
In Vitro Anticancer Activity of Methanolic Extract of Justicia adhatoda Leaves with Special Emphasis on Human Breast Cancer Cell Line

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Phytochemical Composition of Methanolic Extract of J. adhatoda
2.2. DPPH Radical Scavenging Assay
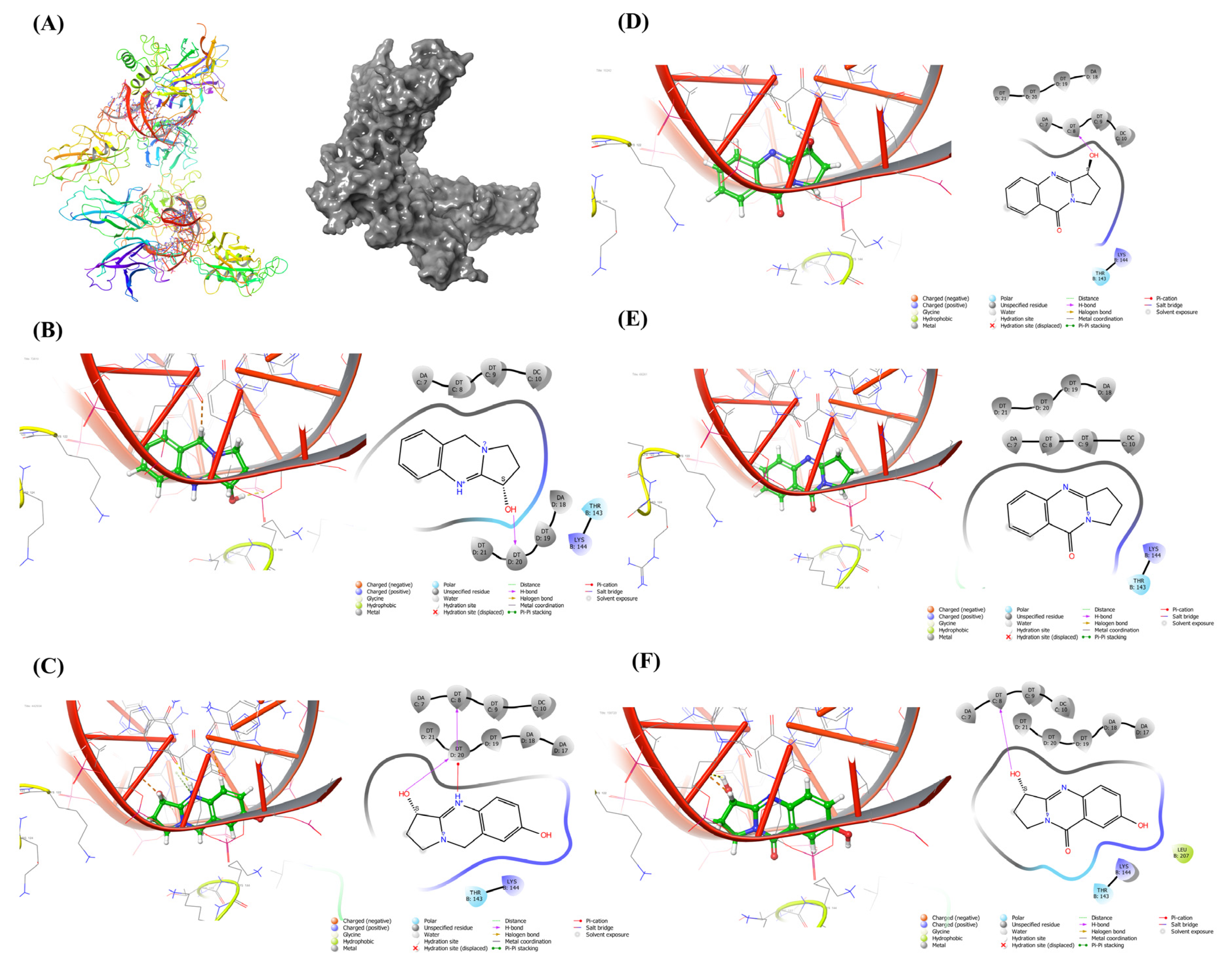
2.3. Docking Study
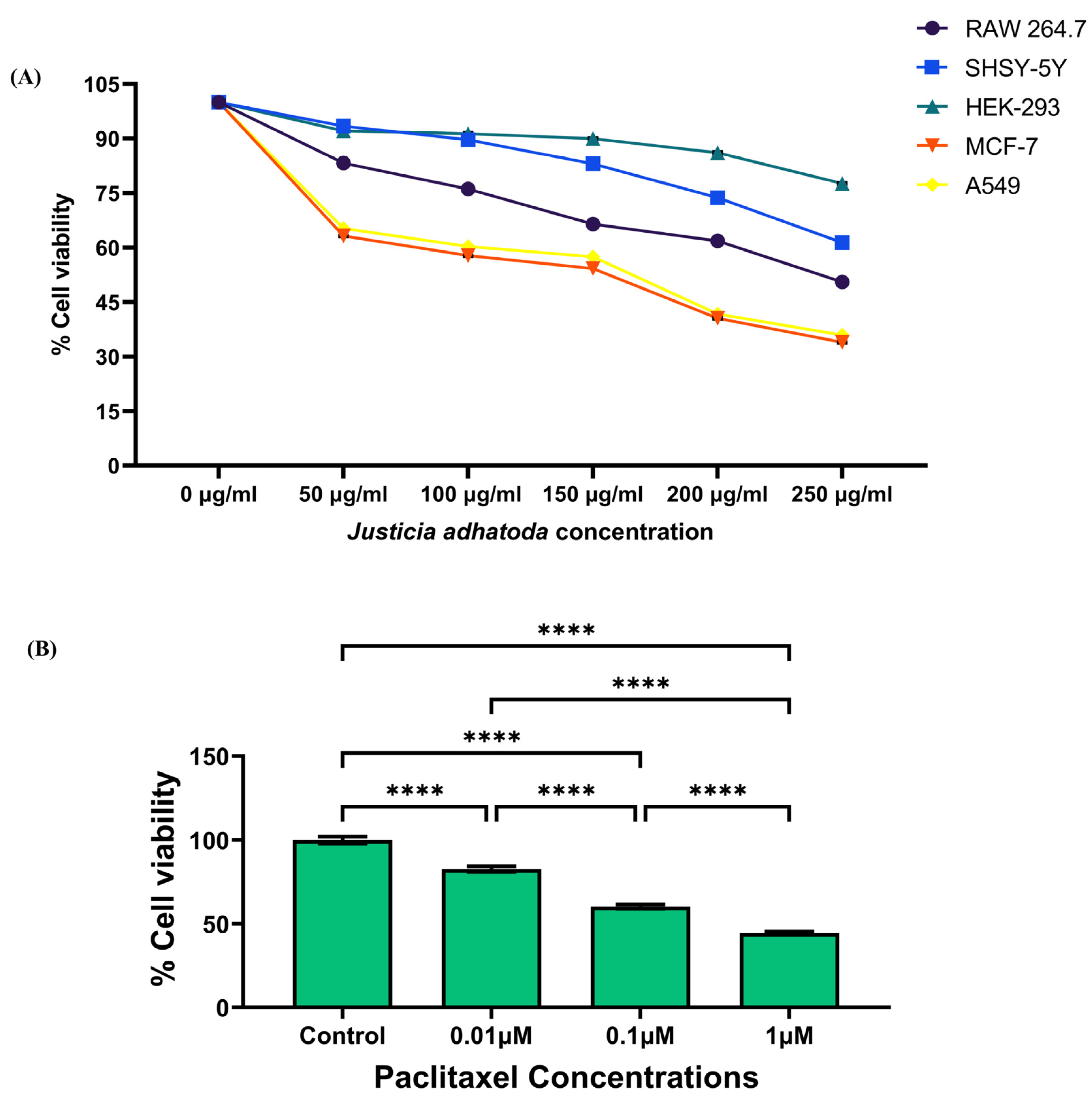
2.4. Evaluation of the Cytotoxicity of J. adhatoda Leaf Extract on Different Cell Lines
2.5. J. adhatoda Leaf Extract Induces NO Production in MCF-7 Cell Line
2.6. J. adhatoda Leaf Extract Induces ROS Production in MCF-7 Cell Line
2.7. Effect of J. adhatoda Leaf Extract on Mitochondrial Membrane Potential (ΔΨM)
2.8. Effect of J. adhatoda Leaf Extract on Glutathione (GSH) and Glutathione Disulfide (GSSG) Level
2.9. Effect of J. adhatoda Leaf Extract on Superoxide Dismutase Activity
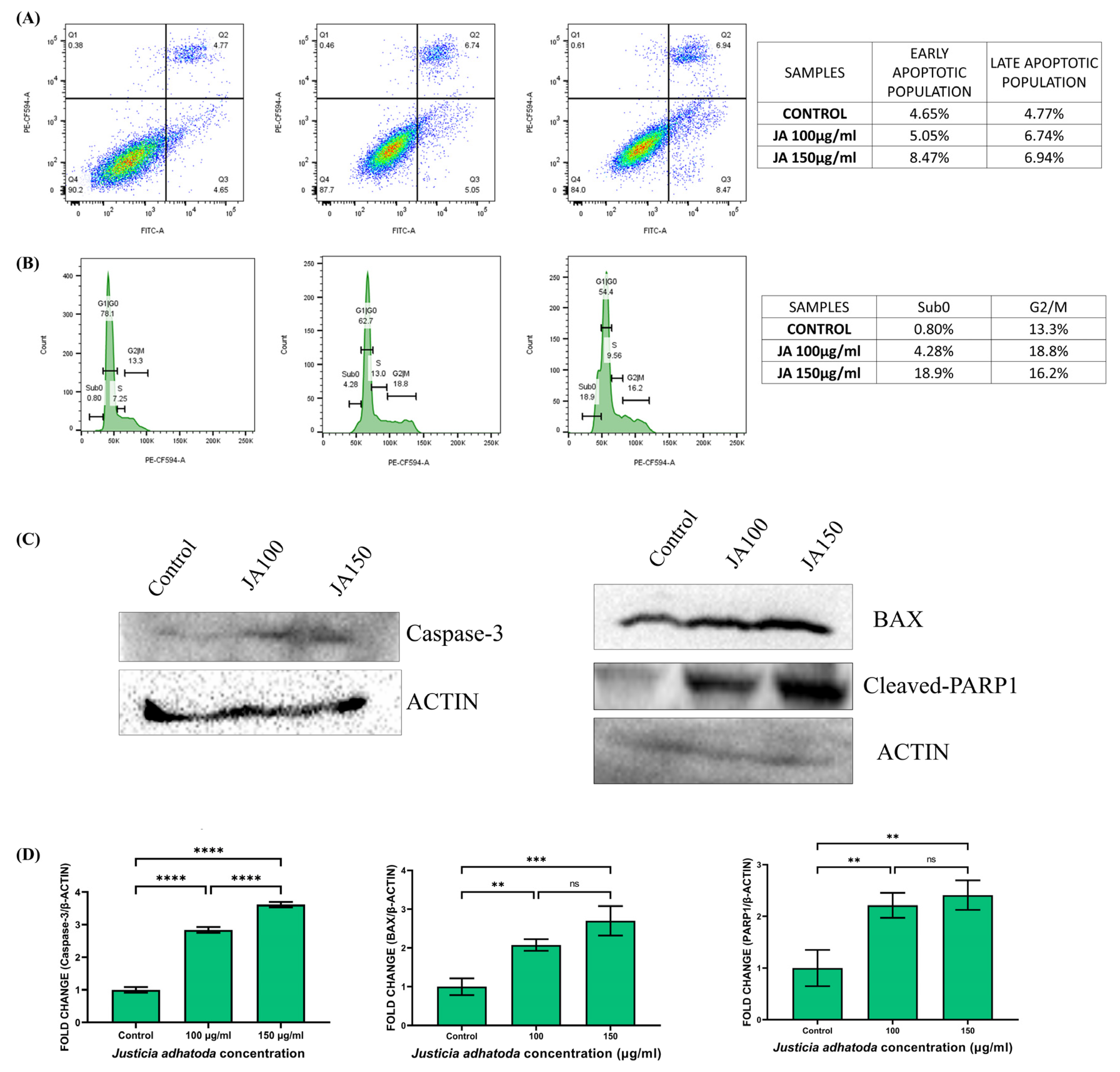
2.10. J. adhatoda Leaf Extract Induces Apoptosis
2.11. J. adhatoda Leaf Extract Induces Sub-G0 and S-Phase Cell Cycle Arrest
2.12. Effect of J. adhatoda Leaf Extract on Apoptosis-Related Protein Expression
2.13. Evaluation of J. adhatoda Leaf Extract on Colony Formation of MCF-7 Cells
2.14. J. adhatoda Leaf Extract Showed Inhibitory Activity in Cell Migration
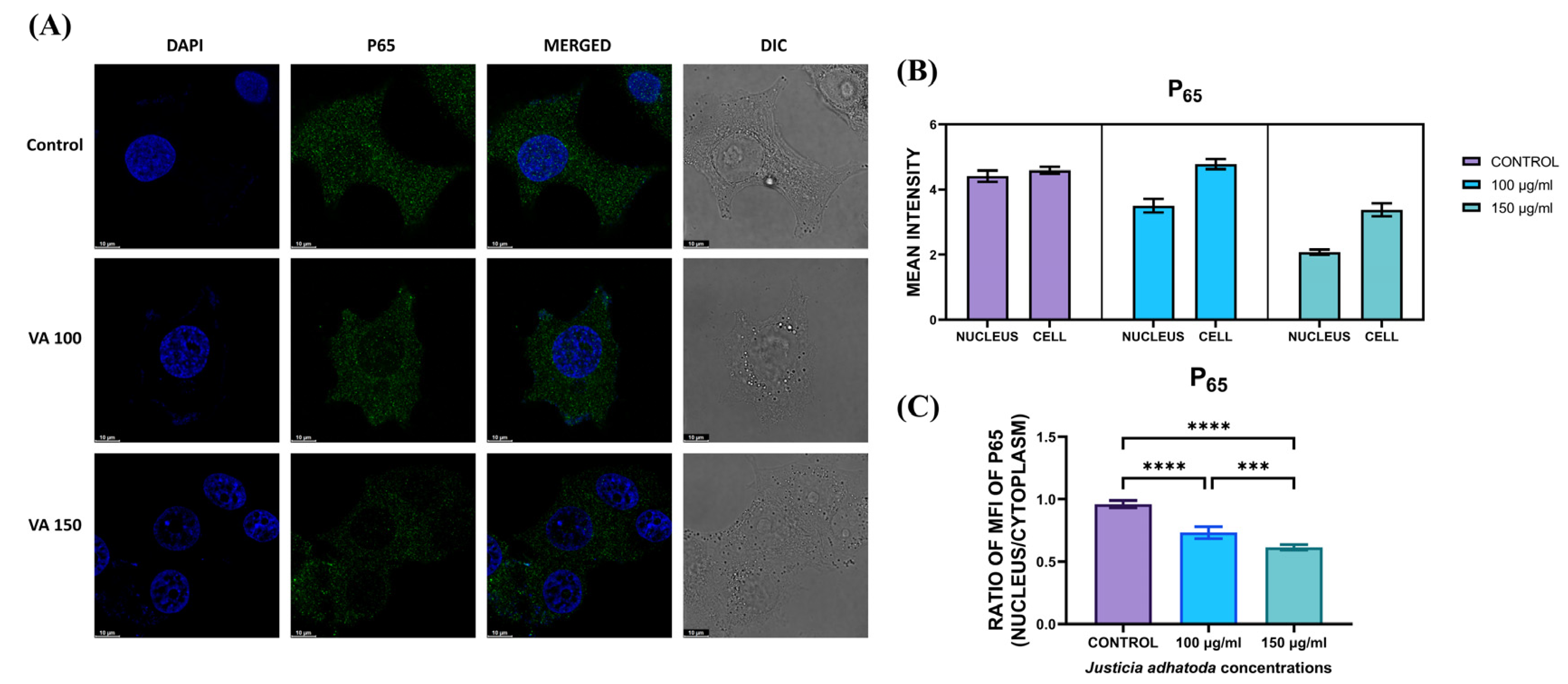
2.15. Evaluation of J. adhatoda Leaf Extract on NF-κB Translocation
3. Discussion
4. Materials and Methods
4.1. Preparation of Mathanolic Extract of J. Adhatoda Leaves
4.2. Analysis of Plant Extract by LC-MS
4.3. DPPH Free Radical Scavenging Activity
4.4. Molecular Docking of the J. adhatoda Leaf Extract towards NF-κB
4.5. Cell Culture
4.6. Measurement of Cell Viability
4.7. Nitric Oxide (NO) Production Assay
4.8. Measurement of Intracellular ROS Generation by DCFDA
4.9. Measurement of Mitochondrial Membrane Potential (ΔΨM) by JC-1
4.10. Quantitative Determination of Glutathione (GSH) and Glutathione Disulfide (GSSG)
4.11. Determination of the Superoxide Dismutase Activity
4.12. Quantification of Apoptotic Cells by Annexin V-PI
4.13. Cell Cycle Analysis
4.14. Western Blot Analysis
4.15. Clonogenic Assay
4.16. Wound Healing/Scratch Assay/Cell Migration Assay
4.17. Translocation of NF-κB
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Global Burden of Disease Cancer Collaboration; Fitzmaurice, C.; Abate, D.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdel-Rahman, O.; Abdelalim, A.; Abdoli, A.; Abdollahpour, I.; et al. Global, regional, and national cancer incidence, mortality, years of life lost, years lived with disability, and disability-adjusted life-years for 29 cancer groups, 1990 to 2017: A systematic analysis for the global burden of disease study. JAMA Oncol. 2019, 5, 1749–1768. [Google Scholar] [PubMed] [Green Version]
