
A Bifunctional Peptide Conjugate That Controls Infections of Erwinia amylovora in Pear Plants


 ,
,  ,
,  , and
, and
Abstract
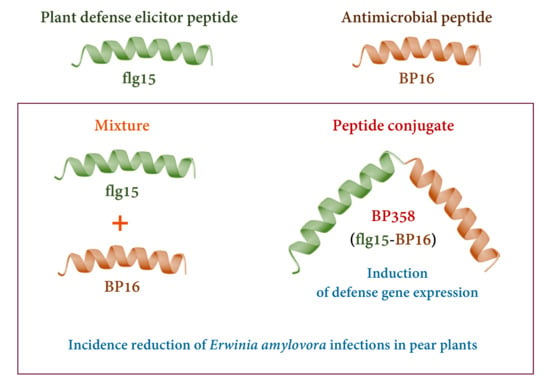
:
1. Introduction
2. Results
2.1. Design and Solid-Phase Synthesis of the Peptides
2.2. In Vitro Biological Activity of Peptides
2.3. Effect of Peptides on Defense Gene Expression of Tomato Plants
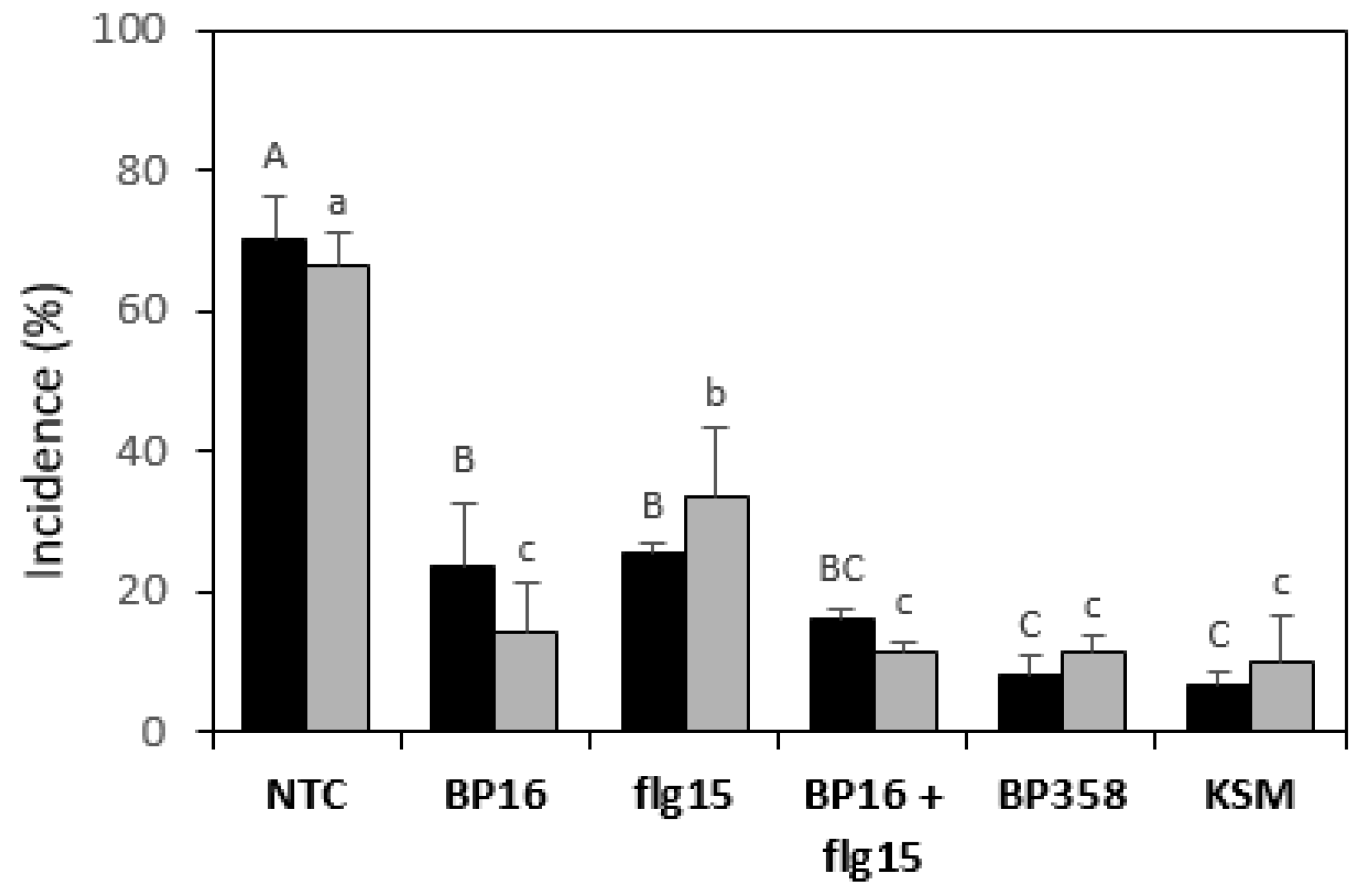
2.4. Activity of Peptides in Planta
3. Discussion
4. Materials and Methods
4.1. General Methods
4.2. General Procedure for the Solid-Phase Synthesis of Peptides
4.3. Bacterial Strains and Growth Conditions
4.4. Antibacterial Activity
4.5. Analysis of Bactericidal Activity
4.6. Hemolytic Activity
4.7. Effect of Peptide Infiltration on Tobacco Leaves
4.8. Effect of Peptide Treatment on Induction of Defense Gene Expression of Tomato Plants
4.9. In Planta Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Oh, C.S.; Beer, S.V. Molecular genetics of Erwinia amylovora involved in the development of fire blight. FEMS Microbiol. Lett. 2005, 253, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferre, R.; Badosa, E.; Feliu, L.; Planas, M.; Montesinos, E.; Bardají, E. Inhibition of plant-pathogenic bacteria by short synthetic cecropin A-melittin hybrid peptides. Appl. Environ. Microbiol. 2006, 72, 3302–3308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aćimović, S.G.; Zeng, Q.; McGhee, G.C.; Sundin, G.W.; Wise, J.C. Control of fire blight (Erwinia amylovora) on apple trees with trunk-injected plant resistance inducers and antibiotics and assessment of induction of pathogenesis-related protein genes. Front. Plant Sci. 2015, 6, 1–10. [Google Scholar]
- Badosa, E.; Montesinos, L.; Camó, C.; Ruz, L.; Cabrefiga, J.; Francés, J.; Gascón, B.; Planas, M.; Feliu, L.; Montesinos, E. Control of fire blight infections with synthetic peptides that elicit plant defense responses. J. Plant Pathol. 2017, 99, 65–73. [Google Scholar]
- Sundin, G.W.; Bender, C.L. Ecological and genetic analysis of copper and streptomycin resistance in Pseudomonas syringae pv. syringae. Appl. Environ. Microbiol. 1993, 59, 1018–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidaver, A.K. Uses of antimicrobials in plant agriculture. Clin. Infect. Dis. 2002, 34, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Ordax, M.; Marco-Noales, E.; López, M.M.; Biosca, E.G. Survival strategy of Erwinia amylovora against copper: Induction of the viable-but-nonculturable state. Appl. Environ. Microbiol. 2006, 72, 3482–3488. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.B.; Temple, T.N. Evaluation of strategies for fire blight control in organic pome fruit without antibiotics. Plant Dis. 2013, 97, 402–409. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Sundin, G.W.; Wang, N. Antibiotic resistance in plant-pathogenic bacteria. Annu. Rev. Phytopathol. 2018, 56, 161–180. [Google Scholar] [CrossRef]
