
Healthy Properties of a New Formulation of Pomegranate-Peel Extract in Mice Suffering from Experimental Autoimmune Encephalomyelitis

 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
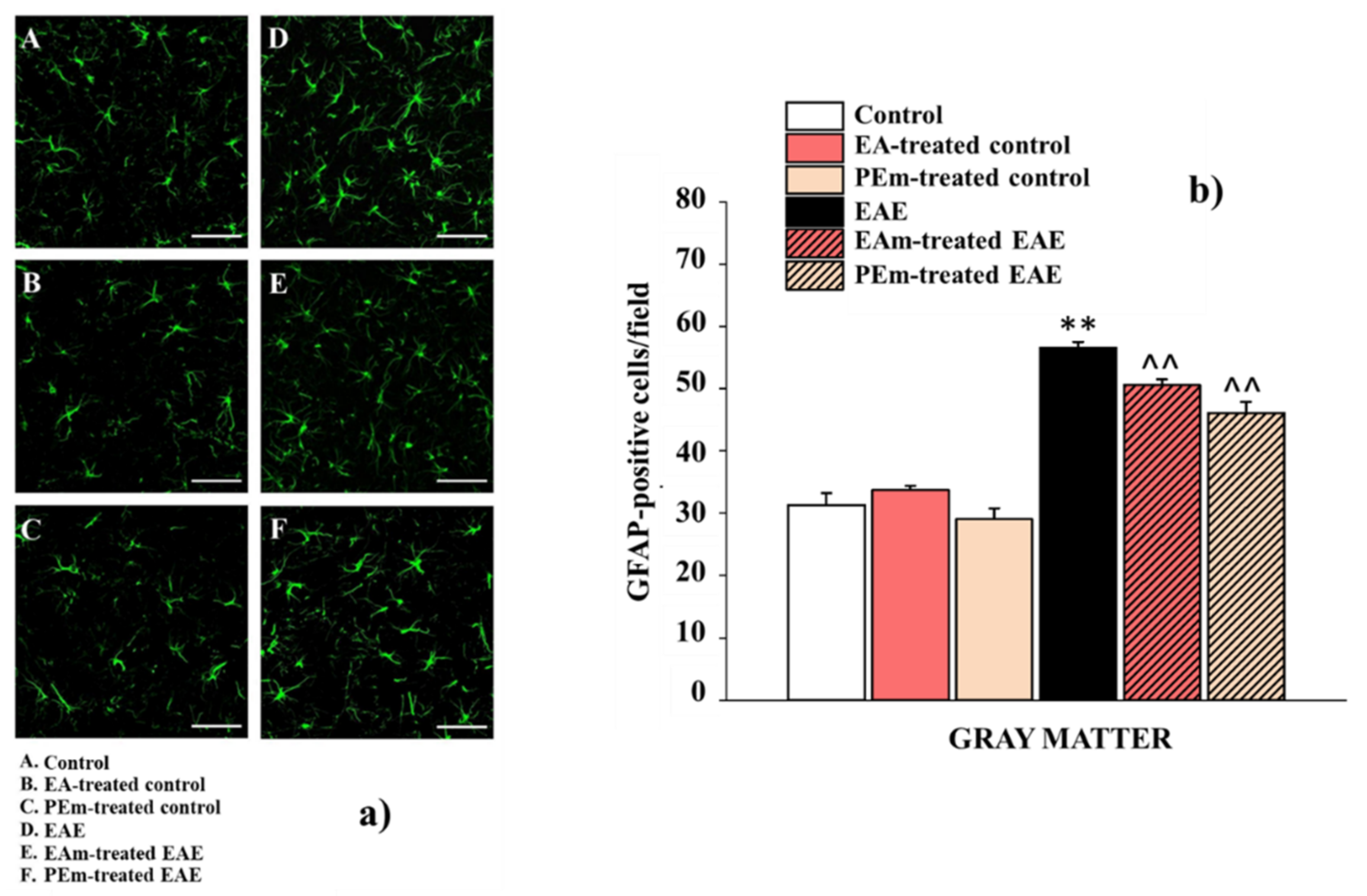
2. Results
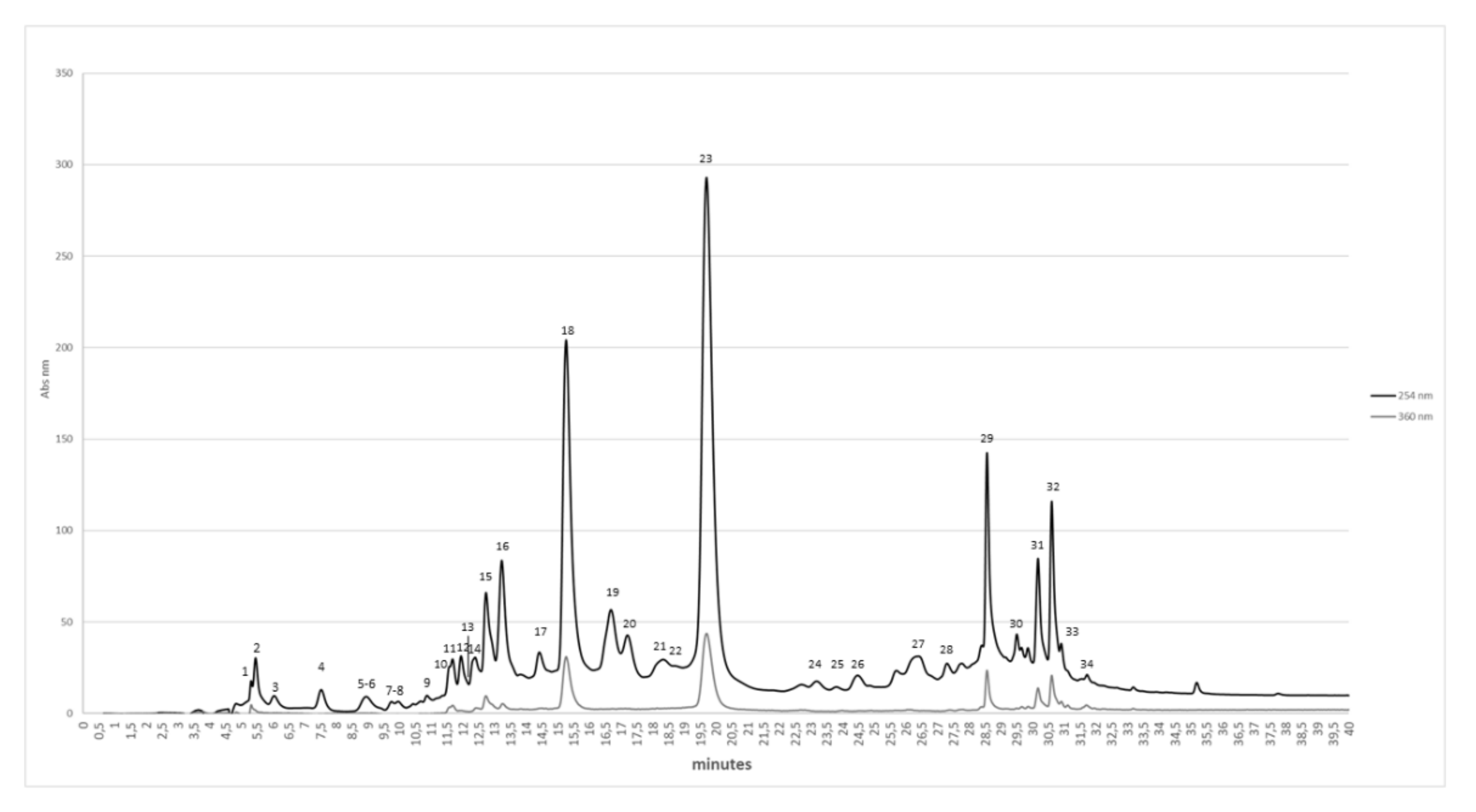
2.1. Chemical and Chromatographic Characterization of Pem
2.2. Pharmacological Activity of PEm and EAm in EAE Mice: “In Vivo” Effects
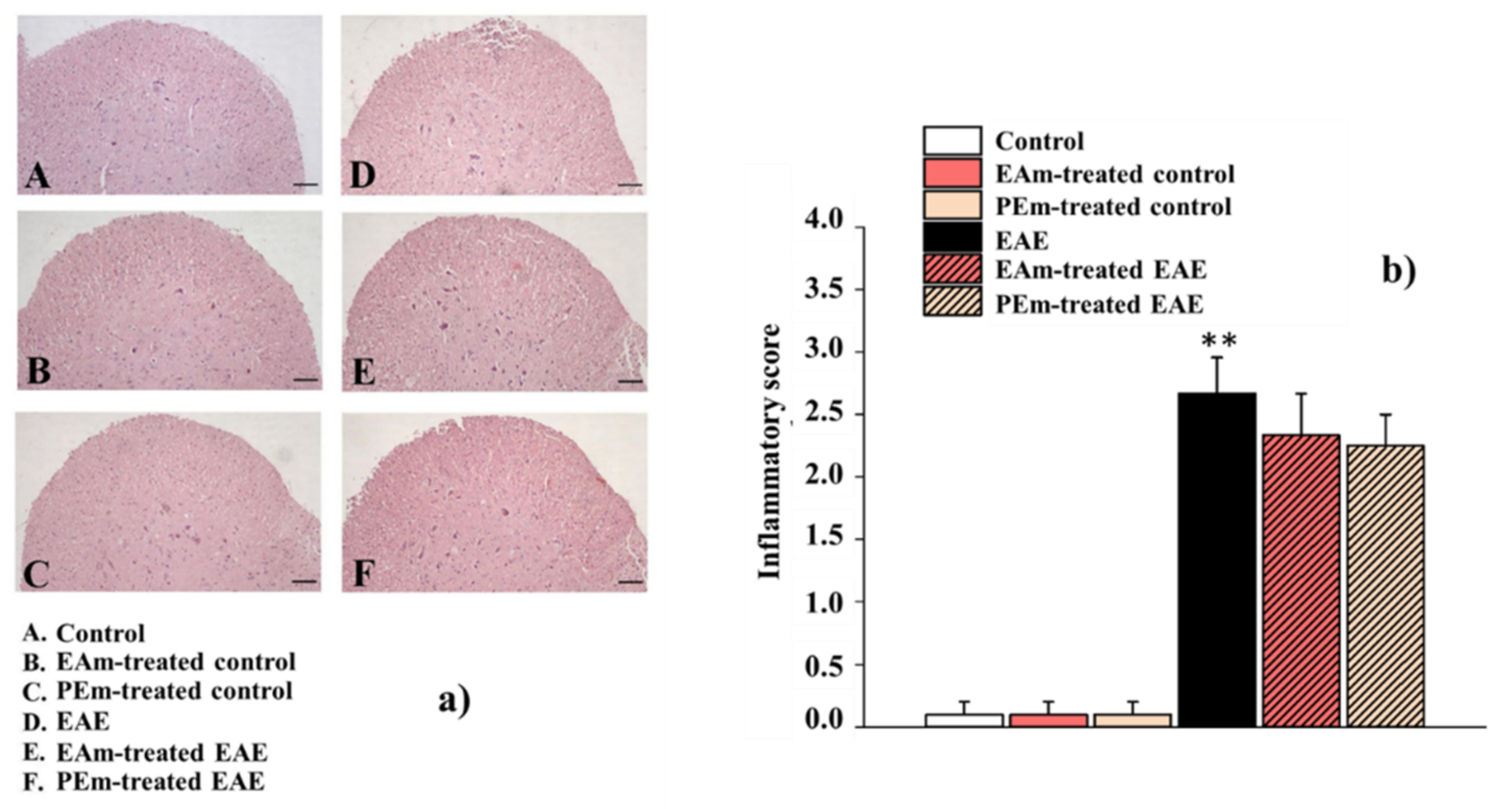
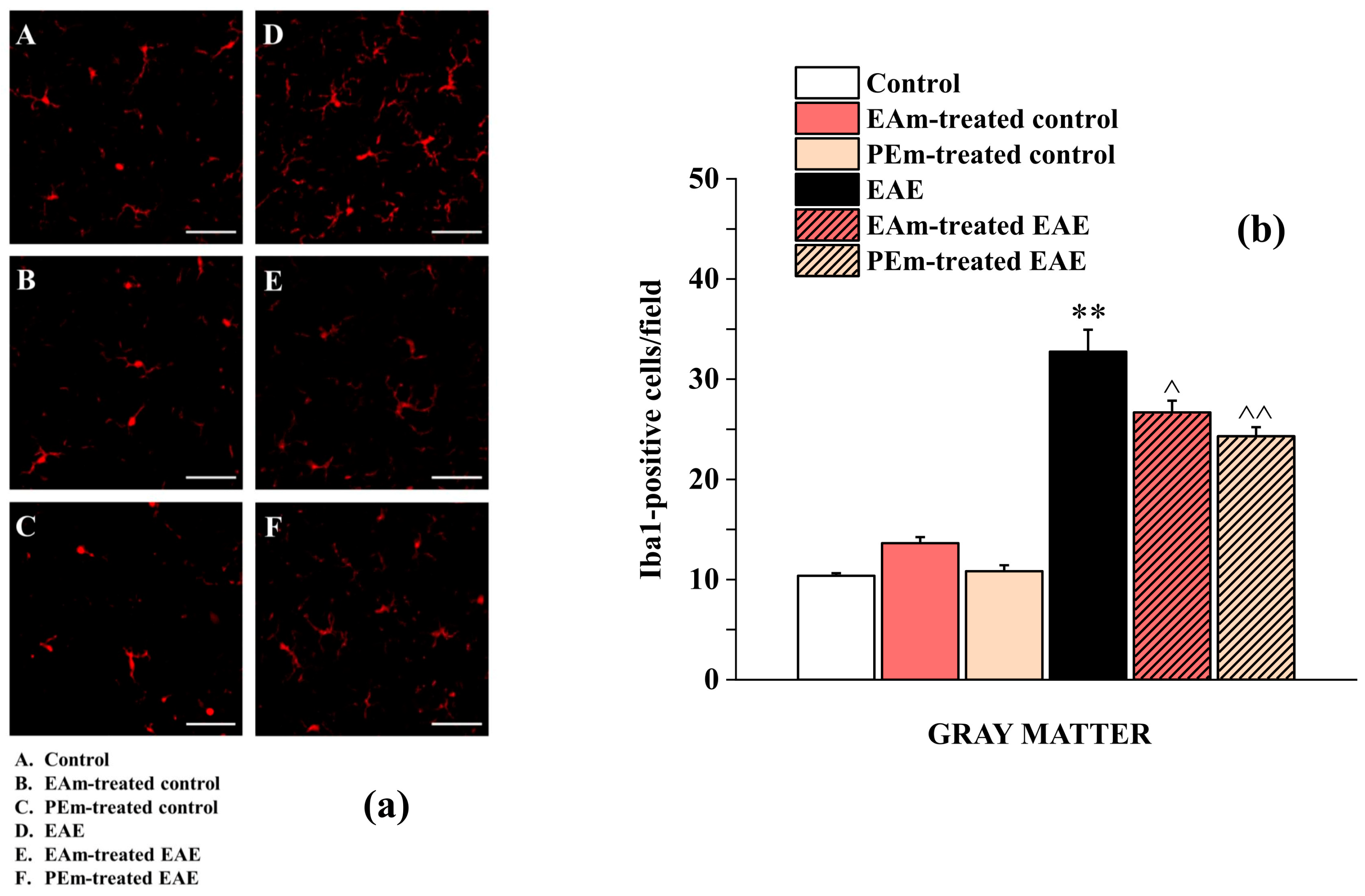
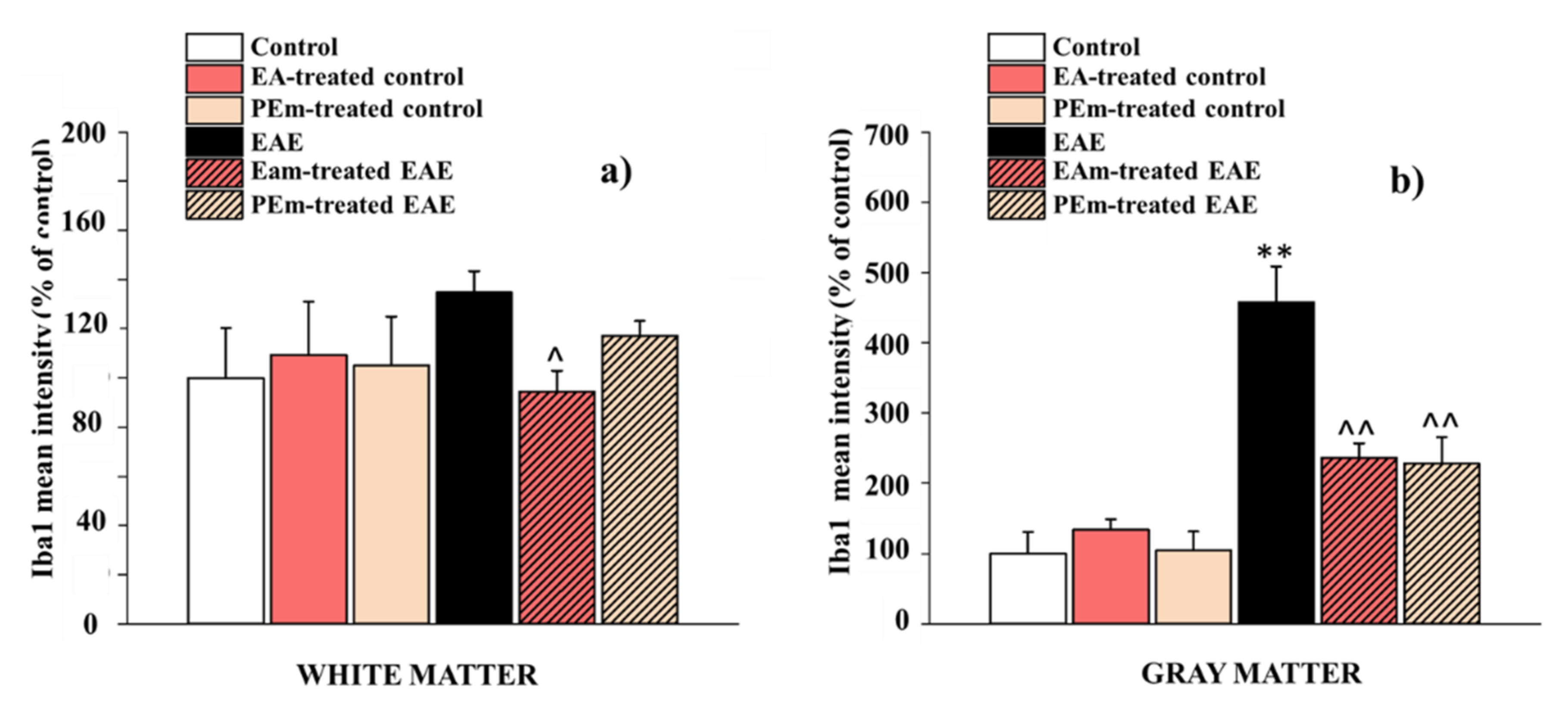
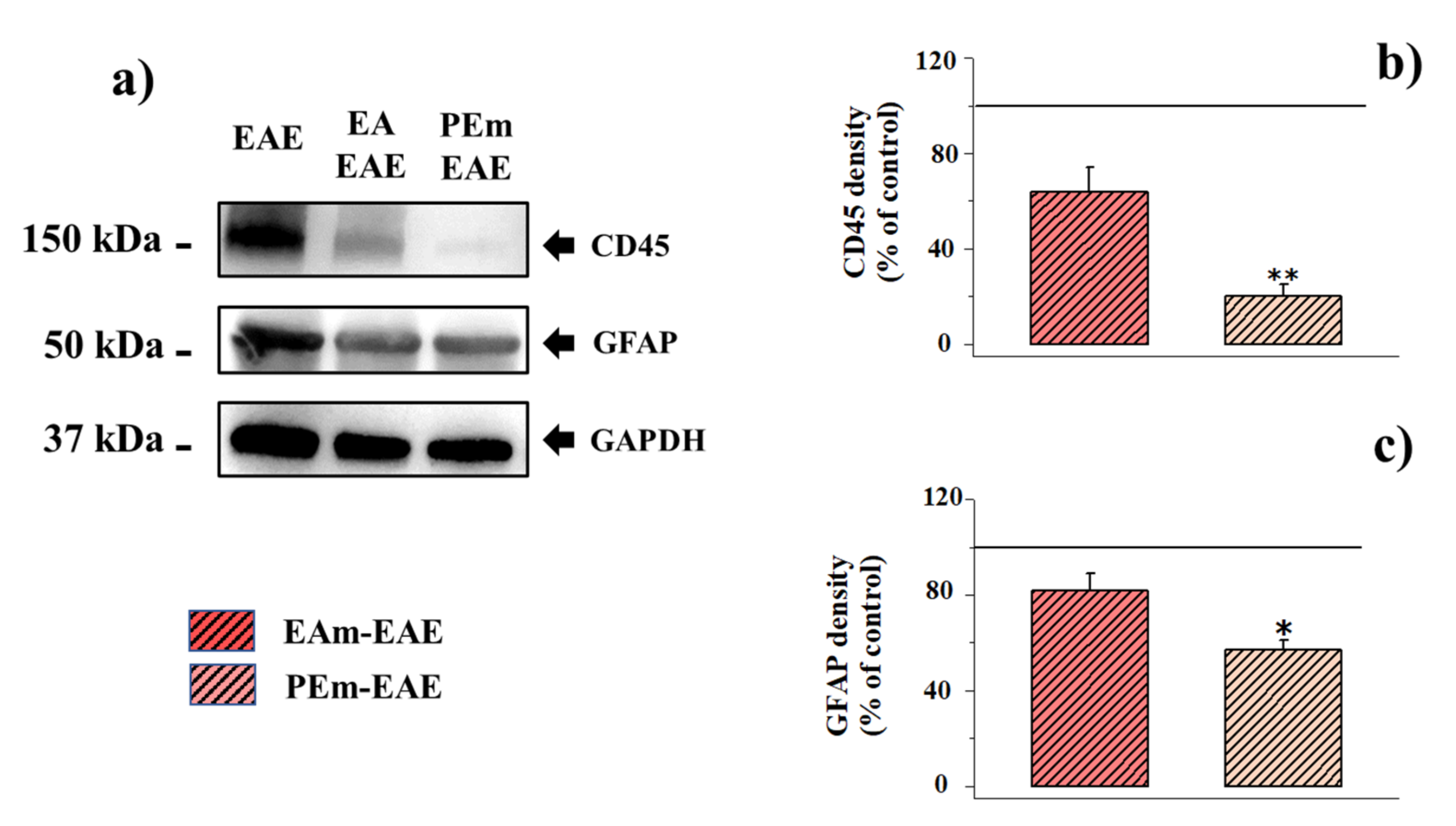
2.3. Pharmacological Activity of PEm Compared to EAm in EAE Mice: “Ex Vivo” Assessments
3. Discussion
4. Materials and Methods
4.1. Pomegranate Peels
4.2. Preparation of the Aqueous Peel Extract by Pulsed Ultrasound-Assisted Extraction (PUAE)
4.3. Formulation of the Aqueous Peel Extract by Spray-Drying Technology
4.4. Preliminary Characterization of the Aqueous Peel Extract Solid Microdispersion (PEm)
4.5. Chromatographic Characterization of the Pomegranate Phyto-Complex (PE and PEm): HPLC–DAD Analysis
4.6. Chromatographic Characterization of the Pomegranate Phyto-Complex (PE and PEm): HPLC–MS Analysis
4.7. Animals and EAE Induction
4.8. Animal Treatment
4.9. Western Blot Analysis
4.10. Histological Analysis
4.11. Immunohistochemistry
4.12. Calculation and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| MS | multiple sclerosis |
| EA | ellagic acid |
| ETs | ellagitannins |
| PE | pomegranate extract |
| PEm | pomegranate extract solid microdispersion |
| EAm | EA solid microdispersion |
| EAE | experimental autoimmune encephalomyelitis |
| PUAE | Pulsed Ultrasound-Assisted Extraction |
| LM | low methoxylated |
| d.p.i. | days post immunization |
| CNS | central nervous system |
| LFB | Luxol Fast Blue |
| H&E | Hematoxylin and Eosin |
| Iba1 | Ionized Calcium Binding Adaptor 1 |
| GFAP | glial fibrillar acid protein |
| RSA | radical scavenging activity |
References
- Farinotti, M.; Vacchi, L.; Simi, S.; Di Pietrantonj, C.; Brait, L.; Filippini, G. Dietary Interventions for Multiple Sclerosis. Cochrane Database Syst. Rev. 2012, 12, CD004192. [Google Scholar] [CrossRef] [PubMed]