- Coughlin, S.S. Epidemiology of breast cancer in women. Adv. Exp. Med. Biol. 2019, 1152, 9–29. [Google Scholar] [PubMed]
- Harbeck, N.; Penault-Llorca, F.; Cortes, J.; Gnant, M.; Houssami, N.; Poortmans, P.; Ruddy, K.; Tsang, J.; Cardoso, F. Breast cancer. Nat. Rev. Dis. Prim. 2019, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Khamis, A.A.A.; Ali, E.M.M.; El-Moneim, M.A.A.; Abd-Alhaseeb, M.M.; El-Magd, M.A.; Salim, E.I. Hesperidin, piperine and bee venom synergistically potentiate the anticancer effect of tamoxifen against breast cancer cells. Biomed. Pharmacother. 2018, 105, 1335–1343. [Google Scholar] [CrossRef] [PubMed]
- Lukong, K.E. Understanding breast cancer–The long and winding road. BBA Clin. 2017, 7, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.; Pan, H.; Godwin, J.; Gray, R.; Arriagada, R.; Raina, V.; Abraham, M.; Medeiros Alencar, V.H.; Badran, A.; Bonfill, X.; et al. Long-term effects of continuing adjuvant tamoxifen to 10 years versus stopping at 5 years after diagnosis of oestrogen receptor-positive breast cancer: ATLAS, a randomised trial. Lancet 2013, 381, 805–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bournine, L.; Bensalem, S.; Fatmi, S.; Bedjou, F.; Mathieu, V.; Iguer-Ouada, M.; Kiss, R.; Duez, P. Evaluation of the cytotoxic and cytostatic activities of alkaloid extracts from different parts of Peganum harmala L. (Zygophyllaceae). Eur. J. Integr. Med. 2017, 9, 91–96. [Google Scholar] [CrossRef]
- Greenwell, M.; Rahman, P. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar]
- Fridlender, M.; Kapulnik, Y.; Koltai, H. Plant derived substances with anti-cancer activity: From folklore to practice. Front. Plant Sci. 2015, 6, 799. [Google Scholar] [CrossRef]
- Alam, S.; Katiyar, D.; Goel, R.; Vats, A.; Mittal, A. Role of herbals in cancer management. J. Phytopharm. 2013, 2, 46–51. [Google Scholar] [CrossRef]
- Jain, S.K. Dictionary of Indian Folk Medicine and Ethnobotany; Deep Publications: New Delhi, India, 1991. [Google Scholar]
- Chowdhury, I.I.; Rahman, A.; Hashem, M.A.; Bhuiyan, M.M.H.; Hajjar, D.; Alelwani, W.; Makki, A.A.; Haque, A.; Tangpong, J.; Bin Bakhtiar, M.T. Supplements of an aqueous combination of Justicia adhatoda and Ocimum tenuiflorum boost antioxidative effects and impede hyperlipidemia. Anim. Model. Exp. Med. 2020, 3, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Bobkov, V.; van der Woning, B.; de Haard, H. Display Technologies for Generation of Ig Single Variable Domains; Methods in Molecuar Biology Series; Humana Press: New York, NY, USA, 2018; Volume 1827, pp. 129–144. [Google Scholar]
