
Comprehensive Analysis of the Structure and Allergenicity Changes of Seafood Allergens Induced by Non-Thermal Processing: A Review


1
School of Food Science and Engineering, South China University of Technology, Guangzhou 510641, China
2
Academy of Contemporary Food Engineering, South China University of Technology, Guangzhou 510006, China
*
Author to whom correspondence should be addressed.
Molecules 2022, 27(18), 5857; https://doi.org/10.3390/molecules27185857
Submission received: 21 August 2022
/
Revised: 5 September 2022
/
Accepted: 7 September 2022
/
Published: 9 September 2022
(This article belongs to the Special Issue Food Chemistry in Asia)
Abstract
:Seafood allergy, mainly induced by fish, shrimp, crab, and shellfish, is a food safety problem worldwide. The non-thermal processing technology provides a new method in reducing seafood allergenicity. Based on the structural and antigenic properties of allergenic proteins, this review introduces current methods for a comprehensive analysis of the allergenicity changes of seafood allergens induced by non-thermal processing. The IgE-binding capacities/immunoreactivity of seafood allergens are reduced by the loss of conformation during non-thermal processing. Concretely, the destruction of native structure includes degradation, aggregation, uncoiling, unfolding, folding, and exposure, leading to masking of the epitopes. Moreover, most studies rely on IgE-mediated assays to evaluate the allergenic potential of seafood protein. This is not convincing enough to assess the effect of novel food processing techniques. Thus, further studies must be conducted with functional assays, in vivo assays, animal trials, simulated digestion, and intestinal microflora to strengthen the evidence. It also enables us to better identify the effects of non-thermal processing treatment, which would help further analyze its mechanism.
1. Introduction
Seafood, including fish, shrimp, crab, squid, algae, shellfish, and others [1], are popular for their palatability and great nutritional value, such as protein, vitamins, calcium, and irons. Furthermore, eating seafood generates various positive effects, such as antioxidant, anti-cancer, regulation of blood sugar, and others [2,3,4]. However, most of the seafoods (e.g., shrimp, fish, crab, and squid) are known as highly allergenic foodstuffs, likely to trigger allergic symptoms (including asthma, rhinitis, abdominal cramps, vomiting, itchy skin, and even anaphylactic shock in severe cases) upon ingestion [5,6]. As such, the allergic threat of seafood is an obstacle to developing the seafood industry [7]. Recently, the number of food-allergy cases has been continuously rising [8,9], and among them, seafood allergy is relatively severe, especially in Asian countries [10].
For consumers, the immunoglobulin (IgE)-mediated type of food allergy occurs most frequently [11]. When the patients are first exposed to the allergen, their plasma cells would produce specific IgE. Then, the antibodies would attach to the fragment crystallizable receptors (FcR), which are present on the mast cells. Subsequently, mast cells would release mediators to trigger numerous allergic reactions after regaining exposure to the same allergen [12]. So far, however, the most effective therapy for food-allergy patients seems to be avoiding ingesting them. Therefore, how to reduce or eliminate the potential sensitization capacities of seafood has become a hot topic.
Traditionally, heat treatment is a common method to tackle allergic food problems, because their structure generally would be destroyed in high temperature, and even denatured. However, due to the heat stability of some seafood allergens (e.g., tropomyosin and parvalbumin), thermal treatment cannot make a huge difference in their allergenicity [13]. Sometimes, heat treatment even causes an increase in allergenicity [14]. Although extreme thermal conditions may obtain effective results [15], they could result in irreversible losses in food sensory properties and nutritional value. Moreover, chemical and biological methods are also ineffective and uncontrollable in reducing allergenic potential of seafood allergens [16,17,18]. Therefore, novel non-thermal processing technologies are urgently required to solve the challenge of seafood allergies.
Non-thermal processing technologies, including high pressure processing (HPP), cold plasma (CP), ultrasound, pulsed electric field (PEF), and ultraviolet (UV), are considered as effective solutions to food allergens [19,20]. It was reported that side effects of some food allergens were largely reduced under non-thermal processing treatment [21,22,23,24,25,26]. Moreover, new processing technologies also demonstrate their unique effects on the heat-resistance characteristics of seafood allergens. For example, Jin et al. [27] used HPP to process the main allergens of squid (Todarodes pacificus) and found that its immunoreactivity decreased. Ekezie et al. [28] found that the CP jet had a good reducing effect on the IgE-binding capacity of the main allergen of shrimp (Litopenaeus vannamei). Shriver et al. [29] also found that UV could change the IgE/IgG-binding capacity of shrimp (Litopenaeus setiferus) allergens. Furthermore, the non-thermal treatment also has other modificatory advantages on seafood, such as improving the gel properties of surimi [30,31], increasing the hardness of shrimp [32], enhancing the efficacy of active substances [33], and prolonging the shelf life [34]. Therefore, non-thermal processing technology can provide a new possibility for reduction in seafood allergenicity.
The structure of allergens is an important factor that conserves its allergenic potency. For instance, the tropomyosins of crustaceans present a visible α-helices structure, while the arginine kinase of crab and parvalbumin of fish show a compact globular structure. Their integrity of unique structure is always related to their allergenicity. In the studies of seafood allergens treated by HPP, researchers mainly focused on the changes in secondary structure and tertiary structure of allergenic proteins, and thus inferred the mechanism about reducing allergenicity from the structural changes [35,36,37]. For example, Jin et al. [27] suggested that HPP might reduce the immunoreactivity of the tropomyosin of squid by expanding and modifying its secondary structure (53% α-helix was converted into β-sheet and random coils at 600 MPa for 20 min). Therefore, the relationship between the structure and allergenicity of seafood allergens is a crucial way to explore the mechanism of novel processing techniques. As non-thermal processing techniques have a variety of physicochemical effects, the analysis and impacts of structural changes of seafood allergens after non-thermal processing treatment are not comprehensive enough. It is still unclear how the protein structure unfolds or folds during the treatment. In addition, the approaches to access the impact on allergenicity still rely on an IgE/IgG-mediated assay. It is an acceptable but not comprehensive evaluation on allergenicity. The measures of the allergenicity of seafood after treatment require higher weight of evidence (WOE) determinations for verification.
Therefore, focusing on the insights into seafood allergens treated by non-thermal processing and the weakness of structure-allergenicity analysis, this review summarizes the modificatory effect on structure and allergenicity of seafood allergens after non-thermal processing treatment. In addition, new methods of comprehensive analysis are concluded in this review (shown in Figure 1). It would help to establish the relationship between structure and allergenicity, which is of great significance for thoroughly understanding the follow-up study about treating the seafood allergen.
2. Analysis of Structure Changes of Seafood Allergens Induced by Non-Thermal Processing
Most non-thermal processing techniques would break the non-covalent bonds of proteins, leading to the destruction of structure and conformation. Previous studies showed that the immunoreactivity would be influenced once the processing modified the structure of proteins [38,39]. The vast majority of seafood allergens are proteins. Commonly, seafood allergens include tropomyosin (e.g., shrimp, squid, crab, abalone, and fish), parvalbumin (e.g., fish), arginine kinase (e.g., crayfish and lobster), and hemocyanin (e.g., octopus and scallop) (http://allergen.org, accessed on 1 March 2022). In essence, proteins are polypeptide chains formed by the covalent linkage of amino acids, and each molecule has its unique three-dimensional structure. Therefore, the structure of proteins is generally divided into the primary structure, secondary structure, tertiary structure, and quaternary structure [40]. The mechanism of allergen-antibody binding is closely related to its linear epitopes and conformational epitopes [41]. Between them, linear epitopes are peptides composed of 10~20 consecutive amino acids on the amino acid sequence, while conformational epitopes are regions formed of 4~6 amino acids in space [42]. There is a closed relationship between structure and epitopes [43]. Undeniably, non-thermal processing usually results in structural changes of the allergen, leading to masking, exposure, or destruction of its epitopes (shown in Table 1) [19]. Consequently, it is necessary to detect the structural properties of allergens after non-thermal processing treatment.
2.1. Primary Structure Changes
The primary structure, namely the sequence of amino acids, is closely related to the epitopes. During non-thermal processing, epitopes would be altered if the primary structure of the allergen is changed, such as chemical modification on amino acid side chains or breakage of peptide bond. As a result, the epitope would be changed, whether the linear or conformational epitope, so that the IgE-binding capacities are influenced by processing techniques [52]. Accordingly, detecting the primary structure of proteins is an effective and direct method to characterize their allergenic potential. In the studies of food allergens, two methods are usually applied to detect it. One is immediate and accurate, namely mass spectrometry (MS). It can help to detect the weight of molecules and identify the sequence of amino acids. In this way, the chemical modification on protein induced by non-thermal processing can be analyzed [53]. For example, Liu et al. [54] reported that the IgG-binding capacity (determined by ELISA and Western Blotting) of food allergen was altered up to 58.21% during 4 min dielectric-barrier discharge (DBD) plasma treatment. This result may be attributed to the oxidation of some amino acids closed to Met, Tyr, and Phe, which MS detected. However, the equipment is expensive and the process is complex. The other one is sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), which is frequently used to visualize the protein profile [55]. During the SDS-PAGE processing, proteins are negatively charged and move directionally in an electric field. Because of the different mobility caused by the molecular weight, proteins with different molecular weights are separated. In this way, it can reflect the change in the protein’s total molecular weight after non-thermal techniques treatment. Although it cannot specifically reflect amino sequence changes, the molecular weight can help infer whether the covalent bond breaks or the protein is degraded during processing.
In the research on non-thermal processing of seafood allergens, SDS-PAGE is used to analyze the purity of allergenic protein or confirm the presence of allergenic protein. Moreover, it can help to determine whether the allergenic protein degraded or aggregated during non-thermal processing [24]. This can be used to predict whether the polypeptide chains have broken. Furthermore, it has been found that non-thermal processing treatment could reduce the intensities of target bands of allergenic proteins, indicating that the allergen may be degraded into small molecules during the processing. For this reason, their primary structure was destroyed, leading to a decrease in IgE-binding capacities [11,28,48,51]. These results also revealed that non-thermal processing techniques could destroy peptide bonds due to the high-pressure extrusion, oxidation, etching of high-energy electrons, cavitation, and other factors. Besides the decrease in band intensities, some results also showed the appearance of new bands, such as a new band at 26 kDa of fish proteins occurring after HPP treatment [44]. This may be attributed to the low handling intensity (200 MPa, 20 min). However, due to the low sensibility, the results of electrophoresis in some time cannot completely and accurately reflect the changes in behaviors of proteins (e.g., degradation or aggregation). For instance, Zhang et al. [39] used high-intensity ultrasound to process the allergen of shrimp. The electrophoresis results merely suggested the decline of the target band intensity, but not the production of other new bands. Contradictorily, the high-performance liquid chromatography (HPLC) results showed the appearance of another fragment. This may be attributed to the large fragments which cannot pass through the pores of the polyacrylamide or the unmatched polyacrylamide concentration of the gel that cannot catch the small fragments [27,56]. In sum, allergens might degrade during the novel processing according to the SDS-PAGE detections.
