
Nitro Fatty Acids (NO2-FAs): An Emerging Class of Bioactive Fatty Acids


1
Department of Chemistry, National and Kapodistrian University of Athens, Panepistimiopolis, 15771 Athens, Greece
2
Center of Excellence for Drug Design and Discovery, National and Kapodistrian University of Athens, 15771 Athens, Greece
*
Author to whom correspondence should be addressed.
Molecules 2021, 26(24), 7536; https://doi.org/10.3390/molecules26247536
Submission received: 6 November 2021
/
Revised: 8 December 2021
/
Accepted: 10 December 2021
/
Published: 13 December 2021
(This article belongs to the Special Issue Lipids–Lipolytic Enzymes–Lipid Analysis: A Themed Issue Dedicated to Prof. Edward A. Dennis for His 80th Birthday)
Abstract
:Unsaturated nitro fatty acids (NO2-FAs) constitute a category of molecules that may be formed endogenously by the reaction of unsaturated fatty acids (UFAs) with secondary species of nitrogen monoxide and nitrite anions. The warhead of NO2-FAs is a nitroalkene moiety, which is a potent Michael acceptor and can undergo nucleophilic attack from thiol groups of biologically relevant proteins, showcasing the value of these molecules regarding their therapeutic potential against many diseases. In general, NO2-FAs inhibit nuclear factorκ-B (NF-κB), and simultaneously they activate nuclear factor (erythroid derived)-like 2 (Nrf2), which activates an antioxidant signaling pathway. NO2-FAs can be synthesized not only endogenously in the organism, but in a synthetic laboratory as well, either by a step-by-step synthesis or by a direct nitration of UFAs. The step-by-step synthesis requires specific precursor compounds and is in position to afford the desired NO2-FAs with a certain position of the nitro group. On the contrary, the direct nitration of UFAs is not a selective methodology; thus, it affords a mixture of all possible nitro isomers.
1. Introduction
Nitro fatty acids (NO2-FAs) is a class of compounds that are endogenously formed by the reaction of unsaturated fatty acids (UFAs) [oleic (OA), linoleic (LA), etc.] with secondary products of nitrogen monoxide (NO.) and nitrite anions (NO2−) (Scheme 1), resulting in the formation of a reactive nitroalkene moiety [1,2,3]. These nitrated lipids have been detected and characterized in several tissues and biological fluids employing mass spectrometry-based approaches [4,5,6,7]. A recent review article summarizes the applications of such approaches to identify NO2-FAs and their protein adducts [8]. A very recent study defines the urine nitrolipidome, highlighting the presence of nitro derivatives of conjugated linoleic (cLA) and linolenic acid (cLnA) [9].
The endogenous formation of NO2-FAs begins with the formation of a β-nitroalkyl radical and its reaction to a nitronitrite or a nitrohydroxy intermediate (Scheme 2). The formation of NO2-FAs is achieved, when an elimination of nitrous acid of the nitronitrite ester or a dehydration of the nitrohydroxy compound takes place, respectively [3]. NO2-FAs have at least one nitrated double bond, granting in this way an electrophilic nature to the molecule and for that reason NO2-FAs can react with thiol groups very fast to generate reversible Michael addition products, due to the high reactivity of the electrophilic warhead of these molecules [10,11,12]. To clarify the way the cells signal with NO2-FAs and in order to better understand their pharmaceutical benefits, their biophysical properties and behavior in water have been studied by Vacek et al. [13].
NO2-FAs have been reported to form covalent adducts with cysteine residues of Kelch-like ECH-associated protein (Keap-1), resulting in activation of nuclear factor erythroid 2-related factor 2 (Nrf2). This activation of Nrf2 induces the expression of genes responsible for an overall antioxidant and cell-protecting function [14,15,16]. The covalent interactions between NO2-FAs and the Cys residues are reported to induce anti-inflammatory cell signaling as well [3]. The formation of covalent adducts between NO2-FAs and the protein of interest, via the formation of a new C-S bond, causes the protein to change its overall structure, therefore its function as well. This structural change of the protein is called Post-Translational Modification (PTM) (Figure 1) [3]. Understanding the way NO2-FAs interact with specific residues of the catalytic site of several proteins, residues beyond the catalytic pocket or even non-enzymatic proteins (such as structural proteins or transcriptional factors) each time, is of paramount importance for the Structure-Activity Relationship (SAR), in order to maximize the therapeutic benefits of NO2-FAs and minimize the harmful effects of the diseases.
NO2-FAs are also reported for alkylating important Cys residues of p65 and p50 subunits of nuclear factor κB (NF-κB), preventing the endogenous formation of a variety of pro-inflammatory mediators, responsible for diseases, like vascular inflammation and pro-fibrotic responses [17,18,19].
Besides the ability of the organism to synthesize by itself NO2-FAs during a state of inflammation, NO2-FAs can be synthesized in the laboratory. These nitroalkene-bearing compounds can be synthesized either by a direct nitration of the double bond of the corresponding UFA, where in that case an approximately 1:1 ratio of nitro regio-isomers are formed, or by a step-by-step synthesis. The step-by-step approach involves a Henry reaction between the appropriate aldehyde and the nitro component, depending on the desired nitration position. The final step of this synthetic pathway is an E2 elimination for the formation of the desired nitroalkene moiety, followed by an ester deprotection step. The various synthetic methods developed so far will be discussed extensively, along with the pros and cons of each method.
2. Bioactivities of NO2-FAs
2.1. Interaction of Cysteine Residues with NO2-FAs
Cysteine residues of several proteins may react with numerous electrophilic compounds, due to the high nucleophilic reactivity of the -SH group, in order to form covalent adducts via a Michael addition. The environment where the Cys residue is located plays a predominant role to define the pKa value of the -SH group, which for the residue Cys in aqueous solution at pH 7.2 is 8.33 [20]. The acidity of the -SH proton increases significantly when next to the Cys residue exist basic residues like His, Lys and Arg, metal centers from different enzymes (e.g. the iron metal center of hemoglobin), and different aromatic amino acids, such as Tyr and Trp (pi interaction with the thiol group) [20]. The above factors have as a result the formation of the highly reactive thiolate anion (RS−), which is a much better nucleophile in comparison with the free thiol group of the Cys residue to undergo oxidative or Michael reactions [21,22].
The covalent formation of adducts between several Cys residues and NO2-FAs play a pivotal role to many biochemical pathways with beneficial outcome for the organism. NO2-FAs exhibit potent anti-inflammatory effects in animal models of inflammation [16,17,18]. These electrophilic molecules interact with specific Cys residues of many regulatory proteins, like Keap-1, Nrf2, NF-κB, Peroxisome Proliferator-Activated Receptor γ (PPARγ), Heat Shock Proteins (HSP), 5-Lipoxygenase (5-LO) and others in order to regulate many diseases.
2.2. 10-Nitro Oleic Acid (CXA-10) and Its Derivatives as Anti-Inflammatory and Cytotoxic Agents
One of the most studied NO2-FAs is CXA-10 (1) (10-nitro oleic acid, 10-NO2-OA), which has entered Phase II clinical trial against Pulmonary Arterial Hypertension (PAH), Focal Segmental Glomerulosclerosis (FSGS) and asthma, by Complexa Inc., despite that it was first studied against acute kidney injury [23,24,25]. CXA-10 is the regio-isomer where the nitro group is located only at C10 of OA (Figure 2) exhibiting unique biological activity. Some of the pharmacokinetic (PK) and pharmacodynamic (PD) effects of oral CXA-10 in healthy and obese subjects were described by Jorkasky, et al. in 2019 [26].
NO2-FAs exhibit potent therapeutic properties in models of PAH caused by hypoxia or obesity [27,28]. More specifically, lung macrophage infiltration, expression of pro-inflammatory cytokines and glucose tolerance were improved significantly when NO2-OA was administrated to a PAH murine model. Characteristic lesions of PAH, like ventricular hypertrophy, fibrosis, etc., dropped significantly, unlike some biological markers like Monocyte Chemotactic Protein-1 (MCP-1), Interleukin-6 (IL-6), Tumor Necrosis Factor α (TNFα) and some macrophage markers (F4/80, CD68) levels in blood of High-Fat Diet (HFD) mice were increased. These HFD data showed that inflammation dropped significantly when 10-NO2-OA was administered, demonstrating once again the anti-inflammatory potency of 10-NO2-OA. NO2-FAs have been reported, as well to inhibit significantly the inflammation caused by bleomycin-mediated lung injury in mice by inhibiting Intratracheal Administration of Bleomycin (ITB)-induced pro-inflammatory mediators [29].
Neumann et al. have reported that CXA-10 increases significantly the antiproliferative effects of some known antineoplastic DNA-damaging agents, such as olaparib, cisplatin, doxorubicin against triple-negative breast cancer (TNBC) [30]. As a result, TNBC epithelial cell growth drops significantly by inhibiting the NF-κB signaling pathway. This signaling inhibition by CXA-10 happens due to the PTM effect on Cys residues to NF-κB subunit kinase β (IΚΚβ) and the NF-κB Re-1a protein, preventing this way DNA binding and an upcoming Re-1 protein-induced degradation of proteins. Moreover, CXA-10 inhibited TNFα-induced NF-κB activity, having as a result the suppression of harmful target genes, responsible for the expression of metastasis-related proteins like Intercellular Adhesion Molecule-1 (ICAM-1) and urokinase-type plasminogen activator [31].
A study in 2018 by Freeman and coworkers reported twenty new nitroalkenes as candidates for the activation of Antioxidant Response Element (ARE), which means activation of Nrf2 and inhibition of the NF-κB signaling pathway [32]. For better understanding of the importance of the nitroalkene position or the carbon chain length of the tested nitroalkenes, a variety of nitroalkenes was synthesized. That work aimed to clarify if the synthetic nitroalkenes activate ARE luciferase or inhibit TNF-α induced NF-κB signaling pathway. The importance ofthe terminal or interior position of the nitroalkene was examined as well. The nitroalkenes with the highest ARE activity are shown below, ranked from highest to lowest activity (Figure 3). It is noted that three out of five nitroalkenes have an ω-12 nitration position. Regarding the NF-κB inhibition, two nitroalkenes showed the same inhibition rate with NO2-OA and are shown below (Figure 4).
In 2018, Kühn et al. reported the other regio-isomer of CXA-10, 9-nitro oleic acid (9-NO2-OA) (9) (Figure 5) as a cytotoxic agent against colorectal cancer cell lines HCT-116 and HT-29. The cell viability of these cancer cell lines was suppressed significantly by 9-NO2-OA through a caspase-dependent apoptosis, via the intrisic apoptotic pathway [33]. This effect against cancer cell lines is based on the antioxidant effects of NO2-FAs and the capability of 9-NO2-OA to target mitochondria and deregulate their membrane and respiration, having as a result the final apoptosis of the cells. The entire cytotoxic effect is based on the Complex II inhibition by 9-NO2-OA, which is reversible and pH-dependent. As the pH gets more basic, the R-SH of the Cys residue deprotonates to the more reactive thiolate R-S−, giving this way a higher inhibition rate by 9-NO2-OA, since Michael additions are favored under basic conditions.
In order to understand whether the Michael acceptor is enough for the tested molecules to demonstrate cytotoxic effects against cancer cell lines, the combination of monomethyl fumarate (MMF) and 15-deoxy-delta-12,14-prostaglandin J2 (PGJ2) with HCT-116 cells was studied and compared with 9-NO2-OA’s cytotoxic effect [33]. The cell line was treated with all the three candidates for 48 h and the best results came from 9-NO2-OA, since it showed the highest cytotoxicity of all (IC50 6.5 μM), followed by PGJ2 (IC5017.6 μM), when MMF presented no cytotoxicity at all.
Most recently, fifteen NO2-FAs have been studied for their effects on activation of Nrf2, and inhibition of NF-κB signaling pathway, cyclooxygenase-2 (COX-2), LO and soluble Epoxide Hydrolase (sEH), as well as for their cytotoxic activity on colorectal cancer cells [34]. All the new NO2-FAs were compared with the lead compound 9-NO2-OA (9) (Figure 5).
The structures of NO2-FAs included in this SAR study are shown in Figure 6. The carbon chain length varies between twelve to twenty-two total carbon atoms and the nitro group is inserted from position C4 to C11 each time, close or away from the carboxylic group.
The inhibitory activity of these NO2-FAs against the NF-κB signaling pathway (p65 and p50) in colorectal cancer cells was studied in comparison with 9-NO2-OA [34]. The cytotoxic effect of the tested NO2-FAs against subunit p65 showed that compounds 22 and 23 demonstrated slightly worse activity than 9-NO2-OA. Compounds 14, 15 and 16 exhibited a stronger cytotoxicity than compound 9, with compound 14 being the most potent agent. The same results were demonstrated against subunit p50, where the only difference was the fact that compounds 14, 15 and 16 exhibited a slightly lower potency.
All of the tested NO2-FAs induced Nrf2, as it was demonstrated by Western blot and ELISA [34]. The majority of the tested compounds with a carbon chain length greater than sixteen, led to a significant Nrf2 gene expression. Also, NO2-FAs where the nitroalkene moiety position was greater than six, led to a 3-fold induction of Nrf2. As a negative experiment control, OA was ascertained that did not affect the Nrf2 gene expression at all.
The SAR study against LOs showed that compound 23 exhibited significant inhibition of 5-LO with an IC50 value of 0.3 ± 1.2 μM, while alternating the nitroalkene moiety position did not cause any significant result against 5-LO [34]. Compound 10 exhibited more potent inhibition of 5-LO (IC50 0.4 ± 1.2 μM) in comparison to 9-NO2-OA (9) (IC50 1.1 ± 1.2 μΜ). 12-LO was not inhibited at all, while 15-LO showed a weaker inhibition effect in comparison to 5-LO. The tested NO2-FAs showed an average inhibition of sEH and failed to inhibit COX-2 [34].
Freeman, et al. reported that invivo administration of NO2-OA at nanomolar levels led to the inhibition of atherosclerotic lesion formation and promotion of plaque stability to mice suffering from atherosclerosis [35]. Many antiatherogenic and cardioprotective ω-3 PUFAs [36], that can be endogenously nitrated by the reactive species of nitrogen monoxide, as described before, can explain the beneficial results of these ω-3 fatty acids against cardiovascular diseases. Human atherosclerosis is a typical vessel-derived disease, so NO2-FAs could be an alternative therapy in that case. NO2-OA has also been reported by Braumann et al. that can attenuate myocardial fibrosis in a murine model of dilated cardiomyopathy (DCM) [37].
Several reports state that NO2-OA might exert a beneficial and protective role against cardiovascular diseases [38,39,40]. Lipoprotein-associated phospholipase A2 (Lp-PLA2) is a cardiovascular risk biomarker, due to its link with coronary disease and stroke [41,42]. The way NO2-OA decreases the levels of Lp-PLA2 biomarker in blood can be explained by the simultaneous downregulation of cytokines, causing inflammation and the enchancement of SuperOxide Dismutase (SOD) activity. NO2-OA may be an alternative as a therapeutic compound for encountering Ventricular Tachykardia (VT), since a murine model demonstrated significantly reduced chance of appearing VT, after NO2-OA was administrated [43].
The induction of resistance to ischemic cardiac injury by NO2-FAs via the modulation of mitochondrial respiration at Complex II has been reported in 2016 by Schopfer et al. [44]. More specifically, NO2-OA inhibited in a reversible manner the respiration function of mitochondria cells in rat heart via interactions with Complex II, resulting in downregulation of the superoxide formation.
Another very important metabolic pathway for the regulation of inflammation is induced by cycloxygenase-2 (COX-2)-derived prostaglandins (PGs) and by the synthesis of leukotrienes (LTs), which are dependent on LO [45]. Some eicosanoids, like LTB4 and PGE2, modulate very strongly the activation of leukocytes, a type of cells of the immune system responsible for protecting the organism against infectious diseases and foreign microorganisms [46,47]. In order LTs to be synthesized, arachidonic acid (AA) must be hydrolysed from the sn-2 position of the membrane phospholipids by phospholipase A2 (PLA2), which is then oxidized by 5-LO giving rise to the synthesis of lipid mediators including LTs, PGs and lysophosphatidic acid [48,49]. 5-LO was found to be inhibited by the interaction of some electrophilic lipids with some important catalytic amino acid residues [50], so Awaad et al. studied if NO2-FAs can interact with 5-LO [51]. The PTM mechanism of NO2-OA against 5-LO was clarified with extending studies, including proteomics and mutagenesis. When 5-LO was treated with NO2-OA, covalent adducts between the nitroalkene and different histidine residues (125, 360, 362, 367 and 432) were reported. These residues did not show any adduct formation with OA. These His residues, forming adducts with NO2-OA, do not exist only in 5-LO, but they exist in 12-LO and 15-LO as well, which means NO2-FAs cannot be considered as selective inhibitors over 5-LO. Some Cys residues (159, 300, 416 and 418) consist of a building unit found only in 5-LO enzyme and are very important for the enzyme’s function [52]. A peptide chain of 5-LO, containing Cys 416 and 418, was alkylated by NO2-OA, resulting in an overall enzyme inhibition. In that study, nitro linoleic acid (NO2-LA) was examined as well to check its inhibitory potency against recombinant 5-LO along with NO2-OA. The IC50s of these two compounds against recombinant 5-LO were 0.3 ± 0.06 μM and 0.004 ± 0.002 μM for NO2-OA and NO2-LA, respectively. These IC50 values show a better inhibition of NO2-LA over NO2-OA in the case of recombinant 5-LO.
In 2016, Maucher et al. reported the use of CXA-10 against 5-LO [53], as Awwad did in 2014 [51]. They tested many electrophilic species against 5-LO, including mainly phenols, quinones, nitroalkenes and other electrophilic lipids [53]. They showed that CXA-10, as a potent Michael acceptor, has the capability of forming covalent adducts with the surface residues Cys416 and 418, modulating this way the structure of 5-LO. This structure modulation has the result of 5-LO inhibition and the prevention of the oxidation of AA and its upcoming inflammation storm.
2.3. PPARγ-Regulated Diseases Attenuated by NO2-FAs
PPARs consist of a group of nuclear receptor proteins whose role is the regulation of the expression of genes [54]. More specific, their regulating role is crucial for development, metabolism, cellular differentiation and tumorigenesis [55,56,57,58]. PPARγ is one of the four nuclear receptor proteins (PPARα, β/δ, γ) and is responsible for the fatty acid storage and the metabolism of glucose [59], the regulation of the differentiation of adipocyte [60], the increase of insulin sensitivity and the M2 macrophage activation in mice [61,62].
NO2-FAs may act as partial agonists of PPARγ [62,63,64]. PPARγ unit has the Cys285 residue that forms a covalent adduct with NO2-FAs, explaining the interaction between PPARγ and NO2-FAs. 9- and 10-NO2-OA are potent and selective activating ligands of PPARγ at physiological concentrations (100 nM), while PPARα and PPARδ require higher concentration levels of NO2-OA (~300 nM) to be activated. NO2-OA regio-isomers are reported to be responsible for the induction of PPARγ-dependent adipogenesis and the uptake of deoxyglucose in preadipocytes.