- Eep, D.; Ramanjeet, K.; Seema, D. A review on Justicia adhatoda: A potential source of natural medicine. Afr. J. Plant Sci. 2013, 5, 620–627. [Google Scholar]
- Khan, A.M.; Bhadauria, S. Isolation of Some Potential Phytocompounds from Adhatoda Vasica through Gas Chromatography-Mass Spectroscopy Analysis. Asian J. Pharm. Clin. Res. 2017, 10, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, R.P.; Mehrotra, B. Compendium of Indian Medicinal Plants; Central Drug Research Institute: Uttar Pradesh, India, 1990. [Google Scholar]
- Chopra, N.R.; Chopra, I.C. Indigenous Drugs of India; Academic Publishers: Kolkata, India, 1994. [Google Scholar]
- Gutti, U.; Komati, J.K.; Kotipalli, A.; Saladi, R.G.V.; Gutti, R.K. Justicia adhatoda induces megakaryocyte differentiation through mitochondrial ROS generation. Phytomedicine 2018, 43, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Shamsi, Y.; Nikhat, S. Medicinal benefits of Adhatoda vasica Nees.-in unani and contemporary medicine. Cellmed 2020, 10, 13.1–13.7. [Google Scholar]
- Khan, R.; Shamsi, Y.; Nikhat, S. Modulatory influence of Adhatoda vesica (Justicia adhatoda) leaf extract on the enzymes of xenobiotic metabolism, antioxidant status and lipid peroxidation in mice. Mol. Cell Biochem. 2000, 213, 99–109. [Google Scholar]
- Badmanaban, R.; Prajapati Milan, G.; Patel, C.N.; Sen, D.J.; Bibhuranjan, P. Studies on leaf extracts of drug Punarnava by comparing its antioxidant potential by spectrophotometric method. Res. J. Pharm. Technol. 2010, 3, 566–569. [Google Scholar]
- Gatadi, S.; Lakshmi, T.V.; Nanduri, S. 4(3H)-Quinazolinone derivatives: Promising antibacterial drug leads. Eur. J. Med. Chem. 2019, 170, 157–172. [Google Scholar] [CrossRef]
- Du, H.; Jiang, X.; Ma, M.; Xu, H.; Liu, S.; Ma, F. Novel deoxyvasicinone and tetrahydro-beta-carboline hybrids as inhibitors of acetylcholinesterase and amyloid beta aggregation. Bioorganic Med. Chem. Lett. 2020, 30, 127659. [Google Scholar] [CrossRef]
- Shoaib, A.J.C. A systematic ethnobotanical review of Adhatoda vasica (L.), Nees. Cell. Mol. Biol. 2021, 67, 248–263. [Google Scholar] [CrossRef] [PubMed]
- Avula, B.; Begum, S.; Ahmed, S.; Choudhary, M.I.; Khan, I.A. Quantitative determination of vasicine and vasicinone in Adhatoda vasica by high performance capillary electrophoresis. Pharm.-Int. J. Pharm. Sci. 2008, 63, 20–22. [Google Scholar]