2.2. Secondary Structure Changes
The secondary structure of a protein describes the spatial winding and folding of peptide chains. It is maintained primarily by hydrogen bonds, in the form of α-helix, β-sheet, β-turns, and random coils [57]. Changes in secondary structure may lead to alterations in linear or conformational epitopes of allergens, thus decreasing allergenicity. Furthermore, the secondary structure is also the basis of the tertiary structure; if it changes, it could cause the conformational structure to collapse [58]. Therefore, the determination of secondary structure of the allergens is also important in studies of allergens. At present, the practical detections are circular dichroism (CD) and Fourier transform infrared (FTIR) [59]. The secondary structure has different characteristic absorption peaks in the spectrum of CD and FTIR. Moreover, CD has more restrictions on the sample, including functional group, sample state, molecular weight, and concentration of protein, while FTIR does not. For example, Ekezie et al. [28] applied CD to analyze tropomyosin of king prawn (Litopenaeus vannamei) after CP jet treatment. The results showed that the native allergen was an ordered structure dominated by α-helix (negative absorption peak at 208 and 222 nm), and the intensity of the characteristic peak of α-helix decreased gradually after CP treatment. It was indicated that the secondary structure of tropomyosin changed from α-helix to β-sheet and β-turns (peak area at 180~200 nm increases). This may be due to the oxidative modification of amino acid by free radicals produced by CP treatment, which altered the interaction between protein molecules [53]. As the structure was changed, the IgE-binding capacities of allergens were reduced. Jin et al. [27] treated tropomyosin of squid with HPP and Zhang et al. [39] treated tropomyosin of shrimp with high-intensity ultrasound. They obtained similar results on secondary structure of protein. This may indicate that both HPP and ultrasound broke the inner hydrogen bond of tropomyosin, thus changing the protein molecular structure and affecting its allergenicity. Particularly, the attenuated immune-response of tropomyosin from seafood is often accompanied by a transition from α-helix to β-sheet and random coil [27,28,39,48,51]. In FTIR, the peak of amide I (1600–1700 cm−1) is the best characteristic for the secondary structure of proteins. Zhang et al. [11] found that the intensity of amide I peak in hemocyanin of squid decreased after HPP treatment. It was found that with the increase in pressure to 600 MPa, α-helix content decreased from 32.37% to 23.67%, β-turns content increased from 16.80% to 21.95%, and random coils content increased from 32.02% to 37.54%, which was similar with the report by Dong et al. [48], who applied high-intensity ultrasound to processing proteins of shrimp. With the extension of treatment time, the secondary structure changed more and more significantly. Zhang et al. [39] also found that the transformational degree of secondary structure was positively correlated with ultrasonic power. Hence, the secondary structure of seafood allergens would be markedly influenced after non-thermal treatment, showing the transition from order to disorder.
2.3. Tertiary Structure Changes
The tertiary structure of a protein is the conformation formed by the interaction of the amino acid side chain on the basis of the secondary structure. It mainly relies on secondary bonds, including hydrogen bonds, hydrophobic bonds, and van der Waals forces. Some of them also rely on disulfide bonds to maintain the conformation. The tertiary structure of a protein allows interaction between amino acids that are far apart in the primary structure, which may constitute conformational epitopes of allergens. Therefore, the change of tertiary structure may lead to the destruction of conformational epitopes and thus affect allergenic potential. Fluorescence chromatography is frequently used to characterize the changes in polarity of proteins. Due to their phenyl, Trp, Tyr, and Phe have fluorescent properties [60]. Hence, when the protein’s tertiary structure changes, its polarity would alter because of the phenyl exposure. This may result in a blue or red shift of the maximum emission wavelength of the fluorescence spectrum [61]. Ma et al. [50] treated cod protein with ultrasound. They found that with the increase in ultrasonic power, the inherent fluorescence intensity gradually increased, while the maximum emission wavelength (λmax) blue shifted from 347.5 nm to 344 nm. This result indicated that the chromogenic groups of cod protein were exposed more in the aqueous phase after ultrasonic treatment, increasing the hydrophobicity (non-polarity) inside the protein. In other words, the tertiary structure of the cod protein was destroyed, and the structure unfolded after non-thermal processing. Conversely, another research found that the fluorescence intensity of shrimp allergen (tropomyosin) treated with CP decreased by about 16%, while λmax red shifted from 338 nm to 339 nm [28]. This may be because the CP treatment decreased its particle size, making aggregation or recombination of protein easier. This aggregating behavior would mask the internal chromogenic groups, reducing the number of groups exposed to the aqueous phase. Zhang et al. [45] obtained similar results on the fluorescence intrinsic spectrum of fish parvalbumin treated with HPP. It was also observed that fluorescence intensity decreased with the increase in pressure. In a word, changes in tertiary structure can mask or expose epitopes and alter allergenicity, and the way of fluorescence chromatography can help to distinguish the changes in tertiary structure.
Besides noncovalent bond forces, the disulfide bond of covalent force is integral to maintaining conformation. For arginine kinase of crustaceans, a proven allergen, Han et al. [62] found its secondary and tertiary structure would be loose due to the breakage or formation of the disulfide bond, giving rise to potential allergenicity. The free sulfhydryl content (FSC) is related to breakage or formation of disulfide bond (S-S). Therefore, to a certain degree, FSC can explain the change in 3D structure, including conformational expansion and polymerization [63]. Conformational changes lead to shaping in epitopes, which affect allergenicity. Furthermore, studies have shown that targeted destruction of disulfide bonds can reduce the IgE-binding capacities of food allergens [64]. Under the various effect of non-thermal processes (e.g., oxidation, extrusion, vibration, and cavitation), molecules may aggregate, and free sulfhydryl groups would bind to each other to form disulfide bonds, resulting in a new conformation [11,45,50]. For instance, the FSC of allergen of prawn decreased significantly after CP treatment [28]. This may be caused by the formation of new intramolecular/intermolecular disulfide bonds. In addition, some studies suggested that the oxidation effect may expose the internal sulfhydryl groups and reduce the steric hindrance of producing disulfide bonds [54]. Similar results occurred when allergens were treated with HPP, but it was closely related to the treated pressure. It was found that high pressure treatment (200~400 MPa) could reduce FSC in allergens, but at the higher pressures (500~600 MPa), FSC was higher than it was at low pressure [27,65]. This result indicated that high pressure further changed protein conformation [37]. Accordingly, the stability of disulfide bond would shape the allergenic potential of seafood allergens, and it could be modified under the non-thermal processing treatment, forming a new conformation.
3. Analysis of Allergenic Changes of Seafood Allergens Induced by Non-Thermal Processing
3.1. IgE-Binding Capacities/Immunoreactivity Analysis
Seafood allergy is a type I allergic reaction mediated by IgE. The ability of seafood allergens to combine with human serum is the most direct indicator of immune response, namely, its IgE-binding capacities (antigenic integrity). When applying animal antibodies to conduct, it is called immunoreactivity. They are the ability of IgE/IgG to bind to epitopes [13]. In vitro immunoassays, specific IgE in the serum of allergic patients is used to react with the allergens, following characterization of its IgE-binding capacities through enzyme reactions. Enzyme-linked immunosorbent assay (ELISA), western blotting, and dot blotting are the three most common methods for IgE-binding capacities/immunoreactivity analysis.
3.1.1. ELISA Analysis
ELISA can quantify the IgE-binding capacities/immunoreactivity of antigens, including indirect competition ELISA (IC-ELISA), indirect ELISA, and sandwich ELISA (shown in Figure 2). Due to the false-negative defects, direct ELISA is not accurate for seafood allergens detection [66]. In indirect methods, the processed antigen are coated as a stationary phase, then the serum of patient containing the specific antibodies and the enzyme-labeled secondary antibody would add consecutively [67]. For example, Jin et al. [27] used indirect ELISA (conducted by human serum) to test the tropomyosin of squid (Todarodes pacificus) treated by HPP (600 MPa, 20 °C, and 20 s) and found its IgE-binding capacities decreased by 38%. It could be attributed to the unfolding of protein and modification of secondary structure. In another study, Zhang et al. [11] also used indirect ELISA to test the hemocyanin of squid (Todarodes pacificus) treated by HPP (600 MPa, 25 °C and 20 s) and found its IgE-binding capacities and immunoreactivity decreased by 40% and 22%, respectively. The reason seemed to be obvious that it was the response of structure of allergens to HPP. Owing to the loose structure induced by ultrasound treatment (800 W, 15 min), a 46% reduction in IgE-binding capacities of tropomyosin of shrimp (Exopalaemon modestus) was detected by indirect ELISA [39]. UV also reduced the IgE-binding capacity of shrimp (Litopenaeus setiferus) by 25% using the same detection [29]. IC-ELISA has the process that antigen compete to captured antigen, so that it has an amplification effect and high sensitivity [25]. Sandwich ELISA uses the antibody as the stationary phase, and then adds the treated sample and the enzyme-labeled antibody consecutively. In a recent study, Dong et al. [48] used sandwich ELISA to find that the content of tropomyosin of prawn (Litopenaeus vannamei) decreased by 76% after ultrasound treatment (400 W, 20 min). However, it was worth noting that the results obtained by sandwich ELISA and indirect ELISA were slightly different. In a study, the former method showed that IgE-binding capacities of shrimp tropomyosin reduced by 70% after CP treatment 5 min, while the latter only showed a 40% reduction [68]. This may be due to the structural change of the allergens treated by CP, which reduced their connection to the carrier. In a word, from the results of ELISA, non-thermal processing technologies have a great effect on reducing IgE-binding capacities/immunoreactivity of seafood allergens. Notably, compared to others types of ELISA, the indirect competition ELISA may equip the higher WOE.
3.1.2. Western Blotting Analysis
Compared with ELISA quantitative detection, western blotting is used to qualitatively judge by imprinting depth. It contains information of molecular weight, thus possibly determining which specific protein’s allergenicity changes during non-thermal processing treatment. Although using monoclonal antibodies in ELISA can achieve this effect, the preparation process of antibodies is too cumbersome and expensive. For example, the protein blotting of fish (Hypophthalmichthys molitrix) treated with HPP did not change significantly, indicating that their IgE-binding capacities did not vary greatly [46]. Moreover, in another study, it can be seen that the protein blotting gradually weakened with the increase in ultrasonic intensity, indicating that its IgE-binding capacities gradually decreased. Furthermore, with the rise in ultrasound intensity, the protein blotting slipped down, and the molecular weight became smaller, indicating that the ultrasonic processing caused the degradation of allergic proteins [39]. Therefore, western blotting can show the changes in IgE-binding capacities/immunoreactivity accompanied by information of real-time molecular weight. However, the characterization of IgE-binding capacities/immunoreactivity determined by western blotting and ELISA is not necessarily consistent. For example, in a study, the blotting showed that IgE-binding capacities of allergens disappeared after ultrasound treatment at 800 W. Contradictorily, ELISA results showed the allergens still had high allergenic potency [39]. It was concluded that ultrasound treatment broke the allergens into small fragments (including some linear epitope fragments), which are too small to be captured by the gel during the electrophoresis phase. This indicates that ELISA is more sensitive than western blotting in detecting allergenic potential [69]. Summarily, with the consequences of western blotting, allergens usually degrade after non-thermal treatment, resulting in the reduction in IgE-binding capacity.
