
A Brief Overview of Potential Treatments for Viral Diseases Using Natural Plant Compounds: The Case of SARS-Cov

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
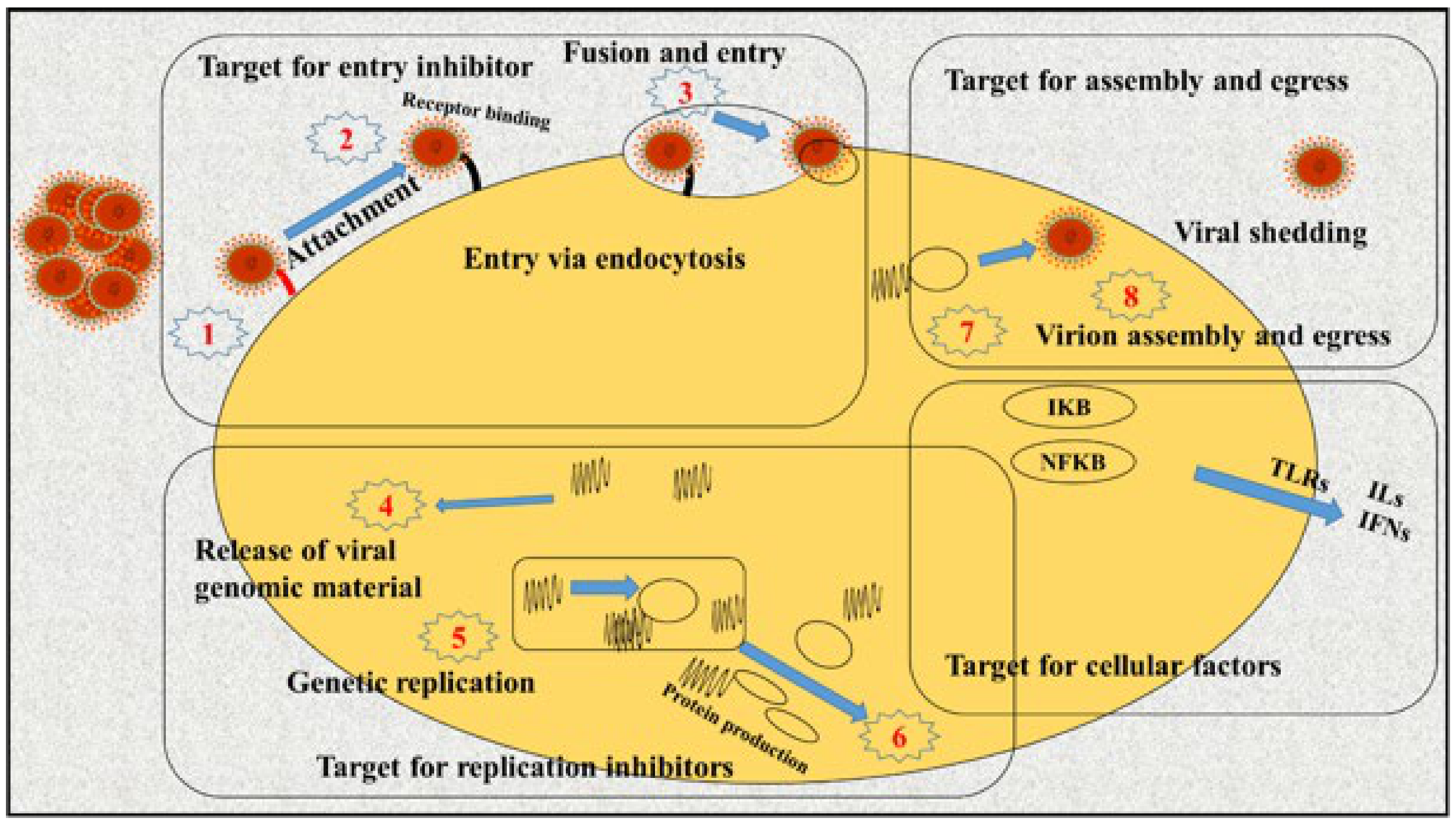
2. A Brief Model of Viral Entering/Replication in Host Cells
3. General Model of SARS-CoV Pathogenicity
4. Replication Inhibitors of SARS-CoV
5. Inhibitors of Assembly and Packaging of SARS-CoV
6. Evidence Supporting the Antiviral Efficacy of Medicinal Plants
7. Plant-Derived Immunomodulators

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Sources | Compounds | Molecular Formula | Lipinski’s Rule of Five | Reference | |
|---|---|---|---|---|---|---|
| Properties | Value | |||||
| Spinacia oleracea, Brassica oleracea, Anethum graveolens, Brassica rapa, Sauropus androgynus | Spinach Cabbage Dill Chinese cabbage Katuk | Kaempferol | C15H10O6 | Molecular weight (<500 Da) | 286.24 | [132,133] |
| LogP (<5) | 1.58 | |||||
| H-bond donor (5) | 4 | |||||
| H-bond acceptor (<10) | 6 | |||||
| Violations | 0 | |||||
| Anethum graveolens, Foeniculum vulgare, Allium cepa, Oregano vulgare, Capsicum annum | Dill Fennel leaves Onion Oregano Chili pepper | Quercetin | C15H10O7 | Molecular weight (<500 Da) | 302.24 | [132] |
| LogP (<5) | 1.23 | |||||
| H-bond donor (5) | 5 | |||||
| H-bond acceptor (<10) | 7 | |||||
| Violations | 0 | |||||
| Olea europaea, Averrhoa belimbi, Capsicum annum, Allium fistulosum | Olive Star fruit Chili pepper Welsh onion/Leek | Luteolin-7-glucoside | C21H20O11 | Molecular weight (<500 Da) | 448.38 | [134] |
| LogP (<5) | 0.16 | |||||
| H-bond donor (5) | 7 | |||||
| H-bond acceptor (<10) | 11 | |||||
| Violations | 2 | |||||
| Curcuma longa, Curcuma xanthorriza | Turmeric Curcuma | Demethoxycurcumine | C20H18O5 | Molecular weight (<500 Da) | 338.35 | [135,136] |
| LogP (<5) | 3 | |||||
| H-bond donor (5) | 2 | |||||
| H-bond acceptor (<10) | 5 | |||||
| Violations | 0 | |||||
| Citrus sinensis | Citrus fruit | Naringenin | C15H12O5 | Molecular weight (<500 Da) | 567.78 | [137] |
| LogP (<5) | 4.33 | |||||
| H-bond donor (5) | 4 | |||||
| H-bond acceptor (<10) | 5 | |||||
| Violations | 1 | |||||
| Averrhoa belimbi, Lycium chinese, Apium graveolens, Olea Europaea | Star fruit Goji berries Celery Olive | Apigenine-7-glucoside | C21H20O10 | Molecular weight (<500 Da) | 432.34 | [138,139,140] |
| LogP (<5) | 0.55 | |||||
| H-bond donor (5) | 6 | |||||
| H-bond acceptor (<10) | 10 | |||||
| Violations | 1 | |||||
| Olea Europaea | Olive | Oleuropein | C19H22O8 | Molecular weight (<500 Da) | 378.37 | [138] |
| LogP (<5) | 1.57 | |||||
| H-bond donor (5) | 3 | |||||
| H-bond acceptor (<10) | 8 | |||||
| Violations | 0 | |||||
| Camellia sinesis | Green tea | Catechin | C15H14O6 | Molecular weight (<500 Da) | 290.27 | [141,142] |
| LogP (<5) | 0.85 | |||||
| H-bond donor (5) | 5 | |||||
| H-bond acceptor (<10) | 6 | |||||
| Violations | 0 | |||||
| Curcuma xanthorriza, Curcuma longa | Turmeric Curcuma | Curcumin | C21H20O6 | Molecular weight (<500 Da) | 368.38 | [135,136] |
| LogP (<5) | 3.03 | |||||
| H-bond donor (5) | 2 | |||||
| H-bond acceptor (<10) | 6 | |||||
| Violations | 0 | |||||
| Zingiber officiale | Ginger | Zingerol | C11H16O3 | Molecular weight (<500 Da) | 196.24 | [36,143,144] |
| LogP (<5) | 1.86 | |||||
| H-bond donor (5) | 2 | |||||
| H-bond acceptor (<10) | 3 | |||||
| Violations | 0 | |||||
| Zingiber officiale | Ginger | Gingerol | C17H26O4 | Molecular weight (<500 Da) | 294.39 | [36,144,145] |
| LogP (<5) | 3.13 | |||||
| H-bond donor (5) | 2 | |||||
| H-bond acceptor (<10) | 4 | |||||
| Violations | 0 | |||||
| Allium sativum | Garlic | Allicin | C6H10OS2 | Molecular weight (<500 Da) | 162.27 | [36] |
| LogP (<5) | 1.61 | |||||
| H-bond donor (5) | 0 | |||||
| H-bond acceptor (<10) | 1 | |||||
| Violations | 0 | |||||
| Camellia sinesis | Green tea | Epicatechin gallate | C22H18O10 | Molecular weight (<500 Da) | 442.37 | [139] |
| LogP (<5) | 1.23 | |||||
| H-bond donor (5) | 7 | |||||
| H-bond acceptor (<10) | 10 | |||||
| Violations | 1 | |||||
| Plant Species and Plant Part | Active Compounds | Coumarins | Extract | Model Organism | Mode of Action/Activity | Ref | |||
|---|---|---|---|---|---|---|---|---|---|
| Terpenes Terpenoids | Flavonoids Flavones | Alkaloids | Stilbenes | ||||||
| Méntha piperíta (whole plant) Lamiaceae | α-pinene β-pinene β-caryophyllene L-Limonene Menthol | Eriocitrin Hesperidin Kaempferol 7-O-rutinoside Luteolin and its derivatives | n/a | Trans-resveratrol | n/a | Ethanol | Vero cell cultures | High antiviral activity | [145,146,147] |
| Thymus vulgaris (whole plant) Lamiaceae | Thymol p-cymene g-erpinene γ-Terpinene Linalool | Rutin Quercetin | n/a | n/a | n/a | Ethanol | Vero cell cultures | High antiviral activity and antioxidant effects | [145,148,149] |
| Desmodium canadense (whole plant) Fabaceae | Sandosaponin B and its derivativesSoyasaponin I Soyasaponin VI | Homoorientin Orientin 2-vicenin Vitexin Isovitexin Rutin Desmodin Homoadonivernite | Indole-3-alkylamine phenylethylamine alkaloids, pyrrolidine alkaloids | n/a | n/a | Ethanol | Vero cell cultures | High antiviral activity | [145,150,151,152,153] |
| Camellia japonica (whole plant, flowers) Theaceae | Oleanane triterpenes 3β,18β-dihydroxy-28-norolean-12-en-16-one 18β-hydroxy-28-norolean-12-ene-3,16-dione | Quercetin Kaempferol Apigenin | Do not produce purine alkaloids | n/a | n/a | Ethanol | Vero cells (African green monkey kidney cell line; ATCC CCR-81) | High antiviral activity on PEDV corona virus Inhibitory effects on key gene and protein synthesis during PEDV replication | [154,155,156,157,158,159] |
| Saposhnikovia divaricate (whole plant) Apiaceae | n/a | n/a | n/a | n/a | cis-3′-Isovaleryl4′-acetylkhellactone Praeruptorin F Praeruptorin B (−)-cis-khellactone | Ethanol | Vero cells (African green monkey kidney cell line; ATCC CCR-81) | High antiviral activity on PEDV corona virus | [155] |
| Quercus ilex L. (Leaves) Fagaceae | kaempferol glycosides (juglanin, kaempferol-3-O-α-L-arabinofuranoside, and afzelin, kaempferol-3-O-α-L-rhamnoside | n/a | n/a | n/a | DMSO | Xenopus oocytes | Inhibits 3a channel protein of coronavirus | [156,157] | |
| Bupleurum sp. (whole plant) Apiaceae | Triterpenoid saponins Saikosaponins 2″-O-Acetylsaikosaponins Prosaikogenins Bupleurosides Etc. | Quercetin Isorhamnetin Narcissin Rutin Eugenin Saikochrome A | n/a | n/a | n/a | DMSO | Human fetal lung fibroblasts (MRC-5; ATCC CCL-171) | Saikosaponins attenuate viral attachment and penetration | [160,161] |
| Houttuynia cordata (whole plant) (Saururaceae) | Cycloart-25-ene-3b,24-diol | Quercetin 7-rhamnoside Hyperin Quercetin Afzelin Rutin | Arisolactams Piperolactam A Caldensin | n/a | n/a | Water | BALB/c mice | Decreases the viral SARS-3CLpro activity Stops viral t RNA polymerase activity (RdRp) Increases the secretion of interleukin (IL)-2 and (IL)-10 | [88,162] |
| Isatis tinctoria (Roots extracts) Brasicaceae | n/a | Hesperetin Quercetin Isoorientin Isovitexin | Indigo Indirubin Indican | Sinigrin | n/a | Water | Vero cells | Cleavage of the activity of SARS-3CLpro enzyme decreased | [163,164] |
| Lycoris radiata (Bulbs) Amaryllidaceae | β-Myrcene A-terpineol Eucalyptol β-cyclocitral | n/a | Lycorine Amaryllidaceae alkaloids Lycoranines | n/a | Ethanol | Vero E6 cells | Exhibits anti-SARS-CoV activity | [16,165,166,167] | |
| Litchi chinensis (seeds) Sapindaceae | 3-Oxotrirucalla-7,24-dien-21-oic acid | Herbacetin Rhoifolin Pectolinarin Quercetin Epigallocatechin gallate Gallocatechin gallate Litchitannins Kaemferol derivatives Epicatechin Cinnamtannin | n/a | n/a | n/a | Water | On model with SARS-CoV 3CLpro | Inhibits SARS-3CLpro activity | [168,169,170,171] |
| Stephania tetrandra S Moore (Roots) Menispermaceae | n/a | n/a | Tetrandrine Fangchinoline, Cepharanthine | n/a | n/a | DMSO | Human cell line MRC-5 cells | Inhibits the expression of HCoV-OC43 spike and nucleocapsid protein. Immunomodulation/ | [129,172] |
| Scutellaria baicalensis (Roots) Lamiaceae | Dodecanedioxins | Scutellarein Baicalin Wogonin Wogonoside | n/a | n/a | n/a | DMSO | Model with SARS-CoV helicase, and nsP13 | Inhibits nsP13 by affecting the ATPase activity | [61,173] |
| Allium sativum (Bulbs) Alliaceae | Nerolidol Phytol Squalene α-pinene Terpinolene Limonene 1,8-cineole γ-terpinene | Catechin Epicatechin | Allicin Ajoene Alliin Diallyl disulfide Diallyl trisulfide | n/a | n/a | Aquaporin | Chicken embryos | Inhibitory effects on avian coronavirus | [174,175,176,177] |
| Artemisia sp. Artemisia absentium (whole plants) Asteraceae | Absinthin Artemisin Scopoletin Artamarin | Rutin Glycosides of quercetin | Artamarin Artamaridin, Artamaridinin, Artamarinin QuebrachitolArtemitin | n/a | n/a | Water | Delayed brain tumor cells | Reduces coronavirus replication | [178,179] |
| Juniperus communis (Fruits) Cupressaceae | Sugiol α-pinene β-pinene | Rutin Scutellarein Quercetin-3-O-rhamnoside quercitrin | n/a | n/a | Umbelliferone | n/a | Protein-molecular docking with network pharmacology analysis | Inhibits the replication, 3CLpro | [180,181] |
| Ecklonia cava (whole plant) Lessoniaceae | n/a | Quercetin | n/a | n/a | n/a | n/a | protein-molecular docking with network pharmacology analysis | PLpro and 3CLpro | [182] |
7.1. Lectins
7.2. Quercetin
7.3. Sulforaphane
7.4. Resveratrol
7.5. Baicalin
7.6. Glycyrrhizin
7.7. Narcissoside
7.8. Curcumin
7.9. Epigallocatechin Gallate
8. Laboratory Evidence Supporting Application of Medicinal Plants Against Respiratory Disorders
8.1. Artemisia annua
8.2. Allium cepa
8.3. Andrographis paniculata
8.4. Aloe vera
8.5. Nigella sativa
8.6. Salvia officinalis
8.7. Toona sinensis
8.8. Eckolina cava
8.9. Isatis indigotica
8.10. Azadirachta indica
8.11. Other Medicinal Herbs
9. In Silico Analysis of Medicinal Plants Role Against SARS-CoV
10. Biotechnological Production of Vaccines
10.1. Molecular Pharming: A Mature Technology to Produce Plant-Derived Pharmaceutical Products
10.2. Hairy Root Culture: A Reliable Method of Producing Pharmaceutical Products
11. Conclusions
12. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amber, R.; Adnan, M.; Tariq, A.; Mussarat, S. A review on antiviral activity of the Himalayan medicinal plants traditionally used to treat bronchitis and related symptoms. J. Pharm. Pharmacol. 2017, 69, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Peeri, N.C.; Shrestha, N.; Rahman, S.; Zaki, R.; Tan, Z.; Bibi, S.; Baghbanzadeh, M.; Aghamohammadi, N.; Zhang, W.; Haque, U. The SARS, MERS and novel coronavirus (COVID-19) epidemics, the newest and biggest global health threats: What lessons have we learned? Int. J. Epidemiol. 2020, 49, 717–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liu, W. Puzzle of highly pathogenic human coronaviruses (2019-nCoV). Protein Cell 2020, 11, 235–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathya, B.; Velpandian, V.; Ramani, M.; Kumar, M.P. A primitive approach on review of Siddha herbs, herbo-mineral formulation exhibiting antiviral activity. Int. J. Pharm. Bio. Sci. 2014, 5, 138–147. [Google Scholar]
- Forman, R.; Shah, S.; Jeurissen, P.; Jit, M.; Mossialos, E. COVID-19 vaccine challenges: What have we learned so far and what remains to be done? Health Policy 2021, 125, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, M.; Saleem, S.; Babar, Z.; Ali, A.; Khan, A.A.; Sardar, Z.; Hamayun, F.; Ali, S.S.; Wei, D.-Q. Phylogenetic Analysis and Structural Perspectives of RNA-Dependent RNA-Polymerase Inhibition from SARs-CoV-2 with Natural Products. Interdiscip. Sci. Comput. Life Sci. 2020, 12, 335–348. [Google Scholar] [CrossRef]
- Khan, A.; Tahir Khan, M.; Saleem, S.; Junaid, M.; Ali, A.; Shujait Ali, S.; Khan, M.; Wei, D.Q. Structural insights into the mechanism of RNA recognition by the N-terminal RNA-binding domain of the SARS-CoV-2 nucleocapsid phosphoprotein. Comput. Struct. Biotechnol. J. 2020, 18, 2174–2184. [Google Scholar] [CrossRef]
- Alrasheid, A.A.; Babiker, M.Y.; Awad, T.A. Evaluation of certain medicinal plants compounds as new potential inhibitors of novel corona virus (COVID-19) using molecular docking analysis. Silico Pharmacol. 2021, 9, 10. [Google Scholar] [CrossRef]
- Khan, M.T.; Zeb, M.T.; Ahsan, H.; Ahmed, A.; Ali, A.; Akhtar, K.; Malik, S.I.; Cui, Z.; Ali, S.; Khan, A.S.; et al. SARS-CoV-2 nucleocapsid and Nsp3 binding: An in silico study. Arch. Microbiol. 2021, 203, 59–66. [Google Scholar] [CrossRef]
- Khan, M.T.; Ali, A.; Wang, Q.; Irfan, M.; Khan, A.; Zeb, M.T.; Zhang, Y.-J.; Chinnasamy, S.; Wei, D.-Q. Marine natural compounds as potents inhibitors against the main protease of SARS-CoV-2—A molecular dynamic study. J. Biomol. Struct. Dyn. 2020, 1–11. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Z. Prevention and treatment of viral respiratory infections by traditional Chinese herbs. Chin. Med. J. 2014, 127, 1344–1350. [Google Scholar]
- Remali, J.; Aizat, W.M. A Review on Plant Bioactive Compounds and Their Modes of Action Against Coronavirus Infection. Front. Pharmacol. 2021, 11, 589044. [Google Scholar] [CrossRef]
