

Beneficial Effects of Phytogenic Feed Additives on Epithelial Barrier Integrity in an In Vitro Co-Culture Model of the Piglet Gut

Abstract
:1. Introduction
2. Results
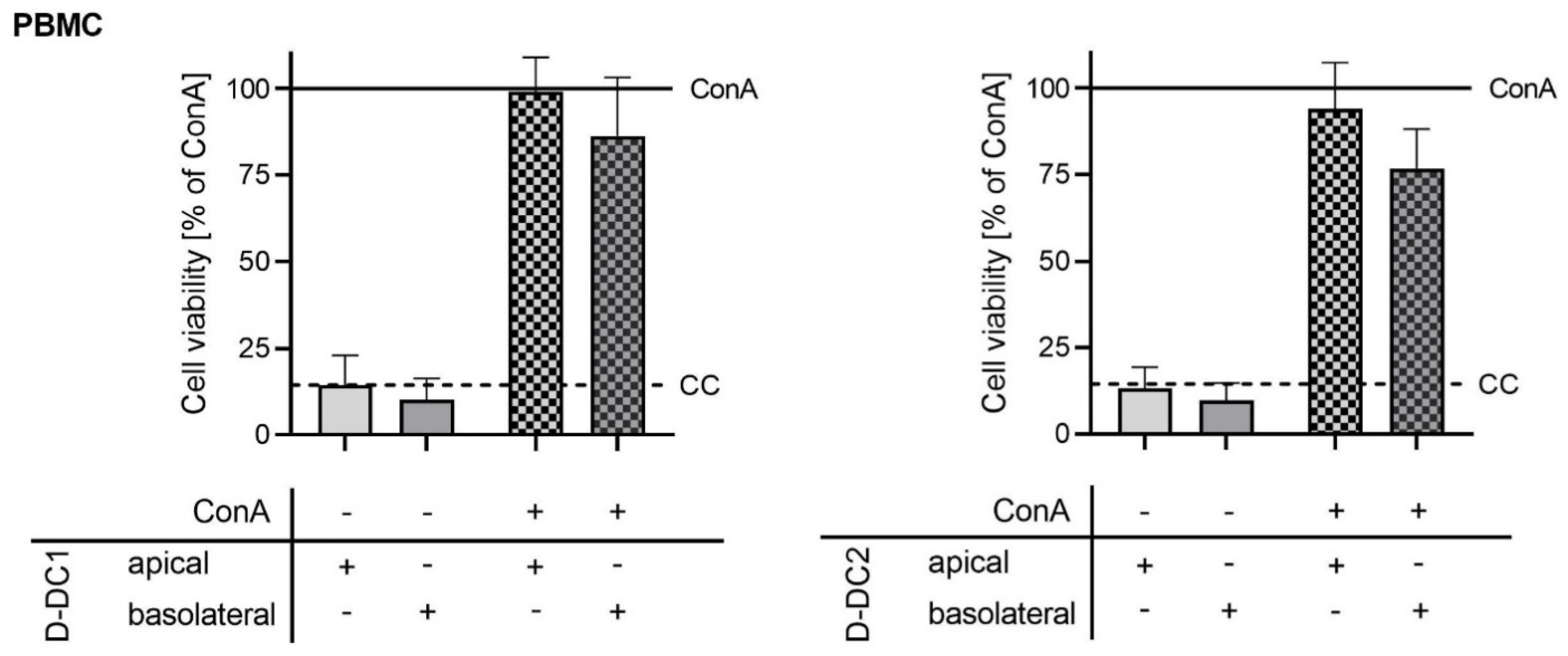
2.1. Co-Culture Model—Barrier Integrity
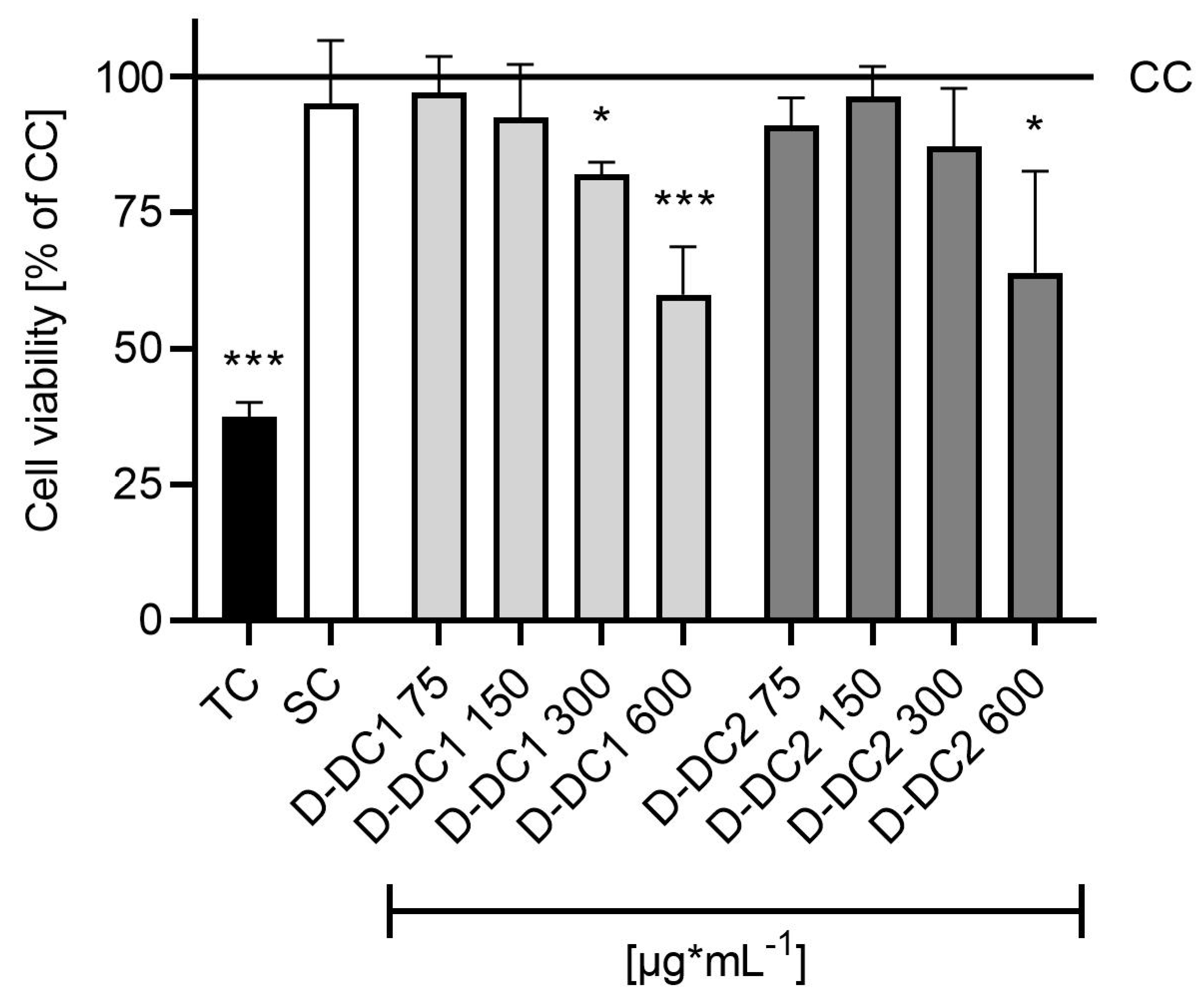
2.2. Co-Culture Model—Cell Viability
3. Discussion
3.1. Effects of Phytogenic Products, Apical or Basolateral Application, and Treatment Time on the In Vitro Co-Culture Model
3.2. Contribution of Phytogenic Components to Observed Effects
3.3. Outlook and Future Use of the Co-Culture Model
4. Materials and Methods
4.1. Routine Maintenance of IPEC-J2 Cultures
4.2. Blood Sampling and Isolation of PBMC
4.3. Co-Culture Model of the Piglet Gut
4.4. Stimulation of the Co-Culture Model
4.5. Cell Viability Testing and Supernatant Samples
4.6. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Cytotoxicity Pre-Trial
Appendix B. Results of Repeated Measures Two-Way ANOVA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | % of Total Variation | p-Value |
|---|---|---|
| Treatment × time | 5.390 | <0.0001 |
| Treatment | 79.09 | <0.0001 |
| Time | 4.592 | <0.0001 |
| Trial | 6.039 | 0.0014 |
References
- Rioja-Lang, F.C.; Connor, M.; Bacon, H.J.; Lawrence, A.B.; Dwyer, C.M. Prioritization of Farm Animal Welfare Issues Using Expert Consensus. Front. Vet. Sci. 2020, 6, 495. [Google Scholar] [CrossRef]
- Liu, Y. Fatty Acids, Inflammation and Intestinal Health in Pigs. J. Anim. Sci. Biotechnol. 2015, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Pluske, J.R.; Turpin, D.L.; Kim, J.C. Gastrointestinal Tract (Gut) Health in the Young Pig. Anim. Nutr. 2018, 4, 187–196. [Google Scholar] [CrossRef]
- Knecht, D.; Cholewińska, P.; Jankowska-Mąkosa, A.; Czyż, K. Development of Swine’s Digestive Tract Microbiota and Its Relation to Production Indices-a Review. Animals 2020, 10, 527. [Google Scholar] [CrossRef] [Green Version]
- Karásková, K.; Suchý, P.; Straková, E. Current Use of Phytogenic Feed Additives in Animal Nutrition: A Review. Czech J. Anim. Sci. 2015, 60, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Stevanović, Z.D.; Bošnjak-Neumüller, J.; Pajić-Lijaković, I.; Raj, J.; Vasiljević, M. Essential Oils as Feed Additives—Future Perspectives. Molecules 2018, 23, 1717. [Google Scholar] [CrossRef] [Green Version]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential Oils as Alternatives to Antibiotics in Swine Production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef]