- Suryanarayana, A.A.; Shankar, S.; Manoharan, J.P.; Subramanian, V. In-Silico analysis and molecular docking studies of phytoconstituents of Justicia adhatoda as potential inhibitors of SARS-CoV2 target proteins. In Proceedings of the First International Conference on Combinatorial and Optimization, ICCAP 2021, Chennai, India, 7–8 December 2021. [Google Scholar]
- Ranganatha, S.; Shruthi, S.D.; Govindappa, M.; Ramachandra, Y.L. In silico studies of NF-κB protein as anti-cancer and anti-inflammatory target. J. Comput. Methods Mol. 2013, 3, 26–33. [Google Scholar]
- Esparza-López, J.; Longoria, O.; La Cruz-Escobar, D.; Neftali, E.; Garibay-Díaz, J.C.; León-Rodríguez, E.; Ibarra-Sánchez MD, J. Paclitaxel resistance is mediated by NF-κB on mesenchymal primary breast cancer cells. Oncol. Lett. 2022, 23, 50. [Google Scholar] [CrossRef] [PubMed]
- Kastrati, I.; Joosten, S.E.P.; Semina, S.E.; Alejo, L.H.; Brovkovych, S.D.; Stender, J.D.; Horlings, H.M.; Kok, M.; Alarid, E.T.; Greene, G.L.; et al. The NF-κB Pathway Promotes Tamoxifen Tolerance and Disease Recurrence in Estrogen Receptor–Positive Breast Cancers. Mol. Cancer Res. 2020, 18, 1018–1027. [Google Scholar] [CrossRef] [Green Version]
- Vannini, F.; Kashfi, K.; Nath, N. The dual role of iNOS in cancer. Redox Biol. 2015, 6, 334–343. [Google Scholar] [CrossRef] [Green Version]
- Britt, K.L.; Cuzick, J.; Phillips, K.-A. Key steps for effective breast cancer prevention. Nat. Cancer 2020, 20, 417–436. [Google Scholar] [CrossRef]
- Doyle, L.A.; Yang, W.; Abruzzo, L.V.; Krogmann, T.; Gao, Y.; Rishi, A.K.; Ross, D.D. A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc. Natl. Acad. Sci. USA 1998, 95, 15665–15670. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Dash, R. Natural Products for the Management and Prevention of Breast Cancer. Evidence-Based Complement. Altern. Med. 2018, 2018, 8324696. [Google Scholar] [CrossRef]
- Preet, R.; Mohapatra, P.; Mohanty, S.; Sahu, S.K.; Choudhuri, T.; Wyatt, M.D.; Kundu, C.N. Quinacrine has anticancer activity in breast cancer cells through inhibition of topoisomerase activity. Int. J. Cancer 2012, 130, 1660–1670. [Google Scholar] [CrossRef]
- Jani, T.S.; DeVecchio, J.; Mazumdar, T.; Agyeman, A.; Houghton, J.A. Inhibition of NF-κB signaling by quinacrine is cytotoxic to human colon carcinoma cell lines and is synergistic in combination with tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) or oxaliplatin. J. Biol. Chem. 2010, 285, 19162–19172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourgeois-Daigneault, M.-C.; St-Germain, L.E.; Roy, D.G.; Pelin, A.; Aitken, A.S.; Arulanandam, R.; Falls, T.; Garcia, V.; Diallo, J.-S.; Bell, J.C. Combination of Paclitaxel and MG1 oncolytic virus as a successful strategy for breast cancer treatment. Breast Cancer Res. 2016, 18, 83. [Google Scholar] [CrossRef] [Green Version]