- Badosa, E.; Ferre, R.; Planas, M.; Feliu, L.; Besalú, E.; Cabrefiga, J.; Bardají, E.; Montesinos, E. A library of linear undecapeptides with bactericidal activity against phytopathogenic bacteria. Peptides 2007, 28, 2276–2285. [Google Scholar] [CrossRef] [PubMed]
- Sarojini, V. Antimicrobial peptides for fire blight control. In Small Wonders: Peptides for Disease Control; ACS Symposium Series; Rajasekaran, K., Cary, J.W., Jaynes, J.M., Montesinos, E., Eds.; American Chemical Society: Washington, DC, USA, 2012; Volume 1095, pp. 397–414. [Google Scholar]
- Badosa, E.; Moiset, G.; Montesinos, L.; Talleda, M.; Bardají, E.; Feliu, L.; Planas, M.; Montesinos, E. Derivatives of the antimicrobial peptide BP100 for expression in plant systems. PLoS ONE 2013, 8, e85515. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.; De Zoysa, G.H.; Sarojini, V. Antimicrobial peptides against Pseudomonas syringae pv. actinidiae and Erwinia amylovora: Chemical synthesis, secondary structure, efficacy, and mechanistic investigations. Biopolymers 2014, 102, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Separovic, F.; O’Brien-Simpson, N.; Wade, J.D. Chemically modified and conjugated antimicrobial peptides against superbugs. Chem. Soc. Rev. 2021, 50, 4932–4973. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial peptides as therapeutic agents: Opportunities and challenges. Crit. Rev. Biotechnol. 2020, 40, 978–992. [Google Scholar] [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Keymanesh, K.; Soltani, S.; Sardari, S. Application of antimicrobial peptides in agriculture and food industry. World J. Microbiol. Biotechnol. 2009, 25, 933–944. [Google Scholar] [CrossRef]
- Montesinos, E.; Badosa, E.; Cabrefiga, J.; Planas, M.; Feliu, L.; Bardají, E. Antimicrobial peptides for plant disease control from discovery to application. In Small Wonders: Peptides for Disease Control; ACS Symposium Series; Rajasekaran, K., Cary, J.W., Jaynes, J.M., Montesinos, E., Eds.; American Chemical Society: Washington, DC, USA, 2012; Volume 1095, pp. 235–261. [Google Scholar]
- Ciumac, D.; Gong, H.; Hu, X.; Lu, J.R. Membrane targeting cationic antimicrobial peptides. J. Colloid Interface Sci. 2019, 537, 163–185. [Google Scholar] [CrossRef]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol. 2016, 16, 232. [Google Scholar] [CrossRef] [Green Version]
- Abdul Malik, N.A.; Kumar, I.S.; Nadarajah, K. Elicitor and receptor molecules: Orchestrators of plant defense and immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.B.; Temple, T.N. Induction of systemic acquired resistance aids restoration of tree health in field-grown pear and apple diseased with fire blight. Plant Dis. 2017, 101, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Felix, G.; Duran, J.D.; Volko, S.; Boller, T. Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 1999, 18, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Robatzek, S.; Bittel, P.; Chinchilla, D.; Köchner, P.; Felix, G.; Shiu, S.H.; Boller, T. Molecular identification and characterization of the tomato flagellin receptor LeFLS2, an orthologue of Arabidopsis FLS2 exhibiting characteristically different perception specificities. Plant Mol. Biol. 2007, 64, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, A.; Neundorf, I. Design and application of antimicrobial peptide conjugates. Int. J. Mol. Sci. 2016, 17, 701. [Google Scholar] [CrossRef] [Green Version]
- Lorenzon, E.N.; Piccoli, J.P.; Santos-Filho, N.A.; Cilli, E.M. Dimerization of antimicrobial peptides: A promising strategy to enhance antimicrobial peptide activity. Protein Pept. Lett. 2019, 26, 98–107. [Google Scholar] [CrossRef]
- Wang, C.; Yang, C.; Chen, Y.C.; Ma, L.; Huang, K. Rational design of hybrid peptides: A novel drug design approach. Curr. Med. Sci. 2019, 39, 349–355. [Google Scholar] [CrossRef]
- Wei, X.B.; Wu, R.J.; Si, D.Y.; Liao, X.D.; Zhang, L.L.; Zhang, R.J. Novel hybrid peptide cecropin A (1-8)-LL37 (17-30) with potential antibacterial activity. Int. J. Mol. Sci. 2016, 17, 983. [Google Scholar] [CrossRef]
- Dong, N.; Li, X.R.; Xu, X.Y.; Lv, Y.F.; Li, Z.Y.; Shan, A.S.; Wang, J.L. Characterization of bactericidal efficiency, cell selectivity, and mechanism of short interspecific hybrid peptides. Amino Acids 2018, 50, 453–468. [Google Scholar] [CrossRef]
- Al Tall, Y.; Abualhaijaa, A.; Alsaggar, M.; Almaaytah, A.; Masadeh, M.; Alzoubi, K.H. Design and characterization of a new hybrid peptide from LL-37 and BMAP-27. Infect. Drug Resist. 2019, 12, 1035–1045. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Zhang, D.; Su, P.; Wei, Y.; Wang, Z.; Wang, P.X.; Dai, C.J.; Gong, G.L. Design, recombinant expression, and antibacterial activity of a novel hybrid magainin-thanatin antimicrobial peptide. Prep. Biochem. Biotech. 2019, 49, 427–434. [Google Scholar] [CrossRef]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W.; Somsri, S.; Aunpad, R. A novel, rationally designed, hybrid antimicrobial peptide, inspired by cathelicidin and aurein, exhibits membrane-active mechanisms against Pseudomonas aeruginosa. Sci. Rep. 2020, 10, 9117. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, J.; Cui, Q.; Jia, B.-Y.; Pei, Z.-H.; Odah, K.A.; Wang, Y.-M.; Dong, W.-L.; Kong, L.-C.; Ma, H.-X. Design and characterization of a novel hybrid antimicrobial peptide OM19R based on oncocin and MDAP-2. Int. J. Pept. Res. Ther. 2020, 26, 1839–1846. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, X.; Zhang, R.; Petitte, J.N.; Si, D.; Li, Z.; Cheng, J.; Du, M. Design and development of a novel peptide for treating intestinal inflammation. Front. Immunol. 2019, 10, 1841. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, B.; Hanif, Q.; Wei, X.; Zhang, L.; Sabir, N.; Li, Z.; Cheng, J.; Khan, S.A.; Basit, A.; Shahid, M.; et al. In vitro impact of yeast expressed hybrid peptide CATH-2TP5 as a prophylactic measure toward sepsis and inflammation. Front. Bioeng. Biotechnol. 2020, 8, 454. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Zhou, T.; Zhang, J.; Xu, J.; Guo, X.; Hu, H.; Zhang, X.; Hu, M.; Li, J.; Yang, W.; et al. Enhanced cell selectivity of hybrid peptides with potential antimicrobial activity and immunomodulatory effect. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129532. [Google Scholar] [CrossRef]
- Che, Y.Z.; Li, Y.R.; Zou, H.S.; Zou, L.F.; Zhang, B.; Chen, G.Y. A novel antimicrobial protein for plant protection consisting of a Xanthomonas oryzae harpin and active domains of cecropin A and melittin. Microb. Biotechnol. 2011, 4, 777–793. [Google Scholar] [CrossRef] [Green Version]
- Granoth, R.; Vadai, E.; Burstein, Y.; Fridkin, M.; Tzehoval, E. Tuftsin-THF-γ2 chimeric peptides: Potential novel immomodulators. Immunopharmacology 1997, 37, 43–52. [Google Scholar] [CrossRef]
- Tiwari, S.K.; Sutyak Noll, K.; Cavera, V.L.; Chikindas, M.L. Improved antimicrobial activities of synthetic-hybrid bacteriocins designed from enterocin E50-52 and pediocin PA-1. Appl. Environ. Microbiol. 2015, 81, 1661–1667. [Google Scholar] [CrossRef] [Green Version]
- Horn, M.; Neundorf, I. Design of a novel cell-permeable chimeric peptide to promote wound healing. Sci. Rep. 2018, 8, 16279. [Google Scholar] [CrossRef]
- Wade, H.M.; Darling, L.E.O.; Elmore, D.E. Hybrids made from antimicrobial peptides with different mechanisms of action show enhanced membrane permeabilization. Biochim. Biophys. Acta Biomembr. 2019, 1861, 182980. [Google Scholar] [CrossRef]
- Sheard, D.E.; O’Brien-Simpson, N.M.; Wade, J.D.; Separovic, F. Combating bacterial resistance by combination of antibiotics with antimicrobial peptides. Pure Appl. Chem. 2019, 91, 199–209. [Google Scholar] [CrossRef]
- Yan, H.; Hancock, R.E.W. Synergistic interactions between mammalian antimicrobial defense peptides. Antimicrob. Agents Chemother. 2001, 45, 1558–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Baeder, D.Y.; Regoes, R.R.; Rolff, J. Combination effects of antimicrobial peptides. Antimicrob. Agents Chemother. 2016, 60, 1717–1724. [Google Scholar] [CrossRef] [Green Version]
- Amso, Z.; Hayouka, Z. Antimicrobial random peptide cocktails: A new approach to fight pathogenic bacteria. Chem. Commun. 2019, 55, 2007–2014. [Google Scholar] [CrossRef]
- Remington, J.M.; Liao, C.; Sharafi, M.; Ste Marie, E.J.; Ferrell, J.B.; Hondal, R.J.; Wargo, M.J.; Schneebeli, S.T.; Li, J. Aggregation state of synergistic antimicrobial peptides. J. Phys. Chem. Lett. 2020, 11, 9501–9506. [Google Scholar] [CrossRef]
