
Anti-Inflammatory Effect of Simonsinol on Lipopolysaccharide Stimulated RAW264.7 Cells through Inactivation of NF-κB Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
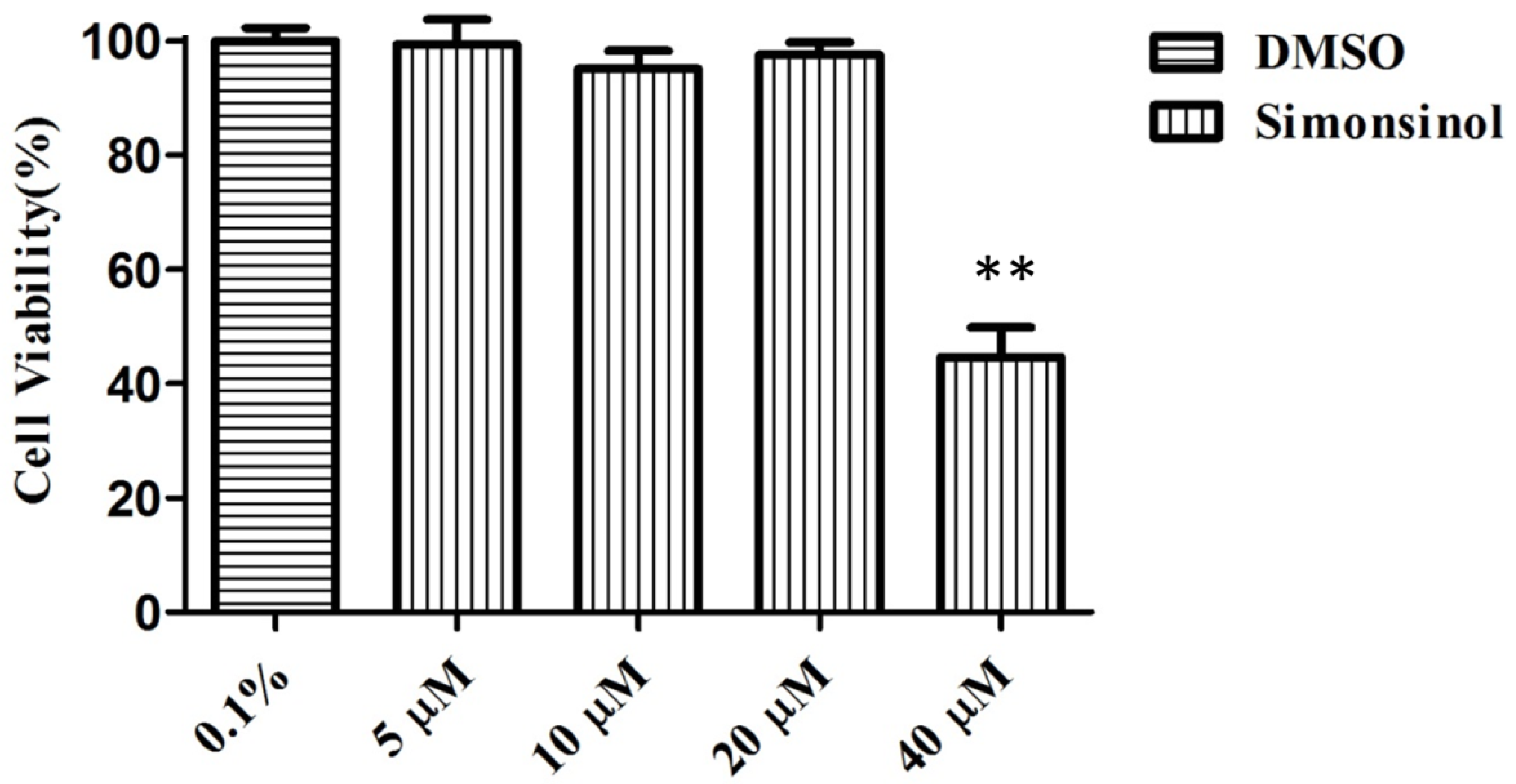
2.1. Effect of Simonsinol on RAW264.7 Cells Viability
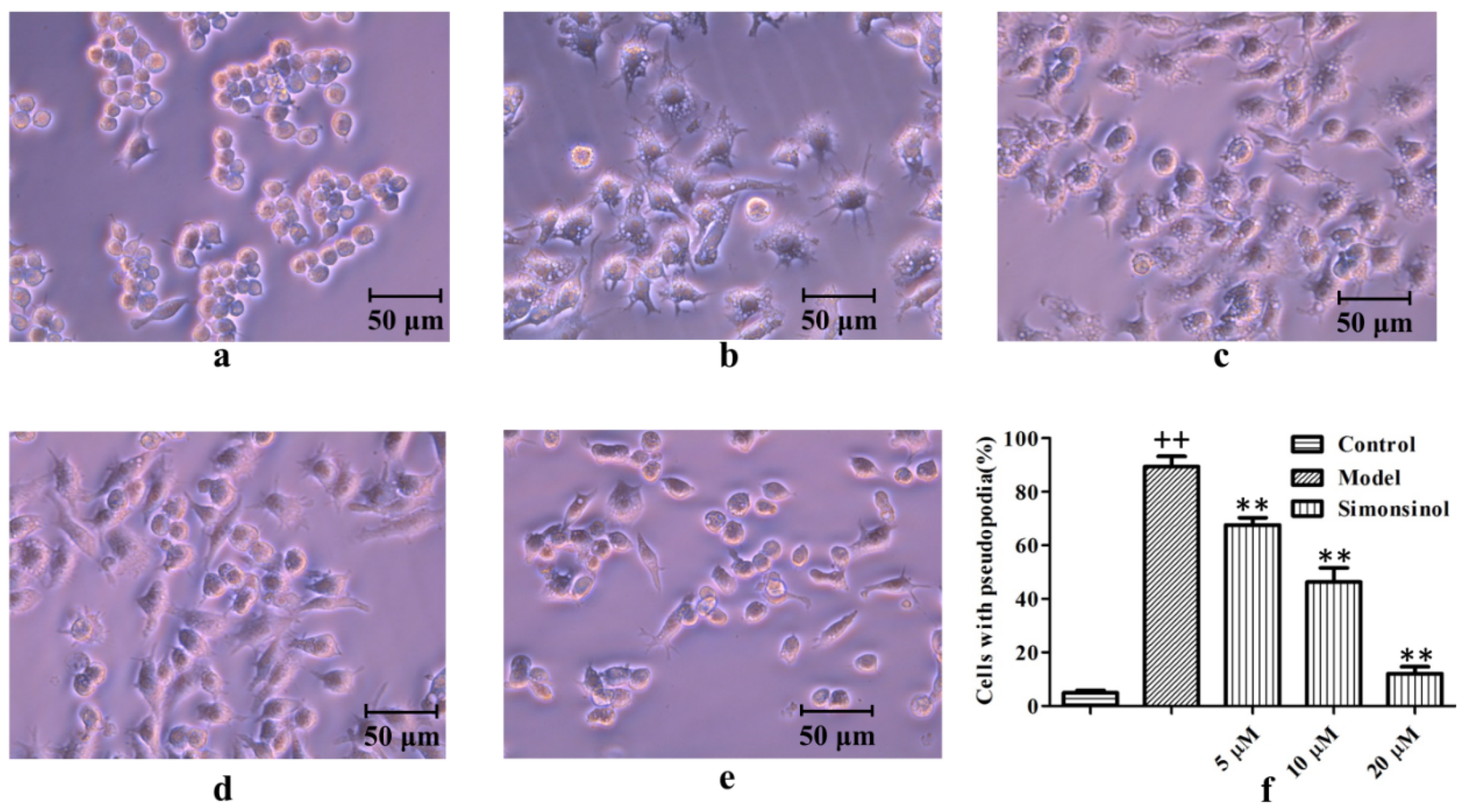
2.2. Effect of Simonsinol on LPS-Stimulated RAW264.7 Cells Morphology
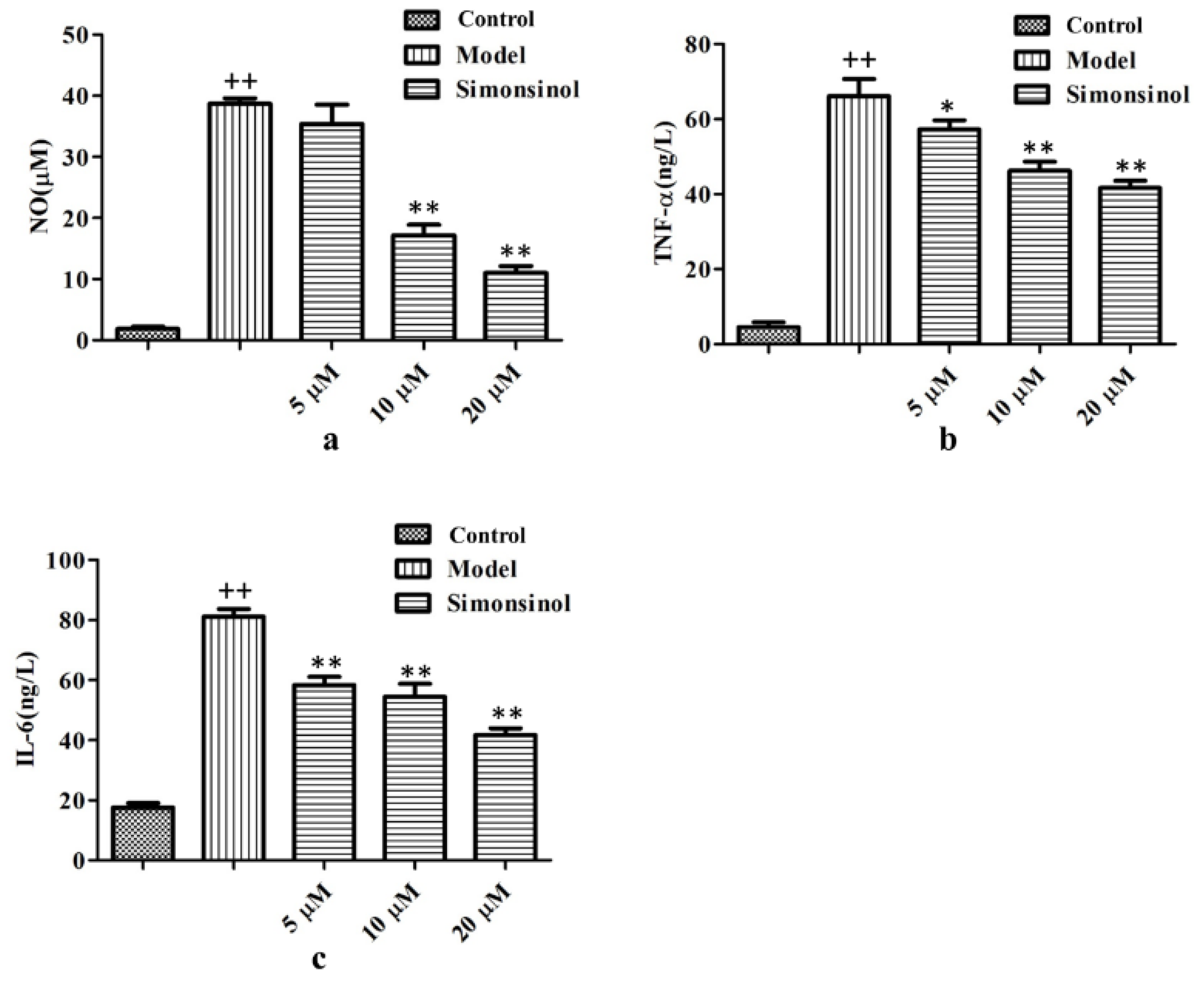
2.3. Simonsinol Inhibits NO, TNF-α, and IL-6 Production in LPS-Stimulated RAW264.7 Cells
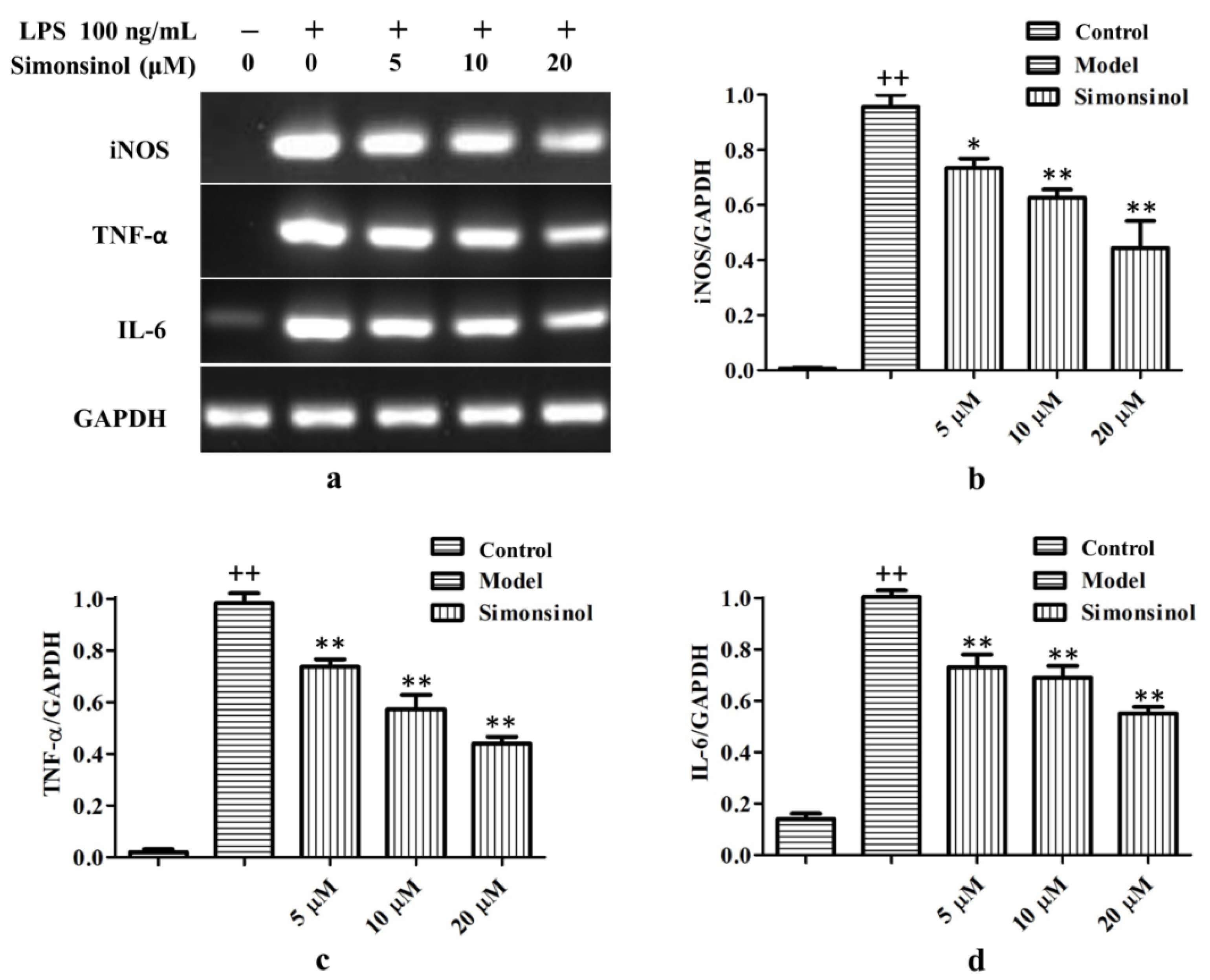
2.4. Simonsinol Suppresses iNOS, TNF-α, and IL-6 Transcription in LPS-Stimulated RAW264.7 Cells
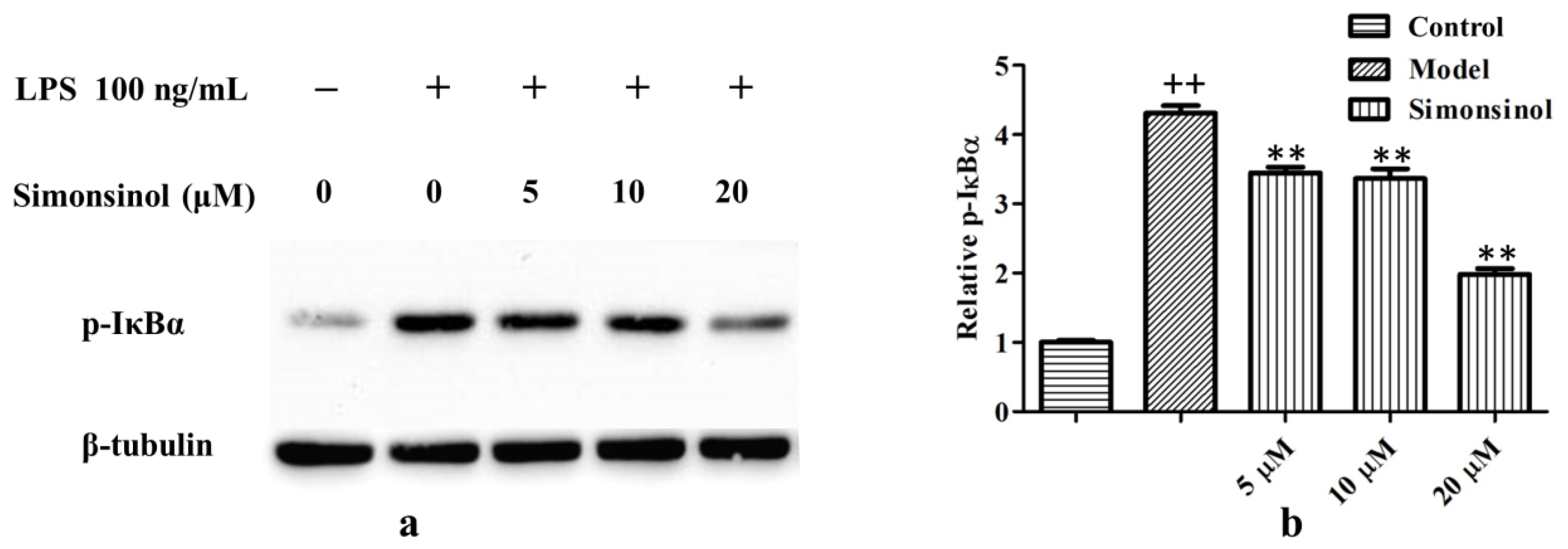
2.5. Simonsinol Suppresses IκBα Phosphorylation in LPS-Stimulated RAW264.7 Cells
3. Discussion and Conclusions
4. Materials and Methods
4.1. General Information
4.2. Reagents
4.3. Plant Material
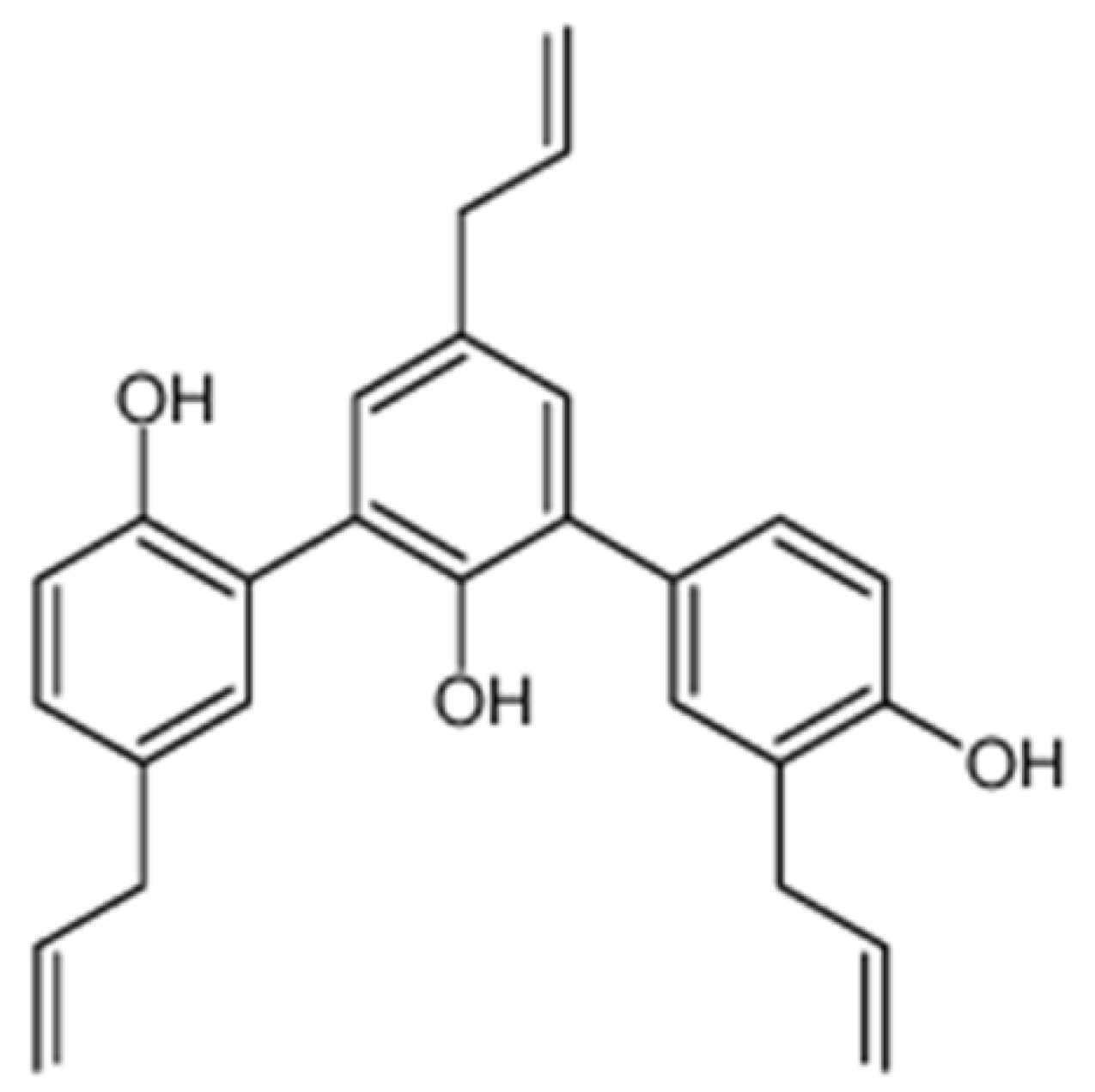