- Zeng, F.; Ju, R.-J.; Liu, L.; Xie, H.-J.; Mu, L.-M.; Lu, W.-L. Efficacy in Treating Lung Metastasis of Invasive Breast Cancer with Functional Vincristine Plus Dasatinib Liposomes. Pharmacology 2018, 101, 43–53. [Google Scholar] [CrossRef]
- Wang, L.; Huang, X.-E.; Cao, J. Clinical Study on Safety of Cantharidin Sodium and Shenmai Injection Combined with Chemotherapy in Treating Patients with Breast Cancer Postoperatively. Asian Pac. J. Cancer Prev. 2014, 15, 5597–5600. [Google Scholar] [CrossRef] [Green Version]
- Ranaware, A.M.; Banik, K.; Deshpande, V.; Padmavathi, G.; Roy, N.K.; Sethi, G.; Fan, L.; Kumar, A.P.; Kunnumakkara, A.B. Magnolol: A Neolignan from the Magnolia Family for the Prevention and Treatment of Cancer. Int. J. Mol. Sci. 2018, 19, 2362. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The mechanism of anticancer action and potential clinical use of kaempferol in the treatment of breast cancer. Biomed. Pharmacother. 2019, 117, 109086. [Google Scholar] [CrossRef] [PubMed]
- Pijuan, J.; Barceló, C.; Moreno, D.F.; Maiques, O.; Sisó, P.; Marti, R.M.; Macià, A.; Panosa, A. In vitro Cell Migration, Invasion, and Adhesion Assays: From Cell Imaging to Data Analysis. Front. Cell Dev. Biol. 2019, 7, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, N.; Argenziano, M.; Gigliotti, C.L.; Ferrara, B.; Boggio, E.; Chiocchetti, A.; Caldera, F.; Trotta, F.; Benetti, E.; Annaratone, L.; et al. Paclitaxel-Loaded Nanosponges Inhibit Growth and Angiogenesis in Melanoma Cell Models. Front. Pharmacol. 2019, 10, 776. [Google Scholar] [CrossRef]
- Mishra, D.; Patel, V.; Banerjee, D. Nitric Oxide and S-Nitrosylation in Cancers: Emphasis on Breast Cancer. Breast Cancer Basic Clin. Res. 2020, 14. [Google Scholar] [CrossRef]
- Qian, Q.; Chen, W.; Cao, Y.; Cao, Q.; Cui, Y.; Li, Y.; Wu, J. Targeting Reactive Oxygen Species in Cancer via Chinese Herbal Medicine. Oxidative Med. Cell. Longev. 2019, 2019, 9240426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salaroglio, I.C.; Gazzano, E.; Abdullrahman, A.; Mungo, E.; Castella, B.; Abd-Ellatef, G.E.; Massaia, M.; Donadelli, M.; Rubinstein, M.; Riganti, C.; et al. Increasing intratumor C/EBP-β LIP and nitric oxide levels overcome resistance to doxorubicin in triple negative breast cancer. J. Exp. Clin. Cancer Res. 2018, 37, 286. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Zhao, B.; Chang, H.; Xiao, M.; Wu, Y.; Liu, Y. Paclitaxel suppresses proliferation and induces apoptosis through regulation of ROS and the AKT/MAPK signaling pathway in canine mammary gland tumor cells. Mol. Med. Rep. 2018, 17, 8289–8299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khongthong, P.; Roseweir, A.K.; Edwards, J. The NF-KB pathway and endocrine therapy resistance in breast cancer. Endocr.-Relat. Cancer 2019, 26, R369–R380. [Google Scholar] [CrossRef] [PubMed]
- Nehra, R.; Riggins, R.B.; Shajahan, A.N.; Zwart, A.; Crawford, A.C.; Clarke, R. BCL2 and CASP8 regulation by NF-κB differentially affect mitochondrial function and cell fate in antiestrogen-sensitive and-resistant breast cancer cells. FASEB J. 2010, 24, 2040–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudevan, S.; Parasivam, R.; Sundar, S.; Velauthan, H.; Ramasamy, V. Investigation of anti-inflammatory and anti-cancer activity of Justicia adathoda metabolites. Pak. J. Pharm. Sci. 2019, 32, 1555–1561. [Google Scholar]
- Yan, N.; Wen, D.S.; Zhao, Y.R.; Xu, S.J. Epimedium sagittatum inhibits TLR4/MD-2 mediated NF-κB signaling pathway with anti-inflammatory activity. BMC Complement. Altern. Med. 2018, 18, 303. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, S.A.R.; Waseem, R.; Mahmood, N.; Hussain, Z.; Khan, Z.A.; Shahzad, S.A.; Yar, M.; Amin, R.; Bukhari, S.A.; Zahoor, A.F.; et al. Phenolic acid content, antioxidant properties, and antibacterial potential of flowers and fruits from selected Pakistani indigenous medicinal plants. ScienceAsia 2013, 39, 340–345. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, S.A.R.; Ali, S.; Sherazi, T.A.; Haq, A.-U.; Saeed, M.; Sulman, M.; Rizwan, M.; Alkahtani, S.; Abdel-Daim, M.M. Antioxidant, antibacterial, and anticancer activities of bitter gourd fruit extracts at three different cultivation stages. J. Chem. 2020, 2020, 7394751. [Google Scholar] [CrossRef]
- Michael, S.; Montag, M.; Dott, W. Pro-inflammatory effects and oxidative stress in lung macrophages and epithelial cells induced by ambient particulate matter. Environ. Pollut. 2013, 183, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Sivalingam, K.; Shibu, M.A.; Liao, P.-H.; Ho, T.-J.; Kuo, W.-W.; Chen, R.-J.; Day, C.-H.; Viswanadha, V.P.; Ju, D.-T. Induction of Autophagy by Vasicinone Protects Neural Cells from Mitochondrial Dysfunction and Attenuates Paraquat-Mediated Parkinson’s Disease Associated α-Synuclein Levels. Nutrients 2020, 12, 1707. [Google Scholar] [CrossRef]
- Wu, J.; Chen, Y.; Bai, L.; Liu, Y.; Fu, X.; Zhu, P.; Li, J.; Chou, J.; Yin, C.; Wang, Y.; et al. Chrysoeriol ameliorates TPA-induced acute skin inflammation in mice and inhibits NF-κB and STAT3 pathways. Phytomedicine 2020, 68, 153173. [Google Scholar] [CrossRef]
