
OAT3 Participates in Drug–Drug Interaction between Bentysrepinine and Entecavir through Interactions with M8—A Metabolite of Bentysrepinine—In Rats and Humans In Vitro
 ,
,
Abstract
:1. Introduction
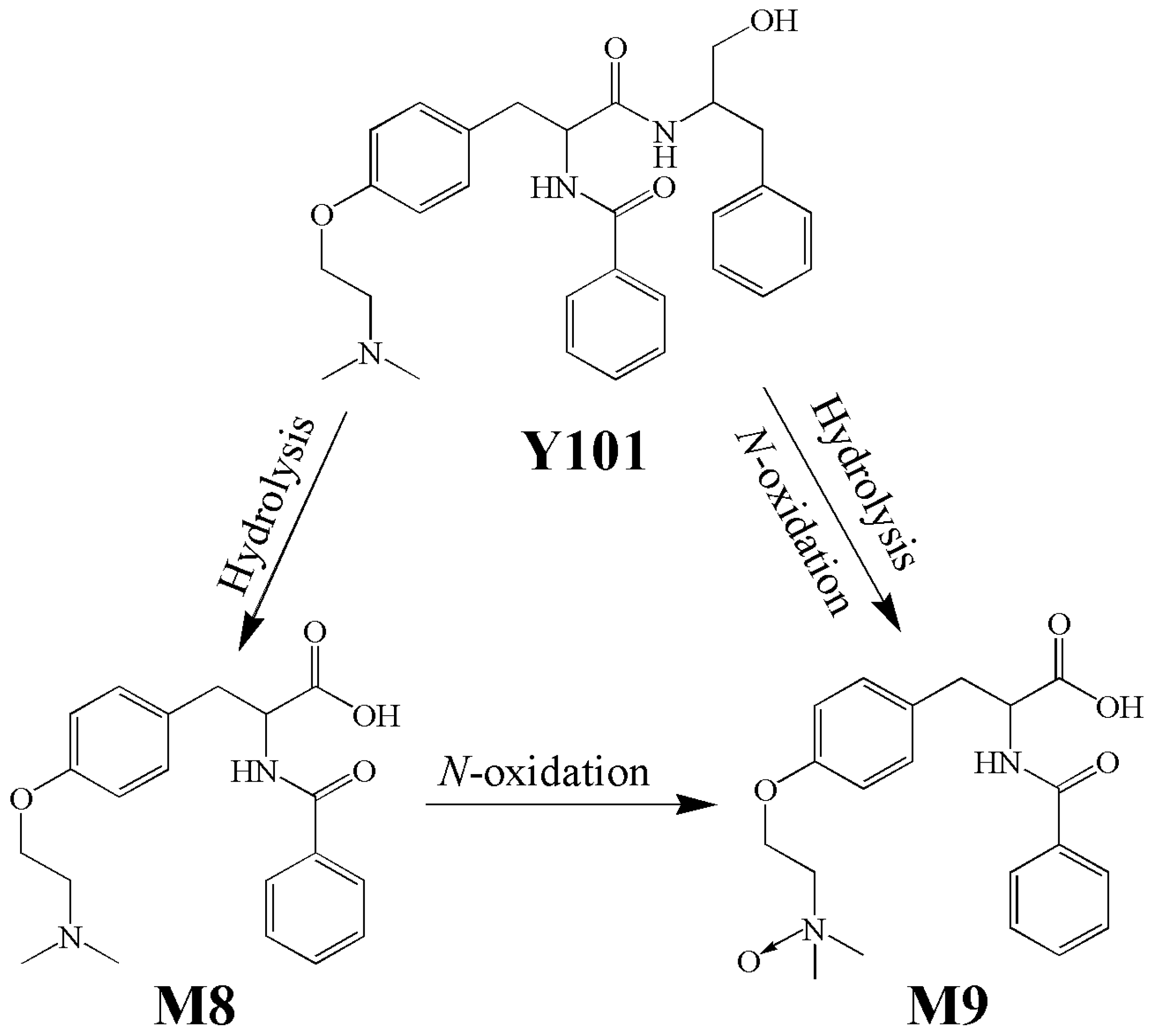
2. Results
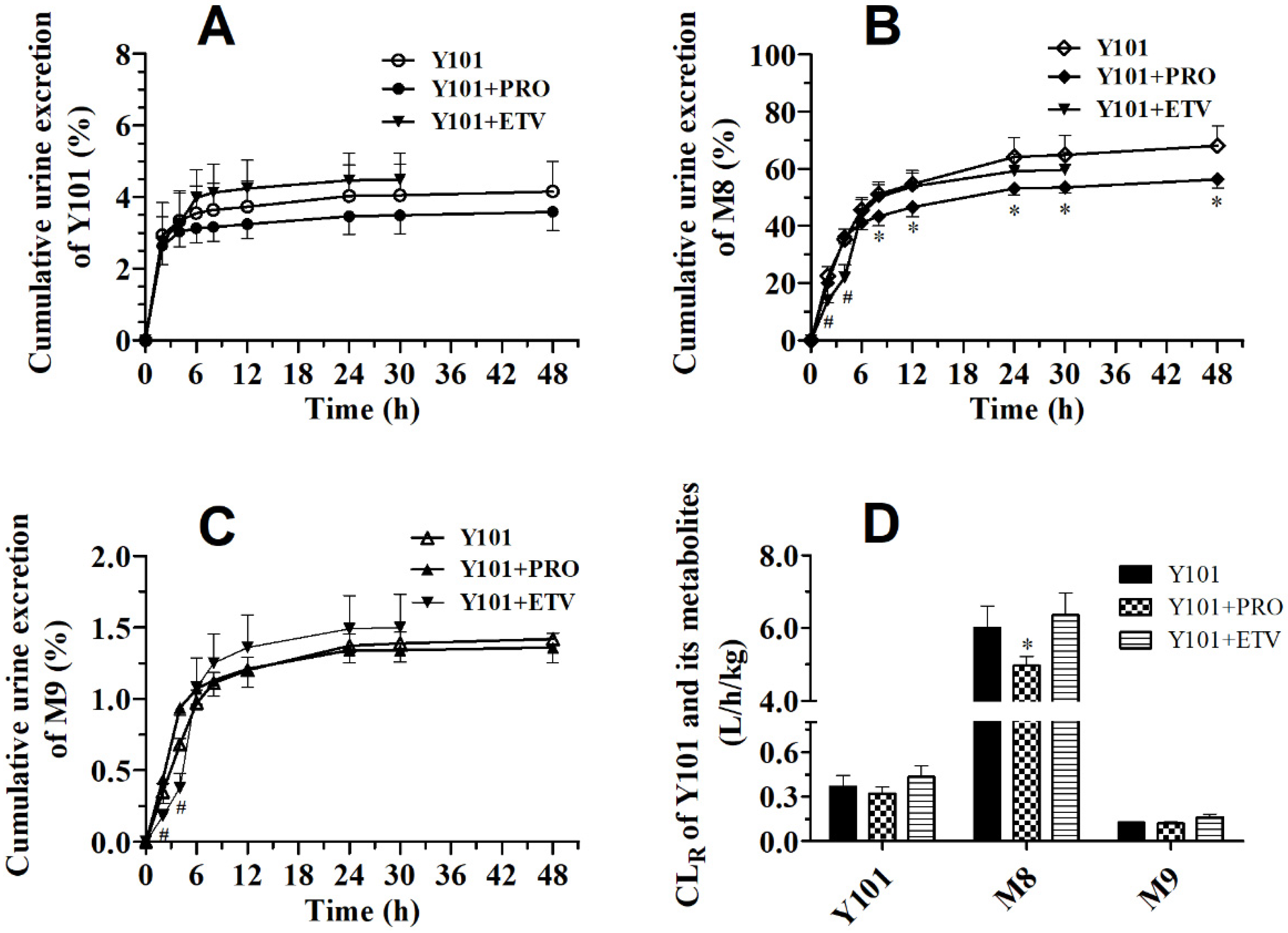
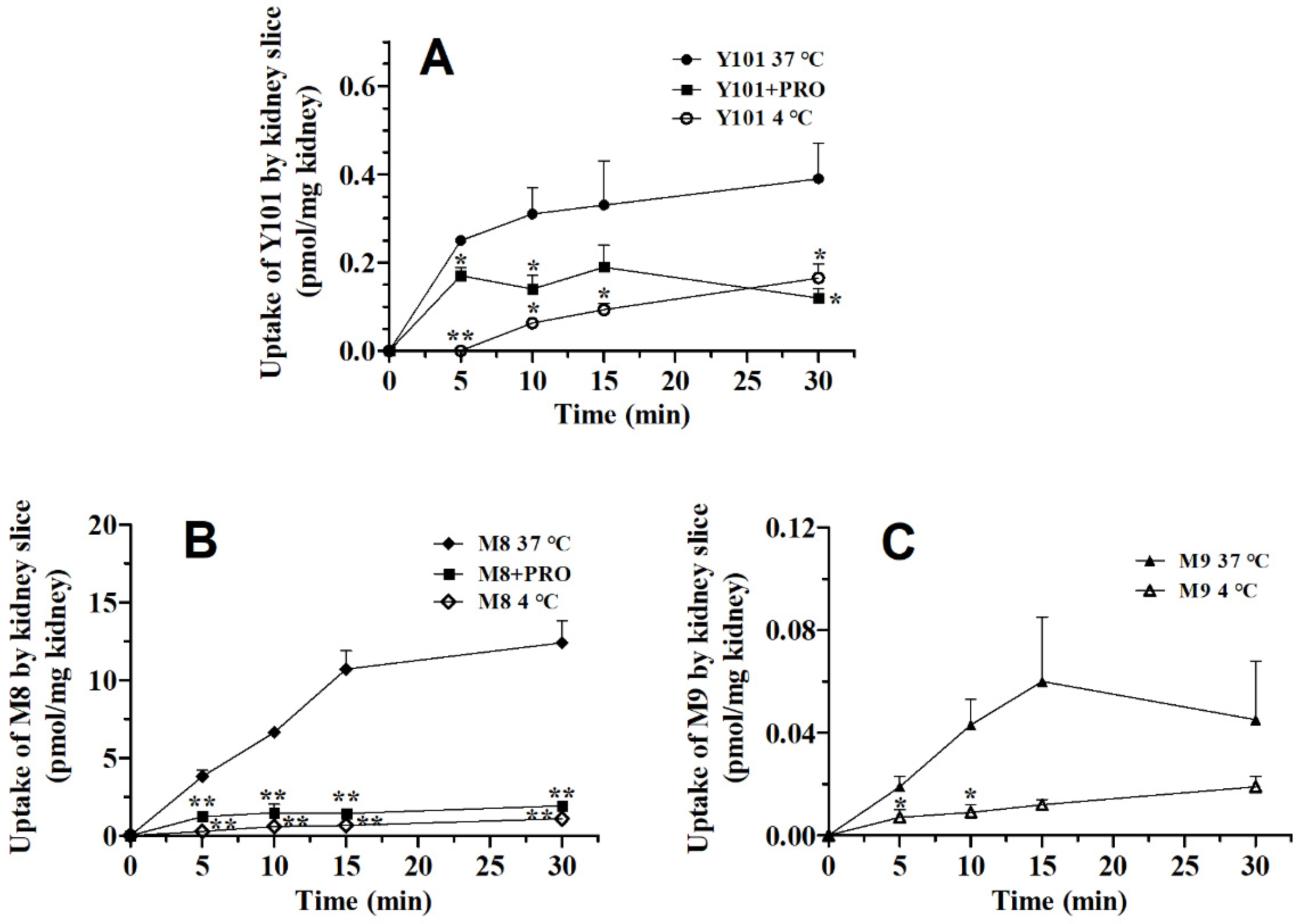
2.1. Effect of PRO on Pharmacokinetics of Y101, M8 and M9 in Rats
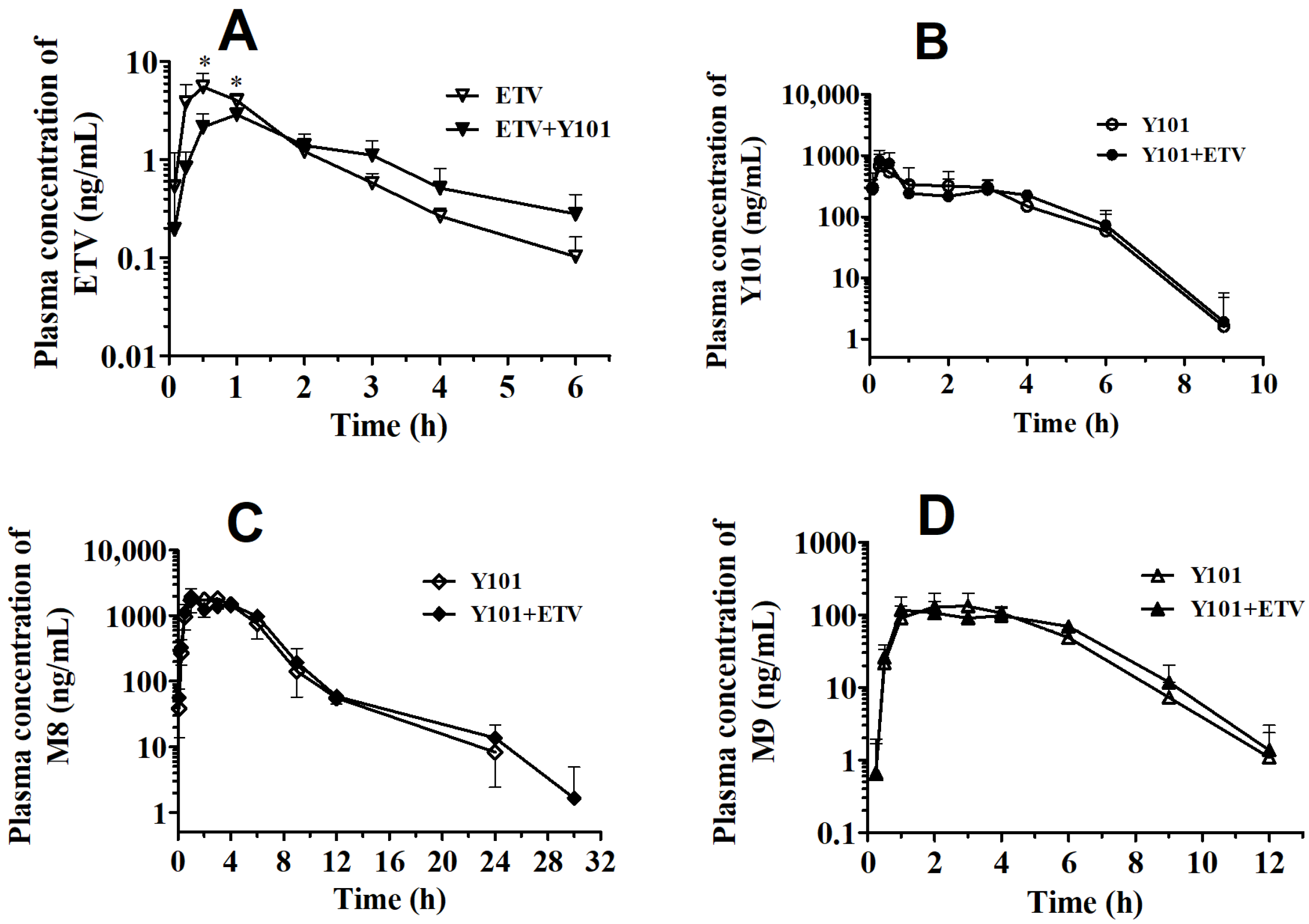
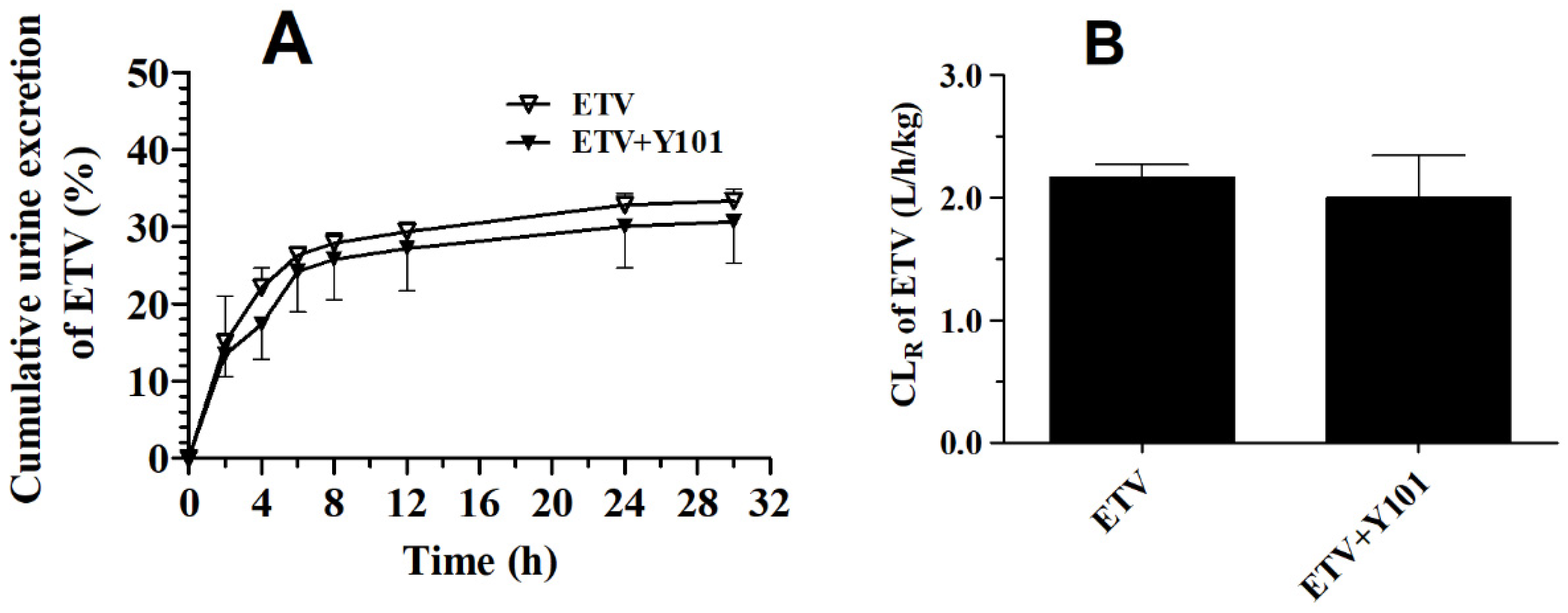
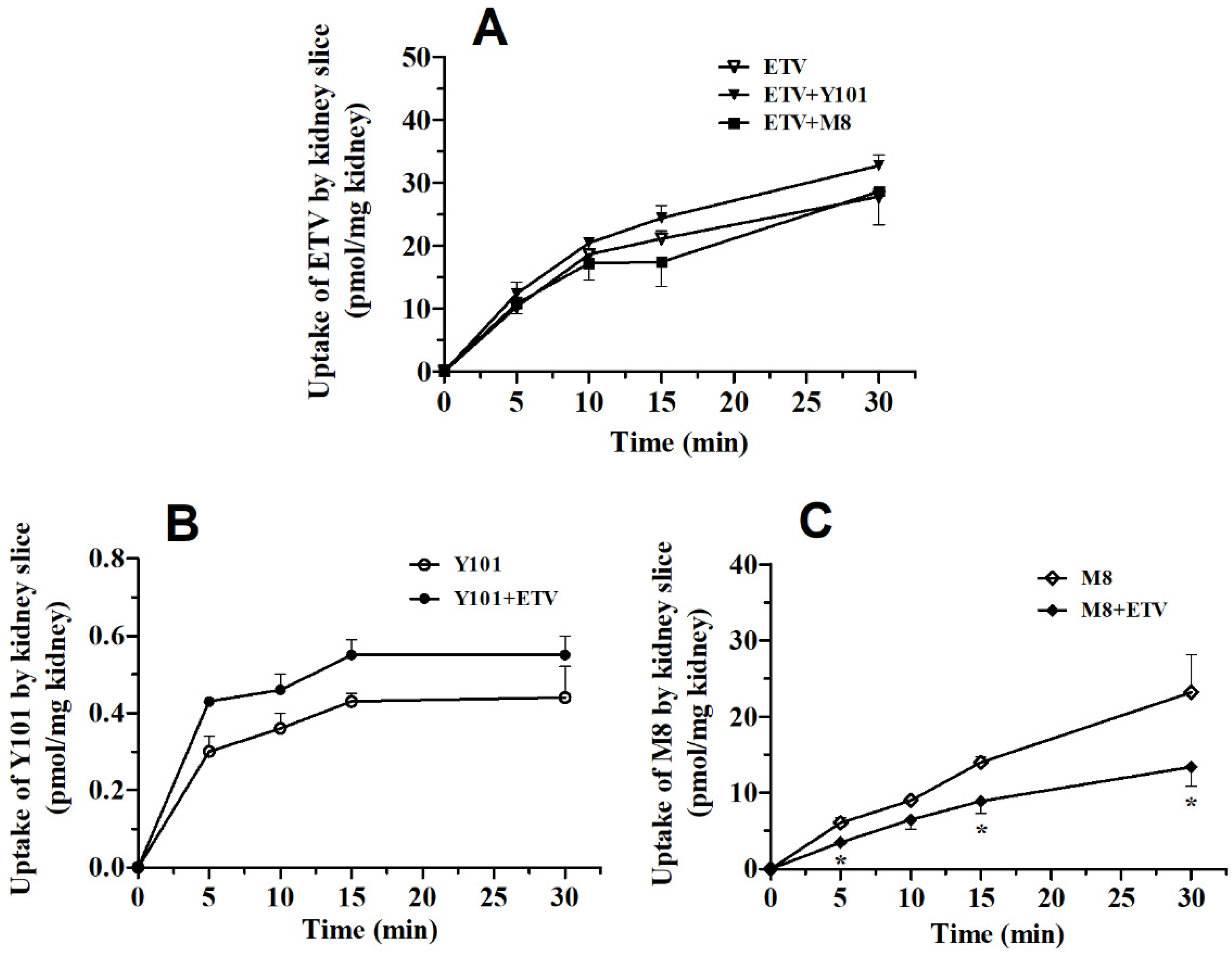
2.2. Pharmacokinetic DDI between Y101 and ETV in Rats
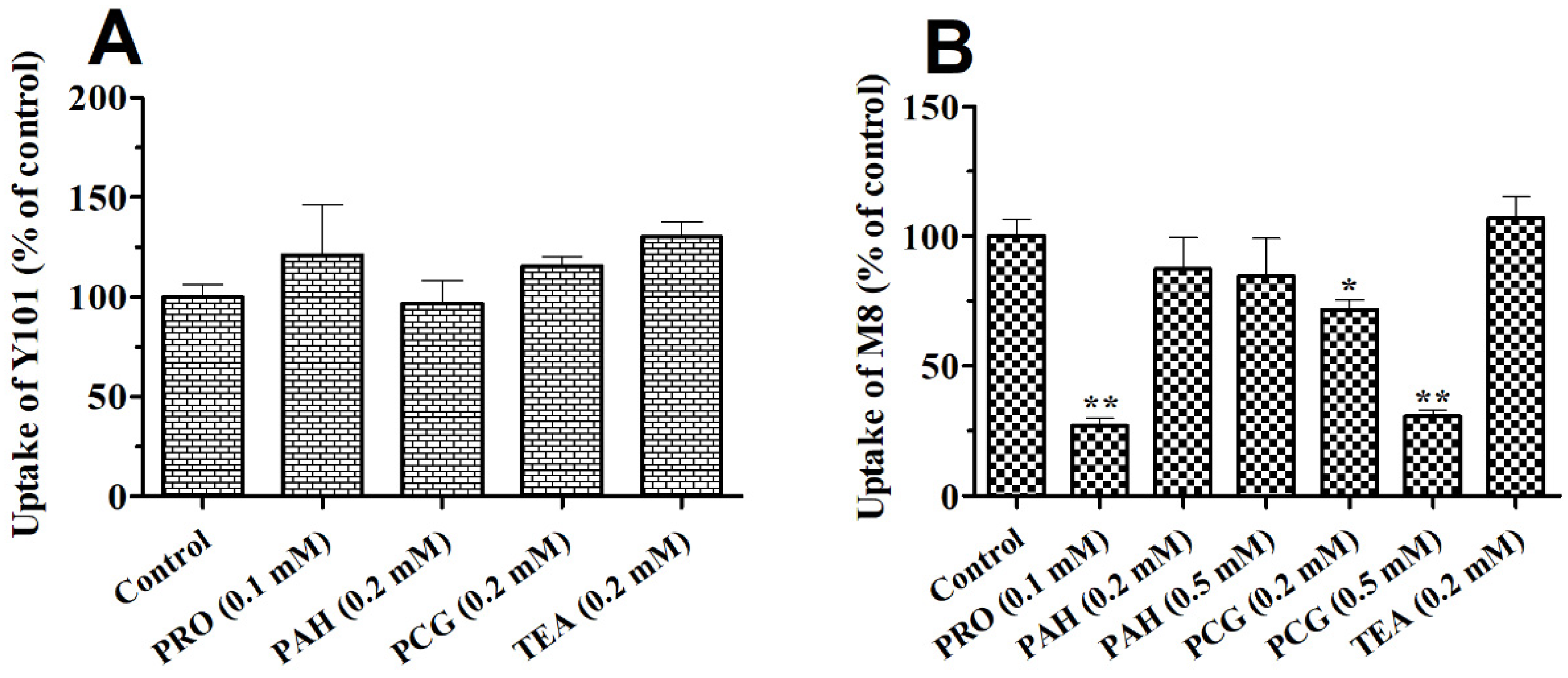
2.3. Effects of OAT Substrates, Inhibitors and ETV on the Uptake of Y101 or M8 in Rat Kidney Slices
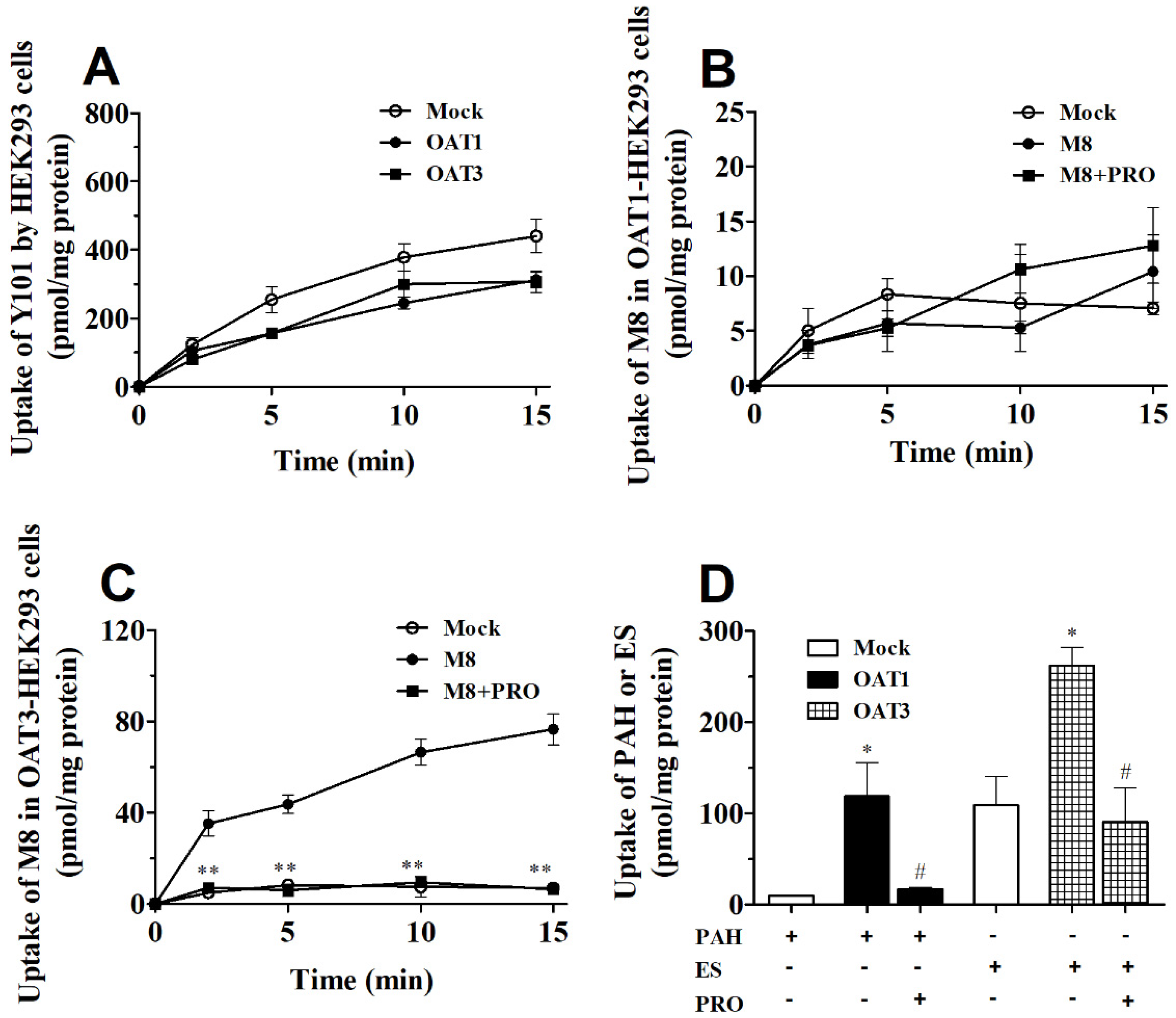
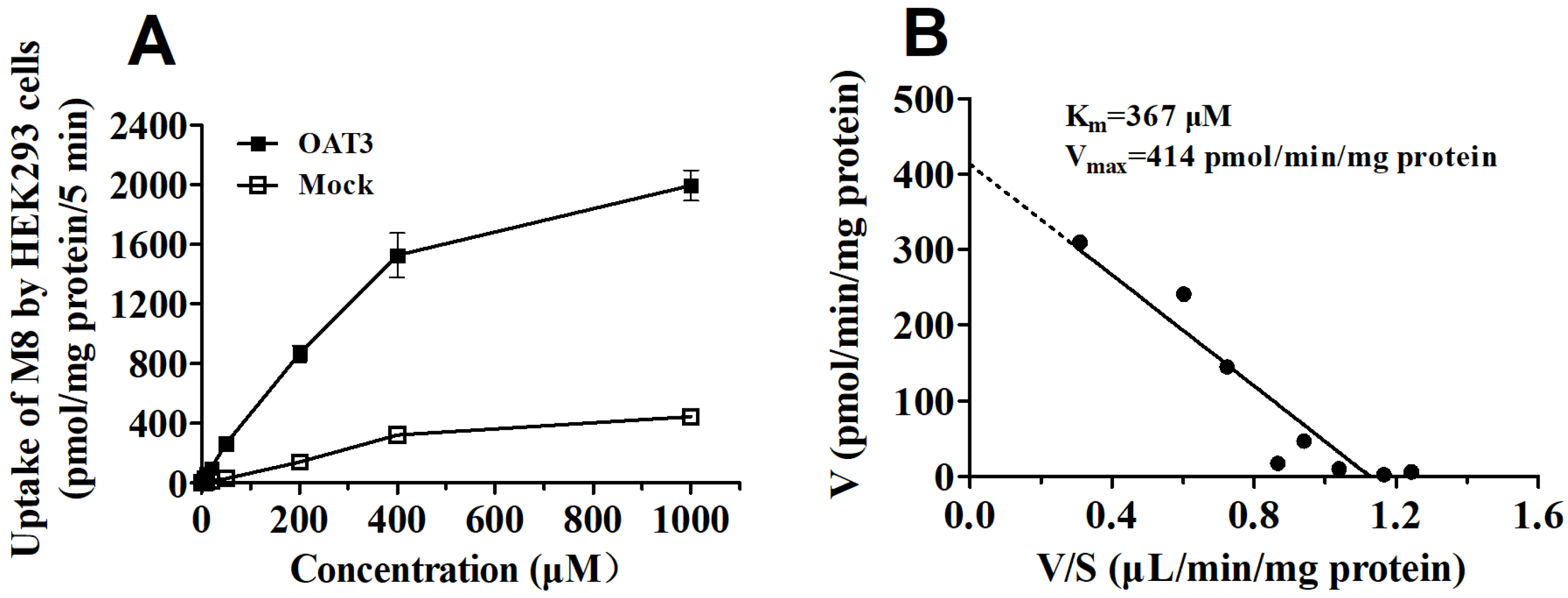
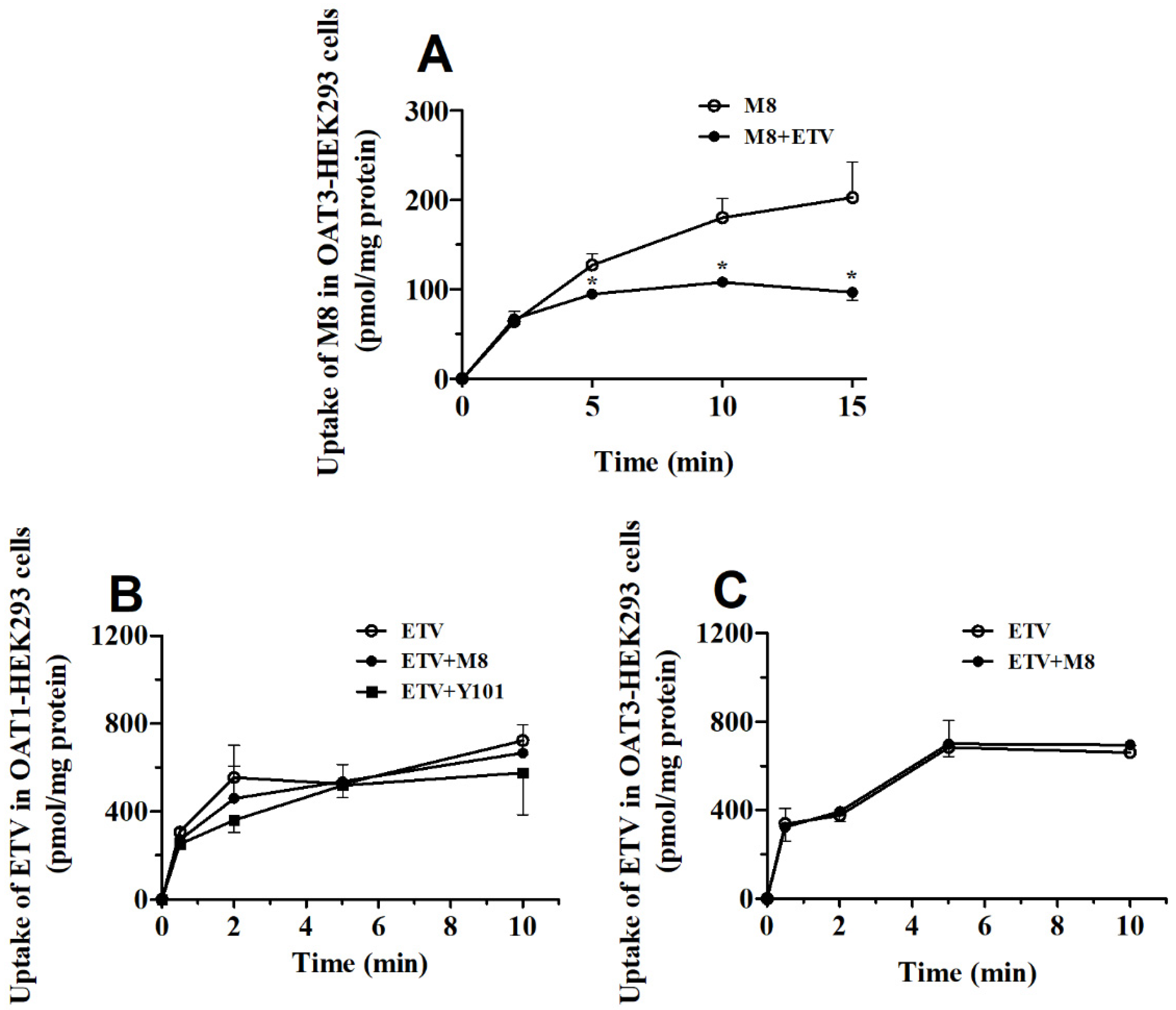
2.4. DDI between Y101 and ETV in hOAT1 and hOAT3-HEK293 Cells
3. Discussion
4. Material and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Pharmacokinetic Study in Rats
4.4. Renal Excretion Study in Rats
4.5. In Vitro Uptake Assay in Kidney Slices
4.6. In Vitro Uptake Assay in hOAT1 and hOAT3-HEK293 Cells
4.7. Sample Preparation and LC-MS/MS Analysis
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Liu, X.; Xue, L.; Zhang, H.; Xu, Q.; Zhang, S.; Ma, S.; Ding, X.; Liu, L.; Dong, J.; Qian, L.; et al. Phase I, first-in-human, single and multiple ascending dose- and food-effect studies to assess the safety, tolerability and pharmacokinetics of a novel anti-hepatitis B virus drug, bentysrepinine (Y101), in healthy Chinese subjects. Clin. Drug Investig. 