Nuclear Translocation of Nuclear Factor Kappa B in First Trimester Deciduas and Chorionic Villi in Early Spontaneous Miscarriage Women

Abstract
:1. Introduction
2. Material and Methods
2.1. Subject Population Examined
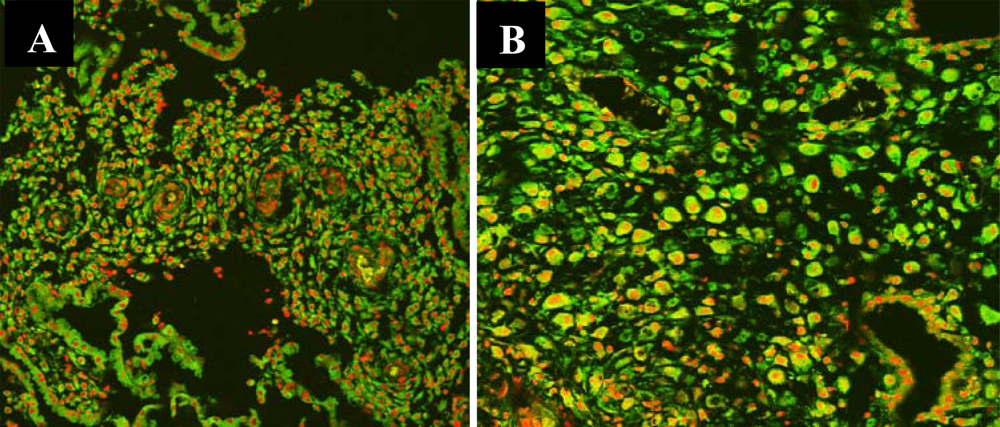
2.2. Measurement of Chorionic Villous and Decidual P65 Component of NF-κB by Confocal Laser Scanning Microscopy
2.3. Assessment of Chorionic Villous and Decidual P65 Component of NF-κB by SP Immunohistochemistry
3. Results
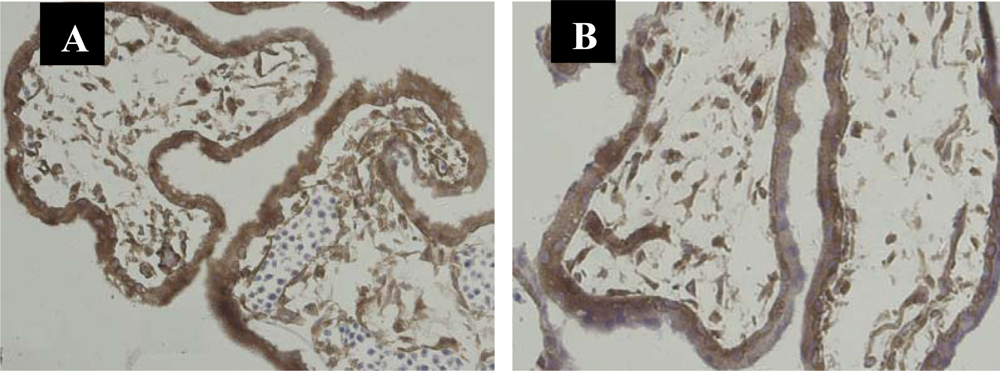
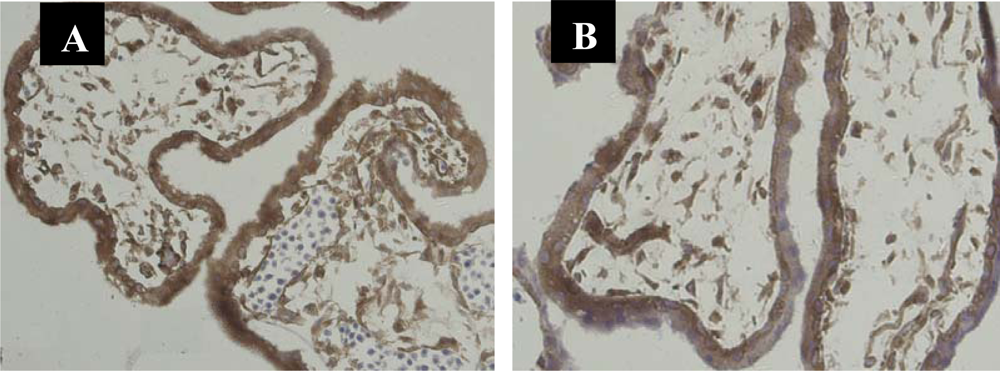
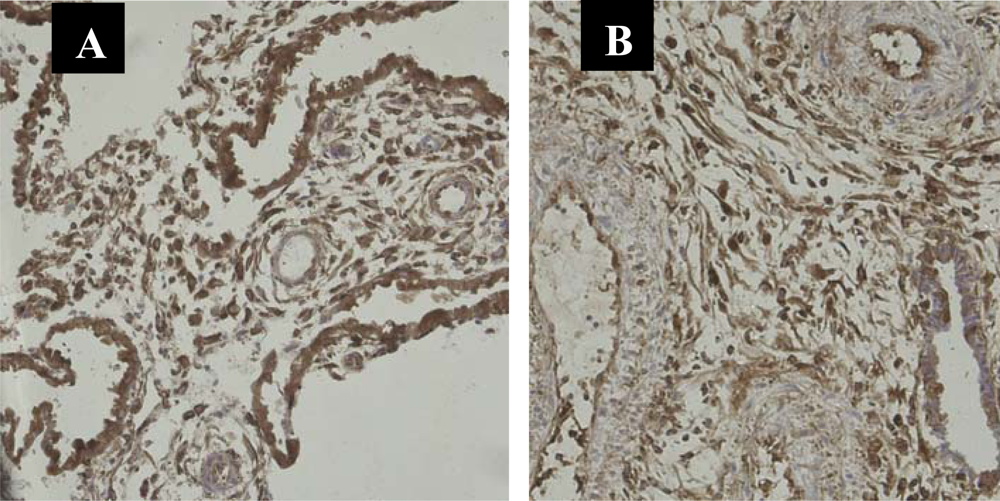
3.1. Immunolocalization of NF-κB Protein P65 in the Human Chorionic Villi and Deciduas
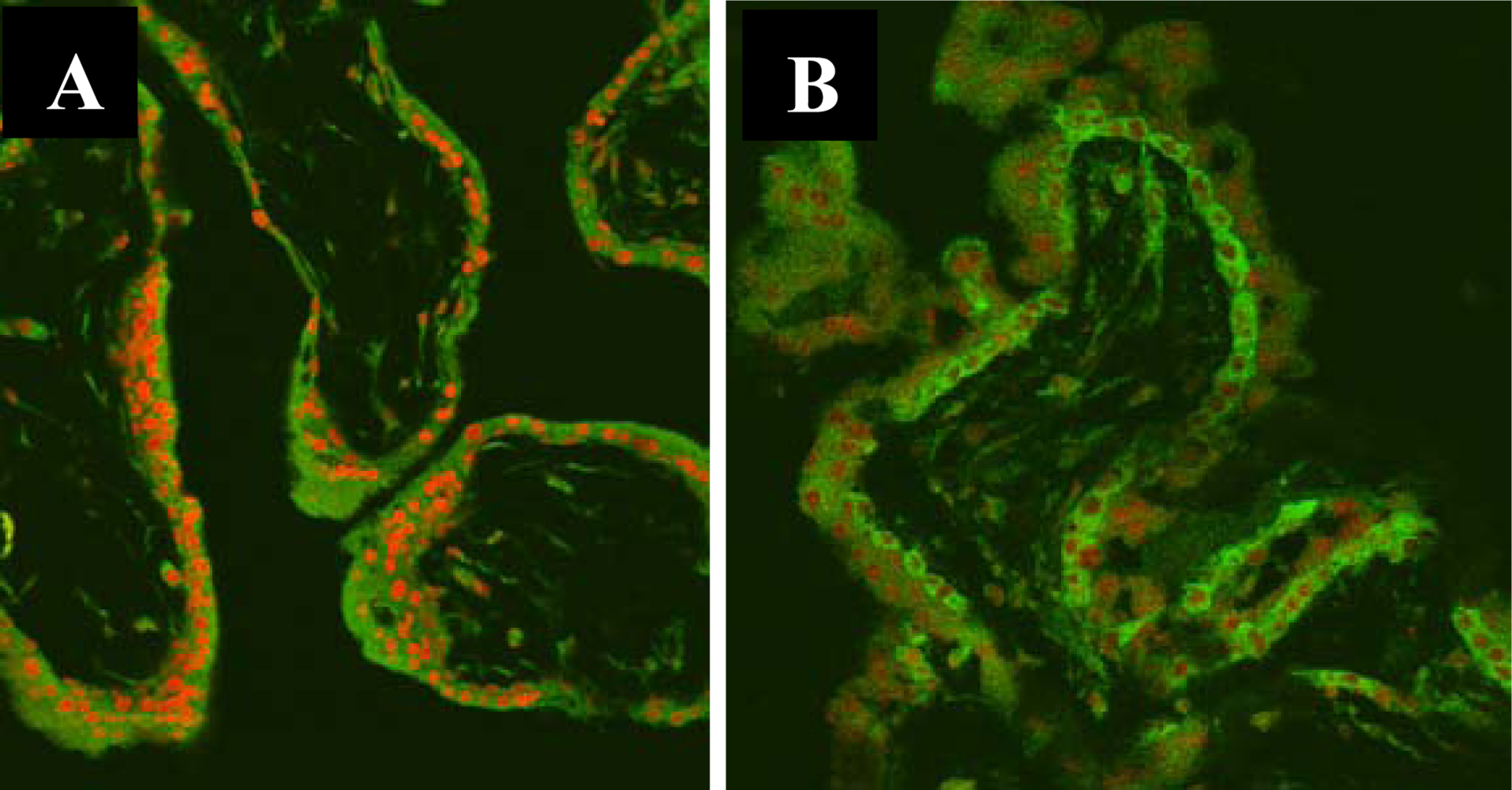
3.2. Nuclear Translocation of the NF-κB Protein P65 in the Human Chorionic Villi
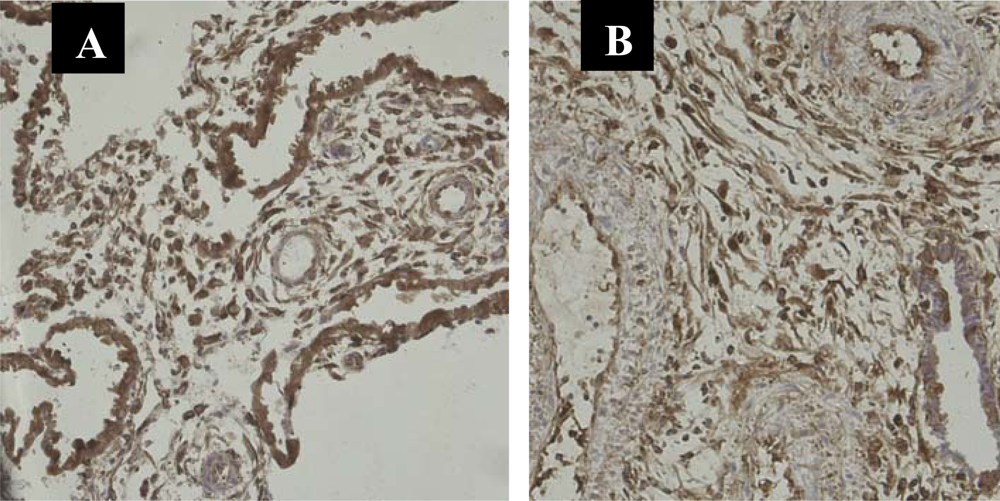
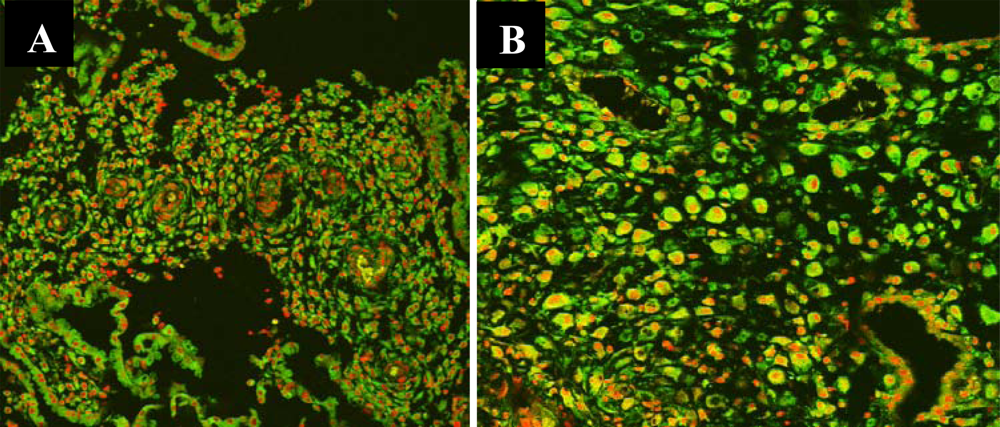
3.3. Nuclear Translocation of the NF-κB Protein P65 in the Human Deciduas
4. Discussion
5. Conclusions
References
- Raghupathy, R. Pregnancy: Success and failure within the Th1/Th2/Th3 paradigm. Semin. Immunol 2001, 13, 219–227. [Google Scholar]
- Chaouat, G; Zourbas, S; Ostojic, S; Lappree-Delage, G; Dubanchet, S; Ledee, N; Martal, J. A brief review of recent data on some cytokine expressions at the materno-foetal interface which might challenge the classical Th1/Th2 dichotomy. J. Reprod. Immunol 2002, 53, 241–256. [Google Scholar]