4.4. Extraction and Isolation
4.5. Cell Culture
4.6. MTT Assay
4.7. Effect of Simonsinol on LPS-Stimulated RAW264.7 Cells Morphology
4.8. Determination of Inflammatory Mediators in LPS-Induced RAW264.7 Cells
4.9. RT-PCR
4.10. Western Blot
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Weiss, U. Inflammation. Nature 2008, 454, 427. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Shawki, S.; Ashburn, J.; Signs, S.A.; Huang, E. Colon cancer: Inflammation-associated cancer. Surg. Oncol. Clin. N. Am. 2008, 27, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Xu, Y.; Shi, Y.; Jiang, C. Inflammation role in sensory neuropathy in Chinese patients with diabetes/prediabetes. Clin. Neurol. Neurosurg. 2018, 166, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Angelotti, F.; Parma, A.; Cafaro, G.; Capecchi, R.; Alunno, A.; Puxeddu, I. One year in review 2017: Pathogenesis of rheumatoid arthritis. Clin. Exp. Rheumatol. 2017, 35, 368–378. [Google Scholar] [PubMed]
- Zhu, G.Z.; Han, X.C.; Wang, H.Z.; Yang, Y.Z.; Gao, Y.; Wang, H.L. Effect of Tripterygium Glycosides Tablets in treating rheumatoid arthritis: A systematic review and Meta-analysis. Zhongguo Zhong Yao Za Zhi 2019, 44, 3358–3364. [Google Scholar]
- Mu, H.; Yao, R.B.; Zhao, L.J.; Shen, S.Y.; Zhao, Z.M.; Cai, H. Sinomenine decreases MyD88 expression and improves inflammation-induced joint damage progression and symptoms in rat adjuvant-induced arthritis. Inflammation 2013, 36, 1136–1144. [Google Scholar] [CrossRef]
- Pan, Z.H.; Cheng, L.; Ning, D.S.; Peng, L.Y.; Fu, Y.X.; Li, L.C. Difengpienols A and B, two new sesqui-neolignans with anti-inflammatory activity from the bark of Illicium difengpi. Phytochem. Lett. 2019, 30, 210–214. [Google Scholar] [CrossRef]