As stated before, NO2-FAs regulate some very important functionalities of the organism like metabolism of glucose, insulin sensitivity, regulation of adipocyte tissue and M2 macrophage activation [59,60,61,62]. For example, partial agonists NO2-OA and NO2-LA activate PPARγ and then CD36 protein expression is regulated, causing the M2 macrophage activation. Insulin sensitivity is increased by the upregulation of PPARγ-dependent expression of glucose transporter type 4 (GLUT4) and coactivator 1-alpha by the aforementioned NO2-FAs. The expression of GLUT4 and coactivator 1-alpha is responsible for the reduction of fat accumulation, pro-inflammatory cytokines and for the increase of mitochondrial mass. These are some beneficial properties of NO2-FAs which, as stated before, consist of partial agonists of PPARγ and not full agonists like rosiglitazone and pioglitazone. Rosiglitazone and pioglitazone are thiazolidinedione (TZD) drugs, which, during their administration, cause adverse side-effects like edema, adipogenesis and gaining fat, which are not caused when NO2-FAs are administrated in mice [40,65].
In 2010, Schopfer et al. showed that NO2-FAs can decrease the insulin and the glucose levels in mice via PPARγ activation, as TZD rosiglitazone does, but without the severe side-effects of rosiglitazone, which is mainly gaining weight [62]. The NO2-FAs tested in this work formed covalent adducts with Cys285 via a Michael addition. Derivatives of NO2-OA were found to be therapeutically efficient against metabolic syndrome in obese zucker rats [66]. Obese zucker rats (ob/ob rats) are mutant mice that eat excessively due to mutations in the gene, which is responsible for the production of leptin, a hormone responsible for the control of the appetite. Zucker rats are a model of type II diabetes. NO2-OA derivatives managed to decrease dramatically the food intake of these rats, a state which lasted during the whole experiment period (~14 days) and caused a significant decrease of weight in the obese rats. This treatment also decreased the triglycerides found in plasma and normalized the plasma free FAs.
Non-alcoholic fatty liver disease (NAFLD) is the most dominant disease that the liver suffers. NAFLD is a common disease to almost 90% of obese adults and correlates with diseases like obesity, type 2 diabetes and dyslipidemia. The most common mark that indicates NAFLD is the lipid accumulation inside the liver (steatosis) without consuming significant amount of alcohol. Non-alcoholic steatohepatitis (NASH) can lead to severe diseases (cirrhosis, hepatic carcinoma, liver failure) and finally to death [67]. The significance of the discovery of new potent drugs to overcome the harmful effects of NASH has emerged. Based on results from a series of clinical trials, the most potent drugs against NASH are TZDs as insulin sensitizers [68]. To fight NASH and encounter at the same time the side-effects of TZDs, electrophilic NO2-FAs have been found to be efficient agents against NAFLD. Freeman’s work in 2019 shows that the tested electrophilic NO2-FAs are promising drugs against NAFLD, without the adverse side-effects of TZDs [69]. ΝO2-OA has also contributed significantly to energy metabolism and to the inhibition of hepatic triglyceride accumulation [70]. In general, NO2-OA has been reported to benefit the organism to conditions of steatohepatitis and fibrosis.
In chronic kidney disease (CKD), a variety of harmful mechanisms, including oxidant stress and inflammation, correlated with dysregulation of NF-κB pathway and Nrf2 protein. From the other side, some protective mechanisms of the organism, which can be activated by NO2-FAs, can decrease these side-effects. In the study of Jorkasky in 2019 [71], 10-NO2-OA was evaluated for the treatment of uni-nephrectomized mice that were receiving deoxy-corticosterone acetate (DOCA)-high salt diet. 10-NO2-OA increased the heart and kidney weight of the mouse model, as well as it decreased nephrinuria, glomerular hypertrophy and glomerusclerosis. In this study, CXA-10 was compared with enalapril, an anti-hypertensive drug, and it showed some interesting results in which enalapril did not. Glomerular protection, anti-fibrotic and anti-inflammatory effects were observed in the case of CXA-10 but were not observed in the case of enalapril. The endogenous interaction of NO2-FAs with PPARs, whose role is known on the metabolism of glucose and lipids [72], may be the reason for all these beneficial effects of CXA-10 against CKD. Freeman and coworkers have reviewed the beneficial effects of NO2-FAs and other electrophilic compounds against several inflammatory and fibrotic diseases, discussing their characteristics and their pharmacology [73].
In vascular diseases—for example, ischemia-reperfusion injury (IRI or I/R)—it is of importance to regulate apoptosis. Nie and coworkers have demonstrated that CXA-10 is able to weaken OGD/R-induced apoptosis [72]. This effect was achieved by inhibition of Bax translocation and activation and an upcoming mitochondrial apoptotic cascade, and it was proved to be PPARγ-dependent.
In 2009, Gorczynski and coworkers reported the optimum structural features of NO2-FAs as PPARγ agonists, in order to activate PPARγ and exhibit all its beneficial effects, ready to decrease or remove possible inflammation or oxidation stress [74]. The main result of this work is that a 12-nitrated position of the unsaturated acid is essential for the optimum activation of PPARγ. Some NO2-FAs exhibited better ligand affinity than rosiglitazone [75]. Table 1 summarizes the IC50 values of various NO2-FAs measured by a scintillation proximity competitive binding assay. The structure of the optimum agonist, which induces the activation of PPARγ, is shown in Figure 7.
The importance of understanding the mechanism by which PPARγ is activated by agonists is of great importance for encountering inflammation and many other PPARγ-dependent systems. For example, PPARγ is reported to inhibit NF-κB dependent transcriptional activation in skeletal muscle [76]. The transcriptional activation of NF-κB in skeletal muscle is associated with skeletal muscle atrophy and insulin resistance and can be caused due to long-term NF-κB activation [77,78,79]. Thus, NO2-FAs could be alternative therapeutic agents for this type of disease. Administration of NO2-OA has also been shown to be beneficial against Abdominal Aortic Aneurysm (AAA), which is one of the diseases with the highest frequency of death globally [80].
In 2002, Karin and coworkers reviewed a correlation between NF-κB signaling pathway and cancer [81]. Disruption of the physiological regulation of NF-κB by the IKK protein complex (four subunits: IKKα, IKKβ, IKKγ, IKKδ) leads to NF-κB’s activity, which causes the transcription of inflammatory-genes, resulting in the generation of many inflammatory mediators. The NF-κB-induced triple negative breast cancer was also reported previously by Woodcock et al. [31]. PPARγ, as a modulator of cellular differentiation and regulator of tumorigenesis, was found to be expressed in many different types of cancer cells and its activation by agonists leads to either cell proliferation inhibition or cancel cell apoptosis [82,83]. The expression of PPARγ in many different types of cancer shows the complexity and the diversity of the biological action of how nitroalkenes overall attenuate inflammation and cancer [84].
2.4. NO2-FAs as Regulators of Miscellaneous Targets
NO2-FAs have been found to be responsible for the treatment of vascular endothelial cells, resulting in a dose-dependent upregulation of correlated HSPs (HSPA6, HPA1A, DNAJA4, HSPB8) by the expression of Heat Shock Factor 1 and 2 (HSF-1 and HSF-2) [15,85]. Heat Shock Proteins (HSPs) are a family of proteins, which are generated by cells under stress conditions. First, they were described as proteins associated only with heat shock [86], but several reports show a correlation with cold, UV light, and with processes of healing or tissue regeneration [87,88,89].
Another interesting bioactivity of NO2-FAs, demonstrated in a murine model, is the inhibition of sEH with the same way presented before, by forming covalent adducts with Cys521 via a Michael addition to the electrophilic double bond [90]. Administration of these electrophilic compounds results in the reduction of blood pressure to mice suffering from hypertension. Inhibition of sEH prevents hydrolysis of the epoxyeicosatrienoic acid substrates, so their accumulation induces vasodilation lowering blood pressure.
Xanthine oxidoreductase (XOR) is a molybdoflavin protein responsible for the oxidation of hypoxanthine to xanthine and finally to uric acid, resulting in the reduction of NAD+ to NADH [91]. XOR is the enzyme responsible for the regulation of the final process of purine catabolism and mainly generates oxidants responsible for the generation of inflammation and secondary species of nitrogen [92,93,94]. NO2-OA inhibits XOR with a low IC50 value (0.6 μM) and decreases significantly the generation of superoxide O2−. Superoxide is one of the Reactive Oxygen Species (ROS), which in oxidative stress its formation grows and causes harmful effects to proteins, lipids and nucleic acids [95].
As reported by Hansen et al. in 2018, NO2-FAs can be formed in response to virus infection and are able to inhibit stimulator of interferon genes (STING) signaling, leading this way to an anti-inflammatory activity against several diseases, such as, systemic lupus erythematosus, Aicardi-Goutiéres syndrome and STING-associated vasculopathy [96]. With this work, 9-NO2-OA, 10-NO2-OA and NO2-cLA are shown to be capable of targeting STING signaling and reduce the release of type I IFNs in murine and human cells. Thus, these endogenously formed nitrated compounds can overcome STING-induced inflammation.
NO2-FAs have also been demonstrated to be activators of Sirtuin 6 (SIRT6), a protein crucial for both glucose and lipid homeostasis, which is found to be actively involved in maintaining the DNA of the organism stable [97]. In this work, a SAR study using 9-NO2-OA (9), 10-NO2-OA (1), 9-NO2-cLA (25) and 12-NO2-cLA (26) (Figure 8) showed that these NO2-FAs are significant in vitro activators of SIRT6. The formation of a covalent adduct between NO2-FA and Cys18 of SIRT6, caused a change to the conformation of the protein, inducing this way a greater activation of SIRT6 (40-fold at 20 μΜ). This Cys18 residue is present only at the N terminus of SIRT6 and absent to other isoforms of the protein, making this the only way SIRT6 is able to react with these electrophilic compounds.
Pereckova and coworkers recently have shown that NO2-OA is involved in the regulation of stem cell pluripotency and differentiation of mouse Embryonic Stem Cells (mESC) via signal transducer and activator of transcription 3 (STAT3) signaling [98]. NO2-OA was found to modulate pluripotency of mESC via the regulation of STAT3 phosphorylation, as well as to attenuate cardiac differentiation and to guide mESC into neural fate.
A recent report by Mathers and coworkers demonstrated the effect of NO2-FAs on psoriasiform dermatitis [99]. Several murine models of psoriasis were employed in order to study the anti-inflammatory activity of NO2-OA. Oral administration of NO2-OA downregulated the formation of inflammatory cytokines in the skin, and in vitro experiments showed a decrease of the formation of basal IL-6 levels and IL-17A-induced expression of IL-6, via the NF-κB signaling pathway. Furthermore, NO2-OA downregulated the phosphorylation and nuclear translocation of STAT3 by nitroalkylating STAT3 and inhibiting this way the keratinocyte proliferation. By blocking NF-κB and STAT3, NO2-FAs may constitute new therapeutic agents to treat psoriasis.
A mammalian enzyme—namely, Prostaglandin Reductase-1 (PtGR-1), able to metabolize NO2-FAs in vivo—has been identified [100]. PtGR-1 reduces fatty acid nitroalkenes to nitroalkanes, deactivating them and silencing their signaling reactions.
3. Syntheses of NO2-FAs
As described before in Scheme 2, the parent mono- or poly-unsaturated compounds can be nitrated endogenously by the reactive species of nitrogen monoxide and generate nitroalkenes [1,2,3]. However, the importance of NO2-FAs and their potential applications to treat inflammatory diseases or cancer has led to the development of various strategies for their synthesis in the laboratory. The different routes, which involve either direct and non-selective nitration or step-by-step and regioselective nitration, are summarized in Scheme 3.
3.1. Direct Nitration of Unsaturated Lipids Yielding Derivatives of NO2-FAs
The first direct nitration of LA or its ethyl ester by exposure to nitrogen monoxide and oxygen in cyclohexane was reported in 1996 by D’Ischia (Scheme 4) [101].
GC-MS and NMR studies showed the synthesis of an inseparable mixture containing three isomeric compounds: nitroalkenes 28, 29 and nitronitrate derivative 30 (Scheme 4). The structure of these formed nitro derivatives was based on the characteristic for each isomer 1H-NMR data (Figure 9). This mixture of isomers started probably by a first addition of NO2 to one of the double bonds of ethyl linoleate, formed by a possible autoxidation of NO [102].
In 2000, D’Ischia studied the possible nitro products and by-products derived by the nitration of ethyl linoleate (27), yielding different isomers of nitroalkenes (Scheme 5).
The reactivity of this unsaturated ester over different nitrating conditions was extensively described; also, the NMR data of the different formed nitro isomers were recorded [103]. The employed nitrating agents in this work were nitronium tetrafluoroborate (NO2BF4), NO or a NO2− solution. Different regio-isomers of nitroalkenes were formed (compounds 29, 31–33), as well as different types of nitro by-products like: 3-nitro-1,5-hexadienes (34 and 35), 1,5-dinitro-1,3-pentadienes (36 and 37) and four different regio-isomers of nitro alcohols 38–41. In other experiments D’Ischia studied the monounsaturated fatty ester methyl oleate for its nitration reaction properties (Scheme 6). Mainly, a ratio 1:1 of (42 and 43):(44 and 45) was observed by the same nitration procedures followed for ethyl linoleate.
In 2008, D’Ischia reported a way to synthesize 9- and 10-NO2-LA, 48 and 52 respectively, by employing PhSeBr, HgCl2 and AgNO2 in dry MeCN, followed by the treatment of H2O2 in chloroform (Scheme 7) [104].
The synthesis of 10-NO2-LA is similar to that applied for the 9-nitro regio-isomer by only changing the free carboxyl group to an allyl ester, which gets cleaved in a final step by Pd(PPh3)4 and HCOOH in dry tetrahydrofuran (THF). During these studies, three decomposition products of 9-NO2-LA were observed, after incubation in a phosphate buffer of pH 7.4.
All three by-products were UV-Vis active in the TLC and were positive to Griess reagent, indicating the presence of a nitro group. By-products 53–55 are shown in Scheme 8.
When 10-NO2-LA 52 was exposed to aqueous phosphate buffer pH 7.4, only one decay product was observed. (12Z)-9-Hydroxy-10-nitrooctadec-12-enoic acid (56) was fully characterized by 1D and 2D NMR experiments and its structure is shown below in Scheme 9.
In 2013, Schopfer reported the synthesis of a mixture of 9- and 10-NO2-OA (9 and 1, respectively) following a modification of the method described by D’Ischia in 2008, where NaNO2 was used instead of AgNO2 (Scheme 10) [105]. After the consecutive additions for the formation of nitro-selenyl intermediates, the oxidative cleavage of PhSe-group takes place with 30% H2O2 in THF for 1 h.
The reaction mechanism (Scheme 11) begins with the nucleophilic attack of the double bond to the selenium atom, which affords the vicinal bromoselenium [106,107]. After intramolecular nucleophilic attack, a three membered ring, including the positively charged selenium atom, occurs.
Now the nitrating source (AgNO2 or NaNO2) has 1:1 chance to attack the two electrophilic carbon atoms of the three membered ring, resulting in a 1:1 regio-isomer ratio of the formed nitro-selenyl intermediates. The nitro-selenyl compound reacts with hydrogen peroxide to give a compound bearing an oxidized form of selenium, which can now undergo an intramolecular elimination, affording phenylselanol and the desired nitroalkene.
In 2021, Manolikakes and coworkers published a review article summarizing different nitration procedures of alkenes [108]. In this review, different nitrating and oxidizing agents are described, for the direct nitration of an alkene leading finally to the nitroalkenyl moiety, which constitutes the reactive electrophilic moiety of NO2-FAs.
3.2. Step-By-Step Synthesis of NO2-FAs
In 2006, Gorczynski and coworkers reported regio and stereospecific syntheses of (E)-9-NO2-OA and (E)-10-NO2-OA (9 and 1) in eight and nine steps, respectively [94,109]. The retrosynthetic analysis (Scheme 12) of these nitroalkenes showcases the need for the corresponding nitroalkanes or aldehydes, depending on the desired position of the nitro group on the double bond.
For the synthesis of 9-NO2-OA (Scheme 13), methyl 9-nitrononanoate (59) was synthesized in three steps by a Henry reaction between nitromethane and the corresponding aldehyde, an O-acetylation and a reduction using NaBH4 and MeOH (overall yield 74%). Compound 59 was used for a Henry reaction with nonanal under basic conditions. The isolated nitro hydroxy compound 62 was acetylated with acetic anhydride (Ac2O) and 4-dimethylaminopyridine (DMAP) in a first step and treatment with excess of DMAP led to the formation of the nitroalkene moiety. The final step was the acidic hydrolysis of the methyl ester to obtain 9-NO2-OA (9).
The synthesis of 10-NO2-OA (1) is similar with the synthetic pathway presented for the 9-regio-isomer. The required precursor compounds for the Henry reaction are 1-nitrononane (60) and methyl 9-oxononanoate (61) (Scheme 14). Compound 60 can be synthesized from the corresponding precursor aldehyde reacting with nitromethane to give the nitro hydroxy compound and then a subsequent reduction with NaBH4 affords 1-nitrononane.
As described in Gorczynski’s work, 9-oxononanoate (61) can be synthesized in three steps; a reduction of monomethyl azelaic acid with BH3-THF and a PCC oxidation affords one of the starting materials for the Henry reaction [109]. The reported yields for this NO2-FA at the late stages of the synthesis are slightly decreased compared with 9-NO2-OA.
In the same year, Branchaud and coworkers reported useful synthetic routes affording all four isomers of nitro oleic acids: (E), (Z), 9- and 10-NO2-OA [110]. The synthesis of (E)-9-NO2-OA started from 8-bromononan-1-ol (64), with a Jones oxidation, an esterification with allyl alcohol (65) and a nitration with AgNO2 in diethyl ether (Scheme 15). The synthesized nitro allyl ester 66 reacted with nonanal and 1,8-diazabicyclo(5.4.0)undec-7-ene (DBU) to give the nitro hydroxy ester 67 in a good yield. An acid-catalyzed acetylation using Ac2O and p-toluenesulfonic acid (TsOH) gave the nitro acetate compound, which under basic conditions and reflux in benzene afforded nitroalkene 68. A final cleavage of the allyl ester using Pd(PPh3)4 and HCOOH in THF gave 9-NO2-OA.
The needed precursors for the synthesis of 10-NO2-OA were synthesized as shown in Scheme 16. Nitration of 1-bromononane (69) with AgNO2 in diethyl ether gave compound 60. The needed aldehyde 70 was synthesized by ozonolysis of OA (57), followed by reductive workup with Me2S, resulting in a mixture of aldehydes. 9-Oxononanoic acid was isolated and its Steglich esterification with allyl alcohol using N,N’-dicyclohexylcarbodiimide (DCC)/DMAP afforded the desired allyl 9-oxononanoate.
A Henry condensation of 60 with 70 afforded the nitro hydroxy product 71 in 85% yield (Scheme 17). The three final steps of the synthesis are the same as described before with similar yields (85%, 74% and 95% respectively).
In this work, all four possible isomers, (E), (Z), 9-, 10- of NO2-OA were synthesized, isolated and fully characterized by NMR [110]. The NMR data showed that the (E) nitroalkene proton is shifted at 7.10 ppm, unlike the (Ζ) shift that appears at 5.70 ppm, in both 9- and 10-NO2-OA. These NMR shifts can be used as a diagnostic tool.
Branchaud’s protocol has the capability of forming (Z) nitroalkenes from their (E) isomers by first employing Ph2Se2, NaBH4 and acetic acid (AcOH), and in a second step H2O2, affording this way any isomer of the desired nitroalkenes (Scheme 18). The isomer ratio of the observed Z:E nitroalkenes was calculated by 1H-NMR. For compound 73, the isomer ratio was found to be 82:18 and for compound 74 was 85:15.