2020, 40, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Xu, B.; Huang, Z.; Pan, W.; Cao, P.; Liu, C.; Hao, X.; Song, B.; Liang, G. Synthesis and biological evaluation of Matijing-Su derivatives as potent anti-HBV agents. Bioorg. Med. Chem. 2011, 19, 5352–5360. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Huang, Z.; Liu, C.; Cai, Z.; Pan, W.; Cao, P.; Hao, X.; Liang, G. Synthesis and anti-hepatitis B virus activities of Matijing-Su derivatives. Bioorg. Med. Chem. 2009, 17, 3118–3125. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Xu, W.; Li, Y.; Huang, Z.; Liang, G.; Liu, C. In Silico Molecular Docking Study of Repensine and Bentysrepinine against HBV DNA Polymerase. Chin. Herb. Med. 2015, 7, 39–44. [Google Scholar] [CrossRef]
- Yu, S.; Xu, X.; Li, K.; Liang, G.; Huang, Z. Protective effect of phenylalanine dipeptide compounds Y101 on mice models of ANIT hepatic injury. Pharm. J. Chin. PLA 2011, 27, 118–121. [Google Scholar]
- Yu, S.; Xu, X.; Liang, G.; Huang, Z. Protective effects of phenylalanine dipeptide compounds Y101 on liver injury induced by CnoA in mice. Chin. J. New Drugs 2012, 21, 78–87. [Google Scholar]
- Liu, X.; Huang, C.; Xue, L.; Xu, Q.; Xia, W.; Li, X.; Miao, L. Simultaneous determination of bentysrepinine (Y101) and its metabolites M8 and M9 in human plasma by UPLC-MS/MS and its application to a pharmacokinetic study. J. Pharm. Biomed. Anal. 2018, 150, 287–293. [Google Scholar] [CrossRef]
- Gong, M.; Meng, F.; Fan, H.; Dong, S.; Wang, Y.; Li, Y.; Liang, G.; Liu, C. Molecular Simulation Study on Bentysrepinine Metabolites Improving Binding Affinity of HBV DNA Polymerase. Chin. Herb. Med. 2016, 8, 139–142. [Google Scholar] [CrossRef]