- Boggia, R.; Turrini, F.; Villa, C.; Lacapra, C.; Zunin, P.; Parodi, B. Green Extraction from Pomegranate Marcs for the Production of Functional Foods and Cosmetics. Pharmaceuticals 2016, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Parisio, C.; Lucarini, E.; Micheli, L.; Toti, A.; Khatib, M.; Mulinacci, N.; Calosi, L.; Bani, D.; Di Cesare Mannelli, L.; Ghelardini, C. Pomegranate Mesocarp against Colitis-Induced Visceral Pain in Rats: Effects of a Decoction and Its Fractions. Int. J. Mol. Sci. 2020, 21, 4304. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Setzer, W.N.; Nabavi, S.F.; Orhan, I.E.; Braidy, N.; Sobarzo-Sanchez, E.; Nabavi, S.M. Insights Into Effects of Ellagic Acid on the Nervous System: A Mini Review. Curr. Pharm. Des. 2016, 22, 1350–1360. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Turrini, F.; Catena, S.; Zunin, P.; Grilli, M.; Pittaluga, A.M.; Boggia, R. Ellagic Acid a Multi-Target Bioactive Compound for Drug Discovery in CNS? A Narrative Review. Eur. J. Med. Chem. 2019, 183, 111724. [Google Scholar] [CrossRef]
- Ríos, J.-L.; Giner, R.M.; Marín, M.; Recio, M.C. A Pharmacological Update of Ellagic Acid. Planta Med. 2018, 84, 1068–1093. [Google Scholar] [CrossRef] [Green Version]
- Petrou, P.; Ginzberg, A.; Binyamin, O.; Karussis, D. Beneficial Effects of a Nano Formulation of Pomegranate Seed Oil, GranaGard, on the Cognitive Function of Multiple Sclerosis Patients. Mult. Scler. Relat. Disord. 2021, 54, 103103. [Google Scholar] [CrossRef]
- Lu, X.-Y.; Han, B.; Deng, X.; Deng, S.-Y.; Zhang, Y.-Y.; Shen, P.-X.; Hui, T.; Chen, R.-H.; Li, X.; Zhang, Y. Pomegranate Peel Extract Ameliorates the Severity of Experimental Autoimmune Encephalomyelitis via Modulation of Gut Microbiota. Gut Microbes 2020, 12, 1857515. [Google Scholar] [CrossRef]
- AlMatar, M.; Islam, M.R.; Albarri, O.; Var, I.; Koksal, F. Pomegranate as a Possible Treatment in Reducing Risk of Developing Wound Healing, Obesity, Neurodegenerative Disorders, and Diabetes Mellitus. Mini Rev. Med. Chem. 2018, 18, 507–526. [Google Scholar] [CrossRef]
- Akhtar, S.; Ismail, T.; Fraternale, D.; Sestili, P. Pomegranate Peel and Peel Extracts: Chemistry and Food Features. Food Chem. 2015, 174, 417–425. [Google Scholar] [CrossRef]
- Torres-León, C.; Ramírez-Guzman, N.; Londoño-Hernandez, L.; Martinez-Medina, G.A.; Díaz-Herrera, R.; Navarro-Macias, V.; Alvarez-Pérez, O.B.; Picazo, B.; Villarreal-Vázquez, M.; Ascacio-Valdes, J.; et al. Food Waste and Byproducts: An Opportunity to Minimize Malnutrition and Hunger in Developing Countries. Front. Sustain. Food Syst. 2018, 2, 52. [Google Scholar] [CrossRef]