- Chojnacka, K.; Witek-Krowiak, A.; Skrzypczak, D.; Mikula, K.; Młynarz, P. Phytochemicals containing biologically active polyphenols as an effective agent against Covid-19-inducing coronavirus. J. Funct. Foods 2020, 73, 104146. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Alamri, S.; Al-Whaibi, M.H.; Hussain, Z.; Ali, H.M.; El-Zaidy, M.E. A mini-review of anti-hepatitis B virus activity of medicinal plants. Biotechnol. Biotechnol. Equip. 2016, 31, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-Y.; Chen, C.; Zhang, H.-Q.; Guo, H.-Y.; Wang, H.; Wang, L.; Zhang, X.; Hua, S.-N.; Yu, J.; Xiao, P.-G. Identification of natural compounds with antiviral activities against SARS-associated coronavirus. Antivir. Res. 2005, 67, 18–23. [Google Scholar] [CrossRef]
- Pogrebnyak, N.; Golovkin, M.; Andrianov, V.; Spitsin, S.; Smirnov, Y.; Egolf, R.; Koprowski, H. Severe acute respiratory syndrome (SARS) S protein production in plants: Development of recombinant vaccine. Proc. Natl. Acad. Sci. USA 2005, 102, 9062–9067. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-Y.; Ramalingam, S.; Chye, M.-L. Accumulation of recombinant SARS-CoV spike protein in plant cytosol and chloroplasts indicate potential for development of plant-derived oral vaccines. Exp. Biol. Med. 2006, 231, 1346–1352. [Google Scholar] [CrossRef]
- Vignuzzi, M.; López, C.B. Defective viral genomes are key drivers of the virus–host interaction. Nat. Microbiol. 2019, 4, 1075–1087. [Google Scholar] [CrossRef]
- Haslberger, A.; Jacob, U.; Hippe, B.; Karlic, H. Mechanisms of selected functional foods against viral infections with a view on COVID-19: Mini review. Funct. Foods Heal. Dis. 2020, 10, 195. [Google Scholar] [CrossRef]
- Leung, A.K.; Hon, K.; Leong, K.; Sergi, C. Measles: A disease often forgotten but not gone. Hong Kong Med. J. 2018, 24, 512–520. [Google Scholar] [CrossRef] [Green Version]
- Roy, P. Bluetongue Virus Proteins and Particles and Their Role In Virus Entry, Assembly, And Release. Int. Rev. Cytol. 2005, 64, 69–123. [Google Scholar] [CrossRef]
- Li, T.; Peng, T. Traditional Chinese herbal medicine as a source of molecules with antiviral activity. Antivir. Res. 2013, 97, 1–9. [Google Scholar] [CrossRef]
- Xu, Z.; Peng, C.; Shi, Y.; Zhu, Z.; Mu, K.; Wang, X.; Zhu, W. Nelfinavir was predicted to be a potential inhibitor of 2019-nCov main protease by an integrative approach combining homology modelling, molecular docking and binding free energy calculation. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Morales, A.J.; MacGregor, K.; Kanagarajah, S.; Patel, D.; Schlagenhauf, P. Going global—Travel and the 2019 novel coronavirus. Travel Med. Infect. Dis. 2020, 33, 101578. [Google Scholar] [CrossRef]
- Gould Maule, G. Investigation of Host Factors Required for Lymphocytic Choriomeningitis Virus Entry. Master’s Thesis, University of Ottawa, Ottawa, ON, Canada, 2019. [Google Scholar] [CrossRef]
- Coutard, B.; Valle, C.; De Lamballerie, X.; Canard, B.; Seidah, N.; Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antivir. Res. 2020, 176, 104742. [Google Scholar] [CrossRef]
- Vankadari, N.; Wilce, J.A. Emerging COVID-19 coronavirus: Glycan shield and structure prediction of spike glycoprotein and its interaction with human CD26. Emerg. Microbes Infect. 2020, 9, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Sargiacomo, C.; Sotgia, F.; Lisanti, M.P. COVID-19 and chronological aging: Senolytics and other anti-aging drugs for the treatment or prevention of corona virus infection? Aging 2020, 12, 6511–6517. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Prado, E.; Simbaña-Rivera, K.; Gómez-Barreno, L.; Rubio-Neira, M.; Guaman, L.P.; Kyriakidis, N.C.; López-Cortés, A. Clinical, molecular and epidemiological characterization of the SARS-CoV2 virus and the Coronavirus disease 2019 (COVID-19), a comprehensive literature review. Diagn. Microbiol. Infect. Dis. 2020, 98, 115094. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, X.-J. Potential inhibitors against 2019-nCoV coronavirus M protease from clinically approved medicines. J. Genet. Genom. 2020, 47, 119–121. [Google Scholar] [CrossRef]
- Chang, K.-O.; Kim, Y.; Lovell, S.; Rathnayake, A.D.; Groutas, W.C. Antiviral Drug Discovery: Norovirus Proteases and Development of Inhibitors. Viruses 2019, 11, 197. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-Y.; You, Z.; Wang, Q.; Zhou, Z.-J.; Qiu, Y.; Luo, R.; Ge, X.-Y. The epidemic of 2019-novel-coronavirus (2019-nCoV) pneumonia and insights for emerging infectious diseases in the future. Microbes Infect. 2020, 22, 80–85. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, X.; Lu, Y.; Chen, F.; Zhang, W. Clinical characteristics and therapeutic procedure for four cases with 2019 novel coronavirus pneumonia receiving combined Chinese and Western medicine treatment. Biosci. Trends 2020, 14, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Loizzo, M.R.; Saab, A.M.; Tundis, R.; Statti, G.A.; Menichini, F.; Lampronti, I.; Gambari, R.; Cinatl, J.; Doerr, H.W. Phytochemical Analysis andin vitro Antiviral Activities of the Essential Oils of Seven Lebanon Species. Chem. Biodivers. 2008, 5, 461–470. [Google Scholar] [CrossRef]
- Khaerunnisa, S.; Kurniawan, H.; Awaluddin, R.; Suhartati, S.; Soetjipto, S. Potential inhibitor of COVID-19 main protease (Mpro) from several medicinal plant compounds by molecular docking study. Preprints 2020. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, A.E.; Howard-Varona, C.; Needham, D.M.; John, S.G.; Worden, A.Z.; Sullivan, M.B.; Waldbauer, J.R.; Coleman, M.L. Metabolic and biogeochemical consequences of viral infection in aquatic ecosystems. Nat. Rev. Genet. 2020, 18, 21–34. [Google Scholar] [CrossRef]
- Feng, Y.; Gao, G.F. Towards our understanding of SARS-CoV, an emerging and devastating but quickly conquered virus. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 309–327. [Google Scholar] [CrossRef]
- Kpewou, D.E. Antioxidant Micronutrients Intake in People Living with HIV: Implications on Serum Levels and Liver Function. Master’s Thesis, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana, October 2017. [Google Scholar]
- Chantrill, B.H.; Coulthard, C.E.; Dickinson, L.; Inkley, G.W.; Morris, W.; Pyle, A.H. The Action of Plant Extracts on a Bacteriophage of Pseudomonas pyocyanea and on Influenza A Virus. J. Gen. Microbiol. 1952, 6, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Debiaggi, M.; Pagani, L.; Cereda, P.M.; Landini, P.; Romero, E. Antiviral activity of Chamaecyparis lawsoniana extract: Study with herpes simplex virus type 2. Microbiology 1988, 11, 55–61. [Google Scholar]
- Vermani, K.; Garg, S. Herbal medicines for sexually transmitted diseases and AIDS. J. Ethnopharmacol. 2002, 80, 49–66. [Google Scholar] [CrossRef]
- Asres, K.; Bucar, F. Anti-HIV activity against immunodeficiency virus type 1 (HIV-I) and type II (HIV-II) of compounds isolated from the stem bark of Combretum molle. Ethiop. Med. J. 2005, 43, 15–20. [Google Scholar] [PubMed]
- Kotwal, G.J.; Kaczmarek, J.N.; Leivers, S.; Ghebremariam, Y.T.; Kulkarni, A.P.; Bauer, G.; De Beer, C.; Preiser, W.; Mohamed, A.R. Anti-HIV, Anti-Poxvirus, and Anti-SARS Activity of a Nontoxic, Acidic Plant Extract from the Trifollium Species Secomet-V/anti-Vac Suggests That It Contains a Novel Broad-Spectrum Antiviral. Ann. N. Y. Acad. Sci. 2005, 1056, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.H.; Kwon, H.Y.; Kim, H.J.; Chang, E.J.; Kim, M.B.; Yoon, S.K.; Song, E.Y.; Yoon, D.Y.; Lee, Y.H.; Choi, I.S.; et al. Inhibition of hepatitis B virus by an aqueous extract of Agrimonia eupatoria L. Phytotherapy Res. 2005, 19, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.-L.; Lai, Y.-K.; Lin, C.-C.; Chang, J.-M. Inhibition of hepatitis B virus production byBoehmeria nivearoot extract in HepG2 2.2.15 cells. World J. Gastroenterol. 2006, 12, 5721–5725. [Google Scholar] [CrossRef]
- Webster, D.; Taschereau, P.; Lee, T.D.; Jurgens, T. Immunostimulant properties of Heracleum maximum Bartr. J. Ethnopharmacol. 2006, 106, 360–363. [Google Scholar] [CrossRef]
- Shanti, B.M. Perspective of Potential Plants for Medicine from Rajasthan, India. Int. J. Pharm. Res. 2016, 7, 1–6. [Google Scholar]
- Otake, T.; Mori, H.; Morimoto, M.; Ueba, N.; Sutardjo, S.; Kusumoto, I.T.; Hattori, M.; Namba, T. Screening of Indonesian plant extracts for anti-human immunodeficiency virus—Type 1 (HIV-1) activity. Phytotherapy Res. 1995, 9, 6–10. [Google Scholar] [CrossRef]
- Rege, A.; Chowdhary, A.S. Evaluation of Ocimum sanctum and Tinospora cordifolia as probable HIV protease inhibitors. Int. J. of Pharm. Sci. Rev. Res. 2014, 25, 315–318. [Google Scholar]
- Thayil Seema, M.; Thyagarajan, S. Methanol and aqueous extracts of Ocimum kilimandscharicum (Karpuratulasi) inhibits HIV-1 reverse transcriptase in vitro. Int. J. Pharmacogn. Phytochem. Res. 2016, 8, 1099–1103. [Google Scholar]
- Yu, Y.-B. The extracts of Solanum nigrum L. for inhibitory effects on HIV-1 and its essential enzymes. Korean J. Orient Med. Prescr. 2004, 10, 119–126. [Google Scholar]
- Nair, R. HIV-1 reverse transcriptase inhibition by Vitex negundo L. leaf extract and quantification of flavonoids in relation to anti-HIV activity. J. Cell. Mol. Biol. 2012, 10, 53–59. [Google Scholar]
- Castro, F.; Cardoso, A.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Ryu, Y.B.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, J.-Y.; Kim, D.; Naguyen, T.T.H.; Park, S.-J.; Chang, J.S.; Park, K.H. Biflavonoids from Torreya nucifera displaying SARS-CoV 3CLpro inhibition. Bioorg. Med. Chem. 2010, 18, 7940–7947. [Google Scholar] [CrossRef]
- Park, J.-Y.; Ko, J.-A.; Kim, D.W.; Kim, Y.M.; Kwon, H.-J.; Jeong, H.J.; Kim, C.Y.; Park, K.H.; Lee, W.S.; Ryu, Y.B. Chalcones isolated fromAngelica keiskeiinhibit cysteine proteases of SARS-CoV. J. Enzym. Inhib. Med. Chem. 2016, 31, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Ye, S.-Y.; Yang, Y.; Li, Z.-Y. A review on charred traditional Chinese herbs: Carbonization to yield a haemostatic effect. Pharm. Biol. 2019, 57, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Wen, C.-C.; Shyur, L.-F.; Jan, J.-T.; Liang, P.-H.; Kuo, C.-J.; Arulselvan, P.; Wu, J.-B.; Kuo, S.-C.; Yang, N.-S. Traditional Chinese medicine herbal extracts of Cibotium barometz, Gentiana scabra, Dioscorea batatas, Cassia tora, and Taxillus chinensis inhibit SARS-CoV replication. J. Tradit. Complement. Med. 2011, 1, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.W.; Seo, K.H.; Curtis-Long, M.J.; Oh, K.Y.; Oh, J.-W.; Cho, J.K.; Lee, K.H.; Park, K.H. Phenolic phytochemical displaying SARS-CoV papain-like protease inhibition from the seeds of Psoralea corylifolia. J. Enzym. Inhib. Med. Chem. 2013, 29, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Zhen, H.; Fang, F.; Ye, D.-Y.; Shu, S.-N.; Zhou, Y.-F.; Dong, Y.-S.; Nie, X.-C.; Li, G. Experimental study on the action of allitridin against human cytomegalovirus in vitro: Inhibitory effects on immediate-early genes. Antivir. Res. 2006, 72, 68–74. [Google Scholar] [CrossRef]
- Yu, M.-S.; Lee, J.; Lee, J.M.; Kim, Y.; Chin, Y.-W.; Jee, J.-G.; Keum, Y.-S.; Jeong, Y.-J. Identification of myricetin and scutellarein as novel chemical inhibitors of the SARS coronavirus helicase, nsP13. Bioorg. Med. Chem. Lett. 2012, 22, 4049–4054. [Google Scholar] [CrossRef]
- Zhu, Q.-C.; Wang, Y.; Liu, Y.-P.; Zhang, R.-Q.; Li, X.; Su, W.-H.; Long, F.; Luo, X.-D.; Peng, T. Inhibition of enterovirus 71 replication by chrysosplenetin and penduletin. Eur. J. Pharm. Sci. 2011, 44, 392–398. [Google Scholar] [CrossRef]
- Wen, C.-C.; Kuo, Y.-H.; Jan, J.-T.; Liang, P.-H.; Wang, S.-Y.; Liu, H.-G.; Lee, C.-K.; Chang, S.-T.; Kuo, C.-J.; Lee, S.-S.; et al. Specific Plant Terpenoids and Lignoids Possess Potent Antiviral Activities against Severe Acute Respiratory Syndrome Coronavirus. J. Med. Chem. 2007, 50, 4087–4095. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Dong, X.; Kang, T.G. Activity of in vitro anti-influenza virus of arctigenin. Chin. Tradit. Herb. Drugs 2002, 33, 724–725. [Google Scholar]
- Yi, L.; Li, Z.; Yuan, K.; Qu, X.; Chen, J.; Wang, G.; Zhang, H.; Luo, H.; Zhu, L.; Jiang, P.; et al. Small Molecules Blocking the Entry of Severe Acute Respiratory Syndrome Coronavirus into Host Cells. J. Virol. 2004, 78, 11334–11339. [Google Scholar] [CrossRef] [Green Version]
- Zuo, G.-Y.; Li, Z.-Q.; Chen, L.-R.; Xu, X.-J. In Vitro Anti-HCV Activities of Saxifraga Melanocentra and its Related Polyphenolic Compounds. Antivir. Chem. Chemother. 2005, 16, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Zuo, G.; Li, Z.; Chen, L.; Xu, X. Activity of compounds from Chinese herbal medicine Rhodiola kirilowii (Regel) Maxim against HCV NS3 serine protease. Antivir. Res. 2007, 76, 86–92. [Google Scholar] [CrossRef]