3.1.3. Dot Blotting Analysis
Dot blotting is similar to western blotting, in which the depth of blotting determines IgE-binding capacities. However, dot blotting does not require the process of electrophoresis, and it can quickly test the changes in the IgE-binding capacities of the total proteins after non-thermal treatment. The reduction in IgE-binding capacities of UV on shrimp (Litopenaeus setiferus) extractions were verified by dot blotting [29], since without electrophoretic separation, dot blotting characterizes the changes throughout the sample after non-thermal treatment.
3.2. Allergenicity Analysis
The definition of allergenicity means the potential of allergens to induce allergic reactions. It should simulate the process of immune disorders in the body. Thus, the analysis of allergenicity includes in vitro cell model assays (simulating mediators release and epithelial transport), in vivo assays (skin prick tests), and animal trials. Comparing the WOE of each immunological analysis, cell model assays and animal trials have the highest WOE, IgE-binding capacity is acceptable, and immunoreactivity has the lowest WOE [13].
3.2.1. Cell Model Assay Analysis
The cell model assay, namely functional biological assays, are used to evaluate the ability of the allergens to induce a variety of responses in the body, including the release of allergenic mediators and activation of primary T cells (shown in Figure 3) [70,71]. Therefore, the cell assays evaluate the allergenicity at the cellular level and the IgE-binding capacity at the molecular level. Common cell models include mast cell (MC) and basophil cell models, dendritic cell (DC) models, and intestinal epithelial cell (IEC) models [70,71,72,73,74,75].
The MC model represents the ability that allergen stimulates the MC to release allergenic mediators (histamine and β-hexosaminidase) [76]. The common MC line is HMC-1, but its expression of surface Fcε R I receptor is low. Thus, it has not been widely used. Basophil models include human cell lines (KU812 and KU812F) and rat cell lines (RBL-2H3). Moreover, the degree of their activation is reflected by the detection of CD63, histamine, tryptase (TPS), interleukin 4 (IL-4), and β-hexosaminidase (β-hex) [77]. In a study, the KU812 model was used to evaluate the allergenicity of tropomyosin of shrimp, and the results showed that the histamine release, β-hex, CD63, and IL-4 significantly decreased after processing treatment [71]. The same cell model was also used to characterize fish allergens (parvalbumin) [75]. Furthermore, Luo et al. [77] used the RBL-2H3 model to evaluate the allergenicity of parvalbumin of fish after digestion by digestive enzymes.
DCs can function as antigen uptake and delivery [70,78]. In addition, DCs are thought to determine the direction of differentiation of naïve T cells and thus modulate the immune response [79]. Although the relationship between DCs and the allergenic potential of protein is not direct or clear, DCs are responsible for presenting antigen, which is an important process in the sensitization and reaction stages. For this reason, the degree of DCs maturity is also closely related to the allergenicity of proteins [80]. Moreover, the expression level of molecules (e.g., MHC II, CD80, CD86, CD103, IL-6, and IL-2) on the surface of DCs can be used to assist in the analysis of allergenicity of proteins after non-thermal treatment [73,81,82]. For example, Wu et al. [70] used the DCs model to evaluate the allergenicity of parvalbumin of fish and measured the release of inflammatory factors (IL-6 and IL-10). The high levels of inflammatory factors also suggested that fish allergens activated the DCs cells.
The mucosal immune system is the first line of defense of the immune system, and IECs are a vital part of it, which acts as a barrier to food allergens [82]. The transmittance of allergens has also become a subsidiary index to evaluate the allergenicity of food allergens because high transmittance means that more allergens can penetrate IEC, thereby inducing more severe allergic reactions. Generally, the Caco-2 cell model is applied to simulate the transport of food allergens by human IECs [83]. Allergens undergo significant structural changes after transport by Caco-2, especially when decomposed by digestive enzymes [84]. Moreover, the allergic reaction also inhibits proliferation of Caco-2 cell and promotes secretion of cytokine [72]. Zhang et al. [85] found that the allergen of shrimp whose IgE-binding capacities decreased, as detected by ELISA, could reduce the increment of Caco-2 and inhibit the release of IL-8 (the mediator to active the basophils).
Although the immune cell models are perfected, few reports have used them to assess the effectiveness of non-thermal processing techniques on seafood allergens. Cellular models can be employed to assist in explaining the modification of allergenic protein by non-thermal processing treatment at the cellular level, which helps comprehensively assess changes on allergenicity of seafood after non-thermal treatment.
3.2.2. Animal Trials Analysis
An imbalance of immune system would cause allergic reactions. It involves many immunoregulatory processes, including gastrointestinal digestion, epithelial transport, microbiota barriers, interleukin upregulation, and mediator release (shown in Figure 3). Thus, in vitro immune response alone cannot adequately reflect the reducing effect of allergenicity induced by non-thermal processing techniques. However, there is a big risk for an allergic reaction happening in the human body, as well as ethical issues. For this reason, animal models offer a great surrogate. Commonly, animal models include Balb/c mice, C57BL/6 mice, C3H/HeJ mice, and DBA2 mice [86]. They are often selected for several generations before inducing sensitization to certain foods. Specifically, the establishment of the animal model includes the sensitization and challenge phase. In the sensitization phase, mice have been sensitized to the targeted allergen by feeding small amounts of the native allergen and immune adjuvant. This phase usually lasts about a month. Afterwards, going into the challenge phase, sensitized mice are fed large amounts of the treated allergen, which would induce allergic reactions. Finally, the degree of allergic reaction in mice is determined by several indicators, including physiological response, the IgE level in the serum, histamine content, and various cytokines content [82]. The allergenic potential and oral tolerance of food allergens after non-thermal processing treatment can be identified through animal trials. In addition to external allergic symptoms, animal trials also include the content on immunoglobulin, intestine, immune cell, and cytokine. Fu et al. [87] fed Balb/c mice with tropomyosin of shrimp and found that the allergic mice lost weight and developed allergic symptoms, including diarrhea and greasy hair. Notably, the male mice did not show significant allergic reactions. With the appearance of allergic symptoms in the challenge phase, the rising levels of histamine, IgE, and IgG2a in blood indicated that allergic reactions occurred. In addition, the level of some cytokine also rose significantly in the sensitized group, such as Il-4, IL-5, and IL-13. In contrast, level of IL-17 decreased, which would affect the polarity differentiation of ILC 2 cells and generate B cells. Sections of the intestine (duodenum, jejunum, and ileum) also showed rising level of mast cells and accumulation of eosinophils after allergic elicitation [88]. Long et al. [47] also found that the challenged mice did not show allergic symptoms after feeding allergens of shrimp treated by HPP combined with heat. Moreover, compared with the control group (fed with untreated allergens), there was a lower level of histamine expression in their serum. Therefore, the reduction effect on allergenicity of seafood allergens by non-thermal processing treatment can be more directly characterized in animal trials.
3.2.3. Skin Prick Tests (SPT) Analysis
SPT is used to judge whether the patient is allergic to a specific antigen in clinical diagnosis [89]. It applies the allergens on the skin and then uses a needle to pierce it into the skin. If the patient is allergic to this allergen, the allergen will bind to the antibody and stimulate mast cells to release mediators, causing red spots to appear on the skin of patient after a few minutes [90]. Afterward, the diameter of wheals is used to determine the degree of allergic reaction. Therefore, SPT can help evaluate the allergenicity of seafood allergens according to the diameter of wheals. Compared with prolonged animal trials, SPT can be done in minutes [91]. It is worth noting that although the sensitivity of skin prick is high, its specificity is low due to a series of antibodies contained in the body. Lavilla et al. [92] used SPT to determine the allergenicity of food allergen after HPP treatment but found the opposite results to ELISA. This may be related to the low specificity of SPT. However, Gamez et al. [93] adopted shrimp protein extract as the allergen and obtained consistent results between SPT and ELISA. These unexpected results emphasize the importance of in vivo assays in evaluating the effects of non-thermal processing, despite negative bias in SPT tests [89].
3.3. In Vitro Digestibility Analysis
Digestibility is an important index to evaluate the ability to be absorbed in the digestive tract. Generally, food allergens are taken orally, subsequently inducing an allergic reaction in the digestive tract if the proteins maintain a relatively integral structure [64,94,95,96]. In food allergy research, digestibility is usually considered the resistance to digestive enzymes (e.g., pepsin, trypsin, and chymotrypsin). It practices by determining the degree of enzymatic hydrolysis of digestive enzymes to allergens in a specified time. If the allergens have a low resistance to digestive enzymes, their amino acid sequence will be destroyed due to the greater degree of enzymatic hydrolysis. This may result in the loss of epitopes, causing the immune cells cannot to recognize them. Therefore, the allergenicity of food allergens may relate to their digestibility. It determines whether allergens can pass through the gastrointestinal tract and then cause anaphylaxis and inflammatory response. For this reason, it is gradually used as an index or a characterization in food allergens research. Moreover, it also has been recommended as a criterion for evaluating food allergenicity by some international organizations [97]. For instance, Huang et al. [98] studied the relationship between the IgE-binding capacities of tropomyosin of mud crab and its digestibility. The results revealed that tropomyosin showed resistance to pepsin but was relatively easy to degrade by trypsin and chymotrypsin. Furthermore, its allergenicity was gradually decreased with the increasing degree of enzymatic hydrolysis. Lv et al. [67] also found that the IgE-binding capacities of allergens of shrimp decreased with increasing digestion degree. Liu et al. [99] recommended that tropomyosin from Pacific white shrimp (Litopenaeus vannamei) was possibly more allergenic than it from Grass prawn (Penaeus monodon) due to its higher digestion stability as well. Therefore, the digestion ability of proteins is widely used as an indicator related to its allergenic potency in food allergen research, especially seafood allergens. Nevertheless, the results of this indicator are usually affected by many factors, including the pH, the proteolytic, and the enzyme-to-protein ratio, so it does not have a specific criterion. Furthermore, the relationship between the digestibility and allergenicity of food allergens was not fully understood. Some studies even showed no significant correlation between the digestibility of certain food proteins and their allergenicity [100], but this did not prevent it from becoming a worthy reference index in the studies of seafood allergen. In addition, several research about non-thermal processing treatment take advantage of this method to investigate the mechanism of reducing effect on allergenic potency [27,48].