- Guttridge, DC; Albanese, C; Reuther, JY; Pestell, RG; Baldwin, AS. NF-κB controls cell growth and differentiation through transcriptional regulation of cyclin D1. Mol. Cell Biol 1999, 19, 5785–5799. [Google Scholar]
- Barkett, M; Gilmore, TD. Control of apoptosis by Rel/NF-κB transcription factors. Oncogene 1999, 18, 6910–6924. [Google Scholar]
- Barnes, PJ; Karin, M. Mechanisms of disease—nuclear factor-κB—a pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med 1997, 336, 1066–1071. [Google Scholar]
- N’guessan, PD; Hippenstiel, S; Etouem, MO; Zahlten, J; Beermann, W; Lindner, D; Opitz, B; Witzenrath, M; Rosseau, S; Suttorp, N; Schmeck, B. Streptococcus pneumoniae-induced p38 MAPK- and NF-κB-dependent COX-2 expression in human lung epithelium. Am. J. Physiol. Lung Cell Mol. Physiol 2006, 290, L1131–L1138. [Google Scholar]
- Kobielak, A; Fuchs, E. Links between α-catenin, NF-κB, and squamous cell carcinoma in skin. Proc. Natl .Acad. Sci. USA 2006, 103, 2322–2327. [Google Scholar]
- Knofler, M; Mosl, B; Bauer, S; Griesinger, G; Husslein, P. TNF-α/TNFR1 in primary and immortalized first trimester cytotrophoblasts. Placenta 2000, 21, 525–535. [Google Scholar]
- Rezaei, A; Dabbagh, A. T-helper(1) cytokines increase during early pregnancy in women with a history of recurrent spontaneous abortion. Med Sci Monit 2002, 8, CR607–CR610. [Google Scholar]