- Valenzuela-Grijalva, N.V.; Pinelli-Saavedra, A.; Muhlia-Almazan, A.; Domínguez-Díaz, D.; González-Ríos, H. Dietary Inclusion Effects of Phytochemicals as Growth Promoters in Animal Production. J. Anim. Sci. Technol. 2017, 59, 8. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.K.; Xue, H.X.; Zhou, Z.X.; Peng, J. A Carvacrol-Thymol Blend Decreased Intestinal Oxidative Stress and Influenced Selected Microbes without Changing the Messenger RNA Levels of Tight Junction Proteins in Jejunal Mucosa of Weaning Piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Xiang, Q.; Wang, J.; Peng, J.; Wei, H. Oregano Essential Oil Improves Intestinal Morphology and Expression of Tight Junction Proteins Associated with Modulation of Selected Intestinal Bacteria and Immune Status in a Pig Model. Biomed. Res. Int. 2016, 2016, 5436738. [Google Scholar] [CrossRef]
- Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential Oil and Aromatic Plants as Feed Additives in Non-Ruminant Nutrition: A Review. J. Anim. Sci. Biotechnol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Ayrle, H.; Mevissen, M.; Kaske, M.; Nathues, H.; Gruetzner, N.; Melzig, M.; Walkenhorst, M. Medicinal Plants—Prophylactic and Therapeutic Options for Gastrointestinal and Respiratory Diseases in Calves and Piglets? A Systematic Review. BMC Vet. Res. 2016, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.F.; Nyachoti, M. Using Probiotics to Improve Swine Gut Health and Nutrient Utilization. Anim. Nutr. 2017, 3, 331–343. [Google Scholar] [CrossRef]
- Shin, D.; Chang, S.Y.; Bogere, P.; Won, K.H.; Choi, J.Y.; Choi, Y.J.; Lee, H.K.; Hur, J.; Park, B.Y.; Kim, Y.; et al. Beneficial Roles of Probiotics on the Modulation of Gut Microbiota and Immune Response in Pigs. PLoS ONE 2019, 14, e0220843. [Google Scholar] [CrossRef] [Green Version]
- Rajković, E.; Schwarz, C.; Tischler, D.; Schedle, K.; Reisinger, N.; Emsenhuber, C.; Ocelova, V.; Roth, N.; Frieten, D.; Dusel, G.; et al. Potential of Grape Extract in Comparison with Therapeutic Dosage of Antibiotics in Weaning Piglets: Effects on Performance, Digestibility and Microbial Metabolites of the Ileum and Colon. Animals 2021, 11, 2771. [Google Scholar] [CrossRef]
- Rajković, E.; Schwarz, C.; Kapsamer, S.B.; Schedle, K.; Reisinger, N.; Emsenhuber, C.; Ocelova, V.; Roth, N.; Frieten, D.; Dusel, G.; et al. Evaluation of a Dietary Grape Extract on Oxidative Status, Intestinal Morphology, Plasma Acute-Phase Proteins and Inflammation Parameters of Weaning Piglets at Various Points of Time. Antioxidants 2022, 11, 1428. [Google Scholar] [CrossRef]