In 2008, Pellacani, et al. described a facile and highly stereoselective one-pot synthesis of either (E)- or (Z)-nitro alkenes by simply changing solvent and temperature [111]. Several aldehydes and nitroalkanes were combined to give either (E) or (Z) nitroalkenes by the employment of piperidine and molecular sieves. The first method employed CH2Cl2 at r. t. for 30 min and the second method utilized toluene under reflux for 4 h. This methodology was applied only for the formation of simple molecules bearing a nitroalkene moiety without any further functional groups. For example, propanal, pentanal, hexanal and isobutanal were used for a one-pot Henry condensation reaction with nitroethane or 1-nitropropane to afford either (E) or (Z) nitroalkenes.
In 2010, Zanoni reported a much more efficient synthetic route to (E)-12-nitrooctadec-12-enoic acid (24) in 4 steps (overall yield 50%), outperforming the previously reported methodology of Gorczynski [70], which had a 4.8% overall yield (Scheme 19) [112]. This high-yield synthesis of nitroalkene 24 began with a one-pot consecutive Henry reaction of 2-nitrocyclododecanone (75) with hexanal (76), followed by a retro-Claisen ring cleavage, affording this way the resulting nitro hydroxy acid 77. Esterification with diazomethane (CH2NH2) in CH2Cl2 afforded methyl ester 78, which, after dehydration by treatment with trifluoroacetic anhydride (TFAA), led to nitroalkene 79. In this final step of synthesis, Zanoni employed Candida Antartica lipase B (CAL-B) in order to smoothly hydrolyze the methyl ester and avoid the use of harsh conditions, as Gorczynski does. CAL-B is an insensitive pH enzyme, so it can cleave the methyl ester in an organic solvent, like methyl tert-butyl ether (MTBE) [113]. A minimum amount of water exists in the reaction, in order to cleave the intermediate acyl enzyme [114,115].
In 2016, Manolikakes and his coworkers presented the synthesis of a plethora of NO2-FAs [116]. After the preparation of the required starting compounds with either a Victor Meyer reaction or a Kornblum oxidation, the synthetic route began with a nitroaldol reaction between the nitroalkane and the used aldehyde each time, using a catalytic amount of 1,1,3,3-tetramethylguanidine (TMG) as the base (Scheme 20). The dehydration process was promoted by the Burgess reagent in benzene at 80 °C for 2 h, followed by a final deprotection of the allyl ester for the formation of the final NO2-FA. The E:Z 60:40 isomeric ratio for almost all examples turned into greater than 86:14, due to the isomerization of the double bond by the employed Pd(PPh3)4, HCOOH system.
This final isomerization afforded products with a high selectivity of E isomer over Z, providing to the method great selectivity with facile means. In total, seventeen different NO2-FAs, thirteen with the nitro group located close to the carboxyl group, and four with the nitro group far from it, were synthesized and their biological activity was left to be discovered.
Very recently Manolikakes and coworkers reported the one-pot synthesis of NO2-FAs by modifying their pre-existant synthetic methodology to a quicker and easier to use protocol (Scheme 21) [117]. In this methodology, prenyl esters and aldehydes were used as starting materials by using (i) TMG, (ii) TFAA, Et3N and (iii) BF3.OEt2 and affording the desired (E) NO2-FAs.
In 2021, Freeman reported the synthesis of 9- and 12-NO2-LA, as well as 9-nitrorumelenic acid (Scheme 22, Scheme 23 and Scheme 24) [118]. For the synthesis of 9-NO2-LA, starting from ω-bromo carboxylic acid 80 by employing oxalyl chloride (COCl)2, N,N-dimethylformamide (DMF) in tBuOH and then AgNO2 in Et2O, nitro compound 81 was obtained. Then, by employing non-2-enal (82) and Et3N, nitro hydroxy derivative 83 was obtained in 35% yield, which then reacted with TFAA, affording compound 84. Treatment of compound 84, first with Bu4NOAc and then with HCOOH, afforded 9-NO2-cLA (25) in 42% yield. For the synthesis of 12-NO2-LA, ω-UFA 85 was converted into the corresponding tert-butyl ester, and then treated with osmium tetroxide, N-methyl morpholine N-oxide (NMO), and NaIO4, to give aldehyde 86. After a Wittig and a Henry reaction, nitro hydroxy compound 89 was treated with TFAA to afford compound 90. Treatment with Bu4NOAc in Et2O and finally with formic acid afforded 12-NO2-cLA (26). 9-Nitrorumelenic acid (96) was successfully synthesized following similar synthetic steps.
In 2021, Zhang et al. reported the synthesis of 9-nitroeicos-9-en-19-ynoic acid (99), a derivative of 9-NO2-OA, where a terminal triple bond exists (Scheme 25) [119]. Compound 99 may serve as a tool of detecting in general which mammalian proteins can form covalent adducts with the electrophilic center of this NO2-FA. This accessible triple bond suggests a key to access multiple derivatives of the corresponding NO2-FA. The organic base used for the Henry reaction is TMG and the formation of the nitroalkene moiety started by using Ac2O and TsOH. After the initial acetylation reaction was over, Na2CO3 was used in toluene at 90 °C, in order to form the nitrated double bond.
4. Conclusions
NO2-FAs constitute a class of anti-inflammatory agents acting through the interaction with various important proteins, such as Keap-1, Nrf2, NF-κB, HSPs or the PPARγ receptor. Their main mode of interaction is by forming covalent adducts with cysteine residues of important biological targets via a reversible Michael addition. NO2-FAs can be endogenously synthesized by the reaction of UFAs with reactive species of nitrogen monoxide. Despite the endogenous nitration of UFAs, NO2-FAs may be obtained by the direct nitration of UFAs with several nitrating agents, mainly affording mixtures of the corresponding nitro regio-isomers. Another efficient way to obtain NO2-FAs, where the nitro group now has a specific position at the double bond, is by following a step-by-step strategic synthesis, starting from the required aldehydes and nitroalkanes. Efficient one-pot syntheses of NO2-FAs have been developed as well, introducing in this way an easier to use synthetic procedure for the synthesis of a variety of NO2-FAs.
Author Contributions
Conceptualization, G.K.; writing-review and editing, G.S.K. and G.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Stavros Niarchos Foundation (SNF) grant to the National and Kapodistrian University of Athens.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Dedicated to Edward A. Dennis for his 80th birthday. G.S.K. would like to thank SNF for a doctoral fellowship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Delmastro-Greenwood, M.; Hughan, K.S.; Vitturi, D.A.; Salvatore, S.R.; Grimes, G.; Potti, G.; Shiva, S.; Schopfer, F.J.; Gladwin, M.T.; Freeman, B.A.; et al. Nitrite and nitrate-dependent generation of anti-inflammatory fatty acid nitroalkenes. Free Radic. Biol. Med. 2015, 89, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Vitturi, D.A.; Minarrieta, L.; Salvatore, S.R.; Postlethwait, E.M.; Fazzari, M.; Ferrer-Sueta, G.; Lancaster, J.R., Jr.; Freeman, B.A.; Schopfer, F.J. Convergence of biological nitration and nitrosation via symmetrical nitrous anhydride. Nat. Chem. Biol. 2015, 11, 504–510. [Google Scholar] [CrossRef]
- Freeman, B.A.; Baker, P.R.S.; Schopfer, F.J.; Woodcock, S.R.; Napolitano, A.; D’Ischia, M. Nitro-fatty acid formation and signaling. J. Biol. Chem. 2008, 283, 15515–15519. [Google Scholar] [CrossRef] [Green Version]
- Lima, E.S.; Di Mascio, P.; Rubbo, H.; Abdalla, D.S.P. Characterization of linoleic acid nitration in human blood plasma by mass spectrometry. Biochemistry 2002, 41, 10717–10722. [Google Scholar] [CrossRef]
- Baker, P.R.; Lin, Y.; Schopfer, F.J.; Woodcock, S.R.; Groeger, A.L.; Batthyany, C.; Sweeney, S.; Long, M.H.; Iles, K.E.; Baker, L.M.S.; et al. Fatty acid transduction of nitric oxide signaling: Multiple nitrated unsaturated fatty acid derivatives exist in human blood and urine and serve as endogenous peroxisome proliferator-activated receptor ligands. J. Biol. Chem. 2005, 280, 42464–42475. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D.; Zoerner, A.A.; Mitschke, A.; Gutzki, F.-M. Nitro-fatty acids occur in human plasma in the picomolar range: A targeted nitro-lipidomics GC-MS/MS study. Lipids 2009, 44, 855–865. [Google Scholar] [CrossRef]
- Schoeman, J.C.; Harms, A.C.; van Weeghel, M.; Berger, R.; Vreeken, R.J.; Hankemeier, T. Development and application of a UHPLC–MS/MS metabolomics based comprehensive systemic and tissue-specific screening method for inflammatory, oxidative and nitrosative stress. Anal. Bioanal. Chem. 2018, 410, 2551–2568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, T.; Montero-Bullόn, J.-F.; Domingues, P.; Domingues, M.R. Discovery of bioactive nitrated lipids and nitro-lipid-protein adducts using mass spectrometry-based approaches. Redox Biol. 2019, 23, 101106. [Google Scholar] [CrossRef]
- Salvatore, S.R.; Rowart, P.; Schopfer, F.J. Mass spectrometry-based study defines the human urine nitrolipidome. Free Radic. Biol. Med. 2021, 162, 327–337. [Google Scholar] [CrossRef]
- Baker, L.M.S.; Baker, P.R.S.; Golin-Bisello, F.; Schopfer, F.J.; Fink, M.; Woodcock, S.R.; Branchaud, B.P.; Radi, R.; Freeman, B.A. Nitro-fatty acid reaction with glutathione and cysteine. Kinetic analysis of thiol alkylation by a Michael addition reaction. J. Biol. Chem. 2007, 282, 31085–31093. [Google Scholar] [CrossRef] [Green Version]
- Batthyany, C.; Schopfer, F.J.; Baker, P.R.; Duran, R.; Baker, L.M.; Huang, Y.; Cervenansky, C.; Branchaud, B.P.; Freeman, B.A. Reversible post-translational modification of proteins by nitrated fatty acids in vivo. J. Biol. Chem. 2006, 281, 20450–20463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turell, L.; Vitturi, D.A.; Coitiño, E.L.; Lebrato, L.; Möller, M.N.; Sagasti, C.; Salvatore, S.R.; Woodcock, S.R.; Alvarez, B.; Schopfer, F.J. The chemical basis of thiol addition to nitro-conjugated linoleic acid, a protective cell-signaling lipid. J. Biol. Chem. 2017, 292, 1145–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grippo, V.; Mojovic, M.; Pavicevic, A.; Kabelac, M.; Hubatka, F.; Turanek, J.; Zatloukalova, M.; Freeman, B.A.; Vacek, J. Electrophilic characteristics and aqueous behavior of fatty acid nitroalkenes. Redox Biol. 2021, 38, 101756. [Google Scholar] [CrossRef]
- Tsujita, T.; Li, L.; Nakajima, H.; Iwamoto, N.; Nakajima-Takagi, Y.; Ohashi, K.; Kawakami, K.; Kumagai, Y.; Freeman, B.A.; Yamamoto, M.; et al. Nitro-fatty acids and cyclopentenone prostaglandins share strategies to activate the Keap1-Nrf2 system: A study using green fluorescent protein transgenic zebrafish. Genes Cells 2011, 16, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Kansanen, E.; Jyrkkänen, H.K.; Volger, O.L.; Leinonen, H.; Kivelä, A.M.; Häkkinen, S.-K.; Woodcock, S.R.; Schopfer, F.J.; Horrevoets, A.J.; Ylä-Herttuala, S.; et al. Nrf2-dependent and –independent responses to nitro-fatty acids in human endothelial cells: Identification of heat shock response as the major pathway activated by nitro-oleic acid. J. Biol. Chem. 2009, 284, 33233–33241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, M.P.; Rudolph, T.K.; Khoo, N.K.H.; Motanya, U.N.; Golin-Bisello, F.; Wertz, J.W.; Schopfer, F.J.; Ruldoph, V.; Woodcock, S.R.; Bolisetty, S.; et al. Nitro-fatty acid inhibition of Neointima formation after endoluminal vessel injury. Circ. Res. 2009, 105, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Cui, T.; Schopfer, F.J.; Zhang, J.; Chen, K.; Ichikawa, T.; Baker, P.R.S.; Batthyany, C.; Chacko, B.K.; Feng, X.; Patel, R.P.; et al. Nitrated-fatty acids: Endogenous anti-inflammatory signaling mediators. J. Biol. Chem. 2006, 281, 35686–35698. [Google Scholar] [CrossRef] [Green Version]
- Villacorta, L.; Chang, L.; Salvatore, S.R.; Ichikawa, T.; Zhang, J.; Petrovic-Djergovic, D.; Jia, L.; Carlsen, H.; Schopfer, F.J.; Freeman, B.A.; et al. Electrophilic nitro-fatty acids inhibit vascular inflammation by disrupting LPS-dependent TLR4 signalling in lipid rafts. Cardiovasc. Res. 2013, 98, 116–124. [Google Scholar] [CrossRef]
- Ambrozova, G.; Martiskova, H.; Koudelka, A.; Ravekes, T.; Rudolph, T.K.; Klinke, A.; Rudolph, V.; Freeman, B.A.; Woodcock, S.R.; Kubala, L.; et al. Nitro-oleic acid modulates classical and regulatory activation of macrophages and their involvement in profibrotic responses. Free Radic. Biol. Med. 2016, 90, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, T.K.; Freeman, B.A. Transduction of redox signaling by electrophile-protein reactions. Sci. Signal 2009, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Netto, L.E.S.; Antonio de Oliveira, M.; Monteiro, G.; Demasi, A.P.D.; Cussiol, J.R.R.; Discola, K.F.; Demasi, M.; Silva, G.M.; Alves, S.V.; Faria, V.G.; et al. Reactive cysteine in proteins: Protein folding, antioxidant defence, redox signaling and more. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 180–193. [Google Scholar] [CrossRef]
- Derakhshan, B.; Hao, G.; Gross, S.S. Balancing reactivity against selectivity: The evolution of protein S-nitrosylation as an effector of cell signaling by nitric oxide. Cardiovasc. Res. 2007, 75, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Complexa. Complexa Announces Successful Completion of Four Phase 1 Studies of CXA-10 and Planned Phase 2 Initiation in Multiple Orphan Indications. Available online: https://www.complexarx.com/nov12 (accessed on 12 October 2021).
- Chieffo, C.; Botbyl, J.; Perry, K.; Blok, T.M.; Jorkasky, D.K. Use of an obese population in phase 1 to evaluate the pharmacology of oral CXA-10, an endogenous, nitrofatty acid singaling agent. In Proceedings of the Annual Meeting of the American College of Clinical Pharmacology, Bethesda, MD, USA, 24–27 September 2016. [Google Scholar]
- Clinical Trial of CXA-10 against Asthma, Lead Sponsor: University of Colorado, Denver; Anti Inflammatory Lipid Mediators in Asthma: A Double Blind Placebo Control Cross Over, Proof of Concept Study of CXA-10 to Reduce Bronchial Hyperresponsiveness in Obese Asthmatics. Available online: Ichgcp.net/clinical-trials-registry/NCT03762395 (accessed on 12 October 2021).