- Turrini, F.; Zunin, P.; Catena, S.; Villa, C.; Alfei, S.; Boggia, R. Traditional or Hydro-Diffusion and Gravity Microwave Coupled with Ultrasound as Green Technologies for the Valorization of Pomegranate External Peels. Food Bioprod. Process. 2019, 117, 30–37. [Google Scholar] [CrossRef]
- Turrini, F.; Malaspina, P.; Giordani, P.; Catena, S.; Zunin, P.; Boggia, R. Traditional Decoction and PUAE Aqueous Extracts of Pomegranate Peels as Potential Low-Cost Anti-Tyrosinase Ingredients. Appl. Sci. 2020, 10, 2795. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the Rescue of “Old” Metabolites to Understand a “New” Concept: Metabotypes as a Nexus among Phenolic Metabolism, Microbiota Dysbiosis, and Host Health Status. Mol. Nutr. Food Res. 2017, 61, 1500901. [Google Scholar] [CrossRef]
- García-Villalba, R.; Giménez-Bastida, J.A.; Ávila-Gálvez, M.A.; Tomás-Barberán, F.A.; Espín, J.C.; González-Sarrías, A. Ellagitannins and Their Gut Microbiota-Derived Metabolites: Urolithins. In Dietary Polyphenols; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 319–364. ISBN 978-1-119-56375-4. [Google Scholar]
- Zuccari, G.; Baldassari, S.; Ailuno, G.; Turrini, F.; Alfei, S.; Caviglioli, G. Formulation Strategies to Improve Oral Bioavailability of Ellagic Acid. Appl. Sci. 2020, 10, 3353. [Google Scholar] [CrossRef]
- Alfei, S.; Turrini, F.; Catena, S.; Zunin, P.; Parodi, B.; Zuccari, G.; Pittaluga, A.M.; Boggia, R. Preparation of Ellagic Acid Micro and Nano Formulations with Amazingly Increased Water Solubility by Its Entrapment in Pectin or Non-PAMAM Dendrimers Suitable for Clinical Applications. New J. Chem. 2019, 43, 2438–2448. [Google Scholar] [CrossRef]
- Boggia, R.; Turrini, F.; Roggeri, A.; Olivero, G.; Cisani, F.; Bonfiglio, T.; Summa, M.; Grilli, M.; Caviglioli, G.; Alfei, S.; et al. Neuroinflammation in Aged Brain: Impact of the Oral Administration of Ellagic Acid Microdispersion. Int. J. Mol. Sci. 2020, 21, 3631. [Google Scholar] [CrossRef]
- Duncan, I.D.; Radcliff, A.B.; Heidari, M.; Kidd, G.; August, B.K.; Wierenga, L.A. The Adult Oligodendrocyte Can Participate in Remyelination. Proc. Natl. Acad. Sci. USA 2018, 115, E11807–E11816. [Google Scholar] [CrossRef] [Green Version]