- Wang, T.; Yao, W.; Li, J.; Shao, Y.; He, Q.; Xia, J.; Huang, F. Dietary Garcinol Supplementation Improves Diarrhea and Intestinal Barrier Function Associated with Its Modulation of Gut Microbiota in Weaned Piglets. J. Anim. Sci. Biotechnol. 2020, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Chen, G.; Cao, G.; Xu, Y.; Wang, Y.; Yang, C. Effects of Clostridium Butyricum and Enterococcus Faecalis on Growth Performance, Intestinal Structure, and Inflammation in Lipopolysaccharide-Challenged Weaned Piglets. J. Anim. Sci. 2019, 97, 4140–4151. [Google Scholar] [CrossRef]
- Chen, J.; Yu, B.; Chen, D.; Huang, Z.; Mao, X.; Zheng, P.; Yu, J.; Luo, J.; He, J. Chlorogenic Acid Improves Intestinal Barrier Functions by Suppressing Mucosa Inflammation and Improving Antioxidant Capacity in Weaned Pigs. J. Nutr. Biochem. 2018, 59, 84–92. [Google Scholar] [CrossRef]
- Chang, S.Y.; Song, M.H.; Lee, J.H.; Oh, H.J.; Kim, Y.J.; An, J.W.; Go, Y.B.; Song, D.C.; Cho, H.A.; Cho, S.Y.; et al. Phytogenic Feed Additives Alleviate Pathogenic Escherichia Coli-Induced Intestinal Damage through Improving Barrier Integrity and Inhibiting Inflammation in Weaned Pigs. J. Anim. Sci. Biotechnol. 2022, 13, 107. [Google Scholar] [CrossRef]
- Zhai, H.; Liu, H.; Wang, S.; Wu, J.; Kluenter, A.M. Potential of Essential Oils for Poultry and Pigs. Anim. Nutr. 2018, 4, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Christensen, L.G.; Rasmussen, P.B.; Kragh, K.M. In Vitro Immunomodulatory Effects of Thymol and Cinnamaldehyde in a Pig Intestinal Epithelial Cell Line (IPEC-J2). J. Appl. Anim. Nutr. 2020, 8, 127–134. [Google Scholar] [CrossRef]
- Marks, H.; Grześkowiak, Ł.; Martinez-Vallespin, B.; Dietz, H.; Zentek, J. Porcine and Chicken Intestinal Epithelial Cell Models for Screening Phytogenic Feed Additives—Chances and Limitations in Use as Alternatives to Feeding Trials. Microorganisms 2022, 10, 629. [Google Scholar] [CrossRef] [PubMed]
- Caprarulo, V.; Turin, L.; Hejna, M.; Reggi, S.; Dell’Anno, M.; Riccaboni, P.; Trevisi, P.; Luise, D.; Baldi, A.; Rossi, L. Protective Effect of Phytogenic plus Short and Medium-Chain Fatty Acids-Based Additives in Enterotoxigenic Escherichia Coli Challenged Piglets. Vet. Res. Commun. 2022. [Google Scholar] [CrossRef]
- Hejna, M.; Kovanda, L.; Rossi, L.; Liu, Y. Mint Oils: In Vitro Ability to Perform Anti-Inflammatory, Antioxidant, and Antimicrobial Activities and to Enhance Intestinal Barrier Integrity. Antioxidants 2021, 10, 1004. [Google Scholar] [CrossRef]
- Murugesan, G.R.; Syed, B.; Haldar, S.; Pender, C. Phytogenic Feed Additives as an Alternative to Antibiotic Growth Promoters in Broiler Chickens. Front. Vet. Sci. 2015, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Van Der Klis, J.D.; Vinyeta-Punti, E. The Potential of Phytogenic Feed Additives in Pigs and Poultry. In Proceedings of the Congress of the European Society of Veterinary & Comparative Nutrition, Utrecht, The Netherlands, 11–13 September 2014. [Google Scholar]