- Ye, J.; Liu, Q.; Wang, C.; Meng, Q.; Peng, J.; Sun, H.; Kaku, T.; Liu, K. Inhibitory effect of JBP485 on renal excretion of acyclovir by the inhibition of OAT1 and OAT3. Eur. J. Pharm. Sci. 2012, 47, 341–346. [Google Scholar] [CrossRef]
- Huo, X.; Liu, Q.; Wang, C.; Meng, Q.; Sun, H.; Peng, J.; Ma, X.; Sun, P.; Liu, K. Inhibitory effect of valsartan on the intestinal absorption and renal excretion of bestatin in rats. J. Pharm. Sci. 2014, 103, 719–729. [Google Scholar] [CrossRef]
- Huo, X.; Liu, K. Renal organic anion transporters in drug-drug interactions and diseases. Eur. J. Pharm. Sci. 2018, 112, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Meng, Q.; Yang, M.; Liu, D.; Hou, X.; Tang, L.; Wang, X.; Lyu, Y.; Chen, X.; Liu, K.; et al. Current trends in drug metabolism and pharmacokinetics. Acta Pharm. Sin. B 2019, 9, 1113–1144. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.S.Y.; Covert, E.; Wilson, E.; Kottilil, S. Chronic Hepatitis B infection: A review. JAMA 2018, 319, 1802–1813. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.H.; Bifano, M.; Olsen, S.; Smith, R.A.; Zhang, D.; Grasela, D.M.; LaCreta, F. Entecavir pharmacokinetics, safety, and tolerability after multiple ascending doses in healthy subjects. J. Clin. Pharmacol. 2006, 46, 1250–1258. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, C.; Meng, Q.; Liu, Q.; Sun, H.; Peng, J.; Ma, X.; Kaku, T.; Liu, K. OAT1 and OAT3: Targets of drug-drug interaction between entecavir and JBP485. Eur. J. Pharm. Sci. 2013, 48, 650–657. [Google Scholar] [CrossRef]