- Guo, Q.; Zhao, L.; You, Q.; Yang, Y.; Gu, H.; Song, G.; Lu, N.; Xin, J. Anti-hepatitis B virus activity of wogonin in vitro and in vivo. Antivir. Res. 2007, 74, 16–24. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, Y.; Ding, X.-R.; Chen, S.-H.; Yang, J.; Wang, X.-J.; Jia, G.-L.; Chen, H.-S.; Bo, X.-C.; Wang, S.-Q. Protocatechuic aldehyde inhibits hepatitis B virus replication both in vitro and in vivo. Antivir. Res. 2007, 74, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-P.; Zhao, W.; Xue, R.; Zhou, Z.-X.; Liu, F.; Han, Y.-X.; Ren, G.; Peng, Z.-G.; Cen, S.; Chen, H.-S.; et al. Oxymatrine inhibits hepatitis B infection with an advantage of overcoming drug-resistance. Antivir. Res. 2011, 89, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, M.; Jiang, H.; Suzuki, Y.; Li, X.; Xiao, P.; Tanaka, T.; Ling, H.; Yang, B.; Saitoh, H.; Zhang, L.; et al. Procyanidins and butanol extract of Cinnamomi Cortex inhibit SARS-CoV infection. Antivir. Res. 2009, 82, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-Y.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, S.-J.; Kim, D.; Park, K.H.; Lee, W.S.; Ryu, Y.B. Diarylheptanoids from Alnus japonica Inhibit Papain-Like Protease of Severe Acute Respiratory Syndrome Coronavirus. Biol. Pharm. Bull. 2012, 35, 2036–2042. [Google Scholar] [CrossRef] [Green Version]
- Chin, L.W.; Cheng, Y.-W.; Lin, S.-S.; Lai, Y.-Y.; Lin, L.-Y.; Chou, M.-Y.; Yang, C.-C.; Chou, M.-C. Anti-herpes simplex virus effects of berberine from Coptidis rhizoma, a major component of a Chinese herbal medicine, Ching-Wei-San. Arch. Virol. 2010, 155, 1933–1941. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, H.J.; Lee, Y.S. A new Anti-HIV Flavonoid Glucuronide fromChrysanthemum morifolium. Planta Medica 2003, 69, 859–861. [Google Scholar] [CrossRef]
- Zhang, H.; Tan, G.T.; Hoang, V.D.; Van Hung, N.; Cuong, N.M.; Soejarto, D.D.; Pezzuto, J.M.; Fong, H.H.S. Natural Anti-HIV Agents. Part IV. Anti-HIV Constituents fromVaticacinerea1. J. Nat. Prod. 2003, 66, 263–268. [Google Scholar] [CrossRef]
- Yang, X.-W.; Zhao, J.; Cui, Y.-X.; Liu, X.-H.; Ma, C.-M.; Hattori, M.; Zhang, L.-H. Anti-HIV-1 Protease Triterpenoid Saponins from the Seeds of Aesculuschinensis. J. Nat. Prod. 1999, 62, 1510–1513. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Li, S.Y.; Huang, R.L.; Wu, M.D.; Huang, H.C.; Lee, K.H. Schizarin B, C, D, and E, Four New Lignans from Kadsuram atsudai and Their Antihepatitis Activities. J. Nat. Prod. 2001, 64, 487–490. [Google Scholar] [CrossRef]
- Kuo, Y.-C.; Kuo, Y.-H.; Lin, Y.-L.; Tsai, W.-J. Yatein from Chamaecyparis obtusa suppresses herpes simplex virus type 1 replication in HeLa cells by interruption the immediate-early gene expression. Antivir. Res. 2006, 70, 112–120. [Google Scholar] [CrossRef]
- Cheng, H.-Y.; Lin, T.-C.; Yang, C.-M.; Wang, K.-C.; Lin, L.-T.; Lin, C.-C. Putranjivain A from Euphorbia jolkini inhibits both virus entry and late stage replication of herpes simplex virus type 2 in vitro. J. Antimicrob. Chemother. 2004, 53, 577–583. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.-C.; Lin, L.-C.; Tsai, W.-J.; Chou, C.-J.; Kung, S.-H.; Ho, Y.-H. Samarangenin B from Limonium sinense Suppresses Herpes Simplex Virus Type 1 Replication in Vero Cells by Regulation of Viral Macromolecular Synthesis. Antimicrob. Agents Chemother. 2002, 46, 2854–2864. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.-C.; Kuo, Y.-C.; Chou, C.-J. Anti-Herpes Simplex Virus Type-1 Flavonoids and a New Flavanone from the Root of Limonium sinense. Planta Medica 2000, 66, 333–336. [Google Scholar] [CrossRef]
- Parida, M.; Upadhyay, C.; Pandya, G.; Jana, A. Inhibitory potential of neem (Azadirachta indica Juss) leaves on Dengue virus type-2 replication. J. Ethnopharmacol. 2002, 79, 273–278. [Google Scholar] [CrossRef]
- Eze, D.C.; Okwor, E.C.; Ehirim, C.H.; Ibu, J.O.; Shoyinka, S.V.O. Comparative evaluation of Moringa oleifera and vacci-boost immuno-modulators in chickens experimentally infected with Newcastle Disease Virus (Kudu 113 Strain). South Asian J. Exp. Biol. 2014, 4, 42–47. [Google Scholar]
- Murakami, A.; Kitazono, Y.; Jiwajinda, S.; Koshimizu, K.; Ohigashi, H. Niaziminin, a thiocarbamate from the leaves of Moringa oleifera, holds a strict structural requirement for inhibition of tumor-promotor-induced Epstein-Barr virus activation. Planta Med. 1998, 64, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Jayasundar, R.; Ghatak, S.; Makhdoomi, M.A.; Luthra, K.; Singh, A.; Velpandian, T. Challenges in integrating component level technology and system level information from Ayurveda: Insights from NMR phytometabolomics and anti-HIV potential of select Ayurvedic medicinal plants. J. Ayurveda Integr. Med. 2019, 10, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Vickers, N.J. Animal Communication: When I’m Calling You, Will You Answer Too? Curr. Biol. 2017, 27, R713–R715. [Google Scholar] [CrossRef] [PubMed]
- Beula, J.M.; Gnanadesigan, M.; Rajkumar, P.B.; Ravikumar, S.; Anand, M. Antiviral, antioxidant and toxicological evaluation of mangrove plant from South East coast of India. Asian Pac. J. Trop. Biomed. 2012, 2, S352–S357. [Google Scholar] [CrossRef]
- Lau, K.M.; Lee, K.M.; Koon, C.M.; Cheung, C.S.; Lau, C.P.; Ho, H.M.; Lee, M.Y.; Au, S.W.; Cheng, C.H.; Lau, C.B.; et al. Immunomodulatory and anti-SARS activities of Houttuynia cordata. J. Ethnopharmacol. 2008, 118, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Tolo, F.M.; Rukunga, G.M.; Muli, F.W.; Njagi, E.N.; Njue, W.; Kumon, K.; Mungai, G.M.; Muthaura, C.N.; Muli, J.M.; Keter, L.K.; et al. Anti-viral activity of the extracts of a Kenyan medicinal plant Carissa edulis against herpes simplex virus. J. Ethnopharmacol. 2006, 104, 92–99. [Google Scholar] [CrossRef]
- Demurtas, O.C.; Massa, S.; Illiano, E.; De Martinis, D.; Chan, P.K.; Di Bonito, P.; Franconi, R. Antigen production in plant to tackle infectious diseases flare up: The case of SARS. Front. Plant. Sci. 2016, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Van Der Meer, F.J.U.M.; De Haan, C.A.M.; Schuurman, N.M.P.; Haijema, B.J.; Verheije, M.H.; Bosch, B.J.; Balzarini, J.; Egberink, H.F. The carbohydrate-binding plant lectins and the non-peptidic antibiotic pradimicin A target the glycans of the coronavirus envelope glycoproteins. J. Antimicrob. Chemother. 2007, 60, 741–749. [Google Scholar] [CrossRef] [Green Version]
- Notka, F.; Meier, G.; Wagner, R. Concerted inhibitory activities of Phyllanthus amarus on HIV replication in vitro and ex vivo. Antivir. Res. 2004, 64, 93–102. [Google Scholar] [CrossRef]
- Das Mahapatra, A.; Bhowmik, P.; Banerjee, A.; Das, A.; Ojha, D.; Chattopadhyay, D. Ethnomedicinal Wisdom. In New Look to Phytomedicine; Elsevier: Amsterdam, The Netherlands, 2019; pp. 35–61. [Google Scholar]
- Mukherjee, H.; Ojha, D.; Bag, P.; Chandel, H.S.; Bhattacharyya, S.; Chatterjee, T.K.; Mukherjee, P.K.; Chakraborti, S.; Chattopadhyay, D. Anti-herpes virus activities of Achyranthes aspera: An Indian ethnomedicine, and its triterpene acid. Microbiol. Res. 2013, 168, 238–244. [Google Scholar] [CrossRef]
- Chen, C.-N.; Lin, C.P.C.; Huang, K.-K.; Chen, W.-C.; Hsieh, H.-P.; Liang, P.-H.; Hsu, J.T.-A. Inhibition of SARS-CoV 3C-like Protease Activity by Theaflavin-3,3′-digallate (TF3). Evid. Based Complement. Altern. Med. 2005, 2, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Arthanari, S.K.; Vanitha, J.; Ganesh, M.; Venkateshwaran, K.; Clercq, D. Evaluation of antiviral and cytotoxic activities of methanolic extract of S. grandiflora (Fabaceae) flowers. Asian Pac. J. Trop. Biomed. 2012, 2, S855–S858. [Google Scholar] [CrossRef]
- Cagno, V.; Civra, A.; Kumar, R.; Pradhan, S.; Donalisio, M.; Sinha, B.N.; Ghosh, M.; Lembo, D. Ficus religiosa L. bark extracts inhibit human rhinovirus and respiratory syncytial virus infection in vitro. J. Ethnopharmacol. 2015, 176, 252–257. [Google Scholar] [CrossRef]
- Jain, M.; Ganju, L.; Katiyal, A.; Padwad, Y.; Mishra, K.; Chanda, S.; Karan, D.; Yogendra, K.; Sawhney, R. Effect of Hippophae rhamnoides leaf extract against Dengue virus infection in human blood-derived macrophages. Phytomedicine 2008, 15, 793–799. [Google Scholar] [CrossRef]
- Yamai, M.; Tsumura, K.; Kimura, M.; Fukuda, S.; Murakami, T.; Kimura, Y. Antiviral Activity of a Hot Water Extract of Black Soybean against a Human Respiratory Illness Virus. Biosci. Biotechnol. Biochem. 2003, 67, 1071–1079. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Aeri, V.; Gaur, P.K.; Jachak, S.M. Phytochemical, Therapeutic, and Ethnopharmacological Overview for a Traditionally Important Herb: Boerhavia diffusa Linn. BioMed. Res. Int. 2014, 2014, 808302. [Google Scholar] [CrossRef] [Green Version]
- Keyaerts, E.; Vijgen, L.; Pannecouque, C.; Van Damme, E.; Peumans, W.; Egberink, H.; Balzarini, J.; Van Ranst, M. Plant lectins are potent inhibitors of coronaviruses by interfering with two targets in the viral replication cycle. Antivir. Res. 2007, 75, 179–187. [Google Scholar] [CrossRef]
- Prathapan, A.; Vineetha, V.P.; Abhilash, P.A.; Raghu, K.G. Boerhaaviadiffusa, L. attenuates angiotensin II-induced hypertrophy in H9c2 cardiac myoblast cells via modulating oxidative stress and down-regulating NF-κβ and transforming growth factor β1. Br. J. Nutr. 2013, 110, 1201–1210. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.Y.; Kumar, V. Mechanism & inhibition kinetics of bioassay-guided fractions of Indian medicinal plants and foods as ACE inhibitors. J. Tradit. Complement. Med. 2019, 9, 73–84. [Google Scholar] [CrossRef]
- Olivieri, F.; Prasad, V.; Valbonesi, P.; Srivastava, S.; Ghosal-Chowdhury, P.; Barbieri, L.; Bolognesi, A.; Stirpe, F. A systemic antiviral resistance-inducing protein isolated fromClerodendrum inermeGaertn. is a polynucleotide:adenosine glycosidase (ribosome-inactivating protein). FEBS Lett. 1996, 396, 132–134. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Standardized Clitoria ternatea leaf extract as hyaluronidase, elastase and matrix-metalloproteinase-1 inhibitor. Indian J. Pharmacol. 2012, 44, 584–587. [Google Scholar] [CrossRef] [Green Version]
- Bigoniya, P.; Patel, K.K.; Pandey, A.; Raj, V. Pharmacological screening of Coriandrum sativum Linn. for hepatoprotective activity. J. Pharm. Bioallied Sci. 2011, 3, 435–441. [Google Scholar] [CrossRef]
- Vimalanathan, S.; Ignacimuthu, S.; Hudson, J.B. Medicinal plants of Tamil Nadu (Southern India) are a rich source of antiviral activities. Pharm. Biol. 2009, 47, 422–429. [Google Scholar] [CrossRef]
- Gilani, A.H.; Khan, A.-U.; Raoof, M.; Ghayur, M.N.; Siddiqui, B.; Vohra, W.; Begum, S. Gastrointestinal, selective airways and urinary bladder relaxant effects of Hyoscyamus niger are mediated through dual blockade of muscarinic receptors and Ca2+channels. Fundam. Clin. Pharmacol. 2008, 22, 87–99. [Google Scholar] [CrossRef]
- He, L.; Qi, Y.; Rong, X.; Jiang, J.; Yang, Q.; Yamahara, J.; Murray, M.; Li, Y. The Ayurvedic MedicineSalacia oblongaAttenuates Diabetic Renal Fibrosis in Rats: Suppression of Angiotensin II/AT1 Signaling. Evid. Based Complement. Altern. Med. 2011, 2011, 807451. [Google Scholar] [CrossRef] [Green Version]
- Ganjhu, R.K.; Mudgal, P.P.; Maity, H.; Dowarha, D.; Devadiga, S.; Nag, S.; Arunkumar, G. Herbal plants and plant preparations as remedial approach for viral diseases. Virus Dis. 2015, 26, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Galani, V.J.; Patel, B.G.; Rana, D.G. Sphaeranthus indicus Linn.: A phytopharmacological review. Int. J. Ayurveda Res. 2010, 1, 247–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, B.K.; Khosa, R.L. Hepatoprotective and antioxidant effect of Sphaeranthus indicus against acetaminophen-induced hepatotoxicity in rats. J. Pharm Sci Res. 2009, 1, 26–30. [Google Scholar]
- Tsai, Y.-C.; Lee, C.-L.; Yen, H.-R.; Chang, Y.-S.; Lin, Y.-P.; Huang, S.-H.; Lin, C.-W. Antiviral Action of Tryptanthrin Isolated from Strobilanthes cusia Leaf against Human Coronavirus NL63. Biomolecules 2020, 10, 366. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.P.S.; Bhowmik, D.; Dutta, A.; Yadav, A.P.; Paswan, S.; Srivastava, S.; Deb, L. Recent trends in potential traditional Indian herbs Emblica officinalis and its medicinal importance. J. Pharmacogn. Phytochem. 2012, 1, 18–28. [Google Scholar]
- Edai, Z.-C.; Fu, W.; Ewan, L.-Y.; Ecai, H.-H.; Ewang, N.; Qi, S.-S.; Du, D.-L. Different Growth Promoting Effects of Endophytic Bacteria on Invasive and Native Clonal Plants. Front. Plant Sci. 2016, 7, 706. [Google Scholar] [CrossRef] [Green Version]
- Puri, A.; Saxena, R.P.; Saxena, K.; Srivastava, V.; Tandon, J. Immunostimulant activity of Nyctanthes arbor-tristis L. J. Ethnopharmacol. 1994, 42, 31–37. [Google Scholar] [CrossRef]
- Ejantan, I.; Eahmad, W.; Ebukhari, S.N.A. Plant-derived immunomodulators: An insight on their preclinical evaluation and clinical trials. Front. Plant Sci. 2015, 6, 655. [Google Scholar] [CrossRef]
- Mohammadi, S.; Jafari, B.; Asgharian, P.; Martorell, M.; Sharifi-Rad, J. Medicinal plants used in the treatment of Malaria: A key emphasis to Artemisia, Cinchona, Cryptolepis and Tabebuia genera. Phytother. Res. 2020, 34, 1556–1569. [Google Scholar] [CrossRef]
- Brattig, N.; Diao, G.-J.; Berg, P. Immunoenhancing effect of flavonoid compounds on lymphocyte proliferation and immunoglobulin synthesis. Int. J. Immunopharmacol. 1984, 6, 205–215. [Google Scholar] [CrossRef]
- Yadav, V.S.; Mishra, K.P.; Singh, D.P.; Mehrotra, S.; Singh, V.K. Immunomodulatory Effects of Curcumin. Immunopharmacol. Immunotoxicol. 2005, 27, 485–497. [Google Scholar] [CrossRef]