- Garner, R.M.; Mould, D.R.; Chieffo, C.; Jorkasky, D.K. Pharmacokinetic and pharmacodynamic effects of oral CXA-10, a nitro fatty acid, after single and multiple ascending doses in healthy and obese subjects. Clin. Transl. Sci. 2019, 12, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Klinke, A.; Μöller, A.; Pekarova, M.; Ravekes, T.; Friedrichs, K.; Berlin, M.; Scheu, K.M.; Kubala, L.; Kolarova, H.; Ambrozova, G.; et al. Protective effects of 10-nitro oleic acid in hypoxia-induced murine model of pulmonary hypertension. Am. J. Respir. Cell. Mol. Biol. 2014, 51, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, E.E.; Baust, J.; Bonacci, G.; Golin-Bisello, F.; Devlin, J.E.; St Croix, C.M.; Watkins, S.C.; Gor, S.; Cantu-Medellin, N.; Weidert, E.R.; et al. Fatty acid nitroalkenes ameliorate glucose intolerance and pulmonary hypertension in high-fat diet-induced obesity. Cardiovasc. Res. 2014, 101, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, M.L.; Abramova, E.; Guo, C.; Gow, J.G.; Murray, A.; Koudelka, A.; Cechova, V.; Freeman, B.A.; Gow, A.J. Fatty acid nitroalkenes inhibit the inflammatory response to bleomycin-mediated lung injury. Toxicol. Appl. Pharmacol. 2020, 407, 115236. [Google Scholar] [CrossRef] [PubMed]
- Alparslan, A.; Skoko, J.J.; Woodcock, C.-S.C.; Wingert, B.M.; Woodcock, S.R.; Normolle, D.; Huang, Y.; Stark, J.M.; Camacho, C.J.; Freeman, B.A.; et al. Electrophilic fatty acids impair RAD51 function and potentiate the effects of DNA-damaging agents on growth of triple-negative breast cells. J. Biol. Chem. 2018, 294, 397–404. [Google Scholar]
- Woodcock, C.-S.C.; Huang, Y.; Woodcock, S.R.; Salvatore, S.R.; Singh, B.; Golin-Bisello, F.; Davidson, N.E.; Neumann, C.A.; Freeman, B.A.; Wendell, S.G. Nitro-fatty acid inhibition of triple-negative breast cancer cell viability, migration, invasion and tumor growth. J. Biol. Chem. 2018, 293, 1120–1137. [Google Scholar] [CrossRef] [Green Version]
- Khoo, N.K.H.; Li, L.; Salvatore, S.R.; Schopfer, F.J.; Freeman, B.A. Electrophilic fatty acid nitroalkenes regulate Nrf2 and NF-κB signaling: A medicinal chemistry investigation of structure-function relationships. Sci. Rep. 2018, 8, 2295–2311. [Google Scholar] [CrossRef]
- Kühn, B.; Brat, C.; Fettel, J.; Hellmuth, N.; Maucher, I.V.; Bulut, U.; Hock, K.J.; Grimmer, J.; Manolikakes, G.; Rühl, M.; et al. Anti-inflammatory nitro-fatty acids suppress tumor growth by triggering mitochondrial dysfunction and activation of the intrinsic apoptotic pathway in colorectal cancer cell lines. Biochem. Pharmacol. 2018, 155, 48–60. [Google Scholar] [CrossRef]
- Hellmuth, N.; Brat, C.; Awad, O.; George, S.; Kahnt, A.; Bauer, T.; Phuoc, H.P.H.; Steinhilber, D.; Angioni, C.; Hassan, M.; et al. Structural modifications yield novel insights into the intriguing pharmacodynamic potential of anti-inflammatory nitro-fatty acids. Front. Pharmacol. 2021, 12, 715076. [Google Scholar] [CrossRef]
- Rudolph, T.K.; Rudolph, V.; Edreira, M.M.; Cole, M.P.; Bonacci, G.; Schopfer, F.J.; Woodcock, R.S.; Franek, A.; Pekarova, M.; Khoo, N.K.H.; et al. Nitro-fatty acids reduce atherosclerosis in apolipoprotein E-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 938–945. [Google Scholar] [CrossRef]
- Marchioli, R.; Barzi, F.; Bomba, E.; Chieffo, C.; Di Gregorio, D.; Di Mascio, R.; Franzosi, M.G.; Geraci, E.; Levantesi, G.; Maggioni, A.P.; et al. Early protection against sudden death by n-3 polyunsaturated fatty acids after myocardial infarction: Time-course analysis of the results of the Gruppo Italiano per lo Studio della Sopravvivenza nell’Infarto Miocardico (GISSI)-Prevenzione. Circulation 2002, 105, 1897–1903. [Google Scholar] [CrossRef] [Green Version]
- Braumann, S.; Schumacher, W.; Im, N.G.; Nettersheim, F.S.; Mehrkens, D.; Bokredenghel, S.; Hof, A.; Nies, R.J.; Adler, C.; Winkels, H.; et al. Nitro-oleic (NO2-OA) improves systolic function in dilated cardiomyopathy by attenuating myocardial fibrosis. Int. J. Mol. Sci. 2021, 22, 9052. [Google Scholar] [CrossRef]
- Wang, G.; Ji, Y.; Li, Z.; Han, X.; Guo, N.; Song, Q.; Quan, L.; Wang, T.; Han, W.; Pang, D.; et al. Nitro-oleic acid downregulates lipoprotein-associated phospholipase A2 expression via the p42/p44 MAPK and NFκB pathways. Sci. Rep. 2014, 4, 4905–4912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coles, B.; Bloodsworth, A.; Clark, S.R.; Lewis, M.J.; Cross, A.R.; Freeman, B.A.; O’Donnell, V.B. Nitrolinoleate inhibits superoxide generation, degranulation, and integrin expression by human neutrophils: Novel anti-inflammatory properties of nitric-oxide-derived reactives species in vascular cells. Circ. Res. 2002, 91, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, V.; Rudolph, T.K.; Schopfer, F.J.; Bonacci, G.; Woodcock, S.R.; Cole, M.P.; Baker, P.R.S.; Ramani, R.; Freeman, B.A. Endogenous generation and protective effects of nitro-fatty acids in a murine model of focal cardiac ischaemia and reperfusion. Cardiovasc. Res. 2010, 85, 155–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatine, M.S.; Morrow, D.A.; O’Donoghue, M.; Jablonski, K.A.; Rice, M.M.; Solomon, S.; Rosenberg, Y.; Domanski, M.J.; Hsia, J. Prognostic utility of lipoprotein-associated phospholipase A2 for cardiovascular outcomes in patients with stable coronary artery disease. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2463–2469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsimikas, S.; Willeit, J.; Knoflach, M.; Mayr, M.; Egger, G.; Notdurfter, M.; Witztum, J.L.; Wiedermann, C.J.; Xu, Q.; Kiechl, S. Lipoprotein-associated phospholipase A2 activity, ferritin levels, metabolic syndrome, and 10-year cardiovascular and non-cardiovascular mortality: Results from the Bruneck study. Eur. Heart J. 2009, 30, 107–115. [Google Scholar] [CrossRef]
- Mollenhauer, M.; Mehrkens, D.; Klinke, A.; Lange, M.; Remane, L.; Friedrichs, K.; Braumann, S.; Geißen, S.; Simsekyilmaz, S.; Nettersheim, F.S.; et al. Nitro-fatty acids suppress ischemic ventricular arrhythmias by preserving calcium homeostasis. Sci. Rep. 2020, 10, 15319. [Google Scholar] [CrossRef]
- Koenitzer, J.R.; Bonacci, G.; Woodcock, S.R.; Chen, C.-S.; Cantu-Medellin, N.; Kelley, E.E.; Schopfer, F.J. Fatty acid nitroalkenes induce resistance to ischemic cardiac injury by modulating mitochondrial respiration at complex II. Redox Biol. 2016, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Montuschi, P. Leukotrienes, antileukotrienes and asthma. Mini Rev. Med. Chem. 2008, 8, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.D. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef] [Green Version]
- Montuschi, P.; Peters-Golden, M.L. Leukotriene modifiers for asthma treatment. Clin. Exp. Allergy 2010, 40, 1732–1741. [Google Scholar] [CrossRef]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.A.; Cao, J.; Hsu, Y.-H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, R.W.; Gale, P.H. Inhibition of mammalian 5-lipoxygenase by aromatic disulfides. J. Biol. Chem. 1985, 260, 11554–11559. [Google Scholar] [CrossRef]
- Awwad, K.; Steinbrink, S.D.; Frömel, T.; Lill, N.; Isaak, J.; Häfner, A.-K.; Roos, J.; Hofmann, B.; Heide, H.; Geisslinger, G.; et al. Electrophilic fatty acid species inhibit 5-lipoxygenase and attenuate sepsis-induced pulmonary inflammation. Antioxid. Redox Signal 2014, 20, 2667–2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häfner, A.K.; Cernescu, M.; Hofmann, B.; Ermisch, M.; Hörnig, M.; Metzner, J.; Schneider, G.; Brutschy, B.; Steinhilber, D. Dimerization of human 5-lipoxygenase. Biol. Chem. 2011, 392, 1011–1097. [Google Scholar] [CrossRef] [Green Version]
- Maucher, I.V.; Rühl, M.; Kretschmer, S.B.M.; Hoffman, B.; Kühn, B.; Fettel, J.; Vogel, A.; Flügel, K.T.; Manolikakes, G.; Hellmuth, N.; et al. Michael acceptor containing drugs are a novel class of 5-lipoxygenase inhibitor targeting the surface cysteines C416 and C418. Biochem. Pharm. 2016, 25, 55–74. [Google Scholar] [CrossRef]
- Michalik, L.; Auwerx, J.; Berger, J.P.; Chatterjee, V.K.; Glass, C.K.; Gonzalez, F.J.; Grimaldi, P.A.; Kadowaki, T.; Lazar, M.A.; O’Rahilly, S.; et al. International Union of Pharmacology. LXI. Peroxisome proliferator-activated receptors. Pharmacol. Rev. 2006, 58, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Anastasi, M.R.; Zhang, V.J.; Russell, D.L.; Robker, R.L. Regulation of fatty acid oxidation in mouse cumulus-oocyte complexes during maturation and modulation by PPAR agonists. PLoS ONE 2014, 9, e87327. [Google Scholar]
- Belfiore, A.; Genua, M.; Malaguarnera, R. PPAR-gamma agonists and their effects on IGF-I receptor signaling: Implications for cancer. PPAR Res. 2009, 2009, 830501. [Google Scholar] [CrossRef] [Green Version]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef] [Green Version]
- Feige, J.N.; Gelman, L.; Michalik, L.; Desvergne, B.; Wahli, W. From molecular action to physiological outputs: Peroxisome proliferator-activated receptors are nuclear receptors at the crossroads of key cellular functions. Prog. Lipid Res. 2006, 45, 120–159. [Google Scholar] [CrossRef]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, E.-K.; Lee, Y.-R.; Kim, Y.-R.; Yeom, J.-Y.; Yoo, C.-H.; Kim, H.-K.; Park, H.-M.; Kang, H.-S.; Kim, J.-S.; Park, H.-M.; et al. NAADP mediates insulin-stimulated glucose uptake and insulin sensitization by PPARγ in adipocytes. Cell Rep. 2012, 2, 1607–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peluso, I.; Morabito, G.; Urban, L.; Ioannone, F.; Serafini, M. Oxidative stress in atherosclerosis development: The central role of LDL and oxidative burst. Endocr. Metab. Immune 2012, 12, 351–360. [Google Scholar] [CrossRef]
- Schopfer, F.J.; Cole, M.P.; Groeger, A.L.; Chen, C.S.; Khoo, N.K.; Woodcock, S.R.; Golin-Bisello, F.; Motanya, U.N.; Li, Y.; Zhang, J.; et al. Covalent peroxisome proliferator-activated receptor gamma adduction by nitro-fatty acids: Selective ligand activity and anti-diabetic signaling actions. J. Biol. Chem. 2010, 285, 12321–12333. [Google Scholar] [CrossRef] [Green Version]
- Villacorta, L.; Schopfer, F.J.; Zhang, J.; Freeman, B.A.; Chen, Y.E. PPAR-gamma and its ligands: Therapeutic implications in cardiovascular disease. Clin. Sci. 2009, 116, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Schopfer, F.J.; Lin, Y.; Baker, P.R.; Cui, T.; Garcia-Barrio, M.; Zhang, J.; Chen, K.; Chen, Y.E.; Freeman, B.A. Nitrolinoleic acid: An endogenous peroxisome proliferator-activated receptor gamma ligand. Proc. Natl. Acad. Sci. USA 2005, 102, 2340–2345. [Google Scholar] [CrossRef] [Green Version]
- Baker, P.R.; Schopfer, F.J.; Sweeney, S.; Freeman, B.A. Red cell membrane and plasma linoleic acid nitration products: Synthesis, clinical identification and quantitation. Proc. Natl. Acad. Sci. USA 2004, 101, 11577–11582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liu, H.; Jia, Z.; Guan, G.; Yang, T. Effects of endogenous PPAR agonist nitro-oleic acid on metabolic syndrome in obese zucker rats. PPAR Res. 2010, 2010, 601562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernon, G.; Baranova, A.; Younossi, Z.M. Systematic review: The epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Aliment. Pharmacol. Ther. 2011, 34, 274–285. [Google Scholar] [CrossRef]
- Ratziu, V. Pharmacological agents for NASH. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 676–685. [Google Scholar] [CrossRef]
- Khoo, K.H.; Fazzari, M.; Chartoumpekis, D.; Li, L.; Guimaraes, D.A.; Arteel, G.E.; Shiva, S.; Freeman, B.A. Electrophilic nitro-oleic acid reverses obesity-induced hepatic steatosis. Redox Biol. 2019, 22, 101132. [Google Scholar] [CrossRef] [PubMed]
- Rom, O.; Xu, G.; Guo, Y.; Zhu, Y.; Wang, H.; Zhang, J.; Fan, Y.; Liang, W.; Lu, H.; Liu, Y.; et al. Nitro-fatty acids protect against steatosis and fibrosis during development of nonalcoholic fatty liver disease in mice. EBioMedicine 2019, 14, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Arbeeny, C.M.; Ling, H.; Smith, M.M.; O’Brien, S.; Wawersik, S.; Ledbetter, S.R.; McAlexander, A.; Schopfer, F.J.; Wilette, R.N.; Jorkasky, D.K. CXA-10, a nitrated fatty acid, is renoprotective in deoxycorticosterone acetate-salt nephropathy. J. Pharmacol. Exp. Ther. 2019, 369, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Nie, H.; Xue, X.; Li, J.; Liu, X.; Shasha, L.; Guan, G.; Liu, H.; Liu, S.; Chen, Z. Nitro-oleic acid attenuates OGD/R-triggered apoptosis in renal tubular cells via inhibition of Bax mitochrondrial translocation in a PPAR-γ-dependent manner. Cell Physiol. Biochem. 2015, 35, 1201–1218. [Google Scholar] [CrossRef]
- Schopfer, F.J.; Vitturi, D.A.; Jorkasky, D.K.; Freeman, B.A. Nitro-fatty acids: New drug candidates for chronic inflammatory and fibrotic diseases. Nitric Oxide 2018, 79, 31–37. [Google Scholar] [CrossRef]