- Chemat, F.; Rombaut, N.; Sicaire, A.-G.; Meullemiestre, A.; Fabiano-Tixier, A.-S.; Abert-Vian, M. Ultrasound Assisted Extraction of Food and Natural Products. Mechanisms, Techniques, Combinations, Protocols and Applications. A Review. Ultrason. Sonochem. 2017, 34, 540–560. [Google Scholar] [CrossRef]
- Chemat, F.; Vian, M.A.; Cravotto, G. Green Extraction of Natural Products: Concept and Principles. Int. J. Mol. Sci. 2012, 13, 8615–8627. [Google Scholar] [CrossRef] [Green Version]
- Turrini, F.; Boggia, R.; Leardi, R.; Borriello, M.; Zunin, P. Optimization of the Ultrasonic-Assisted Extraction of Phenolic Compounds from Oryza Sativa L. ‘Violet Nori’ and Determination of the Antioxidant Properties of Its Caryopses and Leaves. Molecules 2018, 23, 844. [Google Scholar] [CrossRef] [Green Version]
- Šavikin, K.; Živković, J.; Alimpić, A.; Zdunić, G.; Janković, T.; Duletić-Laušević, S.; Menković, N. Activity guided fractionation of pomegranate extract and its antioxidant, antidiabetic and antineurodegenerative properties. Ind. Crops Prod. 2018, 113, 142–149. [Google Scholar] [CrossRef]
- Khatib, M.; Giuliani, C.; Rossi, F.; Adessi, A.; Al-Tamimi, A.; Mazzola, G.; Di Gioia, D.; Innocenti, M.; Mulinacci, N. Polysaccharides from By-Products of the Wonderful and Laffan Pomegranate Varieties: New Insight into Extraction and Characterization. Food Chem. 2017, 235, 58–66. [Google Scholar] [CrossRef]
- Chaouch, M.A.; Benvenuti, S. The Role of Fruit By-Products as Bioactive Compounds for Intestinal Health. Foods 2020, 9, 1716. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.Y.; Choo, W.S.; Young, D.J.; Loh, X.J. Pectin as a Rheology Modifier: Origin, Structure, Commercial Production and Rheology. Carbohydr. Polym. 2017, 161, 118–139. [Google Scholar] [CrossRef] [PubMed]
- Dranca, F.; Oroian, M. Extraction, Purification and Characterization of Pectin from Alternative Sources with Potential Technological Applications. Food Res. Int. 2018, 113, 327–350. [Google Scholar] [CrossRef] [PubMed]
- Johns, T.G.; Kerlero de Rosbo, N.; Menon, K.K.; Abo, S.; Gonzales, M.F.; Bernard, C.C. Myelin Oligodendrocyte Glycoprotein Induces a Demyelinating Encephalomyelitis Resembling Multiple Sclerosis. J. Immunol. 1995, 154, 5536–5541. [Google Scholar]