- Ghiselli, F.; Rossi, B.; Piva, A.; Grilli, E. Assessing Intestinal Health. In Vitro and Ex Vivo Gut Barrier Models of Farm Animals: Benefits and Limitations. Front. Vet. Sci. 2021, 8, 723387. [Google Scholar] [CrossRef]
- Rahman, S.; Ghiboub, M.; Donkers, J.M.; van de Steeg, E.; van Tol, E.A.F.; Hakvoort, T.B.M.; de Jonge, W.J. The Progress of Intestinal Epithelial Models from Cell Lines to Gut-on-Chip. Int. J. Mol. Sci. 2021, 22, 13472. [Google Scholar] [CrossRef]
- Schierack, P.; Nordhoff, M.; Pollmann, M.; Weyrauch, K.D.; Amasheh, S.; Lodemann, U.; Jores, J.; Tachu, B.; Kleta, S.; Blikslager, A.; et al. Characterization of a Porcine Intestinal Epithelial Cell Line for in Vitro Studies of Microbial Pathogenesis in Swine. Histochem. Cell Biol. 2006, 125, 293–305. [Google Scholar] [CrossRef]
- Bachinger, D.; Mayer, E.; Kaschubek, T.; Schieder, C.; König, J.; Teichmann, K. Influence of Phytogenics on Recovery of the Barrier Function of Intestinal Porcine Epithelial Cells after a Calcium Switch. J. Anim. Physiol. Anim. Nutr. 2019, 103, 210–220. [Google Scholar] [CrossRef]
- Springler, A.; Hessenberger, S.; Schatzmayr, G.; Mayer, E. Early Activation of MAPK P44/42 Is Partially Involved in DON-Induced Disruption of the Intestinal Barrier Function and Tight Junction Network. Toxins 2016, 8, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duell, B.L.; Cripps, A.W.; Schembri, M.A.; Ulett, G.C. Epithelial Cell Coculture Models for Studying Infectious Diseases: Benefits and Limitations. J. Biomed. Biotechnol. 2011, 2011, 852419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CenciČ, A.; Langerholc, T. Functional Cell Models of the Gut and Their Applications in Food Microbiology—A Review. Int. J. Food Microbiol. 2010, 141, S4. [Google Scholar] [CrossRef] [PubMed]
- Ponce de León-Rodríguez, M.D.C.; Guyot, J.P.; Laurent-Babot, C. Intestinal in Vitro Cell Culture Models and Their Potential to Study the Effect of Food Components on Intestinal Inflammation. Crit. Rev. Food Sci. Nutr. 2019, 59, 3648–3666. [Google Scholar] [CrossRef]
- Kämpfer, A.A.M.; Urbán, P.; Gioria, S.; Kanase, N.; Stone, V.; Kinsner-Ovaskainen, A. Development of an in Vitro Co-Culture Model to Mimic the Human Intestine in Healthy and Diseased State. Toxicol. Vitr. 2017, 45, 31–43. [Google Scholar] [CrossRef]
- Gu, M.J.; Song, S.K.; Lee, I.K.; Ko, S.; Han, S.E.; Bae, S.; Ji, S.Y.; Park, B.C.; Song, K.D.; Lee, H.K.; et al. Barrier Protection via Toll-like Receptor 2 Signaling in Porcine Intestinal Epithelial Cells Damaged by Deoxynivalnol. Vet. Res. 2016, 47, 25. [Google Scholar] [CrossRef] [Green Version]