- Fan, H.; Gao, Z.; Ji, K.; Li, X.; Wu, J.; Liu, Y.; Wang, X.; Liang, H.; Liu, Y.; Li, X.; et al. The in vitro and in vivo anti-inflammatory effect of osthole, the major natural coumarin from Cnidium monnieri (L.) Cuss, via the blocking of the activation of the NF-κB and MAPK/p38 pathways. Phytomedicine 2019, 58, 152864. [Google Scholar] [CrossRef]
- Gasparrini, M.; Forbes-Hernandez, T.Y.; Giampieri, F.; Afrin, S.; Alvarez-Suarez, J.M.; Mazzoni, L.; Mezzetti, B.; Quiles, J.L.; Battino, M. Anti-inflammatory effect of strawberry extract against LPS-induced stress in RAW 264.7 macrophages. Food Chem. Toxicol. 2017, 102, 1–10. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, A.; Shi, M.; Chen, X.; Liu, R.; Li, T.; Zhang, C.; Zhang, Z.; Zhu, L.; Ju, Z.; et al. Analysis of redox landscapes and dynamics in living cells and in vivo using genetically encoded fluorescent sensors. Nat. Protoc. 2018, 13, 2362–2386. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.; Lee, C.H. Development of Thiolated-Graphene Quantum Dots for Regulation of ROS in macrophages. Pharm Res. 2016, 33, 2736–2747. [Google Scholar] [CrossRef] [PubMed]
- Sivandzade, F.; Bhalerao, A.; Cucullo, L. Analysis of the Mitochondrial Membrane Potential Using the Cationic JC-1 Dyeas a Sensitive Fluorescent Probe. Bio-Protoc. 2019, 9, e3128. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Li, C.; Zhang, S.; Li, L.; Hu, Q.; Ji, S. Ursodeoxycholic Acid Protects Against Arsenic Induced Hepatotoxicity by the Nrf2 Signaling Pathway. Front. Pharmacol. 2020, 11, 594496. [Google Scholar] [CrossRef]
- Al–Asbahy, W.M.; Shamsi, M. Dynamics, Synthesis and characterization of Copper–based anticancer compound; in vitro interaction studies with DNA/HSA, SOD mimetic, cytotoxic activity and molecular docking investigation. J. Biomol. Struct. Dyn. 2021, 39, 1431–1446. [Google Scholar] [CrossRef]
- Buolamwini, J.K. Cell cycle molecular targets in novel anticancer drug discovery. Curr. Pharm. Des. 2000, 6, 379–392. [Google Scholar] [CrossRef]
- Verma, K.A.; Singh, S. Phytochemical analysis and in vitro cytostatic potential of ethnopharmacological important medicinal plants. Toxicol. Rep. 2020, 7, 443–452. [Google Scholar] [CrossRef]