- Mangiardi, M.; Crawford, D.K.; Xia, X.; Du, S.; Simon-Freeman, R.; Voskuhl, R.R.; Tiwari-Woodruff, S.K. An Animal Model of Cortical and Callosal Pathology in Multiple Sclerosis. Brain Pathol. 2011, 21, 263–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binyamin, O.; Larush, L.; Frid, K.; Keller, G.; Friedman-Levi, Y.; Ovadia, H.; Abramsky, O.; Magdassi, S.; Gabizon, R. Treatment of a Multiple Sclerosis Animal Model by a Novel Nanodrop Formulation of a Natural Antioxidant. Int. J. Nanomed. 2015, 10, 7165–7174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojanović, I.; Šavikin, K.; Đedović, N.; Živković, J.; Saksida, T.; Momčilović, M.; Koprivica, I.; Vujičić, M.; Stanisavljević, S.; Miljković, Đ.; et al. Pomegranate Peel Extract Ameliorates Autoimmunity in Animal Models of Multiple Sclerosis and Type 1 Diabetes. J. Funct. Foods 2017, 35, 522–530. [Google Scholar] [CrossRef]
- Kiasalari, Z.; Afshin-Majd, S.; Baluchnejadmojarad, T.; Azadi-Ahmadabadi, E.; Esmaeil-Jamaat, E.; Fahanik-Babaei, J.; Fakour, M.; Fereidouni, F.; Ghasemi-Tarie, R.; Jalalzade-Ogvar, S.; et al. Ellagic Acid Ameliorates Neuroinflammation and Demyelination in Experimental Autoimmune Encephalomyelitis: Involvement of NLRP3 and Pyroptosis. J. Chem. Neuroanat. 2021, 111, 101891. [Google Scholar] [CrossRef]
- Busto, R.; Serna, J.; Perianes-Cachero, A.; Quintana-Portillo, R.; García-Seisdedos, D.; Canfrán-Duque, A.; Paino, C.L.; Lerma, M.; Casado, M.E.; Martín-Hidalgo, A.; et al. Ellagic Acid Protects from Myelin-Associated Sphingolipid Loss in Experimental Autoimmune Encephalomyelitis. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2018, 1863, 958–967. [Google Scholar] [CrossRef]
- Cai, M. Fruit-Based Functional Food. In The Role of Alternative and Innovative Food Ingredients and Products in Consumer Wellness; Chapter 2; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 35–72. ISBN 978-0-12-816453-2. [Google Scholar]
- Turrini, F.; Boggia, R.; Donno, D.; Parodi, B.; Beccaro, G.; Baldassari, S.; Signorello, M.G.; Catena, S.; Alfei, S.; Zunin, P. From Pomegranate Marcs to a Potential Bioactive Ingredient: A Recycling Proposal for Pomegranate-Squeezed Marcs. Eur. Food Res. Technol. 2020, 246, 273–285. [Google Scholar] [CrossRef]
- Fischer, U.A.; Carle, R.; Kammerer, D.R. Identification and Quantification of Phenolic Compounds from Pomegranate (Punica granatum L.) Peel, Mesocarp, Aril and Differently Produced Juices by HPLC-DAD-ESI/MS(n). Food Chem. 2011, 127, 807–821. [Google Scholar] [CrossRef]
