Nonylphenol-induced oxidative stress and cytotoxicity in testicular Sertoli cells. Reprod. Toxicol 2006, 22, 623–630. [Google Scholar]
- Okai, Y; Sato, EF; Higashi-Okai, K; Inoue, M. Effect of endocrine disruptor para-nonylphenol on the cell growth and oxygen radical generation in Escherichia coli mutant cells deficient in catalase and superoxide dismutase. Free Radical Biol. Med 2004, 37, 1412–1418. [Google Scholar]
- Aydogan, M; Korkmaz, A; Barlas, N; Kolankaya, D. The role of vitamin C on endocrine disrupters bisphenol A, nonylphenol and octylphenol induced brain damages of male rats. Toxicol. Lett 2008, 180, S179. [Google Scholar]
- Walsh, RN; Cummins, RA. Open-Field Test—Critical-Review. Psychol Bulletin 1976, 83, 482–504. [Google Scholar]
- Schantz, SL; Widholm, JJ. Cognitive effects of endocrine-disrupting chemicals in animals. Environ. Health Perspect 2001, 109, 1197–1206. [Google Scholar]
- Poimenova, A; Markaki, E; Rahiotis, C; Kitraki, E. Corticosterone-Regulated Actions in the Rat Brain Are Affected by Perinatal Exposure to Low Dose of Bisphenol A. Neuroscience 2010, 167, 741–749. [Google Scholar]
- Palanza, P; Gioiosa, L; Vom Saal, FS; Parmigiani, S. Effects of developmental exposure to bisphenol A on brain and behavior in mice. Environ. Res 2008, 108, 150–157. [Google Scholar]
- Zalko, D; Costagliola, R; Dorio, C; Rathahao, E; Cravedi, J. In Vivo Metabolic Fate of the Xeno-Estrogen 4-n-Nonylphenol in Wistar Rats. Drug Metab. Dispos 2003, 31, 168–178. [Google Scholar]
- Witorsch, RJ. Low-dose in utero effects of xenoestrogens in mice and their relevance to humans: an analytical review of the literature. Food Chem. Toxicol 2002, 40, 905–912. [Google Scholar]
- Sharma, M; Gupta, YK. Chronic treatment with trans rosveratrol prevents intracerebroventricular streptozotocin induced cognitive impairment and oxidative stress in rats. Life Sci 2002, 71, 2489–2498. [Google Scholar]
- Obata, T; Kubota, S. Formation of hydroxy radicals by environmental estrogen-like chemicals in rat striatum. Neurosci. Lett 2000, 296, 41–44. [Google Scholar]
- Glade, MJ. Oxidative stress and cognitive longevity. Nutrition 2010, 26, 595. [Google Scholar]
- Davies, KJA. Oxidative stress, antioxidant defenses, and damage removal, repair, and replacement systems. IUBMB Life 2001, 50, 279. [Google Scholar]
- Franco, R; Sanchez-Olea, R; Reyes-Reyes, EM; Panayiotidis, MI. Environmental toxicity, oxidative stress and apoptosis: Menage a Trois. Mutat. Res 2009, 674, 3–22. [Google Scholar]
- Takahashi, A; Masuda, A; Sun, M; Centonze, VE; Herman, B. Oxidative stress-induced apoptosis is associated with alterations in mitochondrial caspase activity and Bcl-2-dependent alterations in mitochondrial pH (pHm). Brain Res Bulletin 2004, 62, 497–504. [Google Scholar]
- Lu, J; Zheng, YL; Luo, L; Wu, DM; Sun, DX; Feng, YJ. Quercetin reverses D-galactose induced neurotoxicity in mouse brain. Behav. Brain Res 2006, 171, 251–260. [Google Scholar]
- Flohe, L; Otting, F. Superoxide dismutase assays. Methods Enzymol 1984, 105, 93–104. [Google Scholar]
- Claiborne, A. CRC Handbook of Methods for Oxygen Radical Research; CRC Press: Boca Raton, FL, USA, 1985. [Google Scholar]
- Mohandas, J; Marshall, JJ; Duggin, GG; Horvath, JS; Tiller, DJ. Low Activities of Glutathione-Related Enzymes As Factors in the Genesis of Urinary-Bladder Cancer. Cancer Res 1984, 44, 5086–5091. [Google Scholar]
- Mizuno, Y; Ohta, K. Regional Distributions of Thiobarbituric Acid-Reactive Products, Activities of Enzymes Regulating the Metabolism of Oxygen Free-Radicals, and Some of the Related Enzymes in Adult and Aged Rat Brains. J. Neurochem 1986, 46, 1344–1352. [Google Scholar]
- Okhawa, H; Ohishi, N; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem 1979, 95, 351–358. [Google Scholar]
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mao, Z.; Zheng, Y.-L.; Zhang, Y.-Q. Behavioral Impairment and Oxidative Damage Induced by Chronic Application of Nonylphenol. Int. J. Mol. Sci. 2011, 12, 114-127. https://doi.org/10.3390/ijms12010114
Mao Z, Zheng Y-L, Zhang Y-Q. Behavioral Impairment and Oxidative Damage Induced by Chronic Application of Nonylphenol. International Journal of Molecular Sciences. 2011; 12(1):114-127. https://doi.org/10.3390/ijms12010114
Chicago/Turabian StyleMao, Zhen, Yuan-Lin Zheng, and Yan-Qiu Zhang. 2011. "Behavioral Impairment and Oxidative Damage Induced by Chronic Application of Nonylphenol" International Journal of Molecular Sciences 12, no. 1: 114-127. https://doi.org/10.3390/ijms12010114