- Yang, X.; Ma, Z.; Zhou, S.; Weng, Y.; Lei, H.; Zeng, S.; Li, L.; Jiang, H. Multiple drug transporters are involved in renal secretion of entecavir. Antimicrob. Agents Chemother. 2016, 60, 6260–6270. [Google Scholar] [CrossRef] [Green Version]
- Shu, W.; Ma, L.; Hu, X.; Zhang, M.; Chen, W.; Ma, W.; Huang, J.; Li, J. Drug-drug interaction between crizotinib and entecavir via renal secretory transporter OCT2. Eur. J. Pharm. Sci. 2020, 142, 105153. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Prevention, Care and Treatment of Persons with Chronic Hepatitis B Infection; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Huo, X.; Meng, Q.; Wang, C.; Zhu, Y.; Liu, Z.; Ma, X.; Ma, X.; Peng, J.; Sun, H.; Liu, K. Cilastatin protects against imipenem-induced nephrotoxicity via inhibition of renal organic anion transporters (OATs). Acta Pharm. Sin. B 2019, 9, 986–996. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, C.; Liu, Q.; Meng, Q.; Peng, J.; Sun, H.; Ma, X.; Huo, X.; Liu, K. Involvement of organic anion-transporting polypeptides in the hepatic uptake of dioscin in rats and humans. Drug Metab. Dispos. 2013, 41, 994–1003. [Google Scholar] [CrossRef] [Green Version]
- Mandíková, J.; Volková, M.; Pávek, P.; Navrátilová, L.; Hyršová, L.; Janeba, Z.; Pavlík, J.; Bárta, P.; Trejtnar, F. Entecavir interacts with influx transporters hOAT1, hCNT2, hCNT3, but not with hOCT2: The potential for renal transporter-mediated cytotoxicity and drug-drug interactions. Front. Pharmacol. 2016, 6, 304. [Google Scholar] [CrossRef] [Green Version]