- Kouno, I.; Iwamoto, C.; Kameda, Y.; Tanaka, T.; Yang, C.S. A new triphenyl-type neolignan and a biphenylneolignan from the bark of Illicium simonsii. Chem. Pharm. Bull. 1994, 42, 112–114. [Google Scholar] [CrossRef] [Green Version]
- Tak, P.P.; Firestein, G.S. NF-kappa B: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Brown, K.; Gerstberger, S.; Carlson, L.; Franzoso, G.; Siebenlist, U. Control of I kappa B-alpha proteolysis by site-specific, signal-induced phosphorylation. Science 1995, 267, 1485–1488. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.J.; Zhang, T.; Chen, S.X.; Wang, Y.L.; Zhou, C.X.; Mo, J.X.; Wu, Y.J.; Xu, Y.K.; Lin, L.G.; Gan, L.S. Cycloartane triterpenoids from Actaea vaginata with anti-inflammatory effects in LPS-stimulated RAW264.7 macrophages. Phytochemistry 2019, 160, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Yang, J.; Song, P.; Wang, X.; Shi, W. Effects of salvia miltiorrhiza polysaccharides on lipopolysaccharide-induced inflammatory factor release in RAW264.7 cells. J. Interf. Cytok. Res. 2018, 38, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Sang, Y.; Liu, W.; Yu, W.; Wang, X. Anti-inflammatory effects of gingerol on lipopolysaccharide-stimulated RAW 264.7 cells by inhibiting NF-κB signaling pathway. Inflammation 2018, 41, 835–845. [Google Scholar] [CrossRef]
- Zhou, L.; Zhu, D.Y. Neuronal nitric oxide synthase: Structure, subcellular localization, regulation, and clinical implications. Nitric Oxide 2009, 20, 223–230. [Google Scholar] [CrossRef]
- Cheng, X.; Shen, Y.; Li, R. Targeting TNF: A therapeutic strategy for Alzheimer’s disease. Drug Discov. Today 2014, 19, 1822–1827. [Google Scholar] [CrossRef]
- Arias, A.I.; Giles, B.; Eiermann, T.H.; Sterry, W.; Pandey, J.P. Tumor necrosis factor-alpha gene polymorphism in psoriasis. Exp. Clin. Immunogenet. 1997, 14, 118–122. [Google Scholar]