- Nair, A.; Manohar, S.M. A flow cytometric journey into cell cycle analysis. Bioanalysis 2021, 13, 1627–1644. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, G.; Zhu, X.; Jiang, K.; Wu, H.; Deng, G.; Qiu, C. Sodium selenite induces apoptosis via ROS-mediated NF-κB signaling and activation of the Bax–caspase-9–caspase-3 axis in 4T1 cells. J. Cell. Physiol. 2019, 234, 2511–2522. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Tang, S.; Wang, M.; Xu, X.; Yao, C.; Wang, S. MicroRNA-497 induces apoptosis and suppresses proliferation via the Bcl-2/Bax-caspase9-caspase3 pathway and cyclin D2 protein in HUVECs. PLoS ONE 2016, 11, e0167052. [Google Scholar] [CrossRef] [PubMed]
- Nelson, V.K.; Sahoo, N.K.; Sahu, M.; Sudhan, H.H.; Pullaiah, C.P.; Muralikrishna, K.S. In vitro anticancer activity of Eclipta alba whole plant extract on colon cancer cell HCT-116. BMC Complement. Med. Ther. 2020, 20, 355. [Google Scholar] [CrossRef] [PubMed]
- Sinjari, B.; Pizzicannella, J.; D’Aurora, M.; Zappacosta, R.; Gatta, V.; Fontana, A.; Trubiani, O.; Diomede, F. Curcumin/Liposome Nanotechnology as Delivery Platform for Anti-inflammatory Activities via NFkB/ERK/pERK Pathway in Human Dental Pulp Treated With 2-HydroxyEthyl MethAcrylate (HEMA). Front. Physiol. 2019, 10, 633. [Google Scholar] [CrossRef]
- Diomede, F.; Zingariello, M.; Cavalcanti, M.F.X.B.; Merciaro, I.; Pizzicannella, J.; de Isla, N.; Caputi, S.; Ballerini, P.; Trubiani, O. MyD88/ERK/NFkB pathways and pro-inflammatory cytokines release in periodontal ligament stem cells stimulated by Porphyromonas gingivalis. Eur. J. Histochem. 2017, 61, 2791. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | PDB ID | Compounds | Binding Energy (k Cal/mol) |
|---|---|---|---|
| NF-κB | 1LE5 | Vasicine | −8.71 |
| Vasicinol | −8.167 | ||
| Vasicinone | −7.604 | ||
| Deoxyvasicinone | −7.56 | ||
| Vasicinolone | −7.141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.; Singh, R.; Dutta, D.; Chandel, S.; Bhattacharya, A.; Ravichandiran, V.; Sukla, S. In Vitro Anticancer Activity of Methanolic Extract of Justicia adhatoda Leaves with Special Emphasis on Human Breast Cancer Cell Line. Molecules 2022, 27, 8222. https://doi.org/10.3390/molecules27238222
Kumar S, Singh R, Dutta D, Chandel S, Bhattacharya A, Ravichandiran V, Sukla S. In Vitro Anticancer Activity of Methanolic Extract of Justicia adhatoda Leaves with Special Emphasis on Human Breast Cancer Cell Line. Molecules. 2022; 27(23):8222. https://doi.org/10.3390/molecules27238222
Chicago/Turabian StyleKumar, Sonu, Rajveer Singh, Debrupa Dutta, Shivani Chandel, Arka Bhattacharya, Velayutham Ravichandiran, and Soumi Sukla. 2022. "In Vitro Anticancer Activity of Methanolic Extract of Justicia adhatoda Leaves with Special Emphasis on Human Breast Cancer Cell Line" Molecules 27, no. 23: 8222. https://doi.org/10.3390/molecules27238222