- Gu, M.J.; Han, S.E.; Hwang, K.; Mayer, E.; Reisinger, N.; Schatzmayr, D.; Park, B.C.; Han, S.H.; Yun, C.H. Hydrolyzed Fumonisin B 1 Induces Less Inflammatory Responses than Fumonisin B 1 in the Co-Culture Model of Porcine Intestinal Epithelial and Immune Cells. Toxicol. Lett. 2019, 305, 110–116. [Google Scholar] [CrossRef]
- Saleri, R.; Borghetti, P.; Ravanetti, F.; Andrani, M.; Cavalli, V.; De Angelis, E.; Ferrari, L.; Martelli, P. A Co-Culture Model of IPEC-J2 and Swine PBMC to Study the Responsiveness of Intestinal Epithelial Cells: The Regulatory Effect of Arginine Deprivation. Animals 2021, 11, 2756. [Google Scholar] [CrossRef]
- Schott, T.; Reisinger, N.; Teichmann, K.; König, J.; Ladinig, A.; Mayer, E. Establishment of an in Vitro Co-Culture Model of the Piglet Gut to Study Inflammatory Response and Barrier Integrity. Planta Med. 2022, 88, 262–273. [Google Scholar] [CrossRef]
- Flecknell, P. Replacement, Reduction and Refinement*. ALTEX-Altern. Anim. Exp. 2002, 19, 73–78. [Google Scholar]
- Tannenbaum, J.; Bennett, B.T. Russell and Burch’s 3Rs Then and Now: The Need for Clarity in Definition and Purpose. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 120–132. [Google Scholar] [PubMed]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Special Edition; Universities Federation for Animal Welfare: Hertfordshire, UK, 1959. [Google Scholar]
- Kaschubek, T.; Mayer, E.; Rzesnik, S.; Grenier, B.; Bachinger, D.; Schieder, C.; König, J.; Teichmann, K. Effects of Phytogenic Feed Additives on Cellular Oxidative Stress and Inflammatory Reactions in Intestinal Porcine Epithelial Cells. J. Anim. Sci. 2018, 96, 3657–3669. [Google Scholar] [CrossRef] [PubMed]
- Katayama, M.; Fukuda, T.; Okamura, T.; Suzuki, E.; Tamura, K.; Shimizu, Y.; Suda, Y.; Suzuki, K. Effect of Dietary Addition of Seaweed and Licorice on the Immune Performance of Pigs. Anim. Sci. J. 2011, 82, 274–281. [Google Scholar] [CrossRef] [PubMed]
- You, T.; Tang, J.; Yin, S.; Jia, G.; Liu, G.; Tian, G.; Chen, X.; Cai, J.; Kang, B.; Zhao, H. Effect of Dietary Licorice Flavonoids Powder on Performance, Intestinal Immunity and Health of Weaned Piglets. J. Anim. Physiol. Anim. Nutr. 2022, 107, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Caro-Maldonado, A.; Muñoz-Pinedo, C. Dying for Something to Eat: How Cells Respond to Starvation. Open Cell Signal. J. 2011, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Kamiloglu, S.; Sari, G.; Ozdal, T.; Capanoglu, E. Guidelines for Cell Viability Assays. Food Front. 2020, 1, 332–349. [Google Scholar] [CrossRef]