It has been shown that most seafood allergens would be more tolerable to digestive enzymes than non-food allergens [97], such as tropomyosin of crab, a proven allergen, which is hardly digested by pepsin [96]. Hence, digestibility is recognized as an effective indicator of structure and allergenicity of allergen during the non-thermal processing treatment. For example, Jin et al. [27] explored the relationship between IgE-binding capacities of tropomyosin Tod p1 (TMTp1) of squid and its digestibility after HPP treatment. In particular, the digestion stability assay of simulated gastric fluid (SGF) and simulated intestinal fluid (SIF) revealed that with the digestion time increasing, the IgG/IgE special reactivity of TMTp1 decreased, and it was more resistant to pepsin than trypsin. In addition, HPP treatment promoted TMTp1 to be digested due to its structural unfolding and exposure. Coincidentally, Dong et al. [48] also reported that the digestibility of proteins of shrimp increased from 76.42% to 83.95% after 20 min 400 W of ultrasound processing, while the content of tropomyosin (chief allergen) decreased by 76%. These results could help explain the structural modification of ultrasound treatment, including a decrease in α-helix content and an increase in β-sheet. On the other hand, the digestion stability of food proteins is also a critical food property, directly relating to edibility after non-thermal processing. Therefore, digestibility of seafood allergens can not only reflect the allergenicity, but also contribute to explaining the structural changes induced by non-thermal processing technology. It is a worthwhile and valuable experimental indicator in food allergens research.
3.4. Gut Microbiota Analysis
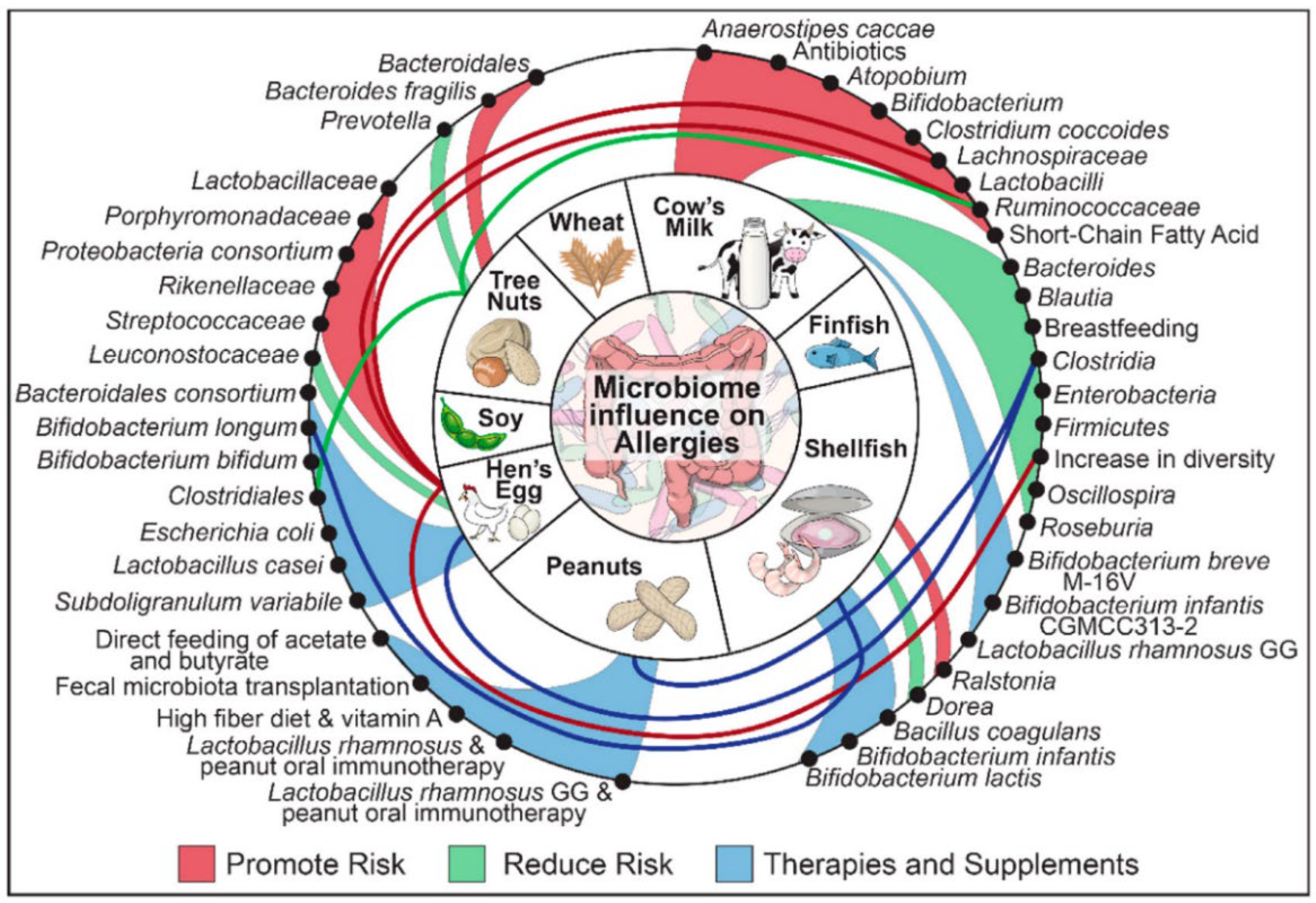
In addition to well-established serum assays and macroscopic animal trials, changes in gut microbes may also provide a new way to characterize food allergic reactions. As an important line of defense against food allergy, the barrier of gut microbiota located in intestine is closely related to the immune system. Some studies have shown that the composition of gut bacteria may promote or suppress food allergies (shown in Figure 4) [101]. The mucosal layer of the intestine is home to a large number of microorganisms (gut microbiota), which play a significant role in regulating the immune system and maintaining the intestinal barrier (including mechanical, biological, chemical, and immune barriers) [102]. It was shown that intestinal microbes could affect food allergic reactions by altering the permeability of intestinal mucosal and state of immune system [103,104]. For example, certain gut bacteria (e.g., Lactobacillus acidophilus) can enhance function of IECs barrier by strengthening their tight junctions. Moreover, short-chain fatty acids produced by bacterial fermentation can also promote the proliferation and differentiation of IECs [105]. Besides playing a role of barrier, the composition of microbiome influences human sensitization to food allergens [106]. It could affect the direction of differentiation of immune cell [107]. For example, the number of Bacteroidetes decreased and Firmicutes increased after a shrimp-stimulated allergic state [87], which would promote the differentiation of ILC2 into Th2 and regulate Treg-mediated immune tolerance [108]. Moreover, Lachnospiraceae, a family in the Clostridia class, also increased significantly, which was associated with oral tolerance [109]. Feehley et al. [110] also pointed out that the Clostridia class affected the food allergic reactions by regulating the level of Treg cells. Similarly, Wu et al. [70] demonstrated that parvalbumin of fish also caused disturbance of intestinal microbiota. At present, although the quantitative and qualitative relationship between them is still unclear, a large number of studies have evidenced that the occurrence of anaphylaxis is inevitably accompanied by significant changes in the composition of intestinal flora [82,111,112]. As an essential part of the body’s immune system and metabolic function, intestinal microbes can be used to characterize the treatment effect and potential safety problems of non-thermal processing more comprehensively.
4. Conclusions and Future Trends
A large number of studies have confirmed that non-thermal processing techniques can reduce the IgE-binding capacities of seafood allergens. However, due to its fuzzy processing mechanism, such a conclusion needs more robust, thorough, and validated assessments to verify it. From the perspective of structure and allergenicity, non-thermal processing would modify the structural integrity of seafood allergens, leading to a reduction in IgE-binding capacities/immunoreactivity. After gathering the available reported research, non-thermal processing techniques had the potential to break down the covalent/non-covalent bond of seafood allergens; through the structural analysis, allergic proteins would undergo degradation, aggregation, uncoiling, unfolding, folding, and exposure behaviors, resulting in a significant change of conformation. Subsequently, these disruptions mask or expose the epitopes, influencing their allergenic potency. Furthermore, structural incompleteness induced by non-thermal processing treatment would alter their resistance to digestive enzymes, especially pepsin. In addition, functional assays, in vivo assays, or animal trials should be conducted to evaluate the allergenicity of treated seafood allergens. Only in this way could the actual reducing effect on allergenicity induced by non-thermal processing be revealed.
In order to use non-thermal processing technologies as new processing ways, it is necessary to use various methods to analyze their processing effect and processing mechanism, which would test their effectiveness, reliability, and safety. The relationship between structure and allergenicity of seafood allergen needs more data and studies to support it. In addition, some other methods of evaluation related to allergenicity need to be perfected, such as Ca2+ binding ability, the interaction of various cytokines, and X-ray diffraction. Only after a comprehensive analysis on the effect of non-thermal processing technologies can they provide practical support for food industries in the future.