- Gorczynski, M.J.; Smitherman, P.K.; Akiyama, T.E.; Wood, H.B.; Berger, J.P.; King, B.S.; Morrow, C.S. Activation of Peroxisome Proliferator-Activated Receptor γ (PPARγ) by nitroalkene fatty acids: Importance of nitration position and degree of unsaturation. J. Med. Chem. 2009, 52, 4631–4639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, R.L.; Wright, M.W.; Gorcjynski, M.J.; Smitherman, P.K.; Akiyama, T.E.; Wood, H.B.; Berger, J.P.; King, B.S.; Morrow, C.S. Differential potencies of naturally occurring regio-isomers of nitrolinoleic acid in PPARγ activation. Biochemistry 2009, 48, 492–498. [Google Scholar] [CrossRef]
- Remels, A.H.V.; Langen, R.C.J.; Gosker, H.R.; Russell, A.P.; Spaapen, F.; Voncken, J.W.; Schrauwen, P.; Schols, W.J. PPARγ inhibits NF-κB dependent transcriptional activity in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2009, 297, 174–183. [Google Scholar] [CrossRef]
- Friedman, J.E.; Kirwan, J.P.; Jing, M.; Presley, L.; Catalano, P.M. Increased skeletal muscle tumor necrosis factor-alpha and impaired insulin signaling persist in obese women with gestational diabetes mellitus 1 year postpartum. Diabetes 2008, 57, 606–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappas, M.; Permezel, M.; Rice, G.E. Release of proinflammatory cytokines and 8-isoprostane from placenta, adipose tissue, and skeletal muscle from normal pregnant women and women with gestational diabetes mellitus. J. Clin. Endocrinol. Metab. 2004, 89, 5627–5633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sriwijitkamol, A.; Christ-Roberts, C.; Berria, R.; Eagan, P.; Pratipanawatr, T.; DeFronzo, R.A.; Mandarino, L.J.; Musi, N. Reduced skeletal muscleinhibitor of κBβcontent is associated with insulin resistance insubjects with type 2 diabetes: Reversal by exercise training. Diabetes 2006, 55, 760–767. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Chang, Z.; Zhao, G.; Lu, H.; Xiong, W.; Liang, W.; Wang, H.; Villacorta, L.; Garcia-Barrio, M.T.; Zhu, T.; et al. Suppression of vascular macrophage activation by nitro-oleic acid and its implication for abdominal aortic aneurysm therapy. Cardiovasc. Drugs Ther. 2021, 35, 939–951. [Google Scholar] [CrossRef]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.-W. NF-κB in cancer: From innocent bystander to major culprit. Nat. Rev. 2002, 2, 301–310. [Google Scholar] [CrossRef]
- Grommes, C.; Landreth, G.E.; Heneka, M.T. Antineoplastic effects of peroxisome proliferator-activated receptor γ agonists. Lancet Oncol. 2004, 5, 419–429. [Google Scholar] [CrossRef]
- Theocharis, S.; Margeli, A.; Vielh, P.; Kouraklis, G. Peroxisome proliferator activated receptor-γ ligands as cell cycle modulators. Cancer Treat. Rev. 2004, 30, 545–554. [Google Scholar] [CrossRef]
- Tachibana, K.; Yamasaki, D.; Ishimoto, K.; Doi, T. The role of PPARs in cancer. PPAR Res. 2008, 2008, 102737. [Google Scholar] [CrossRef] [Green Version]
- Schopfer, F.J.; Cipollina, C.; Freeman, B.A. Formation and signaling actions of electrophilic lipids. Chem. Rev. 2012, 111, 5997–6021. [Google Scholar] [CrossRef] [Green Version]
- Ritossa, F. A new puffing pattern induced by temperature shock and DNP in drosophila. Experientia 1962, 18, 571–573. [Google Scholar] [CrossRef]
- Matz, J.M.; Blake, M.J.; Tatelman, H.M.; Lavoi, K.P.; Holbrook, N.J. Characterization and regulation of cold-induced heat shock protein expression in mouse brown adipose tissue. Am. J. Physiol. 1995, 269, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Ohwatari, N.; Matsumoto, T.; Kosaka, M.; Ohtsuru, A.; Yamashita, S. TGF-beta1 mediates 70-kDa heat shock protein induction due to ultraviolet irradiation in human skin fibroblasts. Pflüg. Arch. 1999, 438, 239–244. [Google Scholar] [CrossRef]
- Laplante, A.F.; Moulin, V.; Auger, F.A.; Landry, J.; Li, H.; Morrow, G.; Tanguay, R.M.; Germain, L. Expression of heat shock proteins in mouse skin during wound healing. J. Histochem. Cytochem. 1998, 46, 1291–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charles, R.L.; Rudyk, O.; Prysyazhna, O.; Kamynina, A.; Yang, J.; Morisseau, C.; Hammock, B.D.; Freeman, B.A.; Eaton, P. Protection from hypertension in mice by the Mediterranean diet is mediated by nitro fatty acid inhibition of soluble epoxide hydrolase. Proc. Natl. Acad. Sci. USA 2014, 111, 8167–8172. [Google Scholar] [CrossRef] [Green Version]
- Kelley, E.E.; Batthyany, C.I.; Hundley, N.J.; Woodcock, S.R.; Bonacci, G.; Del Rio, J.M.; Schopfer, F.J.; Lancaster, J.R., Jr.; Freeman, B.A.; Tarpey, M.M. Nitro-oleic acid, a novel and irreversible inhibitor of xanthine oxidoreductase. J. Biol. Chem. 2008, 283, 36176–36184. [Google Scholar] [CrossRef] [Green Version]
- Kelley, E.E.; Khoo, N.K.; Hundley, N.J.; Malik, U.Z.; Freeman, B.A.; Tarpey, M.M. Hydrogen peroxide is the major oxidant product of xanthine oxidase. Free Radic. Biol. Med. 2010, 48, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, E.E.; Trostchansky, A.; Rubbo, H.; Freeman, B.A.; Radi, R.; Tarpey, M.M. Binding of xanthine oxidase to glycosaminoglycans limits inhibition by oxypurinol. J. Biol. Chem. 2004, 279, 37231–37234. [Google Scholar] [CrossRef] [Green Version]
- Freeman, A.B.; Pekarova, M.; Rubbo, H.; Trostchansky, A. Electrophilic nitro-fatty acids: Nitric oxide and nitrite-derived metabolic and inflammatory signaling mediators. Nitric Oxide 2017, 213–229. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxi. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.L.; Buchan, G.J.; Rühl, M.; Mukai, K.; Salvatore, S.R.; Ogawa, E.; Andersen, S.D.; Iversen, M.B.; Thielke, A.L.; Gunderstofte, C.; et al. Nitro-fatty acids are formed in response to virus infection and are potent inhibitors of STING palmitoylation and signaling. Proc. Natl. Acad. Sci. USA 2018, 115, 7768–7775. [Google Scholar] [CrossRef] [Green Version]
- Carreño, M.; Bresque, M.; Machado, M.R.; Santos, L.; Durán, R.; Vitturi, D.A.; Escande, C.; Denicola, A. Nitro-fatty acids as activators of hSIRT6 deacetylase activity. J. Biol. Chem. 2020, 295, 18355–18366. [Google Scholar] [CrossRef]
- Pereckova, J.; Pekarova, M.; Szamecova, N.; Hoferova, Z.; Kamarytova, K.; Falk, M.; Perecko, T. Nitro-oleic acid inhibits stemness maintenance and enhances neural differentiation of mouse embryonic stem cells via STAT3 signaling. Int. J. Mol. Sci. 2021, 22, 9981. [Google Scholar] [CrossRef]
- Wang, P.; Killeen, M.E.; Sumpter, T.L.; Ferris, L.K.; Falo, L.D., Jr.; Freeman, B.A.; Schopfer, F.J.; Mathers, A.R. Electrophilic nitro-fatty acids suppress psoriasiform dermatitis: STAT3 inhibition as a contributory mechanism. Redox Biol. 2021, 43, 101987. [Google Scholar] [CrossRef] [PubMed]
- Vitturi, D.A.; Chen, C.S.; Woodcock, S.R.; Salvatore, S.R.; Bonacci, G.; Koenitzer, J.R.; Stewart, N.A.; Wakabayashi, N.; Kensler, T.W.; Freeman, B.A.; et al. Modulation of nitro-fatty acid signaling: Prostaglandin reductase-1 is a nitroalkene reductase. J. Biol. Chem. 2013, 288, 25626–25637. [Google Scholar] [CrossRef] [Green Version]
- D’Ischia, M. Oxygen-dependent nitration of ethyl linoleate with nitric oxide. Tetrahedron Lett. 1996, 37, 5773–5774. [Google Scholar] [CrossRef]
- Pryor, W.A.; Lightsey, J.W.; Church, D.F. Reaction of nitrogen dioxide with alkenes and polyunsaturated fatty acids: Addition and hydrogen-abstraction mechanisms. J. Am. Chem. Soc. 1982, 104, 6685–6692. [Google Scholar] [CrossRef]
- Napotalino, A.; Camera, E.; Picardo, M.; D’Ischia, M. Acid-promoted reactions of ethyl linoleate with nitrite ions: Formation and structural characterization of isomeric nitroalkene, nitrohydroxy, and novel 3-Nitro-1,5-hexadiene and 1,5-dinitro-1,3-pentadiene products. J. Org. Chem. 2000, 65, 4583–4860. [Google Scholar]
- Manini, P.; Capelli, L.; Reale, S.; Arzillo, M.; Crescenzi, O.; Napolitano, A.; Barone, V.; D’Ischia, M. Chemistry of nitrated lipids: Remarkable instability of 9-nitrolinoleic acid in neutral aqueous medium and a novel nitronitrate ester product by concurrent autoxidation/nitric oxide-release pathways. J. Org. Chem. 2008, 73, 7517–7525. [Google Scholar] [CrossRef]
- Woodcock, S.R.; Bonacci, G.; Gelhaus, S.L.; Schopfer, F.J. Nitrated fatty acids: Synthesis and measurement. Free Radic. Biol. Med. 2013, 59, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayama, T.; Tomoda, S.; Takeuchi, Y.; Nomura, Y. Synthesis of 2-nitroalkyl phenyl selenides and their conversion to nitroalkenes. Chem. Lett. 1982, 11, 1109–1112. [Google Scholar] [CrossRef]
- Acevedo, C.M.; Kogut, E.F.; Lipton, M.A. Synthesis and analysis of the sterically constrained L-glutamine analogues (3S,4R)-3,4-dimethyl-L-glutamine and (3S,4R)-3,4-dimethyl-L-pyroglutamic acid. Tetrahedron 2001, 57, 6353–6359. [Google Scholar] [CrossRef]
- Hassan, M.; Nde, C.N.; Manolikakes, G. Synthesis of nitroolefins via the direct nitration of alkenes. SynOpen 2021, 5, 229–231. [Google Scholar]
- Gorczynski, M.J.; Huang, J.; King, B. Regio- and Stereospecific syntheses and nitric oxide donor properties of (E)-9 and (E)-10-nitrooctadec-9-enoic acids. Org. Lett. 2006, 8, 2305–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodcock, S.R.; Marwitz, A.J.V.; Bruno, P.; Branchaud, B.P. Synthesis of nitrolipids. All four possible diastereomers of nitrooleic acids: (E)- and (Z)-, 9- and 10-nitro-octadec-9-enoic acids. Org. Lett. 2006, 8, 3931–3934. [Google Scholar] [CrossRef]
- Fioravanti, S.; Pellacani, L.; Tardella, P.A.; Vergari, M.C. Facile and highly stereoselective one-pot synthesis of either (E)- or (Z)-nitro alkenes. Org. Lett. 2008, 10, 1449–1451. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, G.; Bendjeddou, M.V.L.; Porta, A.; Bruno, P.; Vidari, G. Improved synthesis of (E)-12-nitrooctadec-12-enoic acid, a potent PPARγ activator. Development of a “Buffer-free” enzymatic method for hydrolysis of methyl esters. J. Org. Chem. 2010, 75, 8311–8314. [Google Scholar] [CrossRef]
- Uppenberg, J.; Hansen, H.P.; Patkar, S.; Jones, T.A. The sequence, crystal structure determination and refinement of two crystal forms of lipase B from Candida Antarctica. Structure 1994, 2, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Kirk, O.; Christensen, M.W. Lipases from Candida antarctica: Unique biocatalysts from a unique origin. Org. Proc. Res. Dev. 2002, 6, 446–451. [Google Scholar] [CrossRef]
- Branco, R.J.F.; Graber, M.; Denis, V.; Pleiss, J. Molecular mechanism of the hydration of Candida antarctica lipase B in gas phase: Water adsorption isotherms and molecular dynamics simulations. ChemBioChem 2009, 10, 2913–2919. [Google Scholar] [CrossRef] [Green Version]
- Hock, K.J.; Grimmer, J.; Göbel, D.; Gasaya, G.G.T.; Roos, J.; Maucher, I.V.; Kühn, B.; Fettel, J.; Maier, T.J.; Manolikakes, G. Modular regiospecific synthesis of nitrated fatty acids. Synthesis 2016, 49, 615–636. [Google Scholar]
- Hassan, M.; Krieg, S.-C.; Nde, C.-N.; Roos, J.; Maier, T.J.; El Rady, E.A.; Raslan, M.A.; Sadek, K.U.; Manolikakes, G. Streamlined One-pot synthesis of nitro fatty acids. Eur. J. Org. Chem. 2021, 15, 2239–2252. [Google Scholar] [CrossRef]
- Woodcock, S.R.; Salvatore, S.R.; Freeman, B.A.; Schopfer, F.J. Synthesis of 9- and 12-nitro conjugated linoleic acid: Regiospecific isomers of naturally occurring conjugated nitrodienes. Tetrahedron Lett. 2021, 81, 153371. [Google Scholar] [CrossRef]
- Fang, M.Y.; Huang, K.-H.; Tu, W.-J.; Chen, Y.-T.; Pan, P.-Y.; Hsiao, W.-C.; Ke, Y.Y.; Tsou, L.K.; Zhang, M.M. Chemoproteomic profiling reveals cellular targets of nitro-fatty acids. Redox Biol. 2021, 46, 102126. [Google Scholar] [CrossRef]
Scheme 1.
Endogenous formation of nitrogen dioxide, the reactive nitrating agent of UFAs.

Scheme 2.
Endogenous nitration of UFAs [3].
Scheme 2.
Endogenous nitration of UFAs [3].
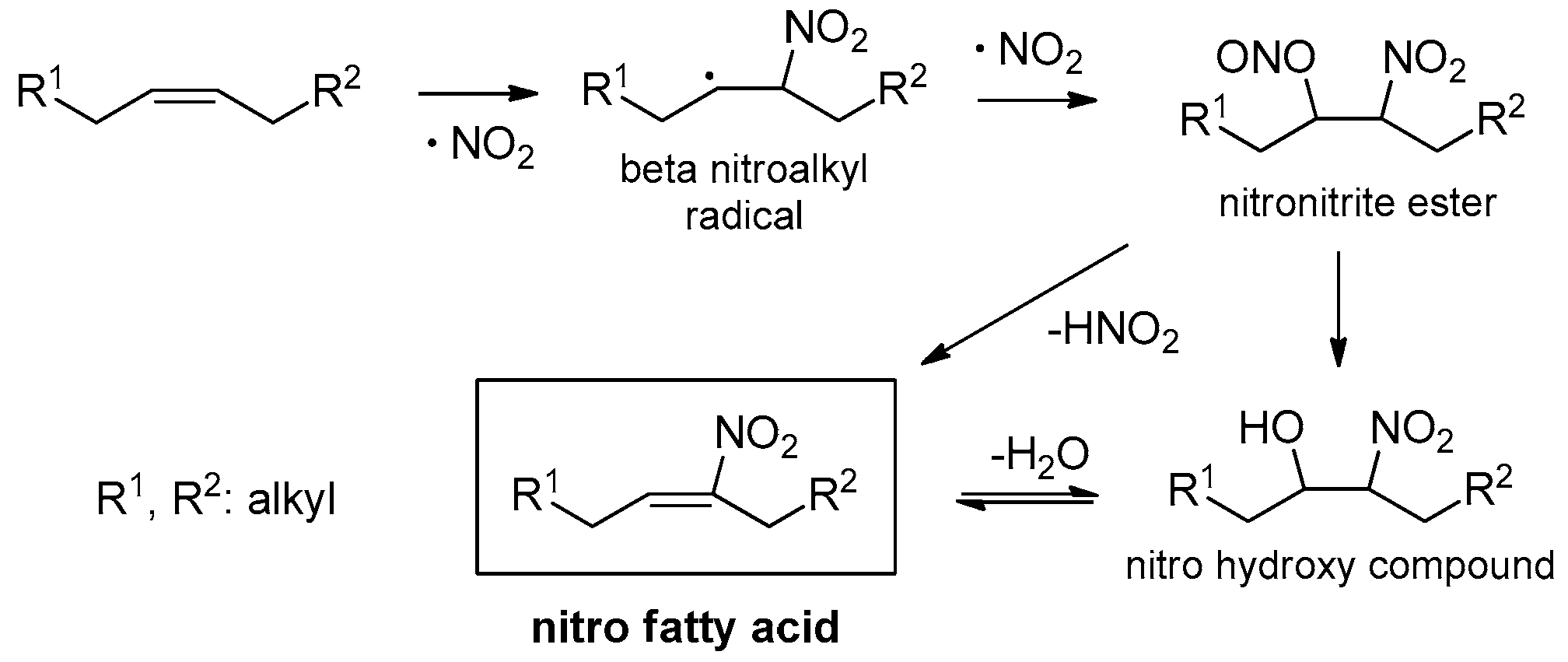
Figure 1.
Post-translational modification of protein of interest by Michael addition [3].
Figure 1.
Post-translational modification of protein of interest by Michael addition [3].
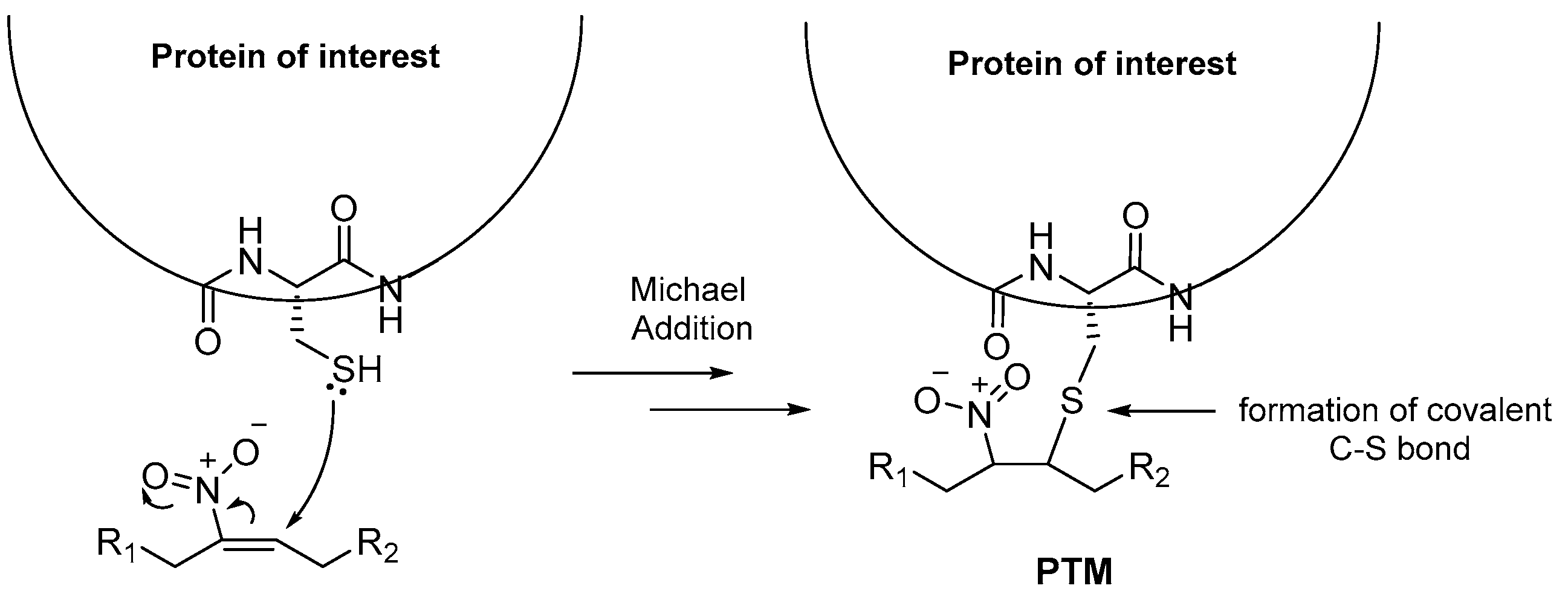
Figure 2.
CXA-10 at ongoing phase II clinical trials as an investigational drug against PAH, FSGS and asthma.
Figure 2.
CXA-10 at ongoing phase II clinical trials as an investigational drug against PAH, FSGS and asthma.
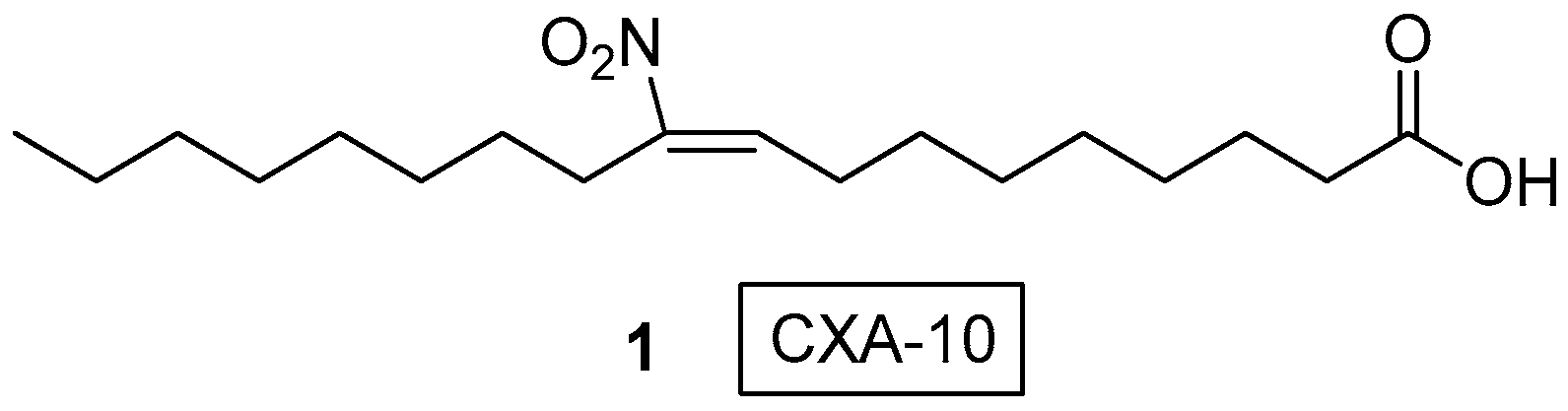
Figure 3.
Activation rate of ARE related to different NO2-FAs.

Figure 4.
Nitroalkenes showing similar deactivation rate of TNFα-induced NF-κB signaling pathway with NO2-OA.
Figure 4.
Nitroalkenes showing similar deactivation rate of TNFα-induced NF-κB signaling pathway with NO2-OA.
Figure 5.
9-NO2-OA as a cytotoxic agent against colorectal cancer cell lines.

Figure 6.
New NO2-FAs for SAR studies.
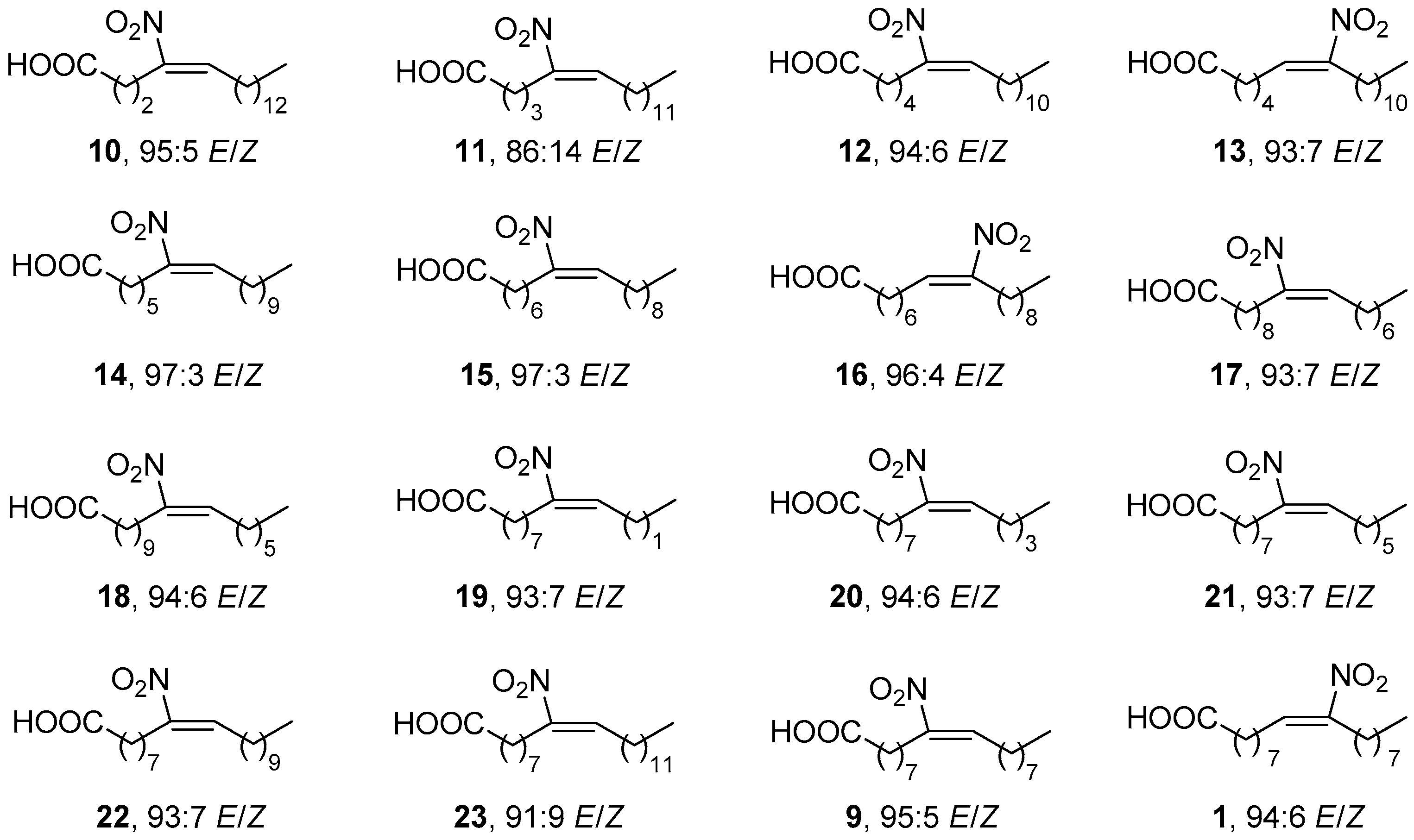
Figure 7.
Structure of the optimum activator of PPARγ reported by Gorczynski [74].