- Hong, D.S.; Angelo, L.S.; Kurzrock, R. Interleukin-6 and its receptor in cancer: Implications for translational therapeutics. Cancer 2007, 110, 1911–1928. [Google Scholar] [CrossRef]
- Haura, E.B.; Livingston, S.; Coppola, D. Autocrine interleukin-6/interleukin-6 receptor stimulation in non-small-cell lung cancer. Clin. Lung Cancer 2006, 7, 273–275. [Google Scholar] [CrossRef]
- Vesely, D.L.; Hoffman, B.; Liebermann, D.A. Phosphatidylinositol 3-kinase/Akt signaling mediates interleukin-6 protection against p53-induced apoptosis in M1 myeloid leukemic cells. Oncogene 2007, 26, 3041–3050. [Google Scholar] [CrossRef] [Green Version]
- Mercurio, F.; Zhu, H.; Murray, B.W.; Shevchenko, A.; Bennett, B.L.; Li, J.W.; Young, D.B.; Barbosa, M.; Mann, M.; Manning, A.; et al. IKK-1 and IKK-2: Cytokine-activated IκB kinases essential for NF-κB activation. Science 1997, 278, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Woronicz, J.D.; Gao, X.; Cao, Z.; Rothe, M.; Goeddel, D.V. IκB