- Matthews, S.J. Entecavir for the treatment of chronic hepatitis B virus infection. Clin Ther. 2006, 28, 184–203. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Meng, Q.; Wang, C.; Liu, Q.; Sun, H.; Kaku, T.; Liu, K. Organic anion transporters involved in the excretion of bestatin in the kidney. Peptides 2012, 33, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Zhang, A.; Liao, C.; Yang, Y.; Zhang, L.; Liu, J.; Xia, Y.; Si, D.; Dong, S.; Liu, C. In vitro metabolism and in vivo pharmacokinetics of bentysrepinine (Y101), an investigational new drug for anti-HBV-infected hepatitis: Focus on interspecies comparison. Xenobiotica 2020, 50, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huo, X.; Wang, C.; Meng, Q.; Liu, Z.; Sun, H.; Tan, A.; Ma, X.; Peng, J.; Liu, K. Organic anion transporters also mediate the drug-drug interaction between imipenem and cilastatin. Asian J. Pharm. Sci. 2020, 15, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Yang, F.; Zhao, L.; Li, C.; Dong, S.; Liu, J.; Fan, H. Effects of verapamil on pharmacokinetics of Y101 in vivo in rats. Chin. Pharmacol. Bull. 2021, 37, 1122–1127. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Parameters | Unit | Y101 | Y101 + PRO | ||||
|---|---|---|---|---|---|---|---|---|
| Y101 | C0 | ng/mL | 16,029 | ± | 7069 | 15,770 | ± | 3818 |
| AUC0-t | ng·h/mL | 2838 | ± | 481 | 2018 | ± | 173 | |
| AUC0-∞ | ng·h/mL | 2855 | ± | 488 | 2031 | ± | 173 | |
| Vd | L/kg | 10.2 | ± | 1.22 | 12.5 | ± | 3.74 | |
| t1/2 | h | 0.798 | ± | 0.119 | 0.706 | ± | 0.227 | |