- Arslan, E; Colakoglu, M; Celik, C; Gezginc, K; Acar, A; Capar, M; Aköz, M; Akyürek, C. Serum TNF-alpha, IL-6, lupus anticoagulant and anticardiolipin antibody in women with and without a past history of recurrent miscarriage. Arch. Gynecol. Obstet 2004, 270, 227–229. [Google Scholar]
- Toder, V; Fein, A; Carp, H; Torchinsky, A. TNF-α in pregnancy loss and embryo maldevelopment: A Mediator of detrimental stimuli or a protector of the fetoplacental unit? J. Assist. Reprod. Genet 2003, 20, 73–81. [Google Scholar]
- Baud, V; Karin, M. Signal transduction by tumor necrosis factor and its relatives. Trends Cell Biol 2001, 11, 372–377. [Google Scholar]
- Yu, XW; Yan, CF; Jin, H; Li, X. Tumor necrosis factor receptor 1 expression and early spontaneous abortion. Int. J. Gynecol. Obestet 2005, 88, 44–48. [Google Scholar]
- Yu, XW; Wang, LQ; Yan, CF; Li, X. Expression and localization of tumor necrosis factor receptor 1 protein in the chorionic villi in early normal and spontaneous abortion. Eur. J. Obstet. Gynecol. Reprod. Biol 2006, 132, 58–63. [Google Scholar]
- Pahl, HL. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar]
- Karin, M; Lin, A. NF-κB at the crossroad of life and death. Nut Immunol 2002, 3, 221–227. [Google Scholar]
- Barkett, M; Gilmore, TD. Control of apoptosis by Rel/NF-κB transcription factors. Oncogene 1999, 18, 6910–6924. [Google Scholar]
- Garcia, KL; Coltrera, MD; Grown, AM. Analysis of proliferative grad using anti PCNA/cyclin monoclonal antibody in fixied, embedded tissues: comparison with flow cytometic analysis. Am. J. Pathol 1989, 134, 733–739. [Google Scholar]