- Vatzia, E.; Pierron, A.; Saalmüller, A.; Mayer, E.; Gerner, W. Deoxynivalenol Affects Proliferation and Expression of Activation-Related Molecules in Major Porcine T-Cell Subsets. Toxins 2019, 11, 644. [Google Scholar] [CrossRef] [Green Version]
- Katial, R.K.; Sachanandani, D.; Pinney, C.; Lieberman, M.M. Cytokine Production in Cell Culture by Peripheral Blood Mononuclear Cells from Immunocompetent Hosts. Clin. Diagn. Lab. Immunol. 1998, 5, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Goyarts, T.; Dänicke, S.; Grove, N.; Tiemann, U.; Rothkötter, H.-J. Institute of Animal Nutrition Methodical Aspects of in Vitro Proliferation of Porcine Blood Lymphocytes When Exposed to Deoxynivalenol (DON). Landbauforsch. Volkenrode 2006, 56, 139–148. [Google Scholar]
- Ali, A.; Tan, H.Y.; Kaiko, G.E. Role of the Intestinal Epithelium and Its Interaction With the Microbiota in Food Allergy. Front. Immunol. 2020, 11, 604054. [Google Scholar] [CrossRef]
- Giovagnoni, G.; Rossi, B.; Tugnoli, B.; Ghiselli, F.; Bonetti, A.; Piva, A.; Grilli, E. Thymol and Carvacrol Downregulate the Expression of Salmonella Typhimurium Virulence Genes during an in Vitro Infection on Caco-2 Cells. Microorganisms 2020, 8, 862. [Google Scholar] [CrossRef] [PubMed]
- Putaala, H.; Nurminen, P.; Tiihonen, K. Effects of Cinnamaldehyde and Thymol on Cytotoxicity, Tight Junction Barrier Resistance, and Cyclooxygenase-1 and -2 Expression in Caco-2 Cells. J. Anim. Feed Sci. 2017, 26, 274–284. [Google Scholar] [CrossRef]
- Pu, J.; Chen, D.; Tian, G.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Huang, Z.; Zhu, L.; Luo, J.; et al. Protective Effects of Benzoic Acid, Bacillus Coagulans, and Oregano Oil on Intestinal Injury Caused by Enterotoxigenic Escherichia Coli in Weaned Piglets. Biomed Res. Int. 2018, 2018, 1829632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, E.; Wang, W.; Gan, L.; Li, Z.; Guo, S.; Guo, Y. Effects of Thymol and Carvacrol Supplementation on Intestinal Integrity and Immune Responses of Broiler Chickens Challenged with Clostridium Perfringens. J. Anim. Sci. Biotechnol. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Du, E.; Yuan, J.; Gao, J.; Wang, Y.L.; Aggrey, S.E.; Guo, Y. Supplemental Thymol and Carvacrol Increases Ileum Lactobacillus Population and Reduces Effect of Necrotic Enteritis Caused by Clostridium Perfringes in Chickens. Sci. Rep. 2017, 7, 7334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memar, M.Y.; Raei, P.; Alizadeh, N.; Aghdam, M.A.; Kafil, H.S. Carvacrol and Thymol: Strong Antimicrobial Agents against Resistant Isolates. Rev. Res. Med. Microbiol. 2017, 28, 63–68. [Google Scholar] [CrossRef]