| CLp | L/h/kg | 8.94 | ± | 1.65 | 12.4 * | ± | 1.01 | |
| CLR | L/h/kg | 0.366 | ± | 0.0742 | 0.317 | ± | 0.0463 | |
| M8 | Cmax | ng/mL | 3887 | ± | 567 | 10,133 ** | ± | 702 |
| Tmax | h | 0.667 | ± | 0.289 | 0.500 | ± | 0 | |
| AUC0-t | ng·h/mL | 8299 | ± | 278 | 21,865 ** | ± | 620 | |
| AUC0-∞ | ng·h/mL | 8356 | ± | 274 | 21,919 ** | ± | 626 | |
| t1/2 | h | 4.07 | ± | 1.11 | 3.45 | ± | 0.955 | |
| CLR | L/h/kg | 6.00 | ± | 0.614 | 4.96 * | ± | 0.265 | |
| M9 | Cmax | ng/mL | 156 | ± | 27.7 | 219 * | ± | 16.5 |
| Tmax | h | 1.17 | ± | 0.289 | 1.33 | ± | 0.289 | |
| AUC0-t | ng·h/mL | 335 | ± | 42.3 | 527 ** | ± | 21.4 | |
| AUC0-∞ | ng·h/mL | 343 | ± | 44.9 | 536 ** | ± | 15.2 | |
| t1/2 | h | 1.01 | ± | 0.143 | 1.32 | ± | 0.414 | |
| CLR | L/h/kg | 0.125 | ± | 0.00356 | 0.120 | ± | 0.00963 | |
| Compound | Parameters | Unit | ETV | Y101 + ETV | ||||
|---|---|---|---|---|---|---|---|---|
| ETV | C0 | ng/mL | 26.4 | ± | 1.63 | 19.1 | ± | 10.0 |
| AUC0-t | ng·h/mL | 9.21 | ± | 2.07 | 6.31 | ± | 1.14 | |
| AUC0-∞ | ng·h/mL | 9.79 | ± | 2.41 | 6.76 | ± | 1.24 | |
| Vd | L/kg | 11.7 | ± | 3.19 | 16.4 | ± | 4.33 | |
| t1/2 | h | 1.28 | ± | 0.233 | 1.28 | ± | 0.393 | |
| CLp | L/h/kg | 6.41 | ± | 1.53 | 9.09 * | ± | 1.52 | |
| CLR | L/h/kg | 2.17 | ± | 0.101 | 2.00 | ± | 0.347 | |
| Compound | Parameters | Unit | Y101 | Y101 + ETV | ||||
| Y101 | C0 | ng/mL | 10,640 | ± | 1350 | 6281 | ± | 3110 |
| AUC0-t | ng·h/mL | 2353 | ± | 272 | 1674 | ± | 462 | |
| AUC0-∞ | ng·h/mL | 2367 | ± | 269 | 1692 | ± | 456 | |
| Vd | L/kg | 10.6 | ± | 2.89 | 15.5 | ± | 7.55 | |