- Aban, M; Cinel, L; Arslan, M; Dilek, U; Kaplanoglu, M; Arpaci, R; Dilek, S. Expression of nuclear factor-κB and placental apoptosis in pregnancies complicated with intrauterine growth restriction and preeclampsia: immunohistochemical study. Tohoku J. Exp. Med 2004, 204, 195–202. [Google Scholar]
- Tsukihara, S; Harada, T; Deura, I; Mitsunari, M; Yoshida, S; Iwabe, T; Terakawa, N. Interleukin-1β-induced expression of IL-6 and production of human chorionic gonadotropin in human trophoblast cells via nuclear factor-kappaB activation. Am. J. Reprod. Immunol 2004, 52, 218–223. [Google Scholar]
- Laird, SM; Tuckerman, EM; Cork, BA; Li, TC. Expression of NF-κB in human endometrium; role in the control of interleukin 6 and leukaemia inhibitory factor production. Mol. Hum. Reprod 2000, 6, 34–40. [Google Scholar]
- Lappas, M; Permezel, M; Georgiou, HM; Rice, GE. Nuclear factor κB regulation of proinflammatory cytokines in human gestational tissues in vitro. Biol. Reprod 2002, 67, 668–673. [Google Scholar]
- King, AE; Critchley, HOD; Kelly, RE. The NF-κB pathway in human endometrium and first trimester decidua. Mol. Hum. Reprod 2001, 7, 175–183. [Google Scholar]
- Kelly, RW; King, AE; Critchley, HO. Cytokine control in human endometrium. Reproduction 2001, 121, 3–19. [Google Scholar]
- McCracken, SA; Gallery, E; Morris, JM. Pregnancy-specific down-regulation of NF-κB expression in T cells in humans is essential for the maintenance of the cytokine profile required for pregnancy success. J. Immunol 2004, 172, 4583–4591. [Google Scholar]