Author Contributions
F.W.: writing—original draft, formal analysis, investigation; H.Z.: formal analysis, investigation; J.-H.C.: supervision, funding acquisition, resources, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The authors are grateful to the National Natural Science Foundation of China (31972205) for its support. This research was also supported by Guangdong Basic and Applied Basic Research Foundation (2021A1515010644).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ruethers, T.; Taki, A.C.; Johnston, E.B.; Nugraha, R.; Le, T.T.K.; Kalic, T.; McLean, T.R.; Kamath, S.D.; Lopata, A.L. Seafood allergy: A comprehensive review of fish and shellfish allergens. Mol. Immunol. 2018, 100, 28–57. [Google Scholar] [CrossRef]
- Nagai, T.; Suzuki, N.; Nagashima, T. Antioxidative activities and angiotensin I-converting enzyme inhibitory activities of enzymatic hydrolysates from commercial kamaboko type samples. Food Sci. Technol. Int. 2006, 12, 335–346. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, C.H.; Chen, S.J. Comparison of active non-volatile taste components in the viscera and adductor muscles of oyster (Ostrea rivularis Gould). Food Sci. Technol. Res. 2013, 19, 417–424. [Google Scholar] [CrossRef]
- Zheng, L.F.; Yu, H.C.; Wei, H.K.; Xing, Q.; Zou, Y.; Zhou, Y.F.; Peng, J. Antioxidative peptides of hydrolysate prepared from fish skin gelatin using ginger protease activate antioxidant response element-mediated gene transcription in IPEC-J2 cells. J. Funct. Foods 2018, 51, 104–112. [Google Scholar] [CrossRef]
- Burks, A.W.; Tang, M.; Sicherer, S.; Muraro, A.; Eigenmann, P.A.; Ebisawa, M.; Fiocchi, A.; Chiang, W.; Beyer, K.; Wood, R.; et al. Icon: Food allergy. J. Allergy Clin. Immunol. 2012, 129, 906–920. [Google Scholar] [CrossRef]
- Ochfeld, E.N.; Pongracic, J.A. Food allergy: Diagnosis and treatment. Allergy Asthma Proc. 2019, 40, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, J.; Selvasekaran, P.; Lokanadham, M.; Chidambaram, R. Food and food products associated with food allergy and food intolerance An overview. Food Res. Int. 2020, 138, 109780. [Google Scholar] [CrossRef]
- Renz, H.; Allen, K.J.; Sicherer, S.H.; Sampson, H.A.; Lack, G.; Beyer, K.; Oettgen, H.C. Food allergy. Nat. Rev. Dis. Primers 2018, 4, 17098. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Cheng, J.H. DNA, protein and aptamer-based methods for seafood allergens detection: Principles, comparisons and updated applications. Crit. Rev. Food Sci. Nutr. 2021, 1–14. [Google Scholar] [CrossRef]
- Shek, L.P.C.; Cabrera-Morales, E.A.; Soh, S.E.; Gerez, I.; Ng, P.Z.; Yi, F.C.; Ma, S.; Lee, B.W. A population-based questionnaire survey on the prevalence of peanut, tree nut, and shellfish allergy in 2 Asian populations. J. Allergy Clin. Immunol. 2010, 126, 324–331.E7. [Google Scholar] [CrossRef]
- Zhang, Y.; Deng, Y.; Zhao, Y. Structure-based modelling of hemocyanin allergenicity in squid and its response to high hydrostatic pressure. Sci. Rep. 2017, 7, 40021. [Google Scholar] [CrossRef] [PubMed]
- Sena-Torralba, A.; Pallas-Tamarit, Y.; Morais, S.; Maquieira, A. Recent advances and challenges in food-borne allergen detection. Trac, Trends Anal. Chem. 2020, 132, 116050. [Google Scholar] [CrossRef]
- Costa, J.; Villa, C.; Verhoeckx, K.; Cirkovic-Velickovic, T.; Schrama, D.; Roncada, P.; Rodrigues, P.M.; Piras, C.; Martin-Pedraza, L.; Monaci, L.; et al. Are Physicochemical Properties Shaping the Allergenic Potency of Animal Allergens? Clin. Rev. Allergy Immunol. 2022, 62, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Kamath, S.D.; Rahman, A.M.A.; Komoda, T.; Lopata, A.L. Impact of heat processing on the detection of the major shellfish allergen tropomyosin in crustaceans and molluscs using specific monoclonal antibodies. Food Chem. 2013, 141, 4031–4039. [Google Scholar] [CrossRef]
- Cuadrado, C.; Cabanillas, B.; Pedrosa, M.M.; Varela, A.; Guillamon, E.; Muzquiz, M.; Crespo, J.F.; Rodriguez, J.; Burbano, C. Influence of thermal processing on IgE reactivity to lentil and chickpea proteins. Mol. Nutr. Food Res. 2009, 53, 1462–1468. [Google Scholar] [CrossRef]
- Lasekan, A.O.; Nayak, B. Effects of buffer additives and thermal processing methods on the solubility of shrimp (Penaeus monodon) proteins and the immunoreactivity of its major allergen. Food Chem. 2016, 200, 146–153. [Google Scholar] [CrossRef]
- Nakamura, A.; Watanabe, K.; Ojima, T.; Ahn, D.H.; Saeki, H. Effect of Maillard reaction on allergenicity of scallop tropomyosin. J. Agric. Food Chem. 2005, 53, 7559–7564. [Google Scholar] [CrossRef]
- Fu, L.; Wang, C.; Wang, J.; Ni, S.; Wang, Y. Maillard reaction with ribose, galacto-oligosaccharide or chitosan-oligosaccharide reduced the allergenicity of shrimp tropomyosin by inducing conformational changes. Food Chem. 2019, 274, 789–795. [Google Scholar] [CrossRef]
- Ekezie, F.G.C.; Cheng, J.H.; Sun, D.W. Effects of nonthermal food processing technologies on food allergens: A review of recent research advances. Trends Food Sci. Technol. 2018, 74, 12–25. [Google Scholar] [CrossRef]
- Dasanayaka, B.P.; Li, Z.; Pramod, S.N.; Chen, Y.; Khan, M.U.; Lin, H. A review on food processing and preparation methods for altering fish allergenicity. Crit. Rev. Food Sci. Nutr. 2020, 62, 1951–1970. [Google Scholar] [CrossRef]
- Prieto, N.; Burbano, C.; Iniesto, E.; Rodriguez, J.; Cabanillas, B.; Crespo, J.F.; Pedrosa, M.M.; Muzquiz, M.; Del Pozo, J.C.; Linacero, R.; et al. A novel proteomic analysis of the modifications induced by high hydrostatic pressure on hazelnut water-soluble proteins. Foods 2014, 3, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Maleki, S.J.; Rodriguez, J.; Cheng, H.; Teuber, S.S.; Wallowitz, M.L.; Muzquiz, M.; Pedrosa, M.M.; Linacero, R.; Burbano, C.; et al. Allergenic properties and differential response of walnut subjected to processing treatments. Food Chem. 2014, 157, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Bai, Y.; Gao, J.; Li, X.; Chen, H. Effects of high hydrostatic pressure on the structure and potential allergenicity of the major allergen bovine beta-lactoglobulin. Food Chem. 2017, 219, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Meinlschmidt, P.; Ueberham, E.; Lehmann, J.; Reineke, K.; Schlüter, O.; Schweiggert-Weisz, U.; Eisner, P. The effects of pulsed ultraviolet light, cold atmospheric pressure plasma, and gamma-irradiation on the immunoreactivity of soy protein isolate. Innov. Food Sci. Emerg. Technol. 2016, 38, 374–383. [Google Scholar] [CrossRef]
- Tammineedi, C.V.R.K.; Choudhary, R.; Perez-Alvarado, G.C.; Watson, D.G. Determining the effect of UV-C, high intensity ultrasound and nonthermal atmospheric plasma treatments on reducing the allergenicity of α-casein and whey proteins. LWT-Food Sci. Technol. 2013, 54, 35–41. [Google Scholar] [CrossRef]
- Nooji, J.K. Reduction of Wheat Allergen Potency by Pulsed Ultraviolet Light, High Hydrostatic Pressure, and Non-Thermal Plasma. Master’s Thesis, University of Florida, Gainesville, FL, USA, 2011. [Google Scholar]
- Jin, Y.; Deng, Y.; Qian, B.; Zhang, Y.; Liu, Z.; Zhao, Y. Allergenic response to squid (Todarodes pacificus) tropomyosin Tod p1 structure modifications induced by high hydrostatic pressure. Food Chem. Toxicol. 2015, 76, 86–93. [Google Scholar] [CrossRef]
- Ekezie, F.C.; Sun, D.W.; Cheng, J.H. Altering the IgE binding capacity of king prawn (Litopenaeus vannamei) tropomyosin through conformational changes induced by cold argon-plasma jet. Food Chem. 2019, 300, 125143. [Google Scholar] [CrossRef]
- Shriver, S.; Yang, W.; Chung, S.Y.; Percival, S. Pulsed ultraviolet light reduces immunoglobulin E binding to Atlantic white shrimp (Litopenaeus setiferus) extract. Int. J. Environ. Res. Public Health 2011, 8, 2569–2583. [Google Scholar] [CrossRef]
- Zhou, A.; Lin, L.; Liang, Y.; Benjakul, S.; Shi, X.; Liu, X. Physicochemical properties of natural actomyosin from threadfin bream (Nemipterus spp.) induced by high hydrostatic pressure. Food Chem. 2014, 156, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Ekezie, F.G.; Cheng, J.H.; Sun, D.W. Effects of mild oxidative and structural modifications induced by argon plasma on physicochemical properties of actomyosin from king prawn (Litopenaeus vannamei). J. Agric. Food Chem. 2018, 66, 13285–13294. [Google Scholar] [CrossRef]
- Li, X.Y.; Li, Z.X.; Lin, H.; Samee, H. Effect of power ultrasound on the immunoactivity and texture changes of shrimp (Penaeus vannamei). Czech J. Food Sci. 2011, 29, 508–514. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, B.; Deng, Y.; Zhao, Y. In vitro anti-inflammatory and antioxidant activities and protein quality of high hydrostatic pressure treated squids (Todarodes pacificus). Food Chem. 2016, 203, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Rathod, N.B.; Kulawik, P.; Ozogul, Y.; Ozogul, F.; Bekhit, A.E.A. Recent developments in non-thermal processing for seafood and seafood products: Cold plasma, pulsed electric field and high hydrostatic pressure. Int. J. Food Sci. Technol. 2021, 57, 774–790. [Google Scholar] [CrossRef]
- Grigera, J.R.; McCarthy, A.N. The behavior of the hydrophobic effect under pressure and protein denaturation. Biophys. J. 2010, 98, 1626–1631. [Google Scholar] [CrossRef]
- De Oliveira, F.A.; Neto, O.C.; doc Santos, L.M.R.; Ferreira, E.H.R.; Rosenthal, A. Effect of high pressure on fish meat quality—A review. Trends Food Sci. Technol. 2017, 66, 1–19. [Google Scholar] [CrossRef]