| t1/2 | h | 0.682 | ± | 0.146 | 0.660 | ± | 0.187 | |
| CLp | L/h/kg | 10.7 | ± | 1.15 | 15.5 | ± | 3.76 | |
| CLR | L/h/kg | 0.431 | ± | 0.0921 | 0.476 | ± | 0.0788 | |
| M8 | Cmax | ng/mL | 2665 | ± | 524 | 2520 | ± | 363 |
| Tmax | h | 0.500 | ± | 0.354 | 0.438 | ± | 0.125 | |
| AUC0-t | ng·h/mL | 5828 | ± | 726 | 6096 | ± | 726 | |
| AUC0-∞ | ng·h/mL | 5862 | ± | 735 | 6146 | ± | 742 | |
| t1/2 | h | 1.93 | ± | 0.378 | 2.65 * | ± | 0.384 | |
| CLR | L/h/kg | 6.90 | ± | 0.719 | 6.35 | ± | 0.614 | |
| M9 | Cmax | ng/mL | 134 | ± | 27.9 | 171 | ± | 55.1 |
| Tmax | h | 1.13 | ± | 0.250 | 1.25 | ± | 0.289 | |
| AUC0-t | ng·h/mL | 547 | ± | 196 | 464 | ± | 168 | |
| AUC0-∞ | ng·h/mL | 553 | ± | 196 | 471 | ± | 168 | |
| t1/2 | h | 1.14 | ± | 0.143 | 1.80 * | ± | 0.282 | |
| CLR | L/h/kg | 0.148 | ± | 0.00798 | 0.160 | ± | 0.0246 | |
| Compound | Parameters | Unit | ETV | ETV + Y101 | ||||
|---|---|---|---|---|---|---|---|---|
| ETV | Cmax | ng/mL | 5.54 | ± | 2.08 | 2.94 | ± | 0.351 |
| Tmax | h | 0.5 | ± | 0 | 0.875 | ± | 0.250 | |
| AUC0-t | ng·h/mL | 8.15 | ± | 1.97 | 6.76 | ± | 0.941 | |
| AUC0-∞ | ng·h/mL | 8.49 | ± | 1.98 | 7.41 | ± | 1.33 | |
| t1/2 | h | 1.10 | ± | 0.392 | 1.45 | ± | 0.334 | |
| Compound | Parameters | Unit | Y101 | Y101 + ETV | ||||
| Y101 | Cmax | ng/mL | 668 | ± | 392 | 843 | ± | 418 |
| Tmax | h | 0.250 | ± | 0 | 0.313 | ± | 0.125 | |
| AUC0-t | ng·h/mL | 1574 | ± | 394 | 1617 | ± | 164 | |
| AUC0-∞ | ng·h/mL | 1622 | ± | 368 | 1681 | ± | 183 | |
| t1/2 | h | 0.998 | ± | 0.291 | 1.11 | ± | 0.261 | |
| M8 | Cmax | ng/mL | 2033 | ± | 536 | 1983 | ± | 563 |