- Mena, P.; Calani, L.; Dall’Asta, C.; Galaverna, G.; García-Viguera, C.; Bruni, R.; Crozier, A.; Del Rio, D. Rapid and Comprehensive Evaluation of (Poly)Phenolic Compounds in Pomegranate (Punica granatum L.) Juice by UHPLC-MSn. Molecules 2012, 17, 14821–14840. [Google Scholar] [CrossRef] [Green Version]
- Abdulla, R.; Mansur, S.; Lai, H.; Ubul, A.; Sun, G.; Huang, G.; Aisa, H.A. Qualitative Analysis of Polyphenols in Macroporous Resin Pretreated Pomegranate Husk Extract by HPLC-QTOF-MS. Phytochem. Anal. 2017, 28, 465–473. [Google Scholar] [CrossRef]
- Pender, M.P.; Tabi, Z.; Nguyen, K.B.; McCombe, P.A. The Proximal Peripheral Nervous System Is a Major Site of Demyelination in Experimental Autoimmune Encephalomyelitis Induced in the Lewis Rat by a Myelin Basic Protein-Specific T Cell Clone. Acta Neuropathol. 1995, 89, 527–531. [Google Scholar] [CrossRef]
- Mandolesi, G.; Gentile, A.; Musella, A.; Fresegna, D.; De Vito, F.; Bullitta, S.; Sepman, H.; Marfia, G.A.; Centonze, D. Synaptopathy Connects Inflammation and Neurodegeneration in Multiple Sclerosis. Nat. Rev. Neurol. 2015, 11, 711–724. [Google Scholar] [CrossRef]
- Acharjee, S.; Verbeek, M.; Gomez, C.D.; Bisht, K.; Lee, B.; Benoit, L.; Sharkey, K.A.; Benediktsson, A.; Tremblay, M.-E.; Pittman, Q.J. Reduced Microglial Activity and Enhanced Glutamate Transmission in the Basolateral Amygdala in Early CNS Autoimmunity. J. Neurosci. 2018, 38, 9019–9033. [Google Scholar] [CrossRef]
- Hammond, J.W.; Bellizzi, M.J.; Ware, C.; Qiu, W.Q.; Saminathan, P.; Li, H.; Luo, S.; Ma, S.A.; Li, Y.; Gelbard, H.A. Complement-Dependent Synapse Loss and Microgliosis in a Mouse Model of Multiple Sclerosis. Brain Behav. Immun. 2020, 87, 739–750. [Google Scholar] [CrossRef]
- Medina-Torres, N.; Ayora-Talavera, T.; Espinosa-Andrews, H.; Sánchez-Contreras, A.; Pacheco, N. Ultrasound Assisted Extraction for the Recovery of Phenolic Compounds from Vegetable Sources. Agronomy 2017, 7, 47. [Google Scholar] [CrossRef]
- Millo, G.; Juntavee, A.; Ratanathongkam, A.; Nualkaew, N.; Peerapattana, J.; Chatchiwiwattana, S. Antibacterial Inhibitory Effects of Punica Granatum Gel on Cariogenic Bacteria: An in Vitro Study. Int. J. Clin. Pediatr. Dent. 2017, 10, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Bonfiglio, T.; Olivero, G.; Vergassola, M.; Di Cesare Mannelli, L.; Pacini, A.; Iannuzzi, F.; Summa, M.; Bertorelli, R.; Feligioni, M.; Ghelardini, C.; et