- Fürst, R.; Zündorf, I. Plant-Derived Anti-Inflammatory Compounds: Hopes and Disappointments regarding the Translation of Preclinical Knowledge into Clinical Progress. Mediat. Inflamm. 2014, 2014, 146832. [Google Scholar] [CrossRef] [Green Version]
- Ložienė, K.; Švedienė, J.; Paškevičius, A.; Raudonienė, V.; Sytar, O.; Kosyan, A. Influence of plant origin natural α-pinene with different enantiomeric composition on bacteria, yeasts and fungi. Fitoterapia 2018, 127, 20–24. [Google Scholar] [CrossRef]
- Lecher, J.C.; Diep, N.; Krug, P.W.; Hilliard, J.K. Genistein Has Antiviral Activity against Herpes B Virus and Acts Synergistically with Antiviral Treatments to Reduce Effective Dose. Viruses 2019, 11, 499. [Google Scholar] [CrossRef] [Green Version]
- Malaguarnera, L. Influence of Resveratrol on the Immune Response. Nutrients 2019, 11, 946. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Mishra, K.P.; Ganju, L. Broad-spectrum antiviral properties of andrographolide. Arch. Virol. 2017, 162, 611–623. [Google Scholar] [CrossRef]
- Perricone, C.; Triggianese, P.; Bartoloni, E.; Cafaro, G.; Bonifacio, A.F.; Bursi, R.; Perricone, R.; Gerli, R. The anti-viral facet of anti-rheumatic drugs: Lessons from COVID-19. J. Autoimmun. 2020, 111, 102468. [Google Scholar] [CrossRef]
- Parvez, M.K.; Rehman, T.; Alam, P.; Al-Dosari, M.S.; Alqasoumi, S.I.; Alajmi, M.F. Plant-derived antiviral drugs as novel hepatitis B virus inhibitors: Cell culture and molecular docking study. Saudi Pharm. J. 2019, 27, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Sarker, M.R.; Ming, L.C.; Mohamed, I.N.; Zhao, C.; Sheikh, B.Y.; Tsong, H.F.; Rashid, M.A. Comprehensive Review on Phytochemicals, Pharmacological and Clinical Potentials of Gymnema sylvestre. Front. Pharmacol. 2019, 10, 1223. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, X.; Li, W. Tetrandrine, a Chinese plant-derived alkaloid, is a potential candidate for cancer chemotherapy. Oncotarget 2016, 7, 40800–40815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Kwak, J. Antiviral phenolic compounds from the whole plants of Zostera marina against influenza A virus. Planta Medica 2015, 81, PW_06. [Google Scholar] [CrossRef]
- Naser, B.; Bodinet, C.; Tegtmeier, M.; Lindequist, U. Thuja occidentalis(Arbor vitae): A Review of its Pharmaceutical, Pharmacological and Clinical Properties. Evid. Based Complement. Altern. Med. 2005, 2, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [Green Version]
- Andarwulan, N.; Batari, R.; Sandrasari, D.A.; Bolling, B.; Wijaya, H. Flavonoid content and antioxidant activity of vegetables from Indonesia. Food Chem. 2010, 121, 1231–1235. [Google Scholar] [CrossRef] [Green Version]
- Lama-Muñoz, A.; Contreras, M.D.M.; Espínola, F.; Moya, M.; Romero, I.; Castro, E. Optimization of Oleuropein and Luteolin-7-O-Glucoside Extraction from Olive Leaves by Ultrasound-Assisted Technology. Energies 2019, 12, 2486. [Google Scholar] [CrossRef] [Green Version]
- Cahyono, B.; Ariani, J.; Failasufa, H.; Suzery, M.; Susanti, S.; Hadiyanto, H. Extraction of homologous compounds of curcuminoid isolated from temulawak (Curcuma xanthorriza Roxb.) plant. Rasayan J. Chem. 2019, 12, 7–13. [Google Scholar] [CrossRef]
- Rosidi, A.; Khomsan, A.; Setiawan, B.I.; Riyadi, H.; Briawan, D. Antioxidant Potential of Temulawak (Curcuma xanthorrhiza roxb). Pak. J. Nutr. 2016, 15, 556–560. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The Therapeutic Potential of Naringenin: A Review of Clinical Trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Meirinhos, J.; Silva, B.M.; Valentão, P.; Seabra, R.M.; Pereira, J.A.; Dias, A.; Andrade, P.B.; Ferreres, F. Analysis and quantification of flavonoidic compounds from Portuguese olive (Olea Europaea L.) leaf cultivars. Nat. Prod. Res. 2005, 19, 189–195. [Google Scholar] [CrossRef]
- Nicolì, F.; Negro, C.; Vergine, M.; Aprile, A.; Nutricati, E.; Sabella, E.; Miceli, A.; Luvisi, A.; De Bellis, L. Evaluation of Phytochemical and Antioxidant Properties of 15 Italian Olea europaea L. Cultivar Leaves. Molecules 2019, 24, 1998. [Google Scholar] [CrossRef] [Green Version]
- Miean, K.H.; Mohamed, S. Flavonoid (Myricetin, Quercetin, Kaempferol, Luteolin, and Apigenin) Content of Edible Tropical Plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef]
- De Maat, M.; Pijl, H.; Kluft, C.; Princen, H. Consumption of black and green tea has no effect on inflammation, haemostasis and endothelial markers in smoking healthy individuals. Eur. J. Clin. Nutr. 2000, 54, 757–763. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, R.M.M. Quantification of catechins and caffeine from green tea (Camellia sinensis) infusions, extract, and ready-to-drink beverages. Food Sci. Technol. 2012, 32, 163–166. [Google Scholar] [CrossRef] [Green Version]
- Sanwal, S.K.; Yadav, R.K.; Singh, P.K.; Buragohain, J.; Verma, M.R. Gingerol content of different genotypes of ginger (Zingiber officinale). Indian J. Agric. Sci. 2010, 80, 258. [Google Scholar]
- Li, L.-L.; Cui, Y.; Guo, X.-H.; Ma, K.; Tian, P.; Feng, J.; Wang, J.-M. Pharmacokinetics and Tissue Distribution of Gingerols and Shogaols from Ginger (Zingiber officinale Rosc.) in Rats by UPLC–Q-Exactive–HRMS. Molecules 2019, 24, 512. [Google Scholar] [CrossRef] [Green Version]
- Lelešius, R.; Karpovaitė, A.; Mickienė, R.; Drevinskas, T.; Tiso, N.; Ragažinskienė, O.; Kubilienė, L.; Maruška, A.; Šalomskas, A. In vitro antiviral activity of fifteen plant extracts against avian infectious bronchitis virus. BMC Vet. Res. 2019, 15, 178. [Google Scholar] [CrossRef] [Green Version]
- Dolzhenko, Y.; Bertea, C.M.; Occhipinti, A.; Bossi, S.; Maffei, M. UV-B modulates the interplay between terpenoids and flavonoids in peppermint (Mentha piperita L.). J. Photochem. Photobiol. B Biol. 2010, 100, 67–75. [Google Scholar] [CrossRef]
- Fatih, B.; Madani, K.; Chibane, M.; Duez, P.; Brahmi, F.; Khodir, M.; Mohamed, C.; Pierre, D. Chemical Composition and Biological Activities of Mentha Species. In Aromatic and Medicinal Plants—Back to Nature; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Porte, A.; Godoy, R. Chemical composition of Thymus vulgaris L. (Thyme) essential oil from the Rio de Janeiro state, Brazil. J. Serb. Chem. Soc. 2008, 73, 307–310. [Google Scholar] [CrossRef]
- Gedikoğlu, A.; Sökmen, M.; Çivit, A. Evaluation of Thymus vulgaris and Thymbra spicata essential oils and plant extracts for chemical composition, antioxidant, and antimicrobial properties. Food Sci. Nutr. 2019, 7, 1704–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puodziuniene, G.; Kairyte, V.; Janulis, V.; Razukas, A.; Barsteigiene, Z.; Ragažinskienė, O. Quantitative hplc estimation of flavonoids in showy tick trefoil (Desmodium canadense) herbs. Pharm. Chem. J. 2011, 45, 88–90. [Google Scholar] [CrossRef]
- Batyuk, V.S.V.; Vasil’eva, L.N.; Chernobrovaya, N.V.; Komissarenko, N.F. Flavonoids of Desmodium canadense and their analgesic effect. Khim. Farm. Zh. 1987, 21, 63–67. [Google Scholar]
- Taylor, W.G.; Sutherland, D.H.; Richards, K.W. Soyasaponins and Related Glycosides of Desmodium canadense and Desmodium illinoense. Open Nat. Prod. J. 2009, 2, 59–67. [Google Scholar] [CrossRef]
- Ma, X.; Zheng, C.; Hu, C.; Rahman, K.; Qin, L. The genus Desmodium (Fabaceae)-traditional uses in Chinese medicine, phytochemistry and pharmacology. J. Ethnopharmacol. 2011, 138, 314–332. [Google Scholar] [CrossRef]
- Yang, J.-L.; Ha, T.-K.-Q.; Dhodary, B.; Pyo, E.; Nguyen-Ngoc, H.; Cho, H.; Kim, E.; Oh, W.K. Oleanane Triterpenes from the Flowers ofCamellia japonicaInhibit Porcine Epidemic Diarrhea Virus (PEDV) Replication. J. Med. Chem. 2015, 58, 1268–1280. [Google Scholar] [CrossRef]
- Yang, J.-L.; Ha, T.K.Q.; Oh, W.K. Discovery of inhibitory materials against PEDV corona virus from medicinal plants. Jpn. J. Vet. Res. 2016, 64, S53–S63. [Google Scholar]
- Azuma, C.M.; Dos Santos, F.C.S.; Lago, J.H.G. Flavonoids and fatty acids of Camellia japonica leaves extract. Rev. Bras. Farm. 2011, 21, 1159–1162. [Google Scholar] [CrossRef] [Green Version]
- Itokawa, H.; Nakajima, H.; Ikuta, A.; Iitaka, Y. Two triterpenes from the flowers of Camellia japonica. Phytochemistry 1981, 20, 2539–2542. [Google Scholar] [CrossRef]
- Kato, M.; Ashihara, H. Biosynthesis and Catabolism of Purine Alkaloids in Camellia Plants. Nat. Prod. Commun. 2008, 3, 1934578 0800300907. [Google Scholar] [CrossRef] [Green Version]
- Karioti, A.; Bilia, A.R.; Skaltsa, H. Quercus ilex L.: A rich source of polyacylated flavonoid glucosides. Food Chem. 2010, 123, 131–142. [Google Scholar] [CrossRef]
- Cheng, P.-W.; Huang, L.-T.; Chiang, L.-C.; Lin, C.-C. Antiviral effects of saikosaponins on human coronavirus 229e in vitro. Clin. Exp. Pharmacol. Physiol. 2006, 33, 612–616. [Google Scholar] [CrossRef]
- Yang, F.; Dong, X.; Yin, X.; Wang, W.; You, L.; Ni, J. Radix Bupleuri: A Review of Traditional Uses, Botany, Phytochemistry, Pharmacology, and Toxicology. BioMed. Res. Int. 2017, 2017, 7597596. [Google Scholar] [CrossRef] [Green Version]
- Hemalatha, S.; Kumar, M.; Prasad, S. A current update on the phytopharmacological aspects of Houttuynia cordata Thunb. Pharmacogn. Rev. 2014, 8, 22–35. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-W.; Tsai, F.-J.; Tsai, C.-H.; Lai, C.-C.; Wan, L.; Ho, T.-Y.; Hsieh, C.-C.; Chao, P.-D.L. Anti-SARS coronavirus 3C-like protease effects of Isatis indigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef]
- Speranza, J.; Miceli, N.; Taviano, M.F.; Ragusa, S.; Kwiecień, I.; Szopa, A.; Ekiert, H. Isatis tinctoria L. (Woad): A Review of Its Botany, Ethnobotanical Uses, Phytochemistry, Biological Activities, and Biotechnological Studies. Plants 2020, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, X.-Q.; Yin, Z.-Q.; Wang, Y.; Ye, W.-C. Two New Amaryllidaceae Alkaloids from the Bulbs of Lycoris radiata. Chem. Pharm. Bull. 2009, 57, 610–611. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Zhang, C.; Guo, M. Comparative Analysis of Amaryllidaceae Alkaloids from Three Lycoris Species. Molecules 2015, 20, 21854–21869. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.; Yue, Y.; Shi, M.; Chen, M.; Yang, X.; Wang, L. Exploration of Floral Volatile Organic Compounds in Six Typical Lycoris taxa by GC-MS. Plants 2019, 8, 422. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.J.; Su, X.J.; Yu, H.P.; Li, J.; Qin, Y.J.; Xu, Q.; Luo, W.-S. A study on anti-SARS-CoV 3CL protein of flavonoids from litchi chinensis sonn core. Chin. Pharmacol. 2008, 24, 699–700. [Google Scholar]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.-S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzym. Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimmanpipug, P.; Lee, V.S.; Wolschann, P.; Hannongbua, S. Litchi chinensis-derived terpenoid as anti-HIV-1 protease agent: Structural design from molecular dynamics simulations. Mol. Simul. 2009, 35, 673–680. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Mohamed, G.A. Litchi chinensis: Medicinal uses, phytochemistry, and pharmacology. J. Ethnopharmacol. 2015, 174, 492–513. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Min, J.S.; Jang, M.S.; Lee, J.Y.; Shin, Y.S.; Park, C.M.; Song, J.H.; Kim, H.R.; Kim, S.; Jin, Y.-H.; et al. Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells. Biomolecules 2019, 9, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): A review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J. Pharm. Pharmacol. 2019, 71, 1353–1369. [Google Scholar] [CrossRef] [Green Version]
- Shojai, T.M.; Langeroudi, A.G.; Karimi, V.; Barin, A.; Sadri, N. The effect of Allium sativum (Garlic) extract on infectious bronchitis virus in specific pathogen free embryonic egg. Avicenna J. Phytomed. 2016, 6, 458. [Google Scholar]
- Weber, N.D.; Andersen, D.O.; North, J.A.; Murray, B.K.; Lawson, L.D.; Hughes, B.G. In VitroVirucidal Effects of Allium sativum (Garlic) Extract and Compounds. Planta Med. 1992, 58, 417–423. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Binduga, U.E.; Rybczyńska-Tkaczyk, K.; Leja, M.L.; Gmiński, J. Cytotoxic effects of two extracts from garlic (Allium sativum L.) cultivars on the human squamous carcinoma cell line SCC-15. Saudi J. Biol. Sci. 2018, 25, 1703–1712. [Google Scholar] [CrossRef] [Green Version]
- Pontin, M.; Bottini, R.; Burba, J.L.; Piccoli, P. Allium sativum produces terpenes with fungistatic properties in response to infection with Sclerotium cepivorum. Phytochemistry 2015, 115, 152–160. [Google Scholar] [CrossRef]
- Shin, S.-W. In vitro Effects of Essential Oils from the Aerial Parts of Artemisia annua L. Against Antibiotic-Susceptible and-Resistant Strains of Salmenella typhimurium. Yakhak Hoeji 2007, 51, 355–360. [Google Scholar]
- Nigam, M.; Atanassova, M.; Mishra, A.P.; Pezzani, R.; Devkota, H.P.; Plygun, S.; Salehi, B.; Setzer, W.N.; Sharifi-Rad, J. Bioactive Compounds and Health Benefits of Artemisia Species. Nat. Prod. Commun. 2019, 14, 19850354. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Bais, S.; Gill, N.S.; Rana, N.; Shandil, S. A Phytopharmacological Review on a Medicinal Plant: Juniperus communis. Int. Sch. Res. Not. 2014, 2014, 634723. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Kim, J.H.; Kwon, J.M.; Kwon, H.J.; Jeong, H.J.; Kim, Y.M. Dieckol, a SARS-CoV 3CL inhibitor, isolated from the edible brown algae Ecklonia cava. Phytochemistry 2013, 20, 2539–2542. [Google Scholar]