- Kurpiewska, K.; Biela, A.; Loch, J.I.; Lipowska, J.; Siuda, M.; Lewinski, K. Towards understanding the effect of high pressure on food protein allergenicity: Beta-lactoglobulin structural studies. Food Chem. 2019, 270, 315–321. [Google Scholar] [CrossRef]
- Sathe, S.K.; Liu, C.Q.; Zaffran, V.D. Food Allergy. Annu. Rev. Food Sci. Technol. 2016, 7, 191–220. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Chen, W.; Zhou, P. Conformation stability, in vitro digestibility and allergenicity of tropomyosin from shrimp (Exopalaemon modestus) as affected by high intensity ultrasound. Food Chem. 2018, 245, 997–1009. [Google Scholar] [CrossRef]
- Cianci, M. Introduction to proteins, introduction to proteins: Structure, function, and motion, 2nd edition. Crystallogr. Rev. 2020, 27, 47–50. [Google Scholar] [CrossRef]
- Rahaman, T.; Vasiljevic, T.; Ramchandran, L. Effect of processing on conformational changes of food proteins related to allergenicity. Trends Food Sci. Technol. 2016, 49, 24–34. [Google Scholar] [CrossRef]
- Zhou, F.L.; He, S.D.; Sun, H.J.; Wang, Y.F.; Zhang, Y. Advances in epitope mapping technologies for food protein allergens: A review. Trends Food Sci. Technol. 2021, 107, 226–239. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, Y.; Bi, Y.; Wang, Q.; Cheng, K.-W.; Chen, F. Review: Seafood allergy and potential application of high hydrostatic pressure to reduce seafood allergenicity. Int. J. Food Eng. 2019, 15, 20180392. [Google Scholar] [CrossRef]
- Zhang, Y.; Bi, Y.; Wang, Q.; Cheng, K.W.; Chen, F. Application of high pressure processing to improve digestibility, reduce allergenicity, and avoid protein oxidation in cod (Gadus morhua). Food Chem. 2019, 298, 125087. [Google Scholar] [CrossRef]
- Zhang, H.E.; Liao, H.Q.; Lu, Y.B.; Hu, Y.H.; Yang, H.; Cao, S.Q.; Qi, X.Y. Effects of high hydrostatic pressure on the structural characteristics of parvalbumin of cultured large yellow croaker (Larimichthys crocea). J. Food Process. Preserv. 2020, 44, e14911. [Google Scholar] [CrossRef]
- Liu, R.; Xue, W.T. High-pressure treatment with silver carp (Hypophthalmichthys molitrix) protein and its allergic analysis. High Press. Res. 2010, 30, 438–442. [Google Scholar] [CrossRef]
- Long, F.Y.; Yang, X.; Wang, R.R.; Hu, X.S.; Chen, F. Effects of combined high pressure and thermal treatments on the allergenic potential of shrimp (Litopenaeus vannamei) tropomyosin in a mouse model of allergy. Innov. Food Sci. Emerg. Technol. 2015, 29, 119–124. [Google Scholar] [CrossRef]
- Dong, X.; Wang, J.; Raghavan, V. Effects of high-intensity ultrasound processing on the physiochemical and allergenic properties of shrimp. Innov. Food Sci. Emerg. Technol. 2020, 65, 102441. [Google Scholar] [CrossRef]
- Li, Z.X.; Lin, H.; Cao, L.M.; Jameel, K. Effect of high intensity ultrasound on the allergenicity of shrimp. J. Zhejiang Univ. Sci. B 2006, 7, 251–256. [Google Scholar] [CrossRef]
- Ma, W.; Wang, J.; Xu, X.; Qin, L.; Wu, C.; Du, M. Ultrasound treatment improved the physicochemical characteristics of cod protein and enhanced the stability of oil-in-water emulsion. Food Res. Int. 2019, 121, 247–256. [Google Scholar] [CrossRef]
- Yang, W.W.; Shriver, S.K.; Chung, S.Y.; Percival, S.; Correll, M.J.; Rababah, T.M. In vitro gastric and intestinal digestions of pulsed light-treated shrimp extracts. Appl. Biochem. Biotechnol. 2012, 166, 1409–1422. [Google Scholar] [CrossRef]
- Dong, X.; Wang, J.; Raghavan, V. Critical reviews and recent advances of novel non-thermal processing techniques on the modification of food allergens. Crit. Rev. Food Sci. Nutr. 2021, 61, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Takai, E.; Kitamura, T.; Kuwabara, J.; Ikawa, S.; Yoshizawa, S.; Shiraki, K.; Kawasaki, H.; Arakawa, R.; Kitano, K. Chemical modification of amino acids by atmospheric-pressure cold plasma in aqueous solution. J. Phys. D Appl. Phys. 2014, 47, 285403. [Google Scholar] [CrossRef]
- Liu, Z.W.; Liu, L.J.; Zhou, Y.X.; Tan, Y.C.; Cheng, J.H.; Bekhit, A.E.; Inam-Ur-Raheem, M.; Aadil, R.M. Dielectric-barrier discharge (DBD) plasma treatment reduces IgG binding capacity of beta-lactoglobulin by inducing structural changes. Food Chem. 2021, 358, 129821. [Google Scholar] [CrossRef]
- Faisal, M.; Vasiljevic, T.; Donkor, O.N. A review on methodologies for extraction, identification and quantification of allergenic proteins in prawns. Food Res. Int. 2019, 121, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Shriver, S.K.; Yang, W.W. Thermal and nonthermal methods for food allergen control. Food Eng. Rev. 2011, 3, 26–43. [Google Scholar] [CrossRef]
- Sharafodin, H.; Soltanizadeh, N. Potential application of DBD plasma technique for modifying structural and physicochemical properties of Soy Protein Isolate. Food Hydrocoll. 2022, 122, 107077. [Google Scholar] [CrossRef]
- Lv, L.; Lin, H.; Li, Z.; Wang, J.; Ahmed, I.; Chen, H. Changes of structure and IgE binding capacity of shrimp (Metapenaeus ensis) tropomyosin followed by acrolein treatment. Food Funct. 2017, 8, 1028–1036. [Google Scholar] [CrossRef]
- Vanga, S.K.; Singh, A.; Kalkan, F.; Gariepy, Y.; Orsat, V.; Raghavan, V. Effect of thermal and high electric fields on secondary structure of peanut protein. Int. J. Food Prop. 2015, 19, 1259–1271. [Google Scholar] [CrossRef]
- Tong, P.; Gao, J.Y.; Chen, H.B.; Li, X.; Zhang, Y.; Jian, S.; Wichers, H.; Wu, Z.H.; Yang, A.S.; Liu, F.H. Effect of heat treatment on the potential allergenicity and conformational structure of egg allergen ovotransferrin. Food Chem. 2012, 131, 603–610. [Google Scholar] [CrossRef]
- Bußler, S.; Steins, V.; Ehlbeck, J.; Schlüter, O. Impact of thermal treatment versus cold atmospheric plasma processing on the techno-functional protein properties from Pisum sativum ‘Salamanca’. J. Food Eng. 2015, 167, 166–174. [Google Scholar] [CrossRef]
- Han, X.Y.; Yang, H.; Rao, S.T.; Liu, G.Y.; Hu, M.J.; Zeng, B.C.; Cao, M.J.; Liu, G.M. The maillard reaction reduced the sensitization of tropomyosin and arginine kinase from Scylla paramamosain, simultaneously. J. Agric. Food Chem. 2018, 66, 2934–2943. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.Q.; Chen, H.B.; Gao, J.Y.; Luo, C.P.; Ma, X.J.; Tong, P. High-pressure microfluidisation-induced changes in the antigenicity and conformation of allergen Ara h 2 purified from Chinese peanut. J. Sci. Food Agric. 2011, 91, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Hazebrouck, S.; Guillon, B.; Drumare, M.F.; Paty, E.; Wal, J.M.; Bernard, H. Trypsin resistance of the major peanut allergen Ara h 6 and allergenicity of the digestion products are abolished after selective disruption of disulfide bonds. Mol. Nutr. Food Res. 2012, 56, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Zheng, Y.; Liu, Z.; Deng, Y.; Zhao, Y. Structure and IgE-binding properties of alpha-casein treated by high hydrostatic pressure, UV-C, and far-IR radiations. Food Chem. 2016, 204, 46–55. [Google Scholar] [CrossRef]
- Shimakura, K.; Tonomura, Y.; Hamada, Y.; Nagashima, Y.; Shiomi, K. Allergenicity of crustacean extractives and its reduction by protease digestion. Food Chem. 2005, 91, 247–253. [Google Scholar] [CrossRef]
- Lv, L.T.; Lin, H.; Li, Z.X.; Ahmed, I.; Chen, G.Z. Determining the effect of malondialdehyde on the IgE-binding capacity of shrimp tropomyosin upon in vitro digestion. J. Sci. Food Agric. 2017, 97, 4588–4594. [Google Scholar] [CrossRef]
- Shriver, S.K. Effect of Selected Nonthermal Processing Methods on the Allergen Reactivity of Altantic White Shrimp (Litopenaeus setiferus). Master’s Thesis, University of Florida, Gainesville, FL, USA, 2011. [Google Scholar]
- Lee, J.W.; Kim, J.H.; Yook, H.S.; Kang, K.O.; Lee, S.Y.; Hwang, H.J.; Byun, M.W. Effects of gamma radiation on the allergenic and antigenic properties of milk proteins. J. Food Prot. 2001, 64, 272–276. [Google Scholar] [CrossRef]
- Wu, Y.T.; Lin, H.; Lu, Y.Y.; Huang, Y.H.; Dasanayaka, B.P.; Ahmed, I.; Chen, G.Z.; Chen, Y.; Li, Z.X. Allergenicity determination of Turbot parvalbumin for safety of fish allergy via dendritic cells, RBL-2H3 cell and mouse model. Eur. Food Res. Technol. 2021, 247, 1959–1974. [Google Scholar] [CrossRef]
- Zhang, Z.; Xiao, H.; Zhou, P. Allergenicity suppression of tropomyosin from Exopalaemon modestus by glycation with saccharides of different molecular sizes. Food Chem. 2019, 288, 268–275. [Google Scholar] [CrossRef]
- Teodorowicz, M.; Fiedorowicz, E.; Kostyra, H.; Wichers, H.; Kostyra, E. Effect of Maillard reaction on biochemical properties of peanut 7S globulin (Ara h 1) and its interaction with a human colon cancer cell line (Caco-2). Eur. J. Nutr. 2013, 52, 1927–1938. [Google Scholar] [CrossRef] [Green Version]
- Loschko, J.; Schreiber, H.A.; Rieke, G.J.; Esterhazy, D.; Meredith, M.M.; Pedicord, V.A.; Yao, K.H.; Caballero, S.; Pamer, E.G.; Mucida, D.; et al. Absence of MHC class II on cDCs results in microbial-dependent intestinal inflammation. J. Exp. Med. 2016, 213, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.M.; Yang, Y.; Maleki, S.J.; Alcocer, M.; Xu, S.S.; Shi, C.L.; Cao, M.J.; Liu, G.M. Anti-Food allergic activity of sulfated polysaccharide from gracilaria lemaneiformis is dependent on immunosuppression and inhibition of p38 MAPK. J. Agric. Food Chem. 2016, 64, 4536–4544. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Li, Z.X.; Wu, Y.T.; Guo, Y.M.; Pavase, T.R.; Chen, G.Z.; Zhang, Z.Y.; Lin, H. Comparison of immunological properties of recombinant and natural turbot (Scophthalmus maximus) parvalbumin. Eur. Food Res. Technol. 2021, 247, 2053–2065. [Google Scholar] [CrossRef]