- Nishida, M.; Imura, Y.; Yamamoto, M.; Kobayashi, S.; Yano, Y.; Matsuzaki, K. Interaction of a magainin-PGLa hybrid peptide with membranes: Insight into the mechanism of synergism. Biochemistry 2007, 46, 14284–14290. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Hunter, H.N.; Matsuzaki, K.; Vogel, H.J. Solution NMR studies of amphibian antimicrobial peptides: Linking structure to function? Biochim. Biophys. Acta 2009, 1788, 1639–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolscher, J.; Nazmi, K.; van Marle, J.; van ‘t Hof, W.; Veerman, E. Chimerization of lactoferricin and lactoferrampin peptides strongly potentiates the killing activity against Candida albicans. Biochem. Cell Biol. 2012, 90, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Nazmi, K.; Bolscher, J.G.; Vogel, H.J. Structural and biophysical characterization of an antimicrobial peptide chimera comprised of lactoferricin and lactoferrampin. Biochim. Biophys. Acta 2012, 1818, 762–775. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Li, D.; Zhang, R.; Shuai, L.; Schulze, B.; Kroth, P.G.; Zhan, D.; Wang, D. Defense responses in female gametophytes of Saccharina japonica (Phaeophyta) induced by flg22-derived peptides. J. Appl. Phycol. 2016, 28, 1793–1801. [Google Scholar] [CrossRef] [Green Version]
- Oliveras, À.; Baró, A.; Montesinos, L.; Badosa, E.; Montesinos, E.; Feliu, L.; Planas, M. Antimicrobial activity of linear lipopeptides derived from BP100 towards plant pathogens. PLoS ONE 2018, 13, e0201571. [Google Scholar] [CrossRef]
- Aslam, S.N.; Erbs, G.; Morrissey, K.L.; Newman, M.A.; Chinchilla, D.; Boller, T.; Molinaro, A.; Jackson, R.W.; Cooper, R.M. Microbe-associated molecular pattern (MAMP) signatures, synergy, size and charge: Influences on perception or mobility and host defence responses. Mol. Plant Pathol. 2009, 10, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Danna, C.H.; Millet, Y.A.; Koller, T.; Han, S.W.; Bent, A.F.; Ronald, P.C.; Ausubel, F.M. The Arabidopsis flagellin receptor FLS2 mediates the perception of Xanthomonas Ax21 secreted peptides. Proc. Natl. Acad. Sci. USA 2011, 108, 9286–9291. [Google Scholar] [CrossRef] [Green Version]
- Dobson, A.J.; Purves, J.; Kamysz, W.; Rolff, J. Comparing selection on S. aureus between antimicrobial peptides and common antibiotics. PLoS ONE 2013, 8, e76521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, E.; Colescott, R.L.; Bossinger, C.D.; Cook, P.I. Color test for detection of free terminal amino groups in the solid-phase synthesis of peptides. Anal. Biochem. 1970, 34, 595–598. [Google Scholar] [CrossRef]
- Hazel, W.; Civerolo, E. Procedures for growth and inoculation of Xanthomonas fragariae, causal organism of angular leaf spot of strawberry. Plant Dis. 1980, 64, 178–181. [Google Scholar] [CrossRef]