- Van Damme, E.J.M.; Peumans, W.J.; Barre, A.; Rougé, P. Plant Lectins: A Composite of Several Distinct Families of Structurally and Evolutionary Related Proteins with Diverse Biological Roles. Crit. Rev. Plant Sci. 1998, 17, 575–692. [Google Scholar] [CrossRef]
- Müller, W.E.; Renneisen, K.; Kreuter, M.H.; Schröder, H.C.; Winkler, I. The D-mannose-specific lectin from Gerardia savaglia blocks binding of human immunodeficiency virus type I to H9 cells and human lymphocytes in vitro. J. Acquir. Immune Defic. Syndr. 1988, 1, 453–458. [Google Scholar]
- Hammar, L.; Eriksson, S.; Morein, B. Human Immunodeficiency Virus Glycoproteins: Lectin Binding Properties. AIDS Res. Hum. Retrovir. 1989, 5, 495–506. [Google Scholar] [CrossRef]
- Hansen, J.-E.S.; Nielsen, C.; Heegaard, P.; Mathiesen, L.R.; Nielsen, J.O.; Nielsen, C. Correlation between carbohydrate structures on the envelope glycoprotein gp120 of.HIV-1 and HIV-2 and syncytium inhibition with lectins. AIDS 1989, 3, 635–642. [Google Scholar] [CrossRef]
- Matsui, T.; Kobayashi, S.; Yoshida, O.; Ishii, S.-I.; Abe, Y.; Yamamoto, N. Effects of succinylated concanavalin A on infectivity and syncytial formation of human immunodeficiency virus. Med. Microbiol. Immunol. 1990, 179, 225–235. [Google Scholar] [CrossRef]
- Balzarini, J.; Schols, D.; Neyts, J.; Van Damme, E.; Peumans, W.; De Clercq, E. Alpha-(1-3)- and alpha-(1-6)-D-mannose-specific plant lectins are markedly inhibitory to human immunodeficiency virus and cytomegalovirus infections in vitro. Antimicrob. Agents Chemother. 1991, 35, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Balzarini, J.; Neyts, J.; Schols, D.; Hosoya, M.; Van Damme, E.; Peumans, W.; De Clercq, E. The mannose-specific plant lectins from Cymbidium hybrid and Epipactis helleborine and the (N-acetylglucosamine)n-specific plant lectin from Urtica dioica are potent and selective inhibitors of human immunodeficiency virus and cytomegalovirus replication in vitro. Antivir. Res. 1992, 18, 191–207. [Google Scholar] [CrossRef]
- Balzarini, J.; Hatse, S.; Vermeire, K.; Princen, K.; Aquaro, S.; Perno, C.-F.; De Clercq, E.; Egberink, H.; Mooter, G.V.; Peumans, W.; et al. Mannose-Specific Plant Lectins from the Amaryllidaceae Family Qualify as Efficient Microbicides for Prevention of Human Immunodeficiency Virus Infection. Antimicrob. Agents Chemother. 2004, 48, 3858–3870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krokhin, O.; Li, Y.; Andonov, A.; Feldmann, H.; Flick, R.; Jones, S.; Stroeher, U.; Bastien, N.; Dasuri, K.V.N.; Cheng, K.; et al. Mass Spectrometric Characterization of Proteins from the SARS Virus. Mol. Cell. Proteom. 2003, 2, 346–356. [Google Scholar] [CrossRef]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.-P.; Desprez, P.-Y.; Krtolica, A.; Campisi, J. The Senescence-Associated Secretory Phenotype: The Dark Side of Tumor Suppression. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 99–118. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, L.; Castillo, J.; Quiñones, M.; Garcia-Vallvé, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of Angiotensin-Converting Enzyme Activity by Flavonoids: Structure-Activity Relationship Studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Li, R.; Li, X.; He, J.; Jiang, S.; Liu, S.; Yang, J. Quercetin as an Antiviral Agent Inhibits Influenza A Virus (IAV) Entry. Viruses 2015, 8, 6. [Google Scholar] [CrossRef]
- Häckl, L.; Cuttle, G.; Dovichi, S.S.; Lima-Landman, M.; Nicolau, M. Inhibition of Angiotensin-Converting Enzyme by Quercetin Alters the Vascular Response to Bradykinin and Angiotensin I. Pharmacology 2002, 65, 182–186. [Google Scholar] [CrossRef]
- Thaler, R.; Maurizi, A.; Roschger, P.; Sturmlechner, I.; Khani, F.; Spitzer, S.; Rumpler, M.; Zwerina, J.; Karlic, H.; Dudakovic, A.; et al. Anabolic and Antiresorptive Modulation of Bone Homeostasis by the Epigenetic Modulator Sulforaphane, a Naturally Occurring Isothiocyanate. J. Biol. Chem. 2016, 291, 6754–6771. [Google Scholar] [CrossRef] [Green Version]
- Olesin, E.; Nayar, R.; Saikumar-Lakshmi, P.; Berg, L.J. The Transcription Factor Runx2 Is Required for Long-Term Persistence of Antiviral CD8+ Memory T Cells. ImmunoHorizons 2018, 2, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Müller, L.; Meyer, M.; Bauer, R.N.; Zhou, H.; Zhang, H.; Jones, S.; Robinette, C.; Noah, T.L.; Jaspers, I. Effect of Broccoli Sprouts and Live Attenuated Influenza Virus on Peripheral Blood Natural Killer Cells: A Randomized, Double-Blind Study. PLoS ONE 2016, 11, e0147742. [Google Scholar] [CrossRef] [Green Version]
- Marinella, M.A. Indomethacin and resveratrol as potential treatment adjuncts for SARS-CoV-2/COVID-19. Int. J. Clin. Pract. 2020, 74, e13535. [Google Scholar] [CrossRef]
- Medina-Bolivar, F.; Condori, J.; Rimando, A.M.; Hubstenberger, J.; Shelton, K.; O’Keefe, S.F.; Bennett, S.; Dolan, M.C. Production and secretion of resveratrol in hairy root cultures of peanut. Phytochemistry 2007, 68, 1992–2003. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, X.; Martin, C. Scutellaria baicalensis, the golden herb from the garden of Chinese medicinal plants. Sci. Bull. 2016, 61, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Sowndhararajan, K.; Deepa, P.; Kim, M.; Park, S.J.; Kim, S. Neuroprotective and Cognitive Enhancement Potentials of Baicalin: A Review. Brain Sci. 2018, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Yonesi, M.; Rezazadeh, A. Plants as a prospective source of natural anti-viral compounds and oral vaccines against COVID-19 coronavirus. Preprints 2020. [Google Scholar] [CrossRef]
- Liu, H.; Ye, F.; Sun, Q.; Liang, H.; Li, C.; Li, S.; Lu, R.; Huang, B.; Tan, W.; Lai, L. Scutellaria baicalensis extract and baicalein inhibit replication of SARS-CoV-2 and its 3C-like protease in vitro. J. Enzym. Inhib. Med. Chem. 2021, 36, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.L.; Wahid, F.; Khan, N.; Farooq, U.; Shah, A.J.; Tareen, S.; Ahmad, F.; Khan, T. Inhibitory Effects ofGlycyrrhiza glabraand Its Major Constituent Glycyrrhizin on Inflammation-Associated Corneal Neovascularization. Evid. Based Complement. Altern. Med. 2018, 2018, 8438101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilcher, H. Liquorice may tackle SARS. Nature 2003. [Google Scholar] [CrossRef]
- Chen, H.; Du, Q. Potential natural compounds for preventing SARS-CoV-2 (2019-nCoV) infection. Preprints 2020. [Google Scholar] [CrossRef]
- Dubey, K.; Dubey, R. Computation screening of narcissoside a glycosyloxyflavone for potential novel coronavirus 2019 (COVID-19) inhibitor. Biomed. J. 2020, 43, 363–367. [Google Scholar] [CrossRef]
- Akram, M.; Shahab-Uddin, A.A.; Usmanghani, K.; Hannan, A.; Mohiuddin, E.; Asif, M. Curcuma longa and curcumin: A review article. Rom. J. Biol Plant. Biol 2010, 55, 65–70. [Google Scholar]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Praditya, D.; Kirchhoff, L.; Brüning, J.; Rachmawati, H.; Steinmann, J.; Steinmann, E. Anti-infective Properties of the Golden Spice Curcumin. Front. Microbiol. 2019, 10, 912. [Google Scholar] [CrossRef] [Green Version]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; Da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Bansal, T.; Awasthi, A.; Jaggi, M.; Khar, R.K.; Talegaonkar, S. Pre-clinical evidence for altered absorption and biliary excretion of irinotecan (CPT-11) in combination with quercetin: Possible contribution of P-glycoprotein. Life Sci. 2008, 83, 250–259. [Google Scholar] [CrossRef]
- Shoba, G.; Joy, D.; Joseph, T.; Majeed, M.; Rajendran, R.; Srinivas, P.S.S.R. Influence of Piperine on the Pharmacokinetics of Curcumin in Animals and Human Volunteers. Planta Med. 1998, 64, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Basu, P.; Dutta, S.; Begum, R.; Mittal, S.; Das Dutta, P.; Bharti, A.C.; Panda, C.K.; Biswas, J.; Dey, B.; Talwar, G.P.; et al. Clearance of cervical human papillomavirus infection by topical application of curcumin and curcumin containing polyherbal cream: A phase II randomized controlled study. Asian Pac. J. Cancer Prev. 2013, 14, 5753–5759. [Google Scholar] [CrossRef] [Green Version]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Utomo, R.Y.; Meiyanto, E. Revealing the potency of citrus and galangal constituents to halt SARS-CoV-2 infection. Preprints 2020. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, D.; Stewart, A.J.; Mullen, W.; Burns, J.; Lean, M.E.J.; Brighenti, A.F.; Crozier, A. HPLC-MSnAnalysis of Phenolic Compounds and Purine Alkaloids in Green and Black Tea. J. Agric. Food Chem. 2004, 52, 2807–2815. [Google Scholar] [CrossRef]
- Lucas, L.; Russell, A.; Keast, R. Molecular Mechanisms of Inflammation. Anti-Inflammatory Benefits of Virgin Olive Oil and the Phenolic Compound Oleocanthal. Curr. Pharm. Des. 2011, 17, 754–768. [Google Scholar] [CrossRef]
- Remely, M.; Ferk, F.; Sterneder, S.; Setayesh, T.; Kepcija, T.; Roth, S.; Noorizadeh, R.; Greunz, M.; Rebhan, I.; Wagner, K.-H.; et al. Vitamin E Modifies High-Fat Diet-Induced Increase of DNA Strand Breaks, and Changes in Expression and DNA Methylation of Dnmt1 and MLH1 in C57BL/6J Male Mice. Nutrients 2017, 9, 607. [Google Scholar] [CrossRef] [Green Version]
- Kaihatsu, K.; Yamabe, M.; Ebara, Y. Antiviral Mechanism of Action of Epigallocatechin-3-O-gallate and Its Fatty Acid Esters. Molecules 2018, 23, 2475. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, C.; Hrincius, E.R.; Korte, V.; Mazur, I.; Droebner, K.; Poetter, A.; Dreschers, S.; Schmolke, M.; Planz, O.; Ludwig, S. A polyphenol rich plant extract, CYSTUS052, exerts anti influenza virus activity in cell culture without toxic side effects or the tendency to induce viral resistance. Antivir. Res. 2007, 76, 38–47. [Google Scholar] [CrossRef]
- Barak, V.; Halperin, T.; Kalickman, I. The effect of Sambucol, a black elderberry-based, natural product, on the production of human cytokines: I. Inflammatory cytokines. Eur. Cytokine Netw. 2001, 12, 290–296. [Google Scholar]
- Efferth, T.; Romero, M.R.; Wolf, D.G.; Stamminger, T.; Marin, J.J.G.; Marschall, M. The Antiviral Activities of Artemisinin and Artesunate. Clin. Infect. Dis. 2008, 47, 804–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamoddini, M.K.; Emami, S.A.; Ghannad, M.S.; Sani, E.A.; Sahebkar, A. Antiviral activities of aerial subsets of Artemisia species against Herpes Simplex virus type 1 (HSV1) in vitro. Asian Biomed. 2011, 5, 63–68. [Google Scholar] [CrossRef]
- Khan, M.A.A.; Jain, D.; Bhakuni, R.; Zaim, M.; Thakur, R. Occurrence of some antiviral sterols in Artemisia annua. Plant Sci. 1991, 75, 161–165. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Luthria, D.L.; Sasaki, T.; Heyerick, A. Flavonoids from Artemisia annua L. as Antioxidants and Their Potential Synergism with Artemisinin against Malaria and Cancer. Molecules 2010, 15, 3135–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, B.J. Antioxidants as Potential Therapeutics for Lung Fibrosis. Antioxid. Redox Signal. 2008, 10, 355–370. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Xuan, X.; Yao, W.; Huang, G.; Jin, J. Anti-profibrotic effects of artesunate on bleomycin-induced pulmonary fibrosis in Sprague Dawley rats. Mol. Med. Rep. 2012, 12, 1291–1297. [Google Scholar] [CrossRef] [Green Version]
- Roberfroid, M.B. Inulin-Type Fructans: Functional Food Ingredients. J. Nutr. 2007, 137, 2493S–2502S. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, M.; Lodi, F.; Vera, R.; Villar, I.C.; Cogolludo, A.; Jimenez, R.; Moreno, L.; Romero, M.; Tamargo, J.; Perez-Vizcaino, F.; et al. Quercetin and Isorhamnetin Prevent Endothelial Dysfunction, Superoxide Production, and Overexpression of p47phox Induced by Angiotensin II in Rat Aorta. J. Nutr. 2007, 137, 910–915. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Li, J.; Luo, C.; Liu, H.; Xu, W.; Chen, G.; Liew, O.W.; Zhu, W.; Puah, C.M.; Shen, X.; et al. Binding interaction of quercetin-3-β-galactoside and its synthetic derivatives with SARS-CoV 3CLpro: Structure–activity relationship studies reveal salient pharmacophore features. Bioorg. Med. Chem. 2006, 14, 8295–8306. [Google Scholar] [CrossRef]
- Okhuaroboa, A.; Falodun, J.E.; Erharuyicd, O.; Imiejec, V.; Faloduncde, A.; Langerc, P. Harnessing the medicinal properties of Andrographis paniculata for diseases and beyond: A review of its phytochemistry and pharmacology. Asian Pac. J. Trop. Dis. 2014, 4, 213–222. [Google Scholar] [CrossRef]
- Yarnell, E. Herbs for Viral Respiratory Infections. Altern. Complement. Ther. 2018, 24, 35–43. [Google Scholar] [CrossRef]
- Arora, R.; Chawla, R.; Marwah, R.; Arora, P.; Sharma, R.K.; Kaushik, V.; Goel, R.; Kaur, A.; Silambarasan, M.; Tripathi, R.P.; et al. Potential of Complementary and Alternative Medicine in Preventive Management of Novel H1N1 Flu (Swine Flu) Pandemic: Thwarting Potential Disasters in the Bud. Evid. Based Complement. Altern. Med. 2010, 2011, 586506. [Google Scholar] [CrossRef]
- Coon, J.T.; Ernst, E. Andrographis paniculata in the treatment and prevention of upper respiratory tract infection (URTI): A systematic review of safety and efficacy. Focus Altern. Complement. Ther. 2010, 8, 160–161. [Google Scholar] [CrossRef]