al. Environmental Training Is Beneficial to Clinical Symptoms and Cortical Presynaptic Defects in Mice Suffering from Experimental Autoimmune Encephalomyelitis. Neuropharmacology 2019, 145, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Marino, M.W.; Wong, G.; Grail, D.; Dunn, A.; Bettadapura, J.; Slavin, A.J.; Old, L.; Bernard, C.C. TNF Is a Potent Anti-Inflammatory Cytokine in Autoimmune-Mediated Demyelination. Nat. Med. 1998, 4, 78–83. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Determination | PEm |
|---|---|
| Moisture (%) | 13.67 ± 0.01 |
| Solubility (mg/mL) Bulk density | 50.00 ± 0.10 0.20 ± 0.05 |
| Radical scavenging activity (RSA: mg AEAC/g powder) | 537.43 ± 10.78 |
| SAMPLE | PUNICALAGIN | ELLAGIC ACID |
|---|---|---|
| μg/mL | ||
| PE | 692.2 ± 19.4 | 57.8 ± 2.9 |
| PEM | 103.4 ± 8.2 | 2.8 ± 0.3 |
| * EE %= 14.9 | ||
| Peak | RT (min) | [M-H]− m/z | MS/MS Fragments | Tentatively Identified Compound |
|---|---|---|---|---|
| 1 | 5.3 | 649 | 301,497 | Galloyl-HHDP-gluconate (lagerstannin C) isomer [30] |
| 2 | 5.4 | 481 | 301,275 | HHDP_hexoside [30,31] |
| 3 | 5.9 | 481 | 301,275 | HHDP_hexoside [30,31] |
| 4 | 7.5 | 481 | 301,275 | HHDP_hexoside [30,31] |
| 5 | 8.8 | 783 | 601,721,765,481,301 | bis-HHDP-hexoside (Pedunculagin I isomer) [29,30] |
| 6 | 8.9 | 649 | 301, 497 | Galloyl-HHDP-gluconate (lagerstannin C) isomer [30] |
| 7 | 9.7 | 735 | 675, 657, 603, 543, 381 | Unknown |
| 8 | 9.9 | 645 | 585, 567, 513, 453, 301 | Unknown |
| 9 | 10.8 | 633 | 301, 481, 275, 249 | Galloyl-HHDP-hexoside isomers [29,30] |
| 10 | 11.5 | 781 | 601.721 | Punicalin isomer [30] |
| 11 | 11.6 | 781 | 601.721 | Punicalin isomer [30] |
| 12 | 12 | 783 | 601,721,765,481,301 | bis-HHDP-hexoside (Pedunculagin I isomer) [29,30] |
| 13 | 12.2 | 649 | 301,497 | Galloyl-HHDP-gluconate (lagerstannin C) isomers [30] |
| 14 | 12.5 | 633 | 301,481,275,249 | Galloyl-HHDP-hexoside isomers [29,30] |
| 15 | 12.7 | 1083 | 601,781,721 | Punicalagin isomer [30] |
| 16 | 13.3 | 783 | 301,481,765 | Pedunculagin I isomer [29,30] |
| 17 | 14.4 | 951 | 907,783, 765,481 | HHDP-valoneoyl-glucoside [31] |
| 18 | 15.2 | 1083 | 601, 781, 721 | Punicalagin α [30] |
| 19 | 16.7 | 783 | 481, 301 | bis-HHDP-hexoside (Pedunculagin I isomer) [30] |
| 20 | 17.5 | 633 | 301, 481, 275, 249 | Galloyl-HHDP-hexoside isomers [29,30] |