- Cabrefiga, J.; Montesinos, E. Analysis of aggressiveness of Erwinia amylovora using disease-dose and time relationships. Phytopathology 2005, 95, 1430–1437. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Güell, I.; Cabrefiga, J.; Badosa, E.; Ferre, R.; Talleda, M.; Bardají, E.; Planas, M.; Feliu, L.; Montesinos, E. Improvement of the efficacy of linear undecapeptides against plant-pathogenic bacteria by incorporation of D-amino acids. Appl. Environ. Microbiol. 2011, 77, 2667–2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | tR (min) 1 | Purity (%) 2 | HRMS |
|---|---|---|---|---|
| BP16 | KKLFKKILKKL-NH2 | 6.04 | >99 | 1386.0070 [M + H]+ |
| flg15 | RINSAKDDAAGLQIA-OH | 5.80 | >99 | 1542.8242 [M + H]+ |
| BP358 (flg15-BP16) | RINSAKDDAAGLQIA-KKLFKKILKKL-NH2 | 6.94 | >99 | 1455.9087 [M + 2H]2+ |
| BP359 (BP16-flg15) | KKLFKKILKKL-RINSAKDDAAGLQIA-OH | 6.04 | >99 | 1456.4023 [M + 2H]2+ |
| Peptide | MIC (μM) | Hemolysis 2 (%) | Tobacco Lesion 3(cm) | |||||
|---|---|---|---|---|---|---|---|---|
| Ea 1 | Pss 1 | Psa 1 | Xap 1 | Xf 1 | Xav 1 | 375 μM | 250 μM | |
| BP16 | 25–50 | 6.2–12.5 | 3.1–6.2 | 6.2–12.5 | 12.5–25 | 12.5–25 | 3 ± 3.5 | 0.26 ± 0.1 |
| flg15 | >100 | >100 | >100 | >100 | >100 | >100 | 0 ± 0 | 0 ± 0 |
| BP358 (flg15-BP16) | 3.1–6.2 | 6.2–12.5 | 3.1–6.2 | 3.1–6.2 | 1.6–3.1 | 1.6–3.1 | 5 ± 0.2 | 0.42 ± 0.1 |
| BP359 (BP16-flg15) | > 50 | 12.5–25 | 12.5–25 | 25–50 | 12.5–25 | 3.1–6.2 | 0.8 ± 0.5 | 0.30 ± 0.1 |
| BP16 + flg154 | 12.5–25 | 6.2–12.5 | 3.1–6.2 | 3.1–6.2 | 25–50 | 12.5–25 | 1 ± 0.3 | 0.30 ± 0.1 |
| Reference 1 | Peptides 2,3 | ||||
|---|---|---|---|---|---|
| Genes | ASM | flg15 | BP16 | BP358 | BP16 + flg15 3 |
| Harp | 3.8 | 3.3 | 0.9 | 4.6 | 1.0 |
| PR1 | 2.8 | 30.4 | 0.1 | 0.3 | 0.2 |
| GluA | 4.9 | 12.7 | 0.1 | 4.2 | 2.8 |
| PPO | 5.6 | 4.8 | 1.6 | 0.7 | 0.3 |
| LOX | 18.7 | 1.6 | 0.9 | 2.3 | 0.4 |
| PinII | 2.6 | 5.2 | 0.4 | 0.5 | 1.5 |
| Sub1 | 3.9 | 12.3 | 0.3 | 3.8 | 1.4 |
| ERT3 | 3.9 | 3.6 | 0.2 | 0.9 | 0.9 |
| BCB | 3.5 | 11.3 | 0.6 | 6.1 | 3.5 |
| Osm2 | 1.5 | 11.3 | 0.7 | 2.5 | 0.5 |
| Tas14 | 0.1 | 2.3 | 2.0 | 0.1 | 0.1 |

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caravaca-Fuentes, P.; Camó, C.; Oliveras, À.; Baró, A.; Francés, J.; Badosa, E.; Planas, M.; Feliu, L.; Montesinos, E.; Bonaterra, A. A Bifunctional Peptide Conjugate That Controls Infections of Erwinia amylovora in Pear Plants. Molecules 2021, 26, 3426. https://doi.org/10.3390/molecules26113426
Caravaca-Fuentes P, Camó C, Oliveras À, Baró A, Francés J, Badosa E, Planas M, Feliu L, Montesinos E, Bonaterra A. A Bifunctional Peptide Conjugate That Controls Infections of Erwinia amylovora in Pear Plants. Molecules. 2021; 26(11):3426. https://doi.org/10.3390/molecules26113426
Chicago/Turabian StyleCaravaca-Fuentes, Pau, Cristina Camó, Àngel Oliveras, Aina Baró, Jesús Francés, Esther Badosa, Marta Planas, Lidia Feliu, Emilio Montesinos, and Anna Bonaterra. 2021. "A Bifunctional Peptide Conjugate That Controls Infections of Erwinia amylovora in Pear Plants" Molecules 26, no. 11: 3426. https://doi.org/10.3390/molecules26113426