- Liu, Y.-T.; Chen, H.-W.; Lii, C.-K.; Jhuang, J.-H.; Huang, C.-S.; Li, M.-L.; Yao, H.-T. A Diterpenoid, 14-Deoxy-11, 12-Didehydroandrographolide, in Andrographis paniculata Reduces Steatohepatitis and Liver Injury in Mice Fed a High-Fat and High-Cholesterol Diet. Nutrients 2020, 12, 523. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Xiao, X.; Wei, X.; Li, J.; Yang, J.; Tan, H.; Zhu, J.; Zhang, Q.; Wu, J.; Liu, L. Composition and divergence of coronavirus spike proteins and host ACE2 receptors predict potential intermediate hosts of SARS-CoV-2. J. Med Virol. 2020, 92, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Mpiana, P.T.; Ngbolua, K.-T.-N.; Tshibangu, D.S.T.; Kilembe, J.T.; Gbolo, B.Z.; Mwanangombo, D.T.; Inkoto, C.L.; Lengbiye, E.M.; Mbadiko, C.M.; Matondo, A.; et al. Aloe vera (L.) Burm. F. as a Potential Anti-COVID-19 Plant: A Mini-review of Its Antiviral Activity. Eur. J. Med. Plants 2020, 86–93. [Google Scholar] [CrossRef]
- Velthuis, A.J.W.T.; Worm, S.H.E.V.D.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; Van Hemert, M.J. Zn2+ Inhibits Coronavirus and Arterivirus RNA Polymerase Activity In Vitro and Zinc Ionophores Block the Replication of These Viruses in Cell Culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef]
- Mohammadi, E.; Golchin, M. High protection of mice against Brucella abortus by oral immunization with recombinant probiotic Lactobacillus casei vector vaccine, expressing the outer membrane protein OMP19 of Brucella species. Comp. Immunol. Microbiol. Infect. Dis. 2020, 70, 101470. [Google Scholar] [CrossRef]
- Chang, H.-C.; Hung, W.-C.; Huang, M.-S.; Hsu, H.-K. Extract from the Leaves of Toona sinensis Roemor Exerts Potent Antiproliferative Effect on Human Lung Cancer Cells. Am. J. Chin. Med. 2002, 30, 307–314. [Google Scholar] [CrossRef]
- Yang, H.-L.; Chang, W.-H.; Chia, Y.-C.; Huang, C.-J.; Lu, F.-J.; Hsu, H.-K.; Hseu, Y.-C. Toona sinensis extracts induces apoptosis via reactive oxygen species in human premyelocytic leukemia cells. Food Chem. Toxicol. 2006, 44, 1978–1988. [Google Scholar] [CrossRef]
- Wang, P.-H.; Tsai, M.-J.; Hsu, C.-Y.; Wang, C.-Y.; Hsu, H.-K.; Weng, C.-F. Toona sinensis Roem (Meliaceae) leaf extract alleviates hyperglycemia via altering adipose glucose transporter 4. Food Chem. Toxicol. 2008, 46, 2554–2560. [Google Scholar] [CrossRef]
- Chen, C.-J.; Michaelis, M.; Hsu, H.-K.; Tsai, C.-C.; Yang, K.D.; Wu, Y.-C.; Cinatl, J.; Doerr, H.W. Toona sinensis Roem tender leaf extract inhibits SARS coronavirus replication. J. Ethnopharmacol. 2008, 120, 108–111. [Google Scholar] [CrossRef]
- Qin, G.-W.; Xu, R.-S. Recent advances on bioactive natural products from Chinese medicinal plants. Med. Res. Rev. 1998, 18, 375–382. [Google Scholar] [CrossRef]
- Wu, X.; Qin, G.; Cheung, K.K.; Cheng, K.F. New alkaloids from Isatis indigotica. Tetrahedron 1997, 53, 13323–13328. [Google Scholar] [CrossRef]
- McGovern, S.L.; Shoichet, B.K. Information Decay in Molecular Docking Screens against Holo, Apo, and Modeled Conformations of Enzymes. J. Med. Chem. 2003, 46, 2895–2907. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.O.; Weber, N.D.; Wood, S.G.; Hughes, B.G.; Murray, B.K.; North, J.A. In vitro virucidal activity of selected anthraquinones and anthraquinone derivatives. Antivir. Res. 1991, 16, 185–196. [Google Scholar] [CrossRef]
- Mak, N.-K.; Leung, C.-Y.; Wei, X.-Y.; Shen, X.-L.; Wong, R.N.-S.; Leung, K.-N.; Fung, M.-C. Inhibition of RANTES expression by indirubin in influenza virus-infected human bronchial epithelial cells. Biochem. Pharmacol. 2004, 67, 167–174. [Google Scholar] [CrossRef]
- Roy, S.; Bhattacharyya, P. Possible role of traditional medicinal plant Neem (Azadirachta indica) for the management of COVID-19 infection. Int. J. Res. Pharm. Sci. 2020, 11, 122–125. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Sujarwo, W.; Keim, A.P.; Caneva, G.; Toniolo, C.; Nicoletti, M. Ethnobotanical uses of neem (Azadirachta indica A. Juss.; Meliaceae) leaves in Bali (Indonesia) and the Indian subcontinent in relation with historical background and phytochemical properties. J. Ethnopharmacol. 2016, 189, 186–193. [Google Scholar] [CrossRef]
- Thakurta, P.; Bhowmik, P.; Mukherjee, S.; Hajra, T.K.; Patra, A.; Bag, P.K. Antibacterial, antisecretory and antihemorrhagic activity of Azadirachta indica used to treat cholera and diarrhea in India. J. Ethnopharmacol. 2007, 111, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, U.; Biswas, K.; Chatterjee, R.; Bandyopadhyay, D.; Chattopadhyay, I.; Ganguly, C.K.; Chakraborty, T.; Bhattacharya, K.; Banerjee, R.K. Gastroprotective effect of Neem (Azadirachta indica) bark extract: Possible involvement of H+-K+-ATPase inhibition and scavenging of hydroxyl radical. Life Sci. 2002, 71, 2845–2865. [Google Scholar] [CrossRef]
- Alam, G.; Wahyuono, S.; Ganjar, I.G.; Hakim, L.; Timmerman, H.; Verpoorte, R. Tracheospasmolytic Activity of Viteosin-A and Vitexicarpin Isolated fromVitex trifolia. Planta Med. 2002, 68, 1047–1049. [Google Scholar] [CrossRef] [PubMed]
- Vellingiri, B.; Jayaramayya, K.; Iyer, M.; Narayanasamy, A.; Govindasamy, V.; Giridharan, B.; Ganesan, S.; Venugopal, A.; Venkatesan, D.; Ganesan, H.; et al. COVID-19: A promising cure for the global panic. Sci. Total. Environ. 2020, 725, 138277. [Google Scholar] [CrossRef]
- Dashputre, N.L.; Naikwade, N.S. Immunomodulatory activity of Abutilon indicum linn on albino mice. Int J. Pharm Sci Res. 2010, 1, 178–184. [Google Scholar]
- Grover, A.; Agrawal, V.; Shandilya, A.; Bisaria, V.S.; Sundar, D. Non-nucleosidic inhibition of Herpes simplex virus DNA polymerase: Mechanistic insights into the anti-herpetic mode of action of herbal drug withaferin A. BMC Bioinform. 2011, 12, S22. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Mohan, C.; Dinakar, S.; Anand, T.; Elayaraja, R.; SathiyaPriya, B. Phytochemical, GC-MS analysis and Antibacterial activity of a Medicinal Plant Acalypha indica. Int. J. Pharm. Tech. Res. 2012, 4, 1050–1054. [Google Scholar]
- Dhaarani, V.; Sarvalingam, A.; Rajendran, A. Medicinal uses of psammophytic plants in Tranquebar regions of Tamil Nadu, India. J. Herbs Spices Med. Plants 2018, 24, 282–292. [Google Scholar] [CrossRef]
- Premila, M.; Conboy, L. Ayurvedic Herbs: A Clinical Guide to the Healing Plants of Traditional Indian Medicine. J. Altern. Complement. Med. 2007, 13, 841–842. [Google Scholar] [CrossRef]
- Vaghasiya, Y.; Chanda, S. Screening of some traditionally used Indian plants for antibacterial activity against Klebsiella pneumonia. J. Herb Med. Toxicol. 2009, 3, 161–164. [Google Scholar]
- Kundu, S.; Roy, S.; Lyndem, L.M. Cassia alata L.: Potential role as anthelmintic agent against Hymenolepis diminuta. Parasitol. Res. 2012, 111, 1187–1192. [Google Scholar] [CrossRef]
- Singh, R.P.; Jain, D.A. Evaluation of antimicrobial activity of alcoholic and aqueous extracts of five plants used in traditional medicine in North India. Int. J. Pharm. Tech. Res. 2011, 3, 376–380. [Google Scholar]
- Nayak, V.; Jadhav, V.; Sajjanshetty, M.R. Traditional medicine in the management of Recurrent Tonsillitis—An Ayurvedic Perspective. J. Ayurveda Integr. Med Sci. 2017, 2, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Harwansh, R.K.; Pareta, S.K.; Patra, K.C.; Rahman, M.A. Preliminary phytochemical screening and anthelmintic activity of Chloroxylon swietenia root extract. Int. J. Phytomed. 2010, 2, 255–259. [Google Scholar]
- Jacob, L.; Latha, M.S. In vitro antioxidant activity of Clitoria ternatea Linn. Int. J. Res. Phytochem. Pharmacol. 2013, 3, 35–39. [Google Scholar]
- Dharmananda, S. Tibetan herbal medicine. Int. J. Orient. Med. 2011, 26, 1–13. [Google Scholar]
- Boothapandi, M.; Ramanibai, R. Immunomodulatory activity of Indigofera tinctoria leaf extract on in vitro macrophage responses and lymphocyte proliferation. Int. J. Pharm. Pharm. Sci. 2016, 8, 58–63. [Google Scholar]
- Afzal, U.; Gulfraz, M.; Hussain, S.; Malik, F.; Maqsood, S.; Shah, I.; Mahmood, S. Hepatoprotective effects of Justicia adhatoda L. against carbon tetrachloride (CCl4) induced liver injury in Swiss albino mice. Afr. J. Pharm. Pharmacol. 2013, 7, 8–14. [Google Scholar] [CrossRef] [Green Version]
- John, K.M.; Enkhtaivan, G.; Ayyanar, M.; Jin, K.; Yeon, J.B.; Kim, D.H. Screening of ethnic medicinal plants of South India against influenza (H1N1) and their antioxidant activity. Saudi J. Biol. Sci. 2015, 22, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Katoch, D.; Singh, B.; Arora, S. Seclusion of vasicine-and quinazoline alkaloid from bioactive fraction of Justicia adhathoda and its antioxidant, antimutagenic and anticancerous activities. J. Glob. Biosci. 2016, 5, 3836–3850. [Google Scholar]
- Ravindran, R.; Juliet, S.; Sunil, A.; Kumar, K.A.; Nair, S.N.; Amithamol, K.; Shynu, M.; Rawat, A.K.S.; Ghosh, S. Eclosion blocking effect of ethanolic extract of Leucas aspera (Lamiaceae) on Rhipicephalus (Boophilus) annulatus. Vet. Parasitol. 2011, 179, 287–290. [Google Scholar] [CrossRef]
- Thenmozhi, M.; Dhanalakshmi, M.; Devi, K.M.; Sushila, K.; Thenmozhi, S. Evaluation of hepatoprotective activity of Leucas aspera hydroalcoholic leaf extract during exposure to lead acetate in male albino Wistar rats. Asian J. Pharm Clin. Res. 2013, 6, 78–81. [Google Scholar]
- Kalita, N.; Kalita, M.C. Ethnomedicinal plants of Assam, India as an alternative source of future medicine for treatment of Pneumonia. Int. Res. J. 2014, 3, 76–82. [Google Scholar]
- Sureshkumar, S.; Mishra, S. Hepatoprotective effect of extracts from Pergularia daemia Forsk. J. Ethnopharmacol. 2006, 107, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Naika, R.; Prasanna, K.P.; Ganapathy, P.S. Antibacterial activity of piperlongumine an alkaloid isolated from methanolic root extract of Piper Longum, L. Pharmacophore 2010, 1, 141–148. [Google Scholar]
- Vadakkan, K.; Hemapriya, J.; Selvaraj, V. Quorum quenching intervened in vivo attenuation and immunological clearance enhancement by Solanum torvum root extract against Pseudomonas aeruginosa instigated pneumonia in Sprague Dawley rats. Clin. Phytosci. 2019, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Rahman, M.K.; Abd El-Megeid, A.A. Hepatoprotective effect of soapworts (Saponaria officinalis), pomegranate peel (Punica granatum L) and cloves (Syzygium aromaticum linn) on mice with CCl4 hepatic intoxication. World J. Chem. 2006, 1, 41–46. [Google Scholar]
- Mishra, A.; Kumar, S.; Pandey, A.K. Scientific Validation of the Medicinal Efficacy of Tinospora cordifolia. Sci. World J. 2013, 2013, 292934. [Google Scholar] [CrossRef] [Green Version]
- Tharakan, B.; Dhanasekaran, M.; Manyam, B.V. Antioxidant and DNA protecting properties of anti-fatigue herb Trichopus zeylanicus. Phytotherapy Res. 2005, 19, 669–673. [Google Scholar] [CrossRef] [PubMed]
- More, D.; Shetty, V.; Shah, A.; Mungekar, S.; Kamat, S.K.; Rege, N.N. Effects of polyherbal ayurvedic formulations on non-specific immunity: An experimental study. Indian J. Pharmacol. 2008, 40, S160–S171. [Google Scholar]
- Koh, E.M.; Kim, H.J.; Kim, S.; Choi, W.H.; Choi, Y.H.; Ryu, S.Y.; Kim, Y.S.; Koh, W.S.; Park, S.-Y. Modulation of Macrophage Functions by Compounds Isolated from Zingiber officinale. Planta Med. 2008, 75, 148–151. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.S.; Wang, K.C.; Yeh, C.F.; Shieh, D.E.; Chiang, L.C. Fresh ginger (Zingiber officinale) has anti-viral activity against human respiratory syncytial virus in human respiratory tract cell lines. J. Ethnopharmacol. 2013, 145, 146–151. [Google Scholar] [CrossRef]
- Akoachere, J.-F.T.K.; Ndip, R.N.; Chenwi, E.B.; Ndip, L.M.; Njock, T.E.; Anong, D.N. Antibacterial effect of Zingiber officinale and Garcinia kola on respiratory tract pathogens. East Afr. Med. J. 2002, 79, 588–592. [Google Scholar] [CrossRef] [Green Version]
- Ali, T.; Shahbaz, B.; Suleri, A. Analysis of myths and realities of deforestation in Northwest Pakistan: Implications for forestry extension. Int. J. Agric. Biol. 2006, 8, 107–110. [Google Scholar]
- Abbasi, A.M.; Iqbal, J.; Khan, M.A.; Shah, M.H. Health risk assessment and multivariate apportionment of trace metals in wild leafy vegetables from Lesser Himalayas, Pakistan. Ecotoxicol. Environ. Saf. 2013, 92, 237–244. [Google Scholar] [CrossRef]
- Singh, G.D.; Kaiser, P.; Youssouf, M.S.; Singh, S.; Khajuria, A.; Koul, A.; Bani, S.; Kapahi, B.K.; Satti, N.K.; Suri, K.A.; et al. Inhibition of early and late phase allergic reactions by Euphorbia hirta L. Phytotherapy Res. 2006, 20, 316–321. [Google Scholar] [CrossRef]
- Ko, F.-N.; Huang, T.-F.; Teng, C.-M. Vasodilatory action mechanisms of apigenin isolated from Apium graveolens in rat thoracic aorta. Biochim. Biophys. Acta Gen. Subj. 1991, 1115, 69–74. [Google Scholar] [CrossRef]
- Khare, C.P. Indian Medicinal Plants: An Illustrated Dictionary; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Walter, T.M. Review of Acalypha indica Linn. in Traditional Siddha Medicine. 2007. Available online: https://www.academia.edu/1410079/Review_of_Review_of_Acalypha_indica_Linn_in_Traditional_Siddha_Acalypha_indica_Linn_in_Traditional_Siddha_Medicine (accessed on 5 March 2021).