- Hirano, S; Furutama, D; Hanafusa, T. Physiologically high concentrations of 17 β-estradiol enhance NF-κB activity in human T cells. Am. J. Physiol. Regul. Integr. Comp. Physiol 2007, 292, R1465–R1471. [Google Scholar]
- Lacey, H; Haigh, T; Westwood, M; Aplin, JD. Mesenchymally-derived Insulin-like growth factor 1 provides a paracrine stimulus for trophoblast migration. BMC Dev. Biol 2002, 2, 5–12. [Google Scholar]
- Westwood, M. Role of insulin-like growth factor binding protein 1 in human pregnancy. Rev. Reprod 1999, 4, 160–167. [Google Scholar]
- Chan, CW; Lao, TT; Ho, PC; Sung, EP; Cheung, AY. The effect of mifepristone on the expression of steroid hormone receptors in human decidua and placenta: A randomized placebo-controlled double-blind study. Clin. Endoc. Metab 2003, 88, 5846–5850. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| − | + | ++ | +++ | |
|---|---|---|---|---|
| SM group | 4 | 16 | 9 | 2 |
| Control group | 0 | 4 | 14 | 12 |
| P* | <0.001 |
| stromal cells | Glandular epithelial cells | |||||||
|---|---|---|---|---|---|---|---|---|
| − | + | ++ | +++ | − | + | ++ | +++ | |
| SM group | 0 | 0 | 2 | 29 | 2 | 14 | 9 | 6 |
| Control group | 0 | 5 | 9 | 16 | 0 | 2 | 10 | 8 |
| P* | 0.001 | <0.001 | ||||||
| stromal cells | vessel endothelial cells | syncytiotrophoblast cells | cytotrophoblast cells | |