| Tmax | h | 2.25 | ± | 0.957 | 1.50 | ± | 1.00 | |
| AUC0-t | ng·h/mL | 10,129 | ± | 731 | 10,339 | ± | 572 | |
| AUC0-∞ | ng·h/mL | 10,203 | ± | 709 | 10,385 | ± | 572 | |
| t1/2 | h | 3.44 | ± | 1.52 | 3.64 | ± | 0.521 | |
| M9 | Cmax | ng/mL | 144 | ± | 59.1 | 127 | ± | 50.1 |
| Tmax | h | 3.00 | ± | 0.816 | 2.50 | ± | 1.73 | |
| AUC0-t | ng·h/mL | 641 | ± | 148 | 645 | ± | 120 | |
| AUC0-∞ | ng·h/mL | 645 | ± | 148 | 657 | ± | 122 | |
| t1/2 | h | 1.16 | ± | 0.171 | 1.46 | ± | 0.269 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.; Yang, F.; Yuan, Y.; Li, C.; Huo, X.; Liu, J.; Zhou, S.; Li, W.; Zhang, N.; Liu, J.; et al. OAT3 Participates in Drug–Drug Interaction between Bentysrepinine and Entecavir through Interactions with M8—A Metabolite of Bentysrepinine—In Rats and Humans In Vitro. Molecules 2023, 28, 1995. https://doi.org/10.3390/molecules28041995
Zhang A, Yang F, Yuan Y, Li C, Huo X, Liu J, Zhou S, Li W, Zhang N, Liu J, et al. OAT3 Participates in Drug–Drug Interaction between Bentysrepinine and Entecavir through Interactions with M8—A Metabolite of Bentysrepinine—In Rats and Humans In Vitro. Molecules. 2023; 28(4):1995. https://doi.org/10.3390/molecules28041995
Chicago/Turabian StyleZhang, Aijie, Fanlong Yang, Yang Yuan, Cai Li, Xiaokui Huo, Jing Liu, Shenzhi Zhou, Wei Li, Na Zhang, Jianfeng Liu, and et al. 2023. "OAT3 Participates in Drug–Drug Interaction between Bentysrepinine and Entecavir through Interactions with M8—A Metabolite of Bentysrepinine—In Rats and Humans In Vitro" Molecules 28, no. 4: 1995. https://doi.org/10.3390/molecules28041995