Figure 7.
Structure of the optimum activator of PPARγ reported by Gorczynski [74].
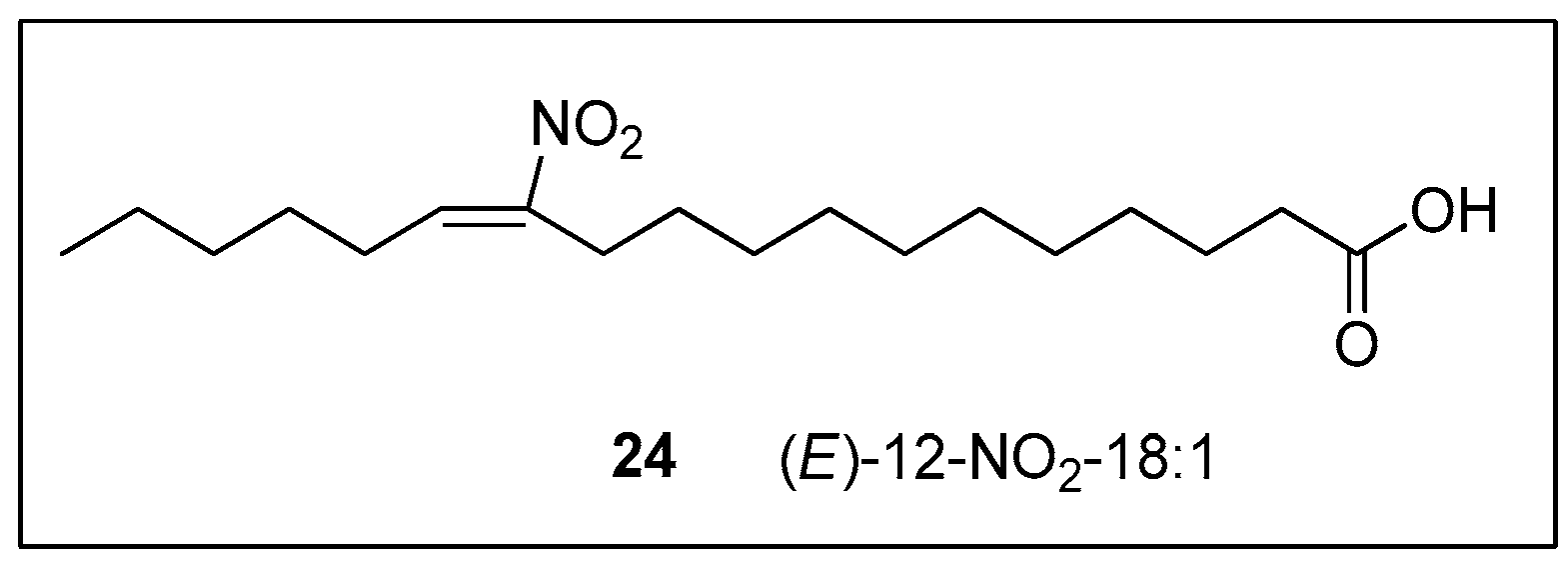
Figure 8.
Structures of the NO2-FAs reported by Denicola and coworkers, which act as activators of SIRT6 [97].
Figure 8.
Structures of the NO2-FAs reported by Denicola and coworkers, which act as activators of SIRT6 [97].

Scheme 3.
Different general strategies for the synthesis of NO2-FAs.
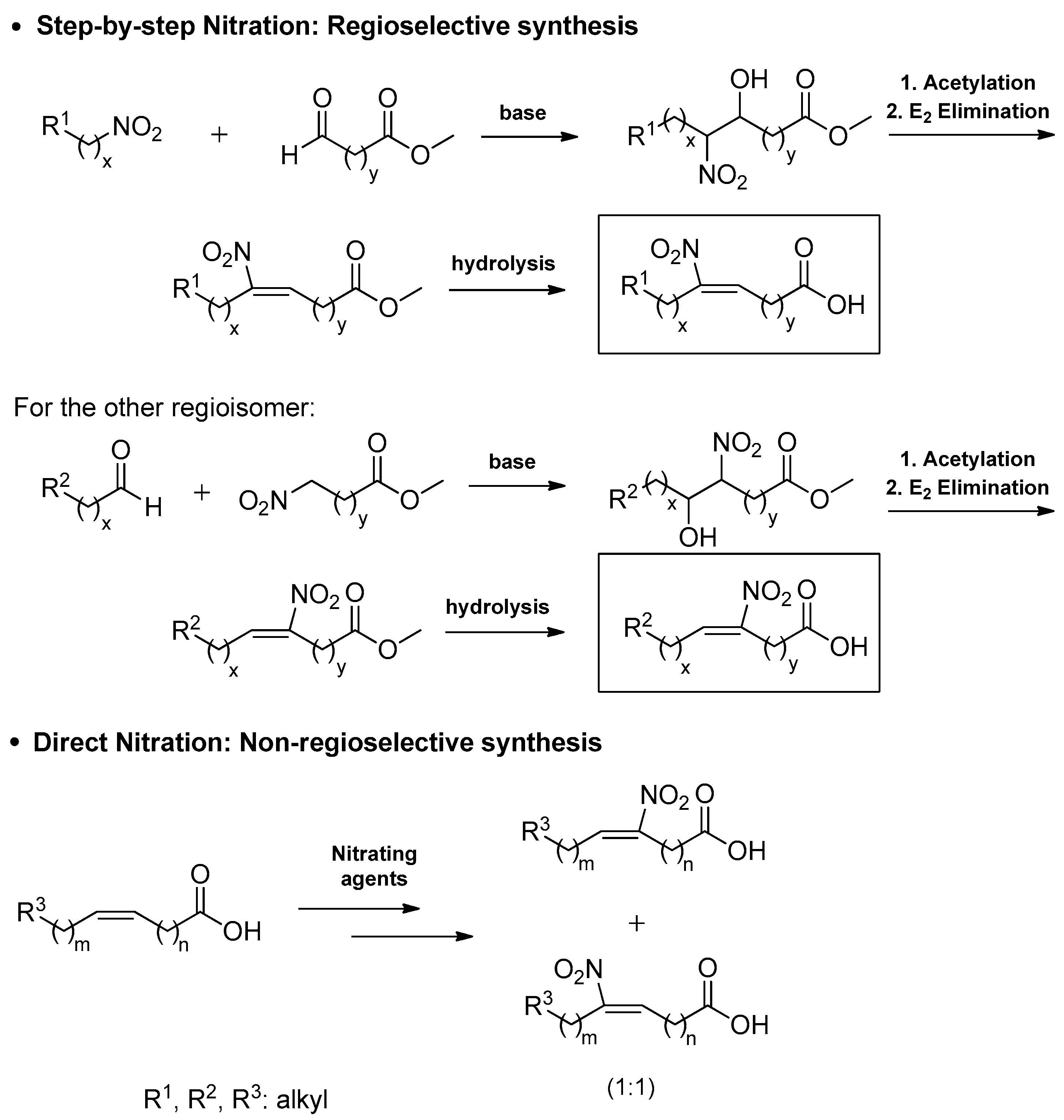
Scheme 4.
Synthesis of nitro derivatives of ethyl linoleate.

Figure 9.
Gas-induced synthesis of nitro isomers and some of their characteristic 1H-NMR shifts [101].
Figure 9.
Gas-induced synthesis of nitro isomers and some of their characteristic 1H-NMR shifts [101].
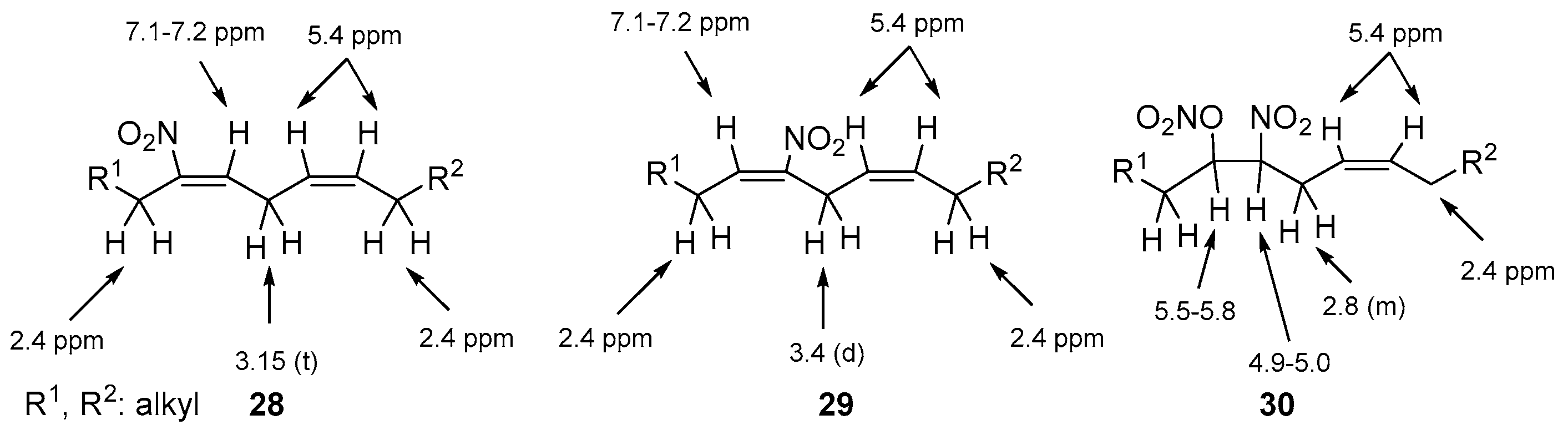
Scheme 5.
Ethyl linoleate and its nitro derivatives including nitroalkenes 29, 31–33.
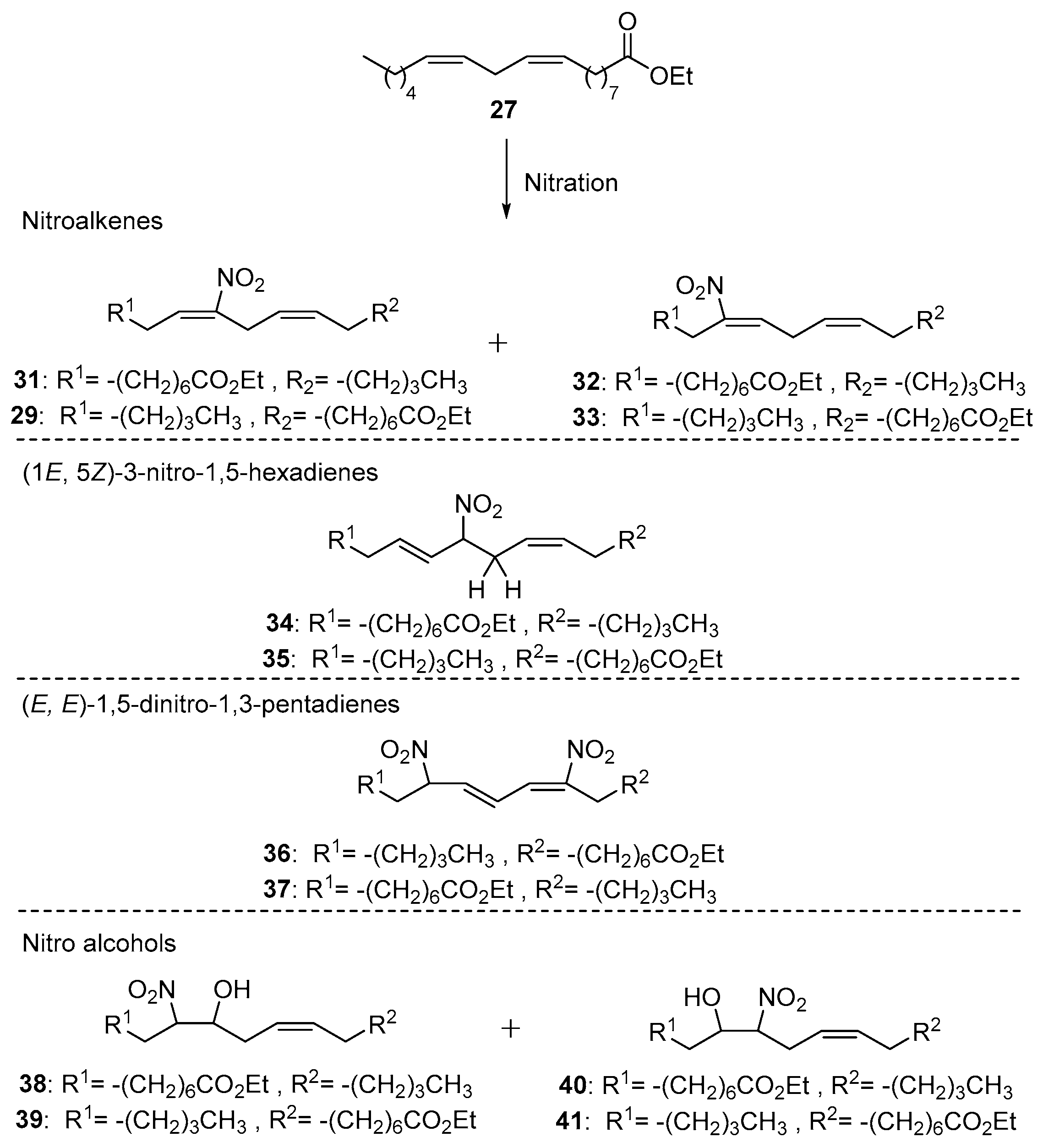
Scheme 6.
Nitroalkenes and by-products formed by nitration of methyl oleate.
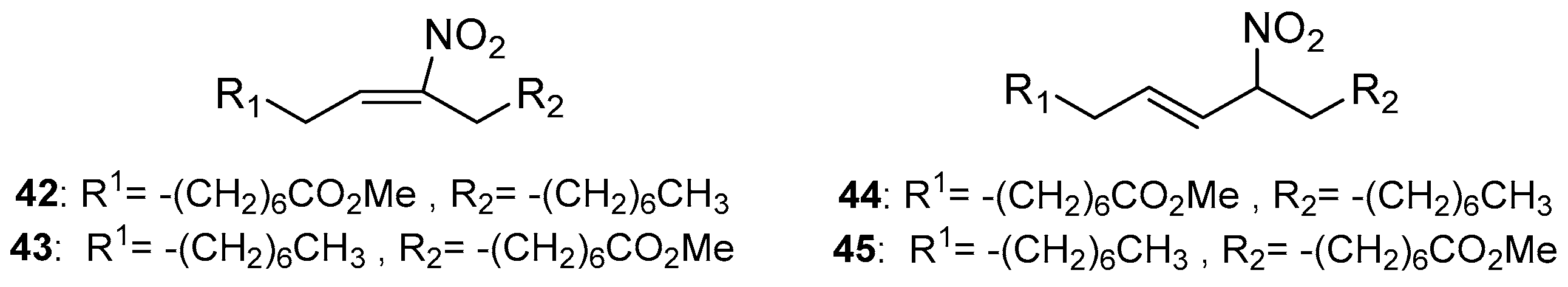
Scheme 7.
Synthesis of 9- and 10-NO2-LA using selenium-based method [104].
Scheme 7.
Synthesis of 9- and 10-NO2-LA using selenium-based method [104].
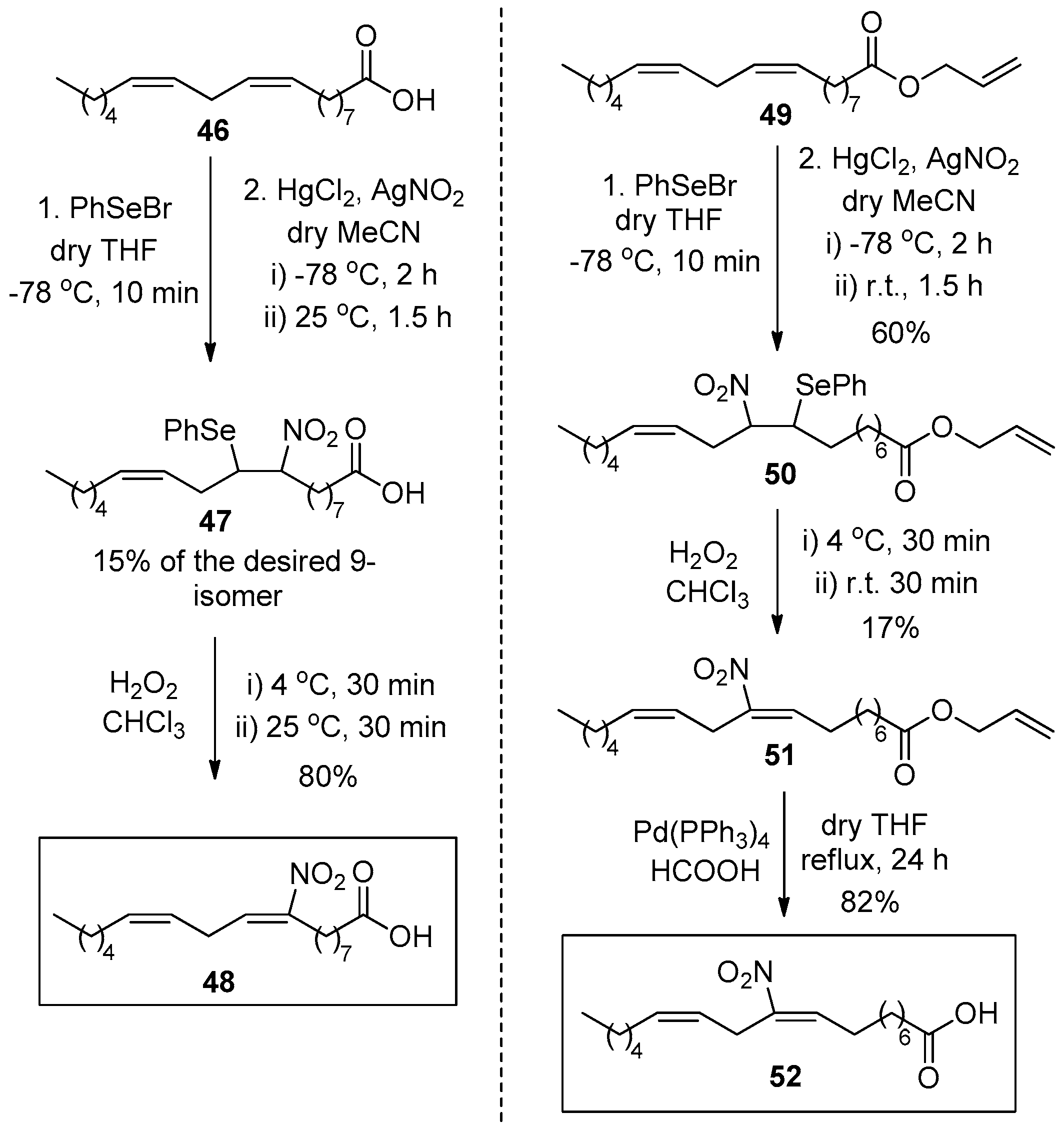
Scheme 8.
Degradation products after incubation of 9-NO2-LA at pH 7.4.

Scheme 9.
Unique decay product of 10-NO2-LA when exposed to pH 7.4.