|---|---|---|---|---|
| SM group | 0.7602 ± 0.0775 | 0.7191 ± 0.0516 | 0.6632 ± 0.0564 | 0.7042 ± 0.0619 |
| Control group | 0.6977 ± 0.0636 | 0.6896 ± 0.0705 | 0.6336 ± 0.0823 | 0.6701 ± 0.0778 |
| T | 3.511 | 1.514 | 1.741 | 1.935 |
| P | 0.001 | 0.135 | 0.086 | 0.057 |
| glandular epithelial cells | stromal cells | Vessel endothelial cells | |
|---|---|---|---|
| SM group | 0.7184 ± 0.0553 | 0.8235 ± 0.0665 | 0.7484 ± 0.0700 |
| Control group | 0.6484 ± 0.0642 | 0.5840 ± 0.0503 | 0.6625 ± 0.0576 |
| T | 4.422 | 15.201 | 5.029 |
| P | <0.001 | <0.001 | <0.001 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, L.-q.; Yu, X.-w.; Yan, C.-f.; Wang, X. Nuclear Translocation of Nuclear Factor Kappa B in First Trimester Deciduas and Chorionic Villi in Early Spontaneous Miscarriage Women. Int. J. Mol. Sci. 2010, 11, 521-531. https://doi.org/10.3390/ijms11020521
Wang L-q, Yu X-w, Yan C-f, Wang X. Nuclear Translocation of Nuclear Factor Kappa B in First Trimester Deciduas and Chorionic Villi in Early Spontaneous Miscarriage Women. International Journal of Molecular Sciences. 2010; 11(2):521-531. https://doi.org/10.3390/ijms11020521
Chicago/Turabian StyleWang, Li-qin, Xue-wen Yu, Chun-fang Yan, and Xiang Wang. 2010. "Nuclear Translocation of Nuclear Factor Kappa B in First Trimester Deciduas and Chorionic Villi in Early Spontaneous Miscarriage Women" International Journal of Molecular Sciences 11, no. 2: 521-531. https://doi.org/10.3390/ijms11020521