kinase-β: NF-κB activation and complex formation with IκB Kinase-α and NIK. Science 1997, 278, 866–869. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, S.S.; Oh, T.H.; Baik, J.S.; Song, G.; Lee, N.H.; Hyun, C.G. Chemical composition, antioxidant, anti-elastase, and anti-inflammatory activities of Illicium anisatum essential oil. Acta Pharm. 2009, 59, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Ito, C.; Itoigawa, M.; Okada, T.; Furukawa, H. Anti-inflammatory activity of phenylpropanoids and phytoquinoids from Illicium species in RBL-2H3 cells. Planta Med. 2007, 73, 662–665. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.Z.; Liu, Y.B.; Yu, S.S.; Qu, J.; Su, D.M. New sesquiterpene lactone and neolignan glycosides with antioxidant and anti-inflammatory activities from the fruits of Illicium oligandrum. Planta Med. 2007, 73, 484–490. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds simonsinol are available from the authors. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.-C.; Pan, Z.-H.; Ning, D.-S.; Fu, Y.-X. Anti-Inflammatory Effect of Simonsinol on Lipopolysaccharide Stimulated RAW264.7 Cells through Inactivation of NF-κB Signaling Pathway. Molecules 2020, 25, 3573. https://doi.org/10.3390/molecules25163573
Li L-C, Pan Z-H, Ning D-S, Fu Y-X. Anti-Inflammatory Effect of Simonsinol on Lipopolysaccharide Stimulated RAW264.7 Cells through Inactivation of NF-κB Signaling Pathway. Molecules. 2020; 25(16):3573. https://doi.org/10.3390/molecules25163573
Chicago/Turabian StyleLi, Lian-Chun, Zheng-Hong Pan, De-Sheng Ning, and Yu-Xia Fu. 2020. "Anti-Inflammatory Effect of Simonsinol on Lipopolysaccharide Stimulated RAW264.7 Cells through Inactivation of NF-κB Signaling Pathway" Molecules 25, no. 16: 3573. https://doi.org/10.3390/molecules25163573