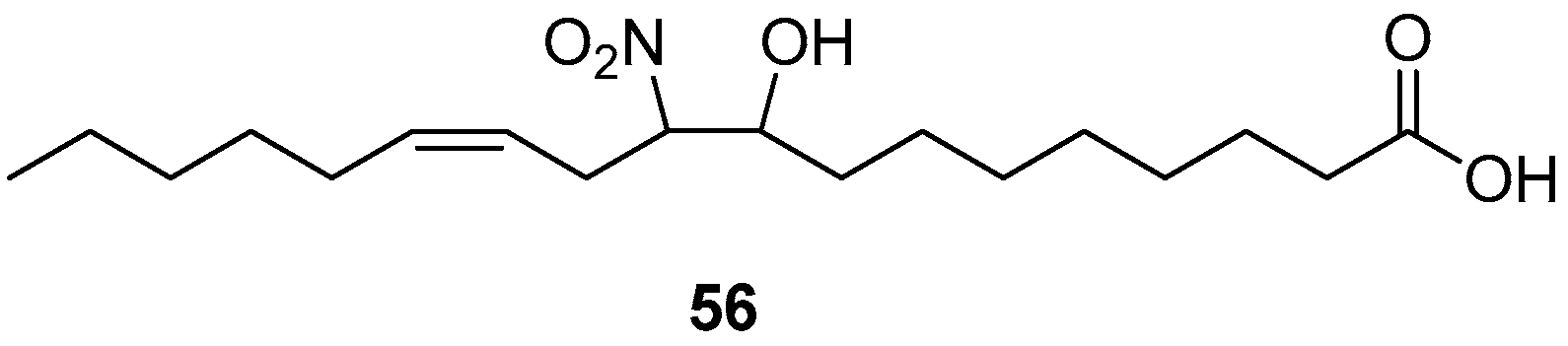
Scheme 10.
Synthesis of a mixture of 9- and 10-NO2-OA by the nitro-selenylation intermediate method [105].
Scheme 10.
Synthesis of a mixture of 9- and 10-NO2-OA by the nitro-selenylation intermediate method [105].
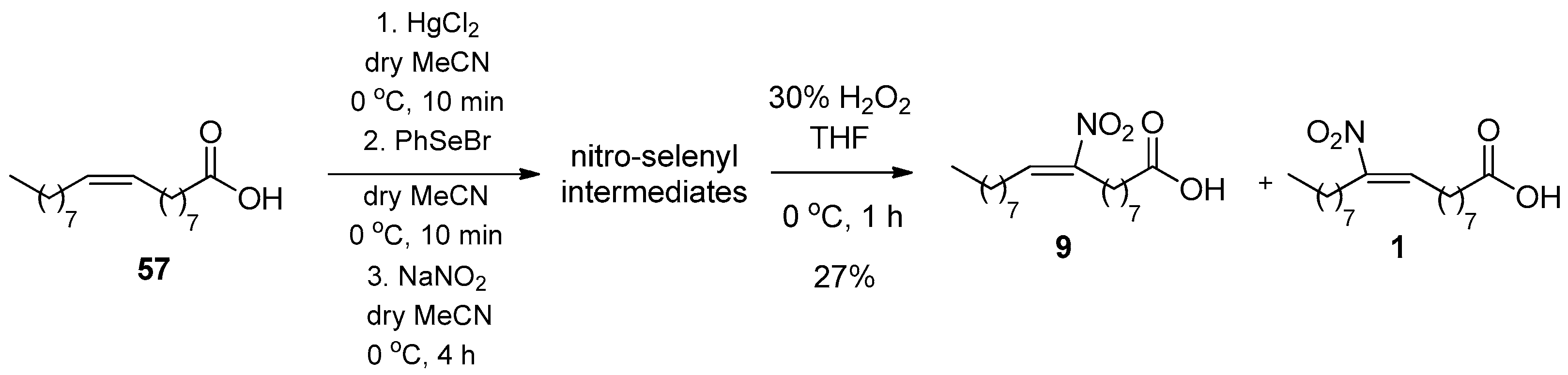
Scheme 11.
Proposed mechanism of the synthesis of 9- and 10-NO2-OA via nitro-selenylation, followed by oxidative elimination.
Scheme 11.
Proposed mechanism of the synthesis of 9- and 10-NO2-OA via nitro-selenylation, followed by oxidative elimination.
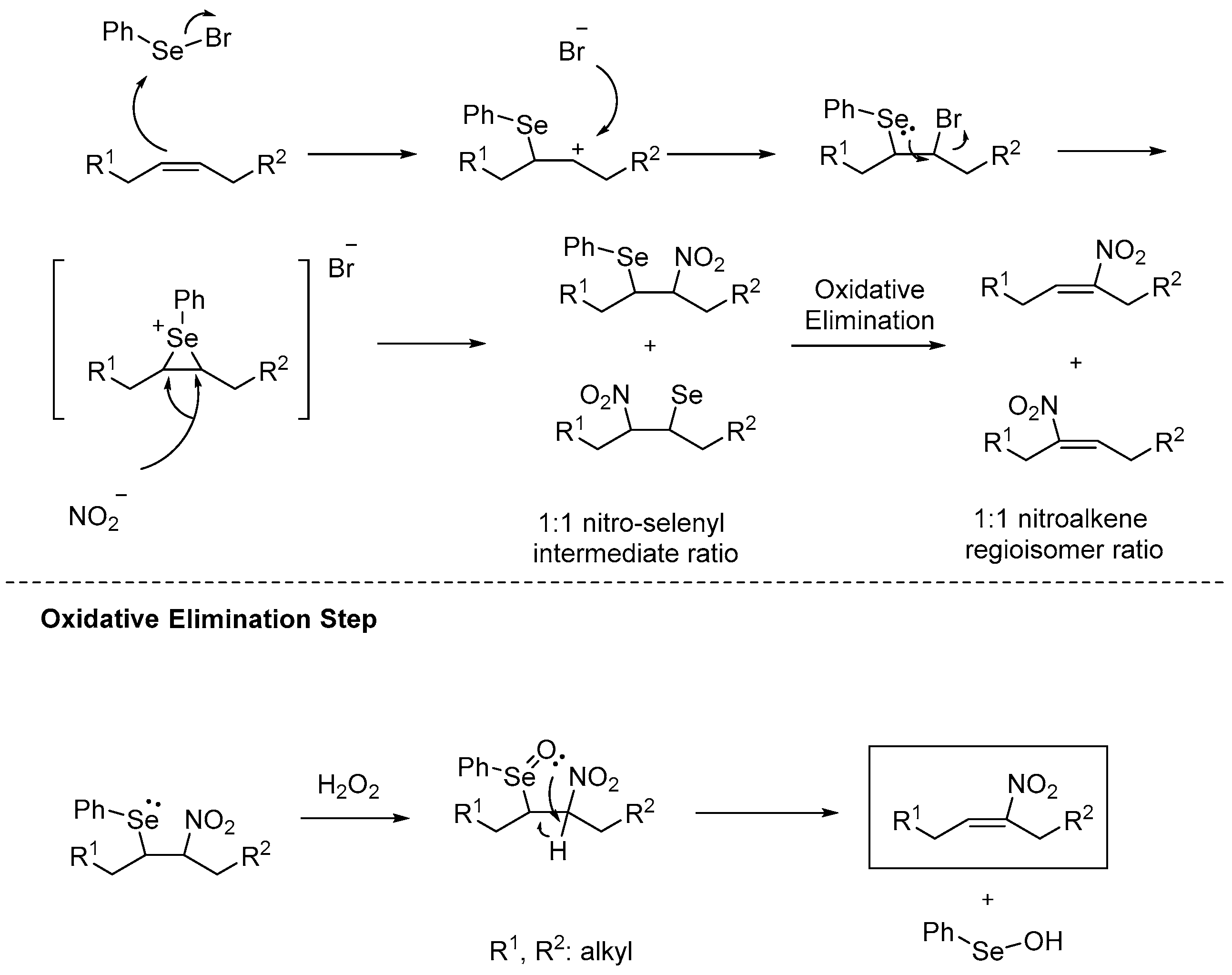
Scheme 12.
Retrosynthetic analysis for 9- and 10-NO2-OA.
Scheme 13.
Regiospecific synthesis of 9-NO2-OA by Gorczynski [109].
Scheme 13.
Regiospecific synthesis of 9-NO2-OA by Gorczynski [109].
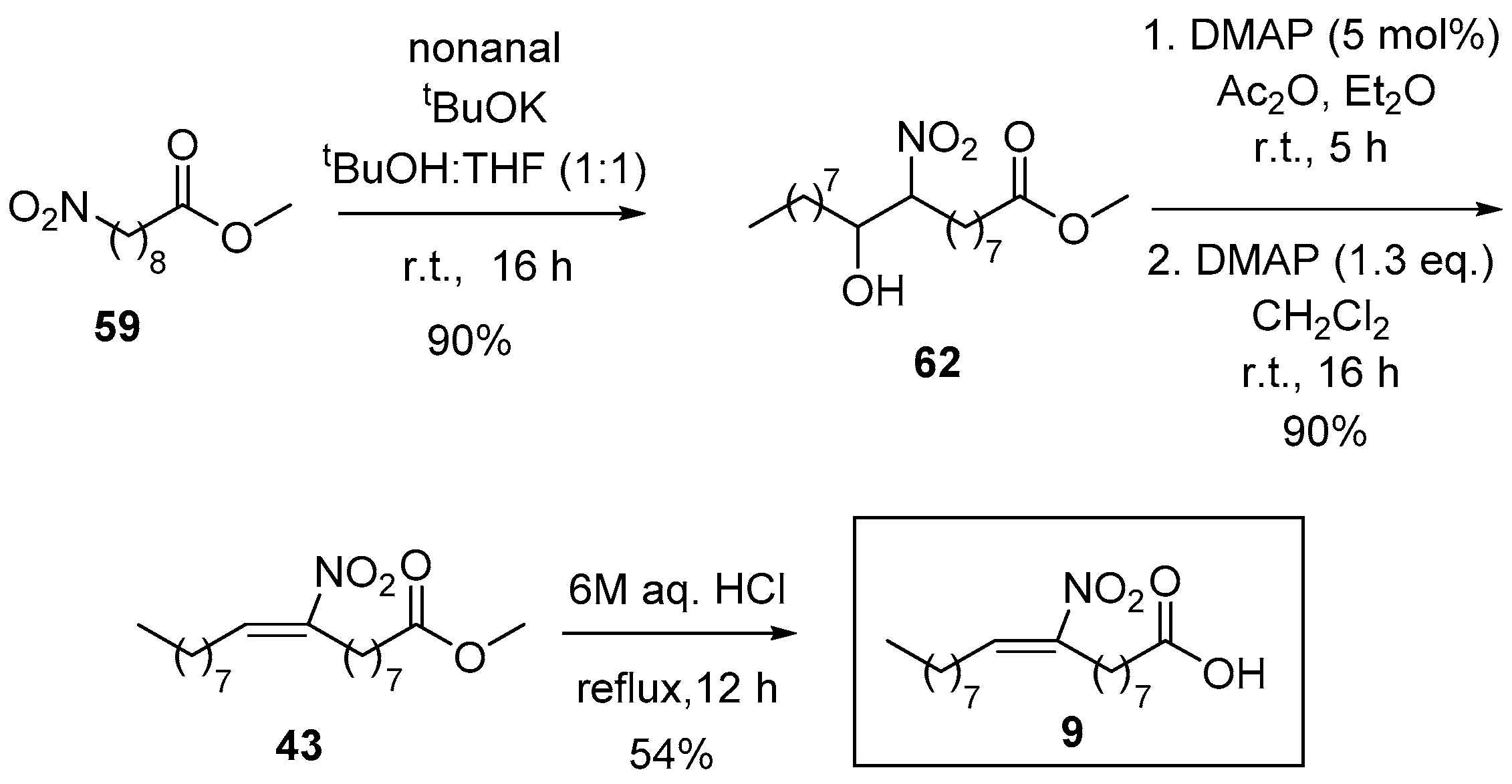
Scheme 14.
Synthetic route for 10-NO2-OA [109].
Scheme 14.
Synthetic route for 10-NO2-OA [109].
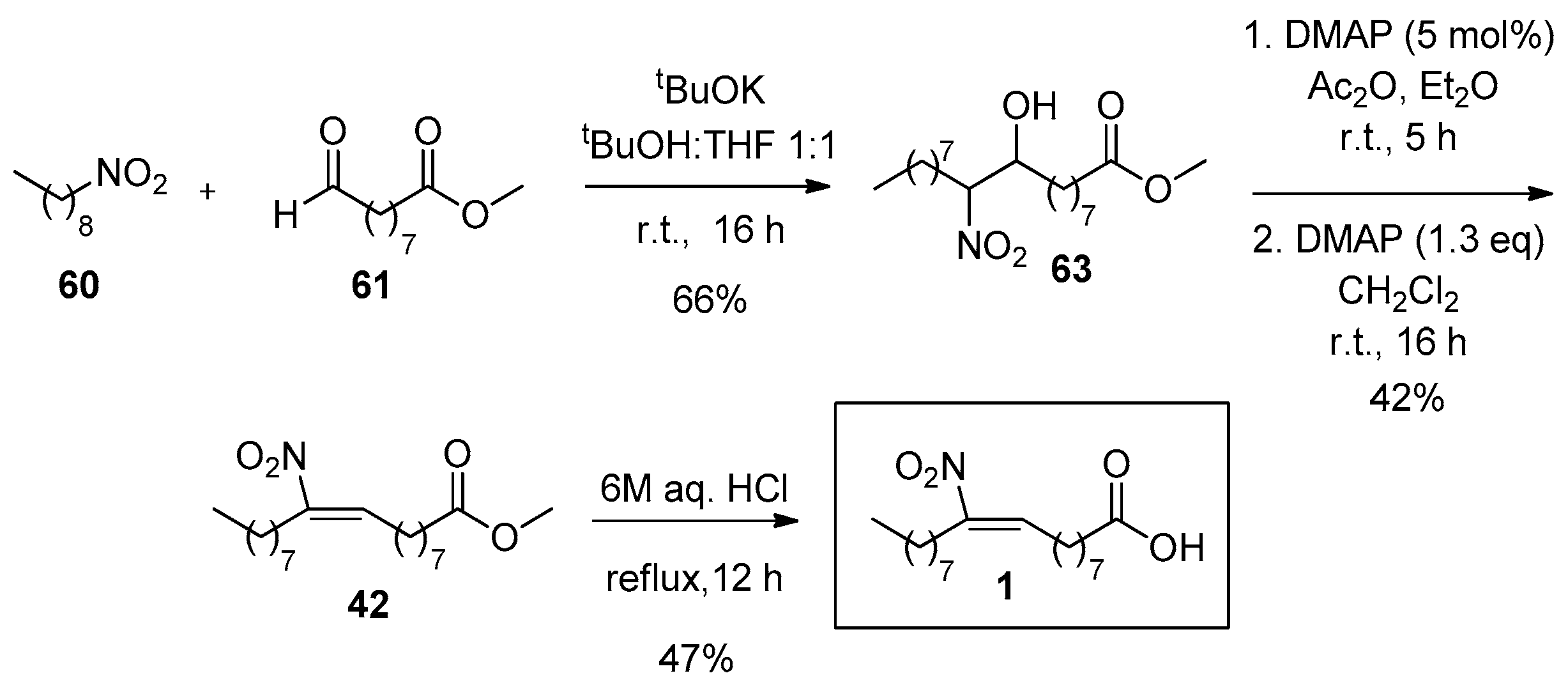
Scheme 15.
Synthesis of 9-NO2-OA by Branchaud’s protocol [110].
Scheme 15.
Synthesis of 9-NO2-OA by Branchaud’s protocol [110].

Scheme 16.
Synthesis of the precursor compounds for 10-NO2-OA [110].
Scheme 16.
Synthesis of the precursor compounds for 10-NO2-OA [110].
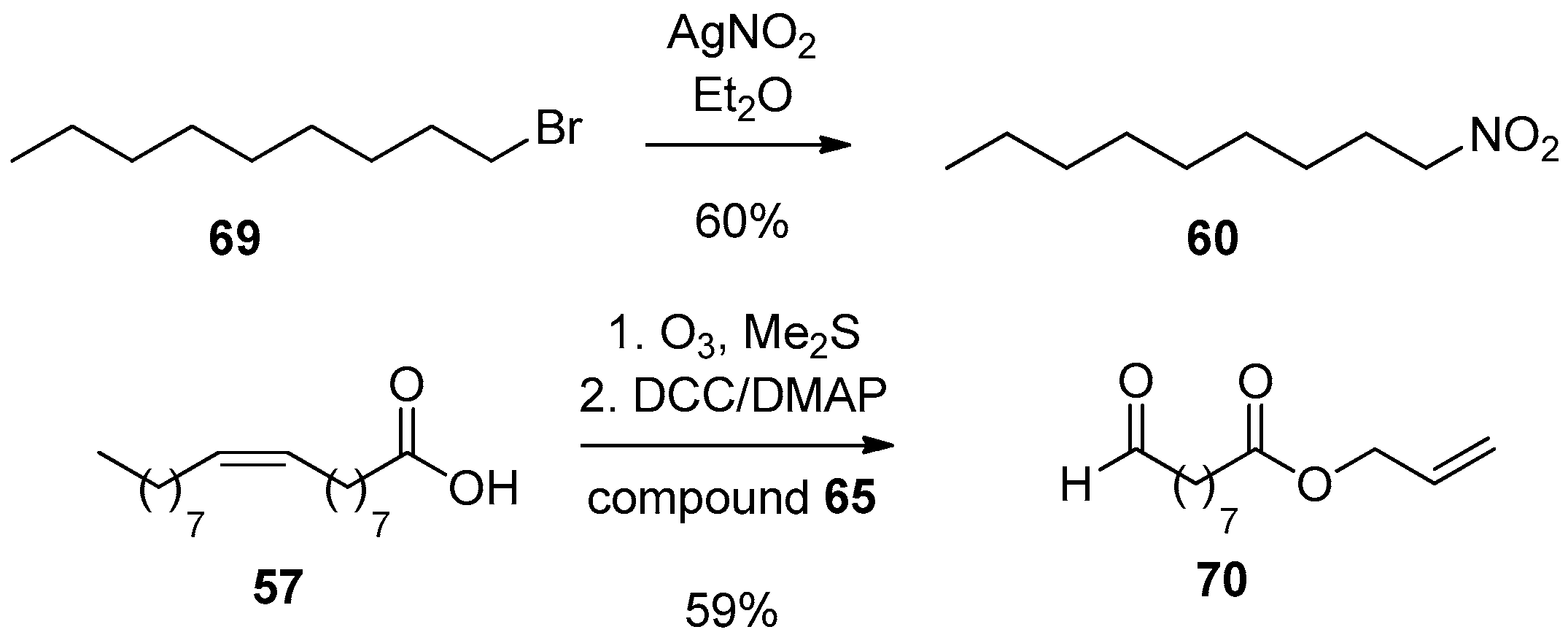
Scheme 17.
Synthesis of 10-NO2-OA by Branchaud’s protocol [110].
Scheme 17.
Synthesis of 10-NO2-OA by Branchaud’s protocol [110].