- Rúa, J.; del Valle, P.; de Arriaga, D.; Fernández-Álvarez, L.; García-Armesto, M.R. Combination of Carvacrol and Thymol: Antimicrobial Activity Against Staphylococcus Aureus and Antioxidant Activity. Foodborne Pathog. Dis. 2019, 16, 622–629. [Google Scholar] [CrossRef]
- Lambert, R.J.W.; Skandamis, P.N.; Coote, P.J.; Nychas, G.-J.E. A Study of the Minimum Inhibitory Concentration and Mode of Action of Oregano Essential Oil, Thymol and Carvacrol. J. Appl. Microbiol. 2001, 91, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Bounar, R.; Krimat, S.; Boureghda, H.; Dob, T. Chemical Analyses, Antioxidant and Antifungal Effects of Oregano and Thyme Essential Oils Alone or in Combination against Selected Fusarium Species. Int. Food Res. J. 2020, 27, 66–77. [Google Scholar]
- Gholami-Ahangaran, M.; Ahmadi-Dastgerdi, A.; Azizi, S.; Basiratpour, A.; Zokaei, M.; Derakhshan, M. Thymol and Carvacrol Supplementation in Poultry Health and Performance. Vet. Med. Sci. 2022, 8, 267–288. [Google Scholar] [CrossRef]
- Abdel-Wareth, A.A.A.; Kehraus, S.; Südekum, K.H. Peppermint and Its Respective Active Component in Diets of Broiler Chickens: Growth Performance, Viability, Economics, Meat Physicochemical Properties, and Carcass Characteristics. Poult. Sci. 2019, 98, 3850–3859. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, R.; Sun, L.; Huang, C.; Wang, C.; Zhang, D.M.; Zhang, T.T.; Du, G.H. Anti-Inflammatory Activity of Methyl Salicylate Glycosides Isolated from Gaultheria Yunnanensis (Franch.) Rehder. Molecules 2011, 16, 3875–3884. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Ponnampalam, E.N.; Pushpakumara, G.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. Cinnamon: A Natural Feed Additive for Poultry Health and Production—A Review. Animals 2021, 11, 2026. [Google Scholar] [CrossRef] [PubMed]
- Abu Hafsa, S.H.; Ibrahim, S.A. Effect of Dietary Polyphenol-Rich Grape Seed on Growth Performance, Antioxidant Capacity and Ileal Microflora in Broiler Chicks. J. Anim. Physiol. Anim. Nutr. 2018, 102, 268–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of Polyphenol-Rich Grape by-Products in Monogastric Nutrition. A Review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Hasan, M.K.; Ara, I.; Mondal, M.S.A.; Kabir, Y. Phytochemistry, Pharmacological Activity, and Potential Health Benefits of Glycyrrhiza Glabra. Heliyon 2021, 7, e07240. [Google Scholar] [CrossRef]
- Federal Act on the Protection of Animals (Animal Protection Act—TSchG)—BGBl. I Nr. 118/2004; Federal Ministry of Agriculture, Forestry, Regions and Water Management: Vienna, Austria, 2004.