| 21 | 18.3 | 783 | 481, 301 | bis-HHDP-hexoside (Pedunculagin I isomer) [30] |
| 22 | 19.0 | 785 | 301, 633, 483, 615 | Digalloyl-HHDP-hexoside (pedunculagin II) [29,31] |
| 23 | 19.6 | 1083 | 601, 781, 721 | Punicalagin β [30] |
| 24 | 23.2 | 801 | 649, 499, 349, 301 | Digalloyl-HHDP-glucoside (punigluconinisomer) [29] |
| 25 | 23.8 | 965 | 933 | Castalagin derivate [29] |
| 26 | 24.5 | 801 | 649, 499, 349, 301 | Digalloyl-HHDP-glucoside (punigluconinisomer) [29] |
| 27 | 26.3 | 785 | 483, 633, 301, 615 | Digalloyl-HHDP-hexoside (pedunculagin II) isomer [29] |
| 28 | 27.1 | 633 | 301, 463, 275, 245 | Corilagin [31] |
| 29 | 28.5 | 463 | 301 | Ellagic acid-glucoside [29] |
| 30 | 29.4 | 614 | 463 | Unknown |
| 31 | 30.2 | 447 | 301 | Ellagic acid-deoxyhexoside [29] |
| 32 | 30.6 | 433 | 301 | Ellagic acid-pentoside [29] |
| 33 | 31.0 | 593 | 285 | Kaempferol rutinoside [29] |
| 34 | 31.5 | 609 | 301 | Rutin [31] |
| CD45/GAPDH Ratio | % of Change | GFAP/GAPDH Ratio | % of Change | |
|---|---|---|---|---|
| Control | 0.083 ± 0.017 | -- | 4.04 ± 0.60 | -- |
| EA-treated control | 0.065 ± 0.023 | −22.87% | 3.43 ± 0.22 | −14.92% |
| PEm-treated control | 0.085 ± 0.022 | +2.23% | 4.20 ± 0.53 | +4.10% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vallarino, G.; Salis, A.; Lucarini, E.; Turrini, F.; Olivero, G.; Roggeri, A.; Damonte, G.; Boggia, R.; Di Cesare Mannelli, L.; Ghelardini, C.; et al. Healthy Properties of a New Formulation of Pomegranate-Peel Extract in Mice Suffering from Experimental Autoimmune Encephalomyelitis. Molecules 2022, 27, 914. https://doi.org/10.3390/molecules27030914
Vallarino G, Salis A, Lucarini E, Turrini F, Olivero G, Roggeri A, Damonte G, Boggia R, Di Cesare Mannelli L, Ghelardini C, et al. Healthy Properties of a New Formulation of Pomegranate-Peel Extract in Mice Suffering from Experimental Autoimmune Encephalomyelitis. Molecules. 2022; 27(3):914. https://doi.org/10.3390/molecules27030914
Chicago/Turabian StyleVallarino, Giulia, Annalisa Salis, Elena Lucarini, Federica Turrini, Guendalina Olivero, Alessandra Roggeri, Gianluca Damonte, Raffaella Boggia, Lorenzo Di Cesare Mannelli, Carla Ghelardini, and et al. 2022. "Healthy Properties of a New Formulation of Pomegranate-Peel Extract in Mice Suffering from Experimental Autoimmune Encephalomyelitis" Molecules 27, no. 3: 914. https://doi.org/10.3390/molecules27030914