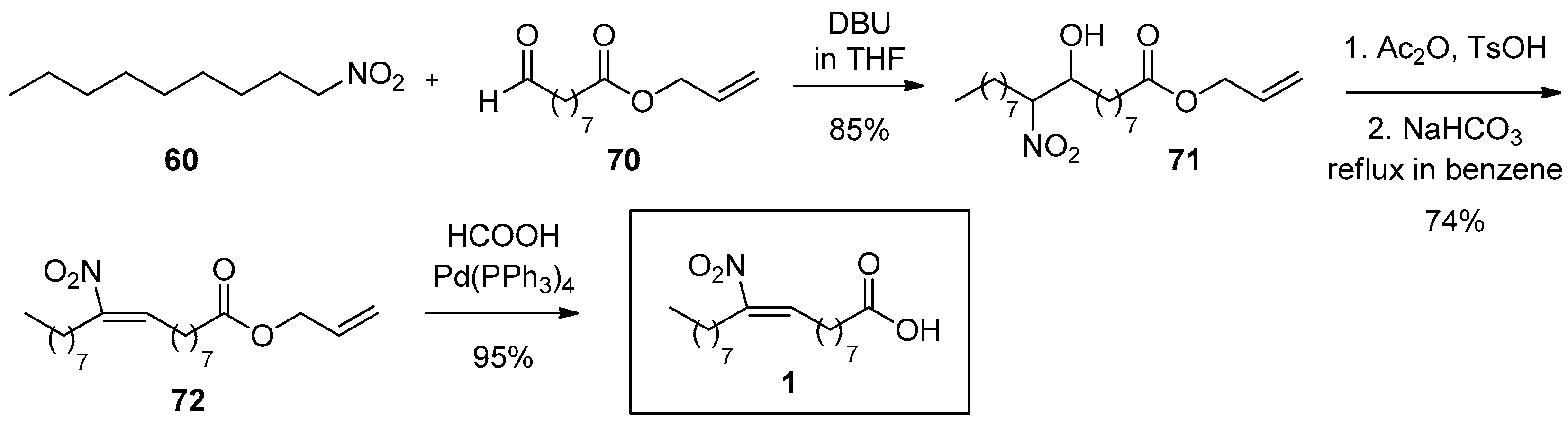
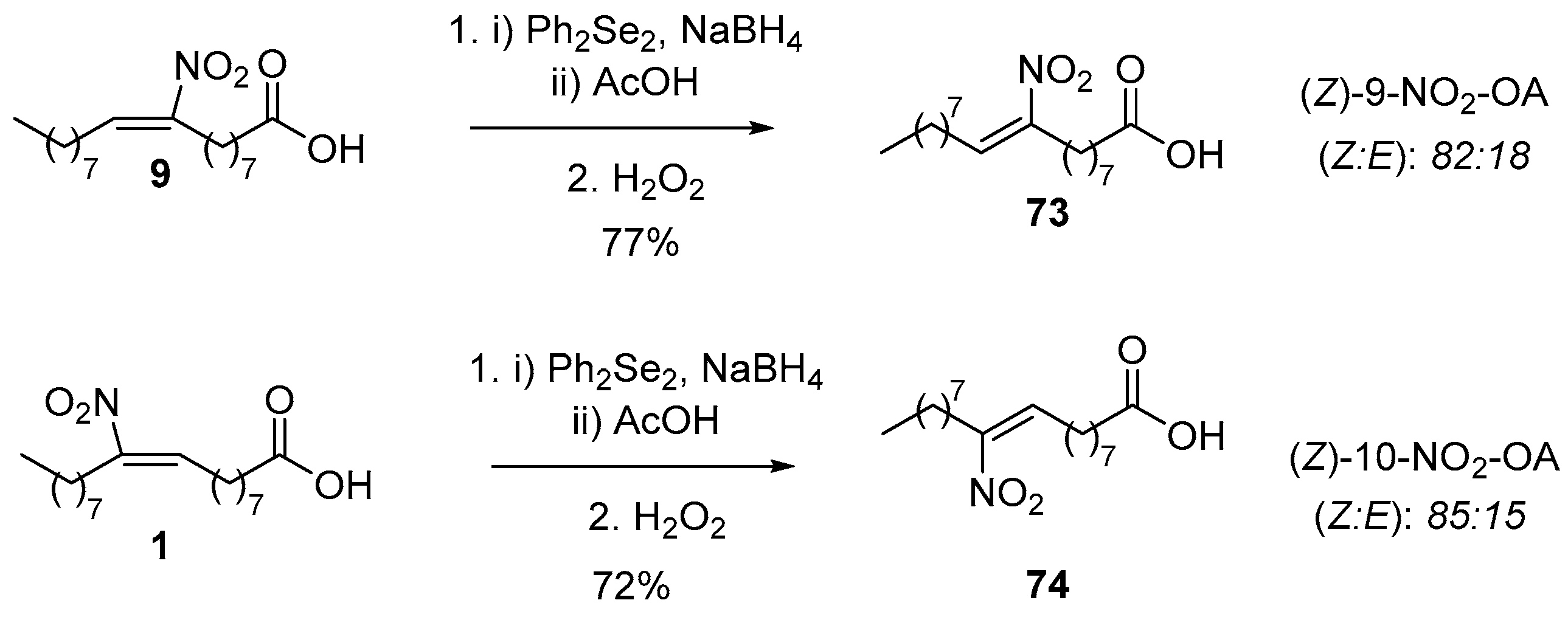
Scheme 19.
Zanoni’s methodology affording nitroalkene 24 in 50% overall yield [112].
Scheme 19.
Zanoni’s methodology affording nitroalkene 24 in 50% overall yield [112].
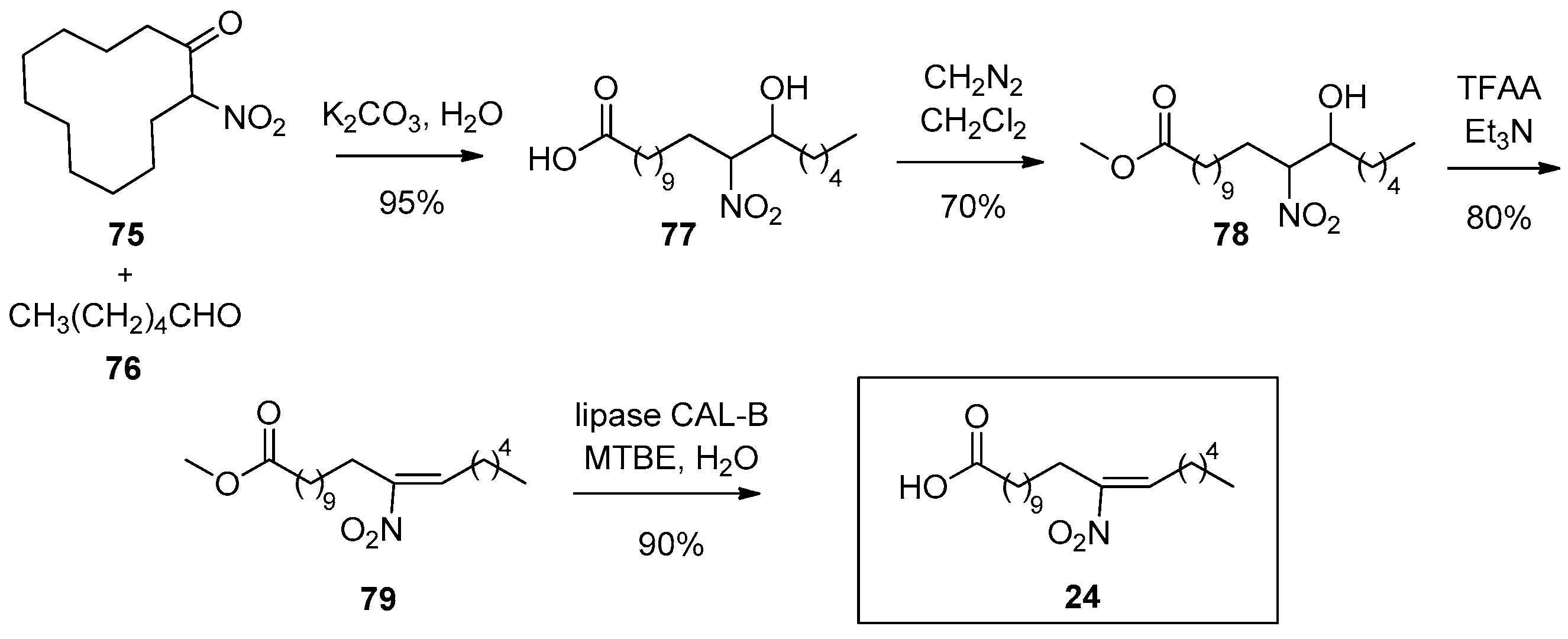
Scheme 20.
Manolikakes’ developed protocol for the synthesis of different NO2-FAs [116].
Scheme 20.
Manolikakes’ developed protocol for the synthesis of different NO2-FAs [116].

Scheme 21.
One-pot transformation for the selective formation of (E) NO2-FAs by Manolikakes [117].
Scheme 21.
One-pot transformation for the selective formation of (E) NO2-FAs by Manolikakes [117].
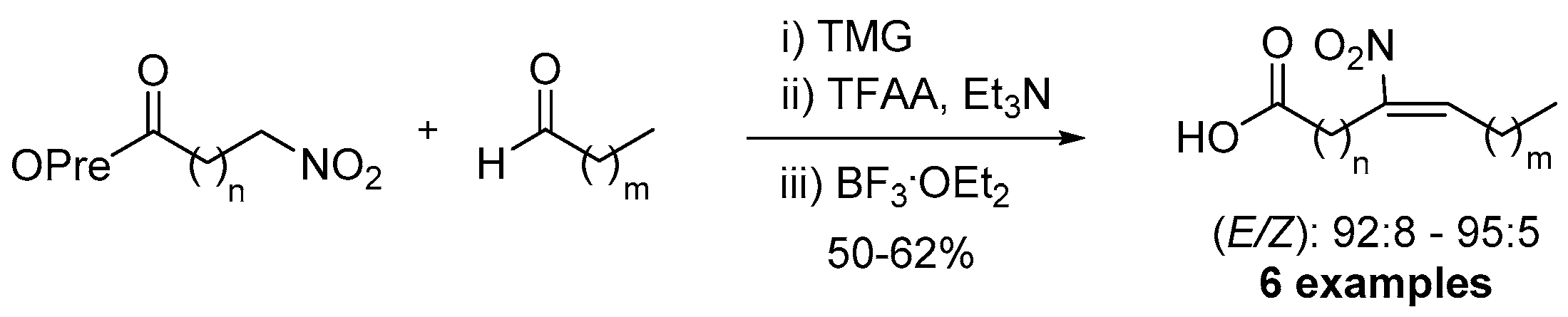
Scheme 22.
Synthesis of 9-NO2-cLA [118].
Scheme 22.
Synthesis of 9-NO2-cLA [118].

Scheme 23.
Synthesis of 12-NO2-cLA [118].
Scheme 23.
Synthesis of 12-NO2-cLA [118].

Scheme 24.
Synthesis of 9-nitrorumelenic acid [118].
Scheme 24.
Synthesis of 9-nitrorumelenic acid [118].
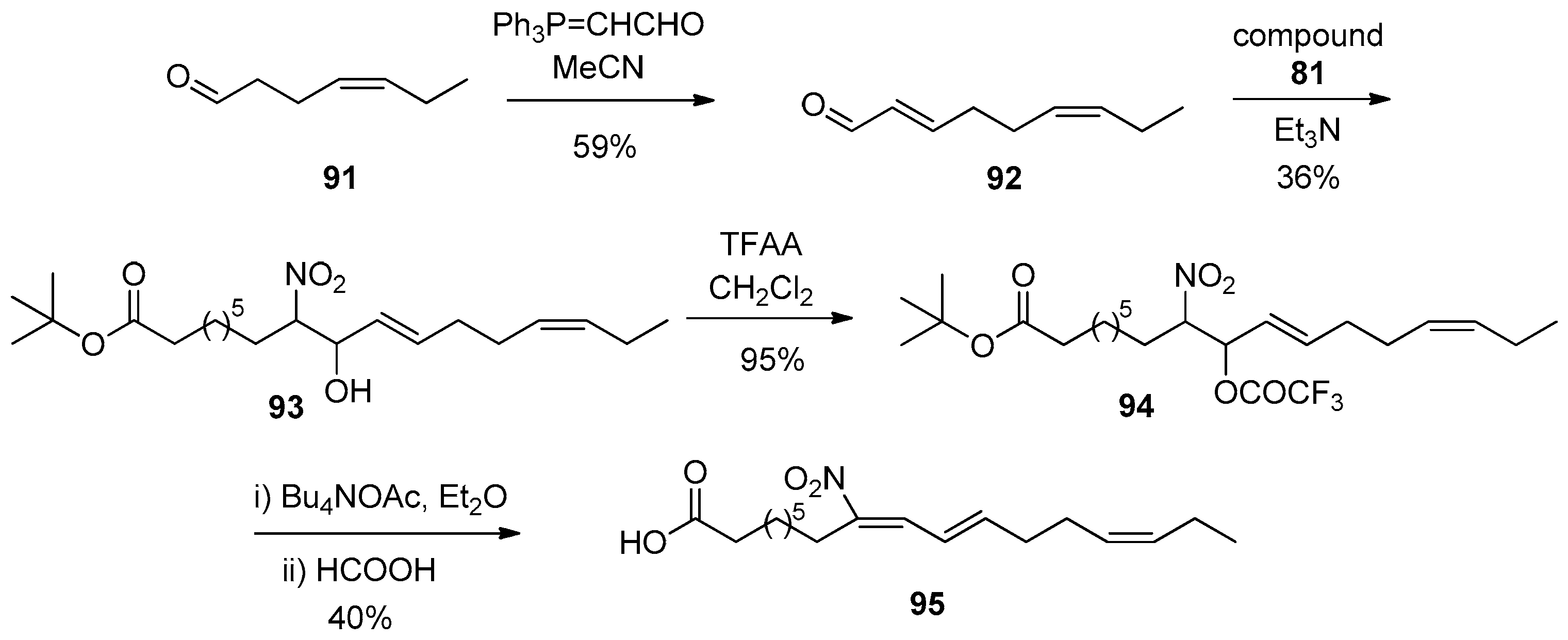
Scheme 25.
Synthesis of 9-nitroeicos-9-en-19-ynoic acid (99) [119].
Scheme 25.
Synthesis of 9-nitroeicos-9-en-19-ynoic acid (99) [119].
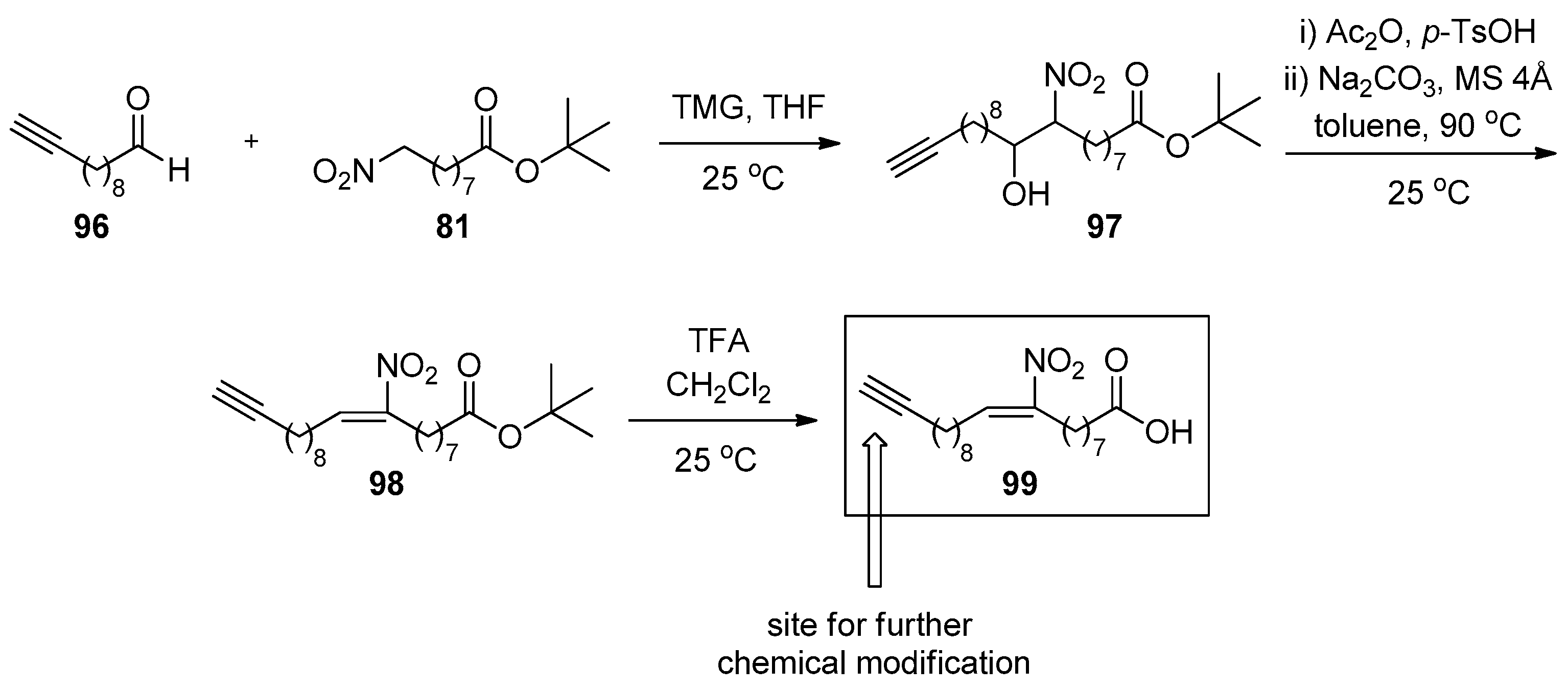

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
NO2-FA-PPAR SAR study. E-12-NO2-18:1 exhibits the highest activity as PPARγ agonist.
| Compound | IC50 (μM) | Ref. | |
|---|---|---|---|
| PPARγ | PPARα | ||
| Rosiglitazone | 0.25 | >15 | [75] |
| E-12-NO2-18:2 | 0.41 | 9.6 | [75] |
| E-9-NO2-18:1 | 0.98 | >15 | [74] |
| E-10-NO2-18:1 | 1.61 | >15 | [74] |
| Z-9/10-NO2-18:1 | 1.73 | 10.8 | [74] |
| E-9-NO2-16:1 | 0.83 | >15 | [74] |
| E-10-NO2-16:1 | 0.63 | >15 | [74] |
| E-5-NO2-18:1 | 1.68 | >15 | [74] |
| E-6-NO2-18:1 | 1.72 | >15 | [74] |
| E-12-NO2-18:1 | 0.039 | 1.6 | [74] |
| E-13-NO2-18:1 | 0.19 | 1.1 | [74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Koutoulogenis, G.S.; Kokotos, G. Nitro Fatty Acids (NO2-FAs): An Emerging Class of Bioactive Fatty Acids. Molecules 2021, 26, 7536. https://doi.org/10.3390/molecules26247536
AMA Style
Koutoulogenis GS, Kokotos G. Nitro Fatty Acids (NO2-FAs): An Emerging Class of Bioactive Fatty Acids. Molecules. 2021; 26(24):7536. https://doi.org/10.3390/molecules26247536
Chicago/Turabian StyleKoutoulogenis, Giorgos S., and George Kokotos. 2021. "Nitro Fatty Acids (NO2-FAs): An Emerging Class of Bioactive Fatty Acids" Molecules 26, no. 24: 7536. https://doi.org/10.3390/molecules26247536