- Hosseinzadeh, H.; Ghenaati, J. Evaluation of the antitussive effect of stigma and petals of saffron (Crocus sativus) and its components, safranal and crocin in guinea pigs. Fitoterapia 2006, 77, 446–448. [Google Scholar] [CrossRef]
- Joshi, V.K.; Joshi, A.; Dhiman, K.S. The Ayurvedic Pharmacopoeia of India, development and perspectives. J. Ethnopharmacol. 2017, 197, 32–38. [Google Scholar] [CrossRef]
- Hirata, N.; Naruto, S.; Inaba, K.; Itoh, K.; Tokunaga, M.; Iinuma, M.; Matsuda, H. Histamine Release Inhibitory Activity of Piper nigrum Leaf. Biol. Pharm. Bull. 2008, 31, 1973–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naseri, M.K.; Yahyavi, H. Antispasmodic effect of Piper nigrum fruit hot water extract on rat ileum. Pak. J. Biol. Sci 2008, 11, 1492–1496. [Google Scholar] [PubMed]
- Bharali, R.; Azad, M.R.H.; Tabassum, J. Chemopreventive Action of Boerhaavia Diffusa on DMDA-induced Skin Carcinogenesis in Mice. Indian J. Physiol. Pharmacol. 2003, 47, 459–464. [Google Scholar] [PubMed]
- Simin, K.; Khaliq-Uz-Zaman, S.M.; Ahmad, V.U. Antimicrobial activity of seed extracts and bondenolide from Caesalpinia bonduc (L.) Roxb. Phytother. Res. 2001, 15, 437–440. [Google Scholar] [CrossRef]
- Shinwari, Z.K. Medicinal plants research in Pakistan. J. Med. Plant. Res. 2010, 4, 161–176. [Google Scholar] [CrossRef]
- Thwe, A.; Arasu, M.V.; Li, X.; Park, C.H.; Kim, S.J.; Al-Dhabi, N.A.; Park, S.U. Effect of Different Agrobacterium rhizogenes Strains on Hairy Root Induction and Phenylpropanoid Biosynthesis in Tartary Buckwheat (Fagopyrum tataricum Gaertn). Front. Microbiol. 2016, 7, 318. [Google Scholar] [CrossRef]
- Shanmugavelu, M. Noikaluku Siddha parikaram. Part I; Indian Medicine-Homeopathy Department, Government of India: New Delhi, India, 2004.
- Moineddin, R.; Nie, J.X.; Domb, G.; Leong, A.M.; Upshur, R.E.G. Seasonality of primary care utilization for respiratory diseases in Ontario: A time-series analysis. BMC Health Serv. Res. 2008, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.K.; Kumar, A.; Misra, N. On the Inhibition of COVID-19 Protease by Indian Herbal Plants: An in Silico Investigation. arXiv 2020, arXiv:2004.03411 2020. [Google Scholar]
- Alsaffar, D.F. In Silico Molecular Docking Studies of Medicinal Arabic Plant-Based Bioactive Compounds as a Promising Drug Candidate against COVID-19. Int. J. Innov. Sci. Res. Technol. 2020, 5, 876–896. [Google Scholar]
- Gautret, P.; Lagier, J.-C.; Parola, P.; Hoang, V.T.; Meddeb, L.; Sevestre, J.; Mailhe, M.; Doudier, B.; Aubry, C.; Amrane, S.; et al. Clinical and microbiological effect of a combination of hydroxychloroquine and azithromycin in 80 COVID-19 patients with at least a six-day follow up: A pilot observational study. Travel Med. Infect. Dis. 2020, 34, 101663. [Google Scholar] [CrossRef]
- Juurlink, D.N. Safety considerations with chloroquine, hydroxychloroquine and azithromycin in the management of SARS-CoV-2 infection. Can. Med. Assoc. J. 2020, 192, E450–E453. [Google Scholar] [CrossRef] [Green Version]
- Gurwitz, D. Angiotensin receptor blockers as tentative SARS-CoV-2 therapeutics. Drug Dev. Res. 2020, 81, 537–540. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Wang, Z.; Zhao, F.; Yang, Y.; Li, J.; Yuan, J.; Wang, F.; Li, D.; Yang, M.; Xing, L.; et al. Treatment of 5 Critically Ill Patients With COVID-19 With Convalescent Plasma. JAMA 2020, 323, 1582. [Google Scholar] [CrossRef]
- Dhama, K.; Sharun, K.; Tiwari, R.; Dadar, M.; Malik, Y.S.; Singh, K.P.; Chaicumpa, W. COVID-19, an emerging coronavirus infection: Advances and prospects in designing and developing vaccines, immunotherapeutics, and therapeutics. Hum. Vaccines Immunother. 2020, 16, 1232–1238. [Google Scholar] [CrossRef] [Green Version]
- Uddin, M.; Mustafa, F.; Rizvi, T.A.; Loney, T.; Al Suwaidi, H.; Al-Marzouqi, A.H.H.; Eldin, A.K.; Alsabeeha, N.; Adrian, T.E.; Stefanini, C.; et al. SARS-CoV-2/COVID-19: Viral Genomics, Epidemiology, Vaccines, and Therapeutic Interventions. Viruses 2020, 12, 526. [Google Scholar] [CrossRef]
- Sanders, J.M.; Monogue, M.L.; Jodlowski, T.Z.; Cutrell, J.B. Pharmacologic Treatments for Coronavirus Disease 2019 (COVID-19). JAMA 2020, 323, 1824–1836. [Google Scholar] [CrossRef]
- Menachery, V.D.; Gralinski, L.E.; Mitchell, H.D.; Dinnon, K.H.; Leist, S.R.; Yount, B.L.; McAnarney, E.T.; Graham, R.L.; Waters, K.M.; Baric, R.S. Combination Attenuation Offers Strategy for Live Attenuated Coronavirus Vaccines. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Buyel, J. Molecular farming—The slope of enlightenment. Biotechnol. Adv. 2020, 40, 107519. [Google Scholar] [CrossRef]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Magnifection?a new platform for expressing recombinant vaccines in plants. Vaccine 2005, 23, 2042–2048. [Google Scholar] [CrossRef]
- A Critical Review of the Concept of Transgenic Plants: Insights into Pharmaceutical Biotechnology and Molecular Farming. Curr. Issues Mol. Biol. 2016, 18, 21–42. [CrossRef] [Green Version]
- Rosales-Mendoza, S. Will plant-made biopharmaceuticals play a role in the fight against COVID-19? Expert Opin. Biol. Ther. 2020, 20, 545–548. [Google Scholar] [CrossRef] [Green Version]
- Malik, S. Production of Plant Derived Natural Compounds through Hairy Root Culture; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Grzegorczyk, I.; Królicka, A.; Wysokińska, H. Establishment of Salvia officinalis L. hairy root cultures for the production of rosmarinic acid. Z. Naturforschung C 2006, 61, 351–356. [Google Scholar] [CrossRef]
- Tavassoli, P.; Afshar, A.S. Influence of different Agrobacterium rhizogenes strains on hairy root induction and analysis of phenolic and flavonoid compounds in marshmallow (Althaea officinalis L.). 3 Biotech 2018, 8, 351. [Google Scholar] [CrossRef]
- Park, C.H.; Zhao, S.; Yeo, H.J.; Park, Y.E.; Baska, T.B.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Comparison of Different Strains of Agrobacterium rhizogenes for Hairy Root Induction and Betulin and Betulinic Acid Production in Morus alba. Nat. Prod. Commun. 2017, 12, 1934578X1701200403. [Google Scholar] [CrossRef] [Green Version]
- Yousefian, S.; Lohrasebi, T.; Farhadpour, M.; Haghbeen, K. Production of phenolic acids in hairy root cultures of medicinal plant Mentha spicata L. in response to elicitors. Mol. Biol. Res. Commun. 2020, 9, 23–34. [Google Scholar]
- Gabr, A.; Sytar, O.; Ahmed, A.R.; Smetanska, I. Production of phenolic acid and antioxidant activity in transformed hairy root cultures of common buckwheat (Fagopyrum Esculentum M.). Aust. J. Basic Appl. Sci. 2012, 6, 577–586. [Google Scholar]
- Zhou, Y.; Hirotani, M.; Yoshikawa, T.; Furuya, T. Flavonoids and phenylethanoids from hairy root cultures of Scutellaria baicalensis. Phytochemistry 1997, 44, 83–87. [Google Scholar] [CrossRef]
- Skala, E.K.A.; Olszewska, M.A.; Kiss, A.K.; WysokiNska, H. Establishment of Hairy Root Cultures of Rhaponticum carthamoides (Willd.) Iljin for the Production of Biomass and Caffeic Acid Derivatives. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, M.; Anbumegala, M.; Surendran, R.; Arun, M.; Shanmugam, G. Elite hairy roots of Raphanus sativus (L.) as a source of antioxidants and flavonoids. 3 Biotech 2018, 8, 128. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Praveen, N.; Kim, E.-H.; Kim, S.-H.; Chung, I.-M. Production of anthraquinones, phenolic compounds and biological activities from hairy root cultures of Polygonum multiflorum Thunb. Protoplasma 2014, 251, 555–566. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Bahramnejad, B.; Baneh, H.D.; Emamifar, A.; Goodwin, P.H. Hairy root culture optimization and resveratrol production from Vitis vinifera subsp. sylvesteris. World J. Microbiol. Biotechnol. 2017, 33, 67. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-W.; Kim, Y.S.; Uddin, R.; Kwon, D.Y.; Kim, Y.B.; Lee, M.Y.; Kim, S.-J.; Park, S.U. Resveratrol Production from Hairy Root Cultures of Scutellaria baicalensis. Nat. Prod. Commun. 2013, 8, 1934578x1300800517. [Google Scholar] [CrossRef] [Green Version]
- Ludwig-Müller, J.; Jahn, L.; Lippert, A.; Püschel, J.; Walter, A. Improvement of hairy root cultures and plants by changing biosynthetic pathways leading to pharmaceutical metabolites: Strategies and applications. Biotechnol. Adv. 2014, 32, 1168–1179. [Google Scholar] [CrossRef]
- Halder, M.; Sarkar, S.; Jha, S. Elicitation: A biotechnological tool for enhanced production of secondary metabolites in hairy root cultures. Eng. Life Sci. 2019, 19, 880–895. [Google Scholar] [CrossRef] [Green Version]
- Vasconsuelo, A.; Boland, R. Molecular aspects of the early stages of elicitation of secondary metabolites in plants. Plant Sci. 2007, 172, 861–875. [Google Scholar] [CrossRef]
- Putalun, W. Technology of compact MAb and its application for medicinal plant breeding named as missile type molecular breeding. Curr. Drug Discov. Technol. 2011, 8, 24–31. [Google Scholar] [CrossRef]
- Hoseinpanahi, B.; Bahramnejad, B.; Majdi, M.; Dastan, D.; Ashengroph, M. The effect of different elicitors on hairy root biomass and resveratrol production in wild Vitis vinifera. Appl. Biotechnol. Rep. 2020, 7, 25–31. [Google Scholar] [CrossRef]





| Plant Species or Plant Organ | Common Name | Active Against | Mode of Action | Compound(s) Isolated/Target | Reference |
|---|---|---|---|---|---|
| Torreya nucifera | Coniferous tree | SARS-CoV | 3CLpro inhibitor | Plant extract | [55] |
| Thuja orientalis | Oriental Arborvitae | SARS-CoV | Viral growth inhibitor | Plant extract | [35] |
| Angelica keiskei | Tomorrow’s leaf | SARS-CoV | 3CLpro inhibitor | Chalcones | [56] |
| Radix glycyrrhizae | Licorice root | SARS-CoV | Inhibits viral replication | Glycyrrhizin | [57] |
| Dioscoreae Rhizoma | Yam Rhizome | SARS-CoV | Viral growth inhibitor | Plant extract | [58] |
| Psoralea corylifolia | Babchi | SARS-CoV | PLpro inhibitor | Plant extract | [59] |
| Allium sativum | Garlic | HCMV | Inhibits viral replication in earlier stages of viral cycle before viral DNA synthesis | Allitridin | [60] |
| Myrica faya | Fire tree | SARS-CoV | Helicase inhibitor | Myricetin | [61] |
| Laggera pterodonta | Curly Blumea | EV71 | Inhibits viral RNA replication | Chrysosplenetin and penduletin | [62] |
| Cassiae semen | n/a | SARS-CoV | Viral growth inhibitor | Plant extract | [58] |
| Cryptomeria japonica | Sugi | SARS-CoV | Viral growth inhibitor | Hydroxy-deoxycryptojaponol | [63] |
| Triterygium regelii | Regel’s threewingnut | SARS-CoV | 3CLpro inhibitor | Celastrol Pristimerin Tingenone Iguesterin | [55] |
| Gentianae Radix | Gentian Root | SARS-CoV | Viral growth inhibitor | Plant extract | [58] |
| Pterocarpus santalinus | Red sandalwood | SARS-CoV | 3CLpro inhibitor | Savinin | [63] |
| Betula pubescens | Downy birch | SARS-CoV | 3CLpro inhibitor | Betulinic acid | [63] |
| Fructus arctii | Burdock | Flu | Inhibits viral replication | Arctigenin | [64] |
| Galla chinensis | Nutgall tree | SARS-CoV | Viral spike protein and Human ACE2 receptors inhibitor | Tetra-O-galloyl- β-d-glucose | [65] |
| Saxifraga melanocentra Rhodiola kirilowii | Saxifrages Golden root | HCV | Inhibits viral NS3 serine protease | Polyphenolic compounds (−)-Epicatechin-3-O-gallate, 3,30-digalloylproprodelphinidin B2, 3,30-digalloylprocyanidin B2, and (−)-epigallocatechin-3-O-gallate | [66,67] |
| Paulownia tomentosa | Princess tree | SARS-CoV | PLpro inhibitor | Flavonoids | [67] |
| Radix scutellariae Salvia miltiorrhiza Ranunculus sieboldii Ranunculus sceleratus Radix sophorae Radix bupleuri | Huang Qin Red sage Buttercup Celery-leaved buttercup Shrubby sophora Thorowax Root | HBV | Inhibits viral DNA polymerase Inhibits viral replication Inhibits viral replication Inhibits viral replication Inhibits viral DNA replication and HBeAg production | Wogonin Protocatechuic aldehyde/Tanshinones Apigenin 40-O-a-rhamnopyranoside, apigenin 7-O-b-glucopyranosyl-40-O-a-rhamnopyranoside, tricin 7-O-b-glucopyranoside, tricin, isoscopoletin Apigenin 4′-O-α-rhamnopyranoside, apigenin 7-O-β-glucopyranosyl-4′-O-a-rhamnopyranoside, tricin 7-O-β-glucopyranoside, tricin, isoscopoletin Saikosaponin C | [16,68,69,70] |
| Scutellaria lateriflora | Blue skullcap | SARS-CoV | Helicase inhibitor | Scutellarein | [71] |
| Loranthi ramus | Mulberry | SARS-CoV | Viral growth inhibitor | Plant extract | [58] |
| Stephania cepharantha | n/a | SARS-CoV | Viral growth inhibitor | Biscoclaurine | [35] |
| Cinnamomum cassia | Chinese cassia | SARS-CoV | 3CLpro inhibitor | Plant extract | [71] |
| Linum usitatissimum | Flax | SARS-CoV | 3CLpro inhibitor | Herbacetin | [55] |
| Alnus japonica | East Asian alder | SARS-CoV | PLpro inhibitor | Plant extract | [72] |
| Laurus nobilis | Bay laurel. | SARS-CoV | Viral growth inhibitor | Plant Extract | [35] |
| Rhizoma coptidis Chrysanthemum Morifolium Vatica cinerea Aesculus chinensis Kadsura matsudai | Huang Lian Florist’s daisy Resak Laut Chinese horse chestnut Kadsura | HIV/SARS-CoV | Inhibits viral DNA synthesis/3CLpro inhibitor Inhibits viral integrase Inhibits viral replication Inhibits viral protease Inhibits viral replication | Berberine Apigenin-7-O-b-D-g-lucopyranoside Vaticinone (23E)-27-nor-3-hydroxycycloart-23-en-25-one Triterpenoid saponins Schizanrin B, C, D, and E | [73,74,75,76,77] |
| Chamaecyparis obtuse Euphorbia jolkini Limonium sinense Ranunculus sieboldii Ranunculus sceleratus Limonium sinense | Hinoki cypress Spurge Girard Buttercup Celery-leaved buttercup Girard | HSV | Inhibits HSV-1 ICP0, ICP4 expression, and as viral DNA synthesis Affects the late stage of HSV-2 Replication Inhibits viral replication Inhibits viral replication Inhibits viral replication | Yatein Putranjivain A Samarangenin B Protocatechuyl aldehyde Isodihydrosyringetin, (−)-epigallocatechin 3-O-gallate, samarangenin B, myricetin, myricetin 3-α-arhamnopyranoside, quercetin 3-O-α-rhamnopyranoside, (−)-epigallocatechin, gallic acid, N-trans-caffeoyltyramine, N-transferuloyltyramine | [16,78,79,80,81] |
| Azadirachta indica | Neem tree | Dengue virus | n/a | Leaf extract (Aqueous) inhibits DEN-2 in vivo | [82] |