- Gilfillan, A.M.; Tkaczyk, C. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 2006, 6, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Chen, G.; Ahmed, I.; Sun, L.; Li, W.; Pavase, T.R.; Li, Z. Immunostimulatory and allergenic properties of emulsified and non-emulsified digestion products of parvalbumin (Scophthalmus maximus) in RBL-2H3 cells and BALB/c mouse models. Food Funct. 2021, 12, 5351–5360. [Google Scholar] [CrossRef] [PubMed]
- Stagg, A.J. Intestinal dendritic cells in health and gut inflammation. Front. Immunol. 2018, 9, 2883. [Google Scholar] [CrossRef] [PubMed]
- Kaiko, G.E.; Horvat, J.C.; Beagley, K.W.; Hansbro, P.M. Immunological decision-making: How does the immune system decide to mount a helper T-cell response? Immunology 2008, 123, 326–338. [Google Scholar] [CrossRef]
- Summers, K.L.; Hock, B.D.; McKenzie, J.L.; Hart, D.N.J. Phenotypic characterization of five dendritic cell subsets in human tonsils. Am. J. Pathol. 2001, 159, 285–295. [Google Scholar] [CrossRef]
- Coombes, J.L.; Siddiqui, K.R.; Arancibia-Carcamo, C.V.; Hall, J.; Sun, C.M.; Belkaid, Y.; Powrie, F. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. J. Exp. Med. 2007, 204, 1757–1764. [Google Scholar] [CrossRef]
- Fu, L.; Song, J.; Wang, C.; Fu, S.; Wang, Y. Bifidobacterium infantis potentially alleviates shrimp tropomyosin-induced allergy by tolerogenic dendritic cell-dependent induction of regulatory T Cells and alterations in gut microbiota. Front. Immunol. 2017, 8, 1536. [Google Scholar] [CrossRef]
- Ding, L.; Wang, L.; Yu, Z.; Zhang, T.; Liu, J. Digestion and absorption of an egg white ACE-inhibitory peptide in human intestinal Caco-2 cell monolayers. Int. J. Food Sci. Nutr. 2016, 67, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huan, F.; Li, M.; Han, T.; Xia, F.; Yang, Y.; Liu, Q.; Chen, G.; Cao, M.; Liu, G. Mapping and IgE-binding capacity analysis of heat/digested stable epitopes of mud crab allergens. Food Chem. 2021, 344, 128735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, H.; Zhang, X.; Zhou, P. Conformation, allergenicity and human cell allergy sensitization of tropomyosin from Exopalaemon modestus: Effects of deglycosylation and Maillard reaction. Food Chem. 2019, 276, 520–527. [Google Scholar] [CrossRef]
- Jimenez-Saiz, R.; Chu, D.K.; Mandur, T.S.; Walker, T.D.; Gordon, M.E.; Chaudhary, R.; Koenig, J.; Saliba, S.; Galipeau, H.J.; Utley, A.; et al. Lifelong memory responses perpetuate humoral Th2 immunity and anaphylaxis in food allergy. J. Allergy Clin. Immunol. 2017, 140, 1604–1615.e1605. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Fu, S.; Wang, C.; Xie, M.; Wang, Y. Yogurt-sourced probiotic bacteria alleviate shrimp tropomyosin-induced allergic mucosal disorders, potentially through microbiota and metabolism modifications. Allergol. Int. 2019, 68, 506–514. [Google Scholar] [CrossRef]
- Lam, Y.F.; Tong, K.K.; Kwan, K.M.; Tsuneyama, K.; Shu, S.A.; Leung, P.S.; Chu, K.H. Gastrointestinal immune response to the shrimp allergen tropomyosin: Histological and immunological analysis in an animal model of shrimp tropomyosin hypersensitivity. Int. Arch. Allergy Immunol. 2015, 167, 29–40. [Google Scholar] [CrossRef]
- Chokshi, N.Y.; Sicherer, S.H. Interpreting IgE sensitization tests in food allergy. Expert Rev. Clin. Immunol. 2016, 12, 389–403. [Google Scholar] [CrossRef]
- Fornadley, J.A. Skin testing for inhalant allergy. Int. Forum Allergy Rhinol. 2014, 4, S41–S45. [Google Scholar] [CrossRef]
- Cabanillas, B.; Cuadrado, C.; Rodriguez, J.; Dieguez, M.C.; Crespo, J.F.; Novak, N. Boiling and pressure cooking impact on IgE reactivity of soybean allergens. Int. Arch. Allergy Immunol. 2018, 175, 36–43. [Google Scholar] [CrossRef]
- Lavilla, M.; Orcajo, J.; Diaz-Perales, A.; Gamboa, P. Examining the effect of High Pressure Processing on the allergenic potential of the major allergen in peach (Pru p 3). Innov. Food Sci. Emerg. Technol. 2016, 38, 334–341. [Google Scholar] [CrossRef]
- Gamez, C.; Zafra, M.P.; Sanz, V.; Mazzeo, C.; Ibanez, M.D.; Sastre, J.; del Pozo, V. Simulated gastrointestinal digestion reduces the allergic reactivity of shrimp extract proteins and tropomyosin. Food Chem. 2015, 173, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Lin, H.; Li, Z.; Nayak, B.; Ahmed, I.; Tian, S.; Chen, G.; Lin, H.; Zhao, J. Structural changes of 2,2’-azobis(2-amidinopropane) dihydrochloride (AAPH) treated shrimp tropomyosin decrease allergenicity. Food Chem. 2019, 274, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Perera, E.; Moyano, F.J.; Rodriguez-Viera, L.; Cervantes, A.; Martinez-Rodriguez, G.; Mancera, J.M. In vitro digestion of protein sources by crude enzyme extracts of the spiny lobster Panulirus argus (Latreille, 1804) hepatopancreas with different trypsin isoenzyme patterns. Aquaculture 2010, 310, 178–185. [Google Scholar] [CrossRef]
- Liu, G.M.; Cao, M.J.; Huang, Y.Y.; Cai, Q.F.; Weng, W.Y.; Su, W.J. Comparative study of in vitro digestibility of major allergen tropomyosin and other food proteins of Chinese mitten crab (Eriocheir sinensis). J. Sci. Food Agric. 2010, 90, 1614–1620. [Google Scholar] [CrossRef]
- FAO; WHO. Evaluation of Allergenicity of Genetically Modified Foods; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; pp. 1–26. [Google Scholar]
- Huang, Y.Y.; Liu, G.M.; Cai, Q.F.; Weng, W.Y.; Maleki, S.J.; Su, W.J.; Cao, M.J. Stability of major allergen tropomyosin and other food proteins of mud crab (Scylla serrata) by in vitro gastrointestinal digestion. Food Chem. Toxicol. 2010, 48, 1196–1201. [Google Scholar] [CrossRef]
- Liu, G.M.; Huang, Y.Y.; Cai, Q.F.; Weng, W.Y.; Su, W.J.; Cao, M.J. Comparative study of in vitro digestibility of major allergen, tropomyosin and other proteins between Grass prawn (Penaeus monodon) and Pacific white shrimp (Litopenaeus vannamei). J. Sci. Food Agric. 2011, 91, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.J.; Abbott, U.R.; Hatzos, C. Digestibility of food allergens and nonallergenic proteins in simulated gastric fluid and simulated intestinal fluid-a comparative study. J. Agric. Food Chem. 2002, 50, 7154–7160. [Google Scholar] [CrossRef] [PubMed]
- Inoue, R.; Sawai, T.; Sawai, C.; Nakatani, M.; Romero-Perez, G.A.; Ozeki, M.; Nonomura, K.; Tsukahara, T. A preliminary study of gut dysbiosis in children with food allergy. Biosci. Biotechnol. Biochem. 2017, 81, 2396–2399. [Google Scholar] [CrossRef]
- Cianci, R.; Pagliari, D.; Piccirillo, C.A.; Fritz, J.H.; Gambassi, G. The microbiota and immune system crosstalk in health and disease. Mediat. Inflamm. 2018, 2018, 2912539. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Sampson, H.A. Food allergy: Epidemiology, pathogenesis, diagnosis, and treatment. J. Allergy Clin. Immunol. 2014, 133, 291–307.e295. [Google Scholar] [CrossRef]
- Aitoro, R.; Paparo, L.; Amoroso, A.; Di Costanzo, M.; Cosenza, L.; Granata, V.; Di Scala, C.; Nocerino, R.; Trinchese, G.; Montella, M.; et al. Gut microbiota as a target for preventive and therapeutic intervention against food allergy. Nutrients 2017, 9, 672. [Google Scholar] [CrossRef] [PubMed]
- Malago, J.J. Contribution of microbiota to the intestinal physicochemical barrier. Benef. Microbes 2015, 6, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Stefka, A.T.; Feehley, T.; Tripathi, P.; Qiu, J.; McCoy, K.; Mazmanian, S.K.; Tjota, M.Y.; Seo, G.Y.; Cao, S.; Theriault, B.R.; et al. Commensal bacteria protect against food allergen sensitization. Proc. Natl. Acad. Sci. USA 2014, 111, 13145–13150. [Google Scholar] [CrossRef] [PubMed]
- Ohnmacht, C.; Park, J.H.; Cording, S.; Wing, J.B.; Atarashi, K.; Obata, Y.; Gaboriau-Routhiau, V.; Marques, R.; Dulauroy, S.; Fedoseeva, M.; et al. The microbiota regulates type 2 immunity through ROR gamma t(+) T cells. Science 2015, 349, 989–993. [Google Scholar] [CrossRef]
- Vatanen, T.; Kostic, A.D.; d’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hamalainen, A.M.; et al. Variation in microbiome LPS immunogenicity contributes to autoimmunity in humans. Cell 2016, 165, 842–853. [Google Scholar] [CrossRef]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by lachnospiraceae and ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Feehley, T.; Plunkett, C.H.; Bao, R.; Hong, S.M.C.; Culleen, E.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Yoon, W.; Lee, S.Y.; Shin, H.S.; Lim, M.Y.; Nam, Y.D.; Yoo, Y. Effects of lactobacillus pentosus in children with allergen-sensitized atopic dermatitis. J. Korean Med. Sci. 2020, 35, e128. [Google Scholar] [CrossRef]
- Muir, A.B.; Benitez, A.J.; Dods, K.; Spergel, J.M.; Fillon, S.A. Microbiome and its impact on gastrointestinal atopy. Allergy 2016, 71, 1256–1263. [Google Scholar] [CrossRef]
- Suther, C.; Moore, M.D.; Beigelman, A.; Zhou, Y. The gut microbiome and the big eight. Nutrients 2020, 12, 3728. [Google Scholar] [CrossRef]
Figure 1.
Analysis for structure and allergenicity changes of seafood allergens induced by non-thermal processing.
Figure 1.
Analysis for structure and allergenicity changes of seafood allergens induced by non-thermal processing.
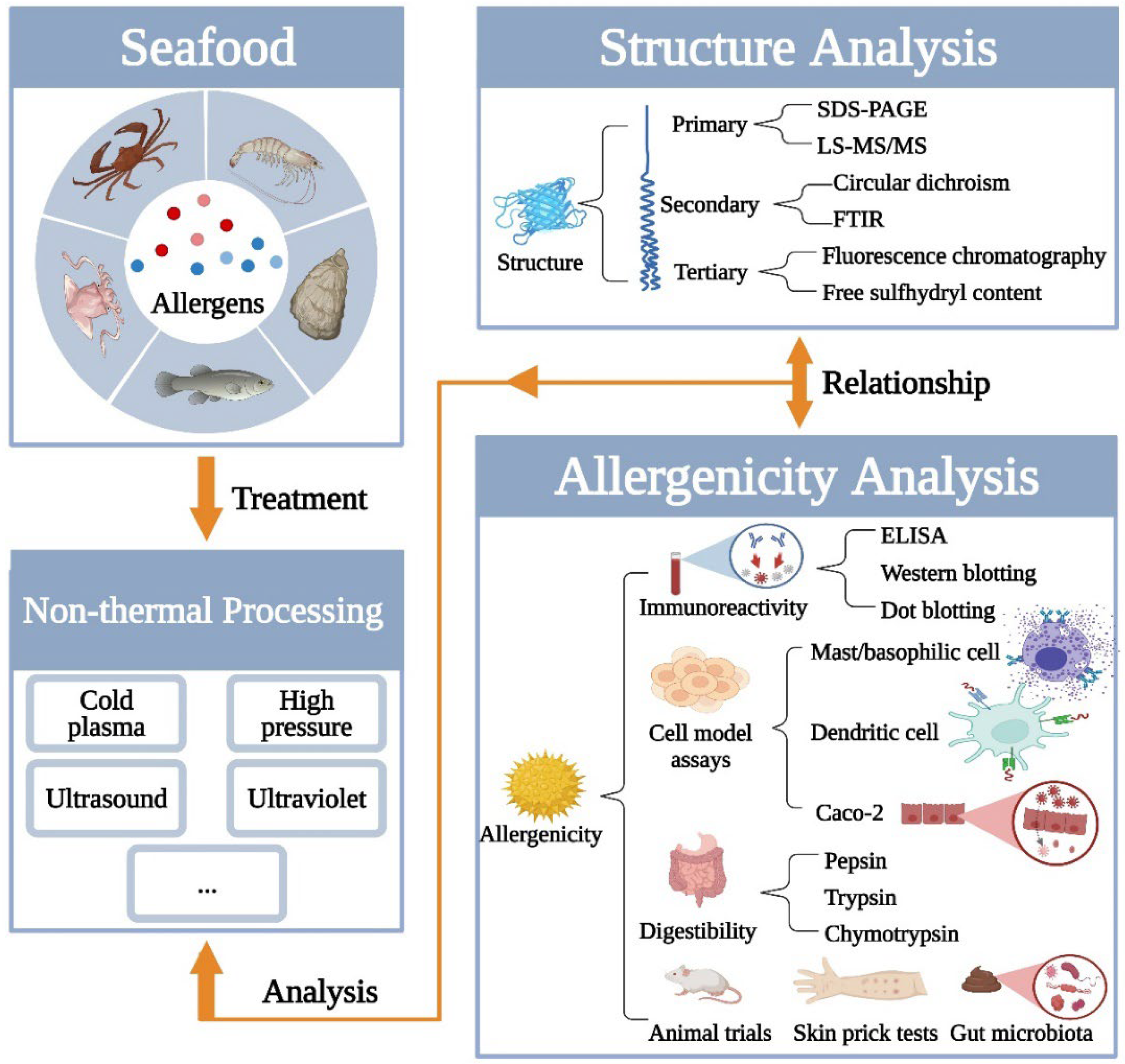
Figure 2.