| Extract D-DC1 | Extract D-DC2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | ConA | api | baso | api + ConA | baso + ConA | api | baso | api + ConA | baso + ConA | ||
| 24 h | TEER [kOhm*cm2] | 12.73 b | 10.52 a | 12.49 b | 12.28 b | 11.08 | 11.19 | 12.71 b | 12.33 b | 10.66 a | 11.31 |
| % of CC | 100.0 | 82.7 | 98.3 | 96.6 | 87.2 | 88.1 | 99.9 | 97.0 | 83.9 | 88.8 | |
| p-value vs. CC | - | 0.0265 | 0.9943 | 0.9017 | 0.0807 | 0.1164 | 0.9999 | 0.9264 | 0.0347 | 0.2206 | |
| % of ConA | 121.0 | 100.0 | 118.8 | 116.8 | 105.4 | 106.4 | 120.9 | 117.2 | 101.4 | 107.5 | |
| p-value vs. ConA | 0.0265 | - | 0.0093 | 0.0185 | 0.5124 | 0.5497 | 0.0180 | 0.0139 | 0.9994 | 0.5787 | |
| 48 h | TEER [kOhm*cm2] | 12.63 b | 9.18 a | 12.29 b | 12.11 b | 10.03 a | 10.33 c | 12.46 b | 11.99 b | 9.66 a | 10.31 c |
| % of CC | 100.0 | 72.7 | 97.3 | 95.9 | 79.5 | 81.8 | 98.6 | 94.9 | 76.5 | 81.7 | |
| p-value vs. CC | - | 0.0001 | 0.8891 | 0.6587 | 0.0004 | 0.0007 | 0.9940 | 0.7701 | 0.0002 | 0.0010 | |
| % of ConA | 137.6 | 100.0 | 134.0 | 132.0 | 109.4 | 112.6 | 135.8 | 130.7 | 105.3 | 112.4 | |
| p-value vs. ConA | 0.0001 | - | 0.0017 | 0.0035 | 0.0507 | 0.0082 | 0.0007 | 0.0301 | 0.2197 | 0.0046 | |
| 72 h | TEER [kOhm*cm2] | 13.05 b | 9.10 a | 12.67 b | 12.43 b | 9.85 a | 9.98 a | 12.83 b | 12.29 b | 9.24 a | 9.81 a |
| % of CC | 100.0 | 69.8 | 97.1 | 95.2 | 75.6 | 76.5 | 98.3 | 94.1 | 70.9 | 75.2 | |
| p-value vs. CC | - | 0.0001 | 0.8359 | 0.2807 | 0.0002 | 0.0005 | 0.9625 | 0.6242 | 0.0008 | 0.0002 | |
| % of ConA | 143.4 | 100.0 | 139.3 | 136.6 | 108.3 | 109.7 | 141.0 | 135.1 | 101.6 | 107.8 | |
| p-value vs. ConA | 0.0001 | - | 0.0004 | 0.0002 | 0.1787 | 0.1014 | 0.0001 | 0.0115 | 0.9849 | 0.1961 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wendner, D.; Schott, T.; Mayer, E.; Teichmann, K. Beneficial Effects of Phytogenic Feed Additives on Epithelial Barrier Integrity in an In Vitro Co-Culture Model of the Piglet Gut. Molecules 2023, 28, 1026. https://doi.org/10.3390/molecules28031026
Wendner D, Schott T, Mayer E, Teichmann K. Beneficial Effects of Phytogenic Feed Additives on Epithelial Barrier Integrity in an In Vitro Co-Culture Model of the Piglet Gut. Molecules. 2023; 28(3):1026. https://doi.org/10.3390/molecules28031026
Chicago/Turabian StyleWendner, Dominik, Theresa Schott, Elisabeth Mayer, and Klaus Teichmann. 2023. "Beneficial Effects of Phytogenic Feed Additives on Epithelial Barrier Integrity in an In Vitro Co-Culture Model of the Piglet Gut" Molecules 28, no. 3: 1026. https://doi.org/10.3390/molecules28031026