| Moringa oleifera | Horseradish tree | HIV/Epstein-Barr virus (EBV) | n/a | Leaves used to inhibit viral replication/leaves and seeds inhibits activity against EBV activation | [83,84] |
| Terminalia bellerica | Myrobalan | HIV-1 Pseudo viruses | n/a | Plant extract against HIV-1 | [85] |
| Rheum palmatum | Chinese rhubarb | SARS-CoV | Viral spike protein and | Emodin | [86] |
| Avicennia marina | Grey mangrove | Hepatitis B virus | Inhibits HBV antigen | n/a | [87] |
| Litchi chinensis | litchi | SARS-CoV | 3CLpro inhibitor | Flavonoids extract | [52] |
| Multiflora Tuber | Tuber fleeceflower | SARS-CoV | Viral spike protein and Human ACE2 receptors inhibitor | Emodin | [86] |
| Canthium coromandelicum | Alston | HIV | - | Leaf extract controls HIV infection | [82] |
| Houttuynia cordata | Fish mint | SARS-CoV | 3CLpro inhibitor and RNA-dependent RNA polymerase (RdRp) inhibitor | Plant extract | [88] |
| Veronica linariifolia | Speedwell | SARS-CoV | Viral growth inhibitor | Luteolin | [62] |
| Carissa edulis | Conkerberry | Herpes simplex virus | Exhibits anti-HSV-1 and -2 properties in vitro and in vivo strongly | n/a | [89] |
| Nicotiana benthamiana | Benth | SARS-CoV | Viral growth inhibitor | NICTABA Lectin | [90] |
| Urtica dioica | Common nettle | SARS-CoV | Viral spike protein inhibitor | Urtica dioica agglutinin | [91] |
| Phyllanthus amarus | Indian gooseberry | Human immunodeficiency virus/hepatitis B virus | Inhibits HIV replication | Plant extract had lost HBV antigen surface | [92] |
| Guazuma ulmifolia Lam | West Indian elm | Polio virus | Extracts inhibits polio replications | n/a | [93] |
| Achyranthus aspera | Chaff-flower | Herpes simplex virus | Inhibits earlier stages of HSV multiplications | n/a | [94] |
| Camelliasinensis | Tea tree | SARS-CoV | 3CLpro inhibitor | Tannic acid/3-isotheaflavin-3-gallate | [95] |
| Sesbania grandiflora | Vegetable hummingbird | Herpes simplex virus | n/a | Extract possesses strong antiviral activity against HSV | [96] |
| Ficus religiosa | Bo tree | Human rhino virus (HRV) and Respiratory syncytial virus (RSV) | n/a | Bark extract endowed with antivirus activity against HRV and RSV | [97] |
| Hippophae rhamnoides | Dengue virus | Significant anti-dengue activity | Leaf extract | [98] | |
| Glycine max(black) | Soybean | Human adenovirus (type 1) | Inhibits human ADV-1 in dose-dependent manner | n/a | [99] |
| Acacia nilotica | Gum arabic tree | HIV-PR | Inhibition | n/a | [100] |
| Allium sativum | Garlic | SARS | Proteolytic and hemagglutinating activity and viral replication | n/a | [101] |
| Andrographispaniculata Boerhaavia diffusa | Green chireta Punarnava | SARS-COV and likely SARS-CoV-2 | Suppression Inhibition | NLRP3, capase-1, and IL-1β ACE | [31,102,103] |
| Clerodendrum inerme | The glory bower | SARS-CoV-2 | Inactivation | Ribosome | [104] |
| Clitoria ternatea | Butterfly pea | n/a | Metalloproteinase inhibitor | ADAM17 | [105] |
| Coriandrum sativum | Coriander | n/a | Inhibition | ACE | [106] |
| Cynara scolymus Cassia occidentalis | Scolymus | n/a | Inhibition | ACE | [102,103] |
| Embelia ribes | White-flowered Embelia | n/a | Inhibition | ACE | [102,103] |
| Eugenia jambolana | Black Plum | n/a | Inhibition | Protease | [49] |
| Euphorbia granulata | Asthma-plant | HIV-1 PR | Inhibition | - | [100] |
| Glycyrrhiza glabra Gymnema sylvestre | Licorice Gurmar | SARS; HIV-1 | Inhibition of viral replication; modulation of membrane fluidity Inhibition of viral DNA synthesis | Glycyrrhizin | [57,107] |
| Hyoscyamus niger | black henbane | n/a | Inhibition and Bronchodilator | Ca2+ | [108] |
| Ocimum kilimandscharicum | Camphor basil | HIV-1 | Inhibition | n/a | [51] |
| Ocimum sanctum | Holy basil | HIV-1 | Inhibition | n/a | [50] |
| Punica granatum | Pomegranate | Human herpes virus-3 | Inhibition | ACE/Phytochemical extract exhibits potential antiviral activity | [102,103] |
| Salacia oblonga | Oblong leaf salacia | n/a | Suppression | Angiotensin II and AT1 | [109] |
| Sambucus ebulus | Danewort | Enveloped virus | Inhibition | n/a | [110] |
| Solanum nigrum | European black nightshade | HIV-1 | n/a | n/a | [52] |
| Sphaeranthus indicus | East Indian globe | Mouse corona virus and Herpes virus | Inhibition | n/a | [111,112] |
| Strobilanthes callosa | Plietesials | HCoV-NL63 | Blocking | n/a | [107,113] |
| Strobilanthes cusia | Kuntze | HCoV-NL63 | Blocking | n/a | [113] |
| Vitex negundo | Five-leaved chaste tree | HIV-1 | Inhibition | n/a | [52,53,113,114] |
| Emblica officinalis | Emblic | Influenza A virus | Prevention of virus adsorption and suppression of virus release | Pentagalloyl glucose | [114] |
| Vitex trifolia | Simpleleaf Chastetree | SARS-COV | Reduction | n/a | [115] |
| Scintific Name | Common Names | Mode of Action | References |
|---|---|---|---|
| Tinospora cordifolia | Heart-leaved moonseed | Chronic fever | [257,258] |
| Andrograhis paniculata | Creat or green chireta | Fever and cold | |
| Cydonia oblonga | Quince | Antioxidant, immune-modulatory, anti-allergic, smooth muscle relaxant, and anti-influenza activity | |
| Zizyphus jujube | Jujube | ||
| Cordia myxa | lasura | ||
| Agastya haritaki | Agastya Rasayana | Upper respiratory infections | |
| Anu Thailam | - | Respiratory infections Fever | |
| Adathoda siddha | Malabar nut | Fever | |
| Bryonia alba | White bryony | Reduce lung inflammation | |
| Rhus toxicodendron | Eastern Poison Oak | Viral infections | |
| Atropa belladonna | Deadly nightshade | Asthma and chronic lung diseases | |
| Bignonia sempervirens | Yellow jessamine | Asthma | |
| Eupatorium perfoliatum | Agueweed | Respiratory symptoms | |
| Kabasura Kudineer | - | Fever, cough, sore throat, shortness of breath | |
| Abutilon indicum | Mallow | Immunomodulatory function | [107,259] |
| Withania somnifera | Ashwagandha | General tonic and to boost immunity/against herpes simplex virus | [260] |
| Hydrastis canadensis | Goldenseal root | Reduces plasma TNF-α, INF-γ, and NO levels; inhibits the T helper-type 2 cytokine profile. | [261] |
| Acalypha indica | Indian Acalypha | Anthelmintic | [262] |
| Achyranthes aspera | Prickly chaff flower | Anti-viral activity | [263] |
| Momordica charantia | bitter melon | Inhibits the release of TNF-α, NO and proliferation of spleen cells induced by PHA and Con A. | [261] |
| Adhatoda vasica | Adulsa | Anti-asthmatic, anti-allergic and anti-tussive activity | [264] |
| Nigella sativa | Black cumin | Reduces the pancreatic ductal adenocarcinoma cell (PDA) synthesis of monocyte chemoattractant protein-1 (MCP-1), TNF- α, IL-1β, and cyclooxygenase (COX)-2, as well as inhibiting the polymorphonuclear leukocyte functions. | [261] |
| Alangium salvifolium | Sage-leaved alangium | Anti-rheumatoid | [265] |
| Urtica dioica | Common Nettle | Reduction of TNF-α and other inflammatory cytokines | [261] |
| Cassia alata | Candle Bush | Anti-helmintic activity | [107,266] |
| Cassia fistula | Golden Shower | Antibacterial activity | [267] |
| Cayratia pedata | Birdfoot Grape | Anti-inflammatory activity | [268] |
| Chloroxylon swietenia | East Indian satinwood | Anti-helmintic activity | [269] |
| Clitoria ternatea linn. | Asian pigeonwings | Anti-oxidant activity | [270] |
| Eugenia singampattiana | Jungle Guava | Anti-inflammatory activity | [107,268] |
| Hippophae rhamnoides | Sea-buckthorn | Eliminating phlegm, stopping coughing, improving digestion, and treating lung diseases | [271] |
| Indigofera tinctoria | True Indigo | Immunomodulatory function | [107,272] |
| Justicia adhatoda | Malabar Nut | Anti-oxidant and anti-mutagenic activity; hepatoprotective activity | [1,236,273,274,275] |
| Leucas aspera | Thumbai | Hepatoprotective activity, acaricidal properties | [107,276,277] |
| Mucuna pruriens Xanthium strumanium | Monkey Tamarind Common Cocklebur | Antibacterial drugs against pneumonia | [278] |
| Pergularia daemia | Pergularia | Hepatoprotective effect | [107,279] |
| Piper longum | Long pepper | Anti-pneumonia drug | [280] |
| Salacia reticulata | Meharimula | Anti-inflammatory activity | [268] |
| Santalum album | White Sandalwood | Anti-inflammation of the lungs, blood, and pus in the sputum | [271] |
| Solanum tornum | Turkey Berry | Anti-pneumonia drug | [281] |
| Solanum suratens | Surattense Nightshade | Anti-viral activity | [263] |
| Solanum xanthocarpum | Yellow-fruit nightshade | Anti-asthmatic | [264,271] |
| Strychnos minor | Snakewood | Anti-inflammatory activity | [268] |
| Strychnosnux vomica | Strychnine tree | Anti-inflammatory activity | [268] |
| Syzygium aromaticum | Caryophyllus | Hepatoprotective properties | [282] |
| Tinospora cordifolia | Moonseed | Anti-pneumonia drug | [283] |
| Trichopus zeylanicus | Arogyapacha | Anti-oxidant and anti-fatigue activity | [284] |
| Tylophora indica | Indian Ipecac | Anti-asthmatic | [264] |
| Verbascum thapsus | Great mullein | Enhancing peroxidase, phenolics, and antioxidant activity | [1] |
| Vitex altissima | Peacock Chaste Tree | Acts against acute inflammation | [285] |
| Vitex trifolia | Hand-of-Mary | Tracheospasmolytic activity | [107,257] |
| Wrightia tinctoria | Pala Indigo | Anti-inflammatory activity | [268] |
| Yuthog’s Bamboo | Bamboo | Reduction of pain and anti-inflammation of the lungs and respiratory tract | [271] |
| Zingiber officinale | Canton Ginger | Modulation of macrophage functions, phagocytic properties, anti-viral activity, and bronchial infections | [1,286,287] |
| Garcinia Kola | Bitter Kola | Anti-bacterial drug against respiratory pathogens | [288] |
| Cymbopogon citratus | West Indian lemon grass | Anti-viral infections | [289] |
| Camellia sinensis | Tea plant | Anti-infective activity | [290] |
| Achillea mellefolium | Yarrow | Protects upper respiratory tract from viral infections | [291] |
| Apium graveolens | Celery | Anti-bacterial and anti-viral agent | [292] |
| Borassus flabellifer | Palmyra Palm | Protects from pulmonary infections; anti-bacterial and anti-viral activity | [293] |
| Caesalpinia bonduc | Grey Nicker | Treatment for asthma (anti-bacterial and anti-viral agent) | [294] |
| Calotropis gigantea | Crown flower | Anti-bacterial and anti-viral agent | [293] |
| Crocus sativus | Saffron Crocus | Treatment for asthma and cough | [295] |
| Euphorbia hirta | Asthma-Plant | Anti-bacterial and anti-viral agent | [291] |
| Piper nigrum | Black pepper | Anti-viral agent | [296] |
| Strychnos potatorum | Clearing-Nut Tree | Treatment for bronchitis | [297] |
| Terminalia bellirica Roxb | Beleric Myrobalan | Effective for asthma | [298] |
| Tylophora indica Merrill | Antamul | Treatment for bronchitis and asthma | [299] |
| Tussilago farfara | Coltsfoot | Treatment for cough and asthma | [300] |
| Thymus linearis | Himalayan Wild Thyme | Anti-viral activity | [290] |
| Senecio chrysanthemoides | Senecio | Treatment for lung diseases | [301] |
| Portulaca oleracea | Duckweed | Anti-inflammatory and anti-asthma properties | [294] |
| Papaver somniferum | Opium Poppy | Treatment for respiratory diseases | [293] |
| Morus laevigata Wall. ex Brandis | White Mulberry | Treatment for cough | [302] |
| Ephedra gerardiana Wall. ex Stapf | Gerard’s Jointfir | Treatment for cough and asthma | [303] |
| Geranium wallichianum | Sylvia’s Surprise | Treatment for cough | [292] |
| Micromeria biflora | Lemon Scented Thyme | Treatment for cough | [295] |
| Picrorhiza kurroa Royle ex. Benth | Kutki | Treatment for asthma and bronchitis | [289] |
| Primula denticulata Sm. | Drumstick Primula | Treatment for cough and bronchitis | [304] |
| Plant Parts | Scientific Name | The Major Phenolic Compound | Formula | Active Sites/Binding Residue/H-Bond Length (Å) |
|---|---|---|---|---|
| Leaves of harsingar | Nyctanthes arbortristis | Nictoflorin | C27H30O15 | N-H--O/GLY-143/2.311 |
| Astragalin | C21H20O11 | O--H/PHE-140/2.197 | ||
| Lupeol | C25H26O4 | N-H--O/THR-26/2.027 | ||
| Giloy | Tinospora cordifolia | Berberine Sitosterol | C28H18NO4 C29H50O | N-H--O/GLY-143/2.540 N-H--O/GLY-143/2.577 O--H/PHE-166/2.080 |
| Aloe vera leaves | Aloe barbadensis | Aloenin Aloesin | C19H22O10 C19H22O9 | O--H/PHE-140/2.151 N-H--O/GLY-143/2.016 N-H--O/GLU-166/2.297 |
| The dried ground rhizome of the turmeric | Curcuma longa | Curcumin | C21H20O6 | N-H--O/GLY-143/2.243 |
| The oil of neem | Azadirachta indica | Nimbin | C30H36O9 | N-H--O/GLY-143/2.161 |
| Steroidal constituents of ashwagandha | Withanolide Withaferin A | C28H38O6 C28H38O6 | O--H/GLU-166/1.991 N-H--O/GLU-166/2.110 N-H--O/GLY-143/2.577 | |
| Constituents of pungent ketones, which result in the strong aroma of ginger | Zingiber officinale | Gingerol Shogaol | C17H26O4 C17H24O3 | O--H/THR-190/2.026 N-H--O/GLY-143/2.289 N-H--O/THR-26/2.398 O--H/THR-24/2.345 |
| Red onion | Allium cepa | Quercetin | C15H10O7 | O--H/THR-26/1.936 |
| Tulsi leaves | Ocimum sanctum | Ursolic acid Apigenin | C30H48O3 C15H10O5 | N-H--O/GLY-143/2.330 O--H/THR-26/1.994 |
| Cannabis extracts | Cannabis sativa | Cannabidiol | C21H30O2 | N-H--O/GLY-143/2.325 |
| Isolated from the plants of both the black and white pepper grains | Piper nigrum | Piperine | C17H19NO3 | N-H--O/THR-26/2.529 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abiri, R.; Abdul-Hamid, H.; Sytar, O.; Abiri, R.; Bezerra de Almeida, E., Jr.; Sharma, S.K.; Bulgakov, V.P.; Arroo, R.R.J.; Malik, S. A Brief Overview of Potential Treatments for Viral Diseases Using Natural Plant Compounds: The Case of SARS-Cov. Molecules 2021, 26, 3868. https://doi.org/10.3390/molecules26133868
Abiri R, Abdul-Hamid H, Sytar O, Abiri R, Bezerra de Almeida E Jr., Sharma SK, Bulgakov VP, Arroo RRJ, Malik S. A Brief Overview of Potential Treatments for Viral Diseases Using Natural Plant Compounds: The Case of SARS-Cov. Molecules. 2021; 26(13):3868. https://doi.org/10.3390/molecules26133868
Chicago/Turabian StyleAbiri, Rambod, Hazandy Abdul-Hamid, Oksana Sytar, Ramin Abiri, Eduardo Bezerra de Almeida, Jr., Surender K. Sharma, Victor P. Bulgakov, Randolph R. J. Arroo, and Sonia Malik. 2021. "A Brief Overview of Potential Treatments for Viral Diseases Using Natural Plant Compounds: The Case of SARS-Cov" Molecules 26, no. 13: 3868. https://doi.org/10.3390/molecules26133868