The type of enzyme-linked immunosorbent assay (ELISA). Note: Ag: Antigen.

Figure 3.
Main mechanisms by which the breakdown of tolerance to food antigens can occur. When food allergens stimulate the mucosal system, the damaged epithelial cells will secrete cytokines, such as IL-33, to induce the differentiation of ILC 2 into Th 2, and secrete related cytokines, such as IL-13. Il-4 inhibits the differentiation of ILC 2 into Treg, ultimately leading to the production of allergen-specific IgE. IgE binds to Fcε R I fragments on mast cells to activate mast cell release mediators and induce food allergy symptoms.
Figure 3.
Main mechanisms by which the breakdown of tolerance to food antigens can occur. When food allergens stimulate the mucosal system, the damaged epithelial cells will secrete cytokines, such as IL-33, to induce the differentiation of ILC 2 into Th 2, and secrete related cytokines, such as IL-13. Il-4 inhibits the differentiation of ILC 2 into Treg, ultimately leading to the production of allergen-specific IgE. IgE binds to Fcε R I fragments on mast cells to activate mast cell release mediators and induce food allergy symptoms.
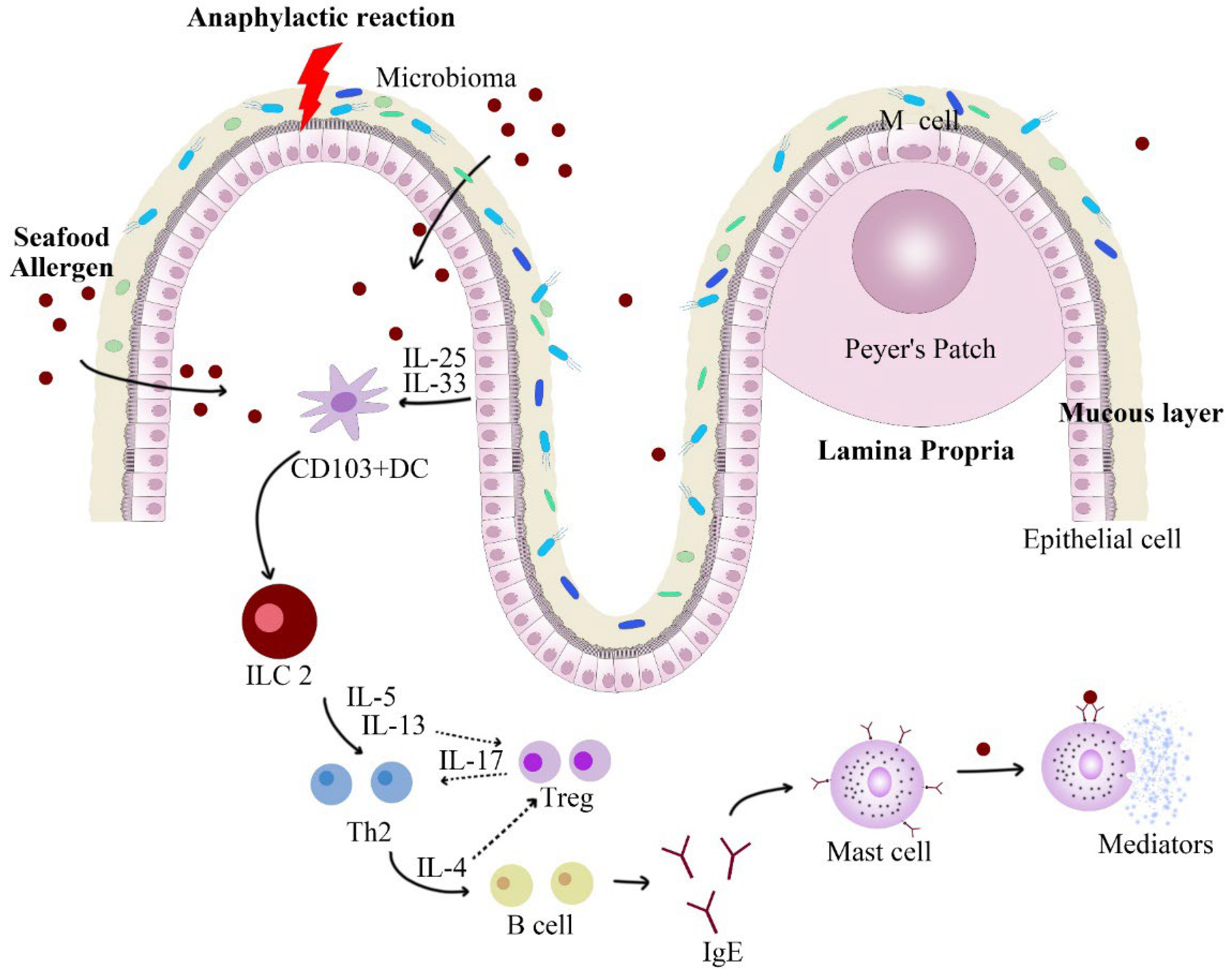
Figure 4.
The relationship between the big eight food allergens and gut microbiota [113]. The advantage of Ralstonia would promote the allergic risk of shellfish, while Dorea would reduce the risk. Moreover, Bacillus coagulans, Bifidobacterium infantis, and Bifidobacterium lactis have the potential for shellfish allergic reactions therapies.
Figure 4.
The relationship between the big eight food allergens and gut microbiota [113]. The advantage of Ralstonia would promote the allergic risk of shellfish, while Dorea would reduce the risk. Moreover, Bacillus coagulans, Bifidobacterium infantis, and Bifidobacterium lactis have the potential for shellfish allergic reactions therapies.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
An overview of studies evaluating the effects of non-thermal processing on the structure of seafood allergens.
Table 1.
An overview of studies evaluating the effects of non-thermal processing on the structure of seafood allergens.
| Non-Thermal Techniques | Target Allergen | Food Material | Treatment Conditions | Evaluations | Results | IgE-Binding Capacities/Immunoreactivity | References |
|---|---|---|---|---|---|---|---|
| High pressure processing (HPP) | Tropomyosin | Squid | 400 MPa, 20 °C, 10 min | SDS-PAGE | No changes | Decrease | [27] |
| CD | α-helix decrease, β-sheet increase | ||||||
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| Hemocyanin | Squid | 200–600 MPa, 25 °C, 20 min | SDS-PAGE | Band intensities decrease | Decrease | [11] | |
| FTIR | α-helix decrease, β-sheet increase | ||||||
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| SAXS | Rg decreased, 〈Nagg〉G and 〈Nagg〉Q increased | ||||||
| NS | Squid | 200–600 MPa, 20 min | SDS-PAGE | Band intensities decrease | NS | [33] | |
| NS | Fish | 200 MPa, 25 °C, 20 min | SDS-PAGE | New bands at 95 and 26 kDa | Decrease | [44] | |
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| Parvalbumin | Fish | 300–600 MPa, 10 min | CD | α-helix decrease | NS | [45] | |
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| Laser Raman spectroscopy | α-helix decrease, β-sheet increase | ||||||
| Actomyosin | Fish | 200–600 MPa, 10–50 min | SDS-PAGE | Band intensities decrease | NS | [30] | |
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| NS | Fish | 100–300 MPa, 20 °C, 10–60 min | SDS-PAGE | No changes | No changes | [46] | |
| CD | Structure changes | ||||||
| Tropomyosin | Shrimp | 100–600MPa, 0–30 min | / | / | Decrease | [47] | |
| Cold plasma (CP) | Tropomyosin | Shrimp | 3–15 min | SDS-PAGE | No changes with DTT, slightly fade without DTT | Decrease | [28] |
| CD | α-helix decrease, β-sheet increase | ||||||
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| Actomyosin | Shrimp | 1–5 min | SDS-PAGE | No changes | NS | [31] | |
| CD | α-helix decrease, β-turns increase | ||||||
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| Ultrasound | Total proteins | Shrimp | 400 W, 20 min | SDS-PAGE | Band intensities decrease | Decrease | [48] |
| FTIR | α-helix increase, β-sheet increase, β-turns decrease | ||||||
| Tropomyosin | Shrimp | 800 W, 30–180 min | SDS-PAGE | Band intensities decrease | Decrease | [49] | |
| Tropomyosin | Shrimp | 100–800 W, 15 min | SDS-PAGE | Band intensities decrease | Decrease | [39] | |
| CD | α-helix decrease, β-sheet and β-turns increase | ||||||
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| NS | Fish | 200–950 W, 60 min | SDS-PAGE | No changes | NS | [50] | |
| FSC | Decrease | ||||||
| SH | Increase | ||||||
| FS | Blue shifts | ||||||
| Ultraviolet (UV) | Tropomyosin | Shrimp | 4 min | SDS-PAGE | Band intensities decrease | Decrease | [51] |
| Tropomyosin | Shrimp | 0–6 min | SDS-PAGE | Band intensities decrease | Decrease | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, F.; Zhong, H.; Cheng, J.-H. Comprehensive Analysis of the Structure and Allergenicity Changes of Seafood Allergens Induced by Non-Thermal Processing: A Review. Molecules 2022, 27, 5857. https://doi.org/10.3390/molecules27185857
AMA Style
Wang F, Zhong H, Cheng J-H. Comprehensive Analysis of the Structure and Allergenicity Changes of Seafood Allergens Induced by Non-Thermal Processing: A Review. Molecules. 2022; 27(18):5857. https://doi.org/10.3390/molecules27185857
Chicago/Turabian StyleWang, Fengqi, Hangyu Zhong, and Jun-Hu Cheng. 2022. "Comprehensive Analysis of the Structure and Allergenicity Changes of Seafood Allergens Induced by Non-Thermal Processing: A Review" Molecules 27, no. 18: 5857. https://doi.org/10.3390/molecules27185857