
Effect of Growth Regulators on Stevia rebaudiana Bertoni Callus Genesis and Influence of Auxin and Proline to Steviol Glycosides, Phenols, Flavonoids Accumulation, and Antioxidant Activity In Vitro


 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Callus Induction of Stevia rebaudiana
2.2. S. rebaudiana Callus-Mass Increase
2.3. NAA and PRO Effect on SG Accumulation in S. rebaudiana Callus
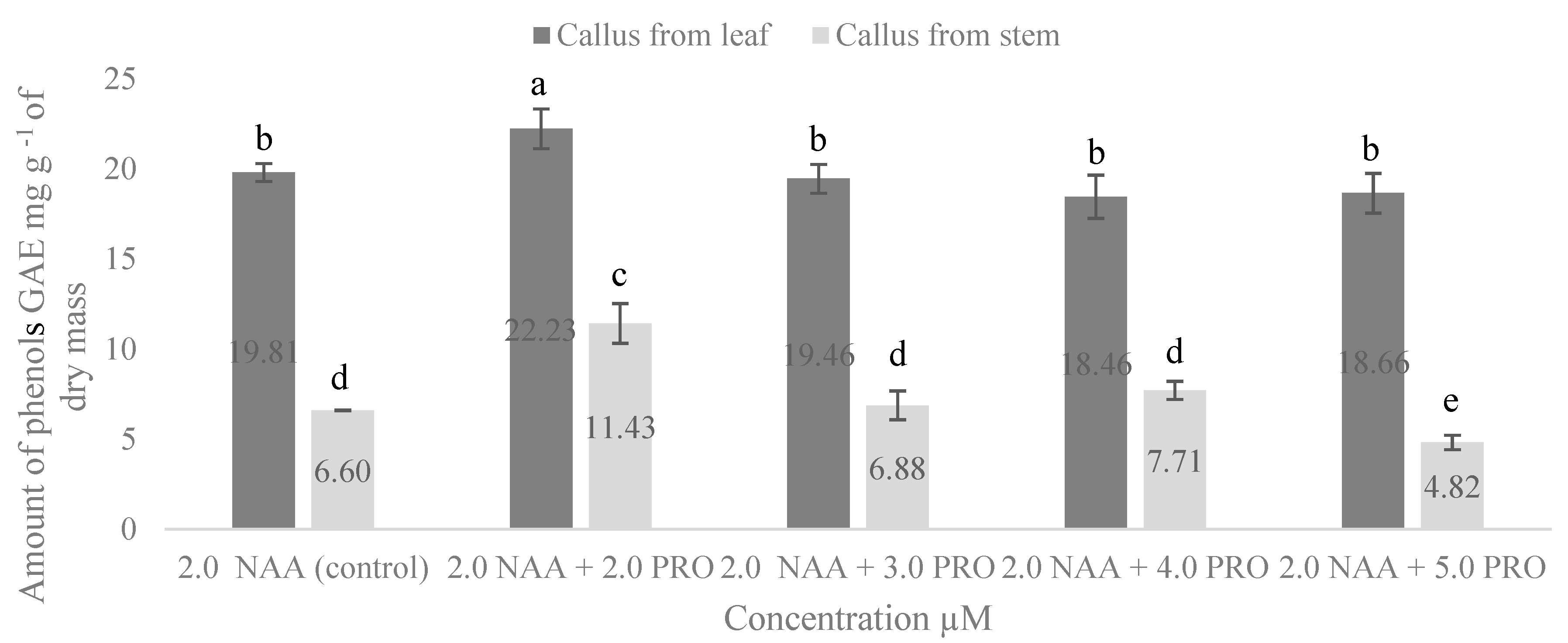
2.4. NAA and PRO Effect on Accumulation of Total Phenolic Compounds and Total Flavonoids in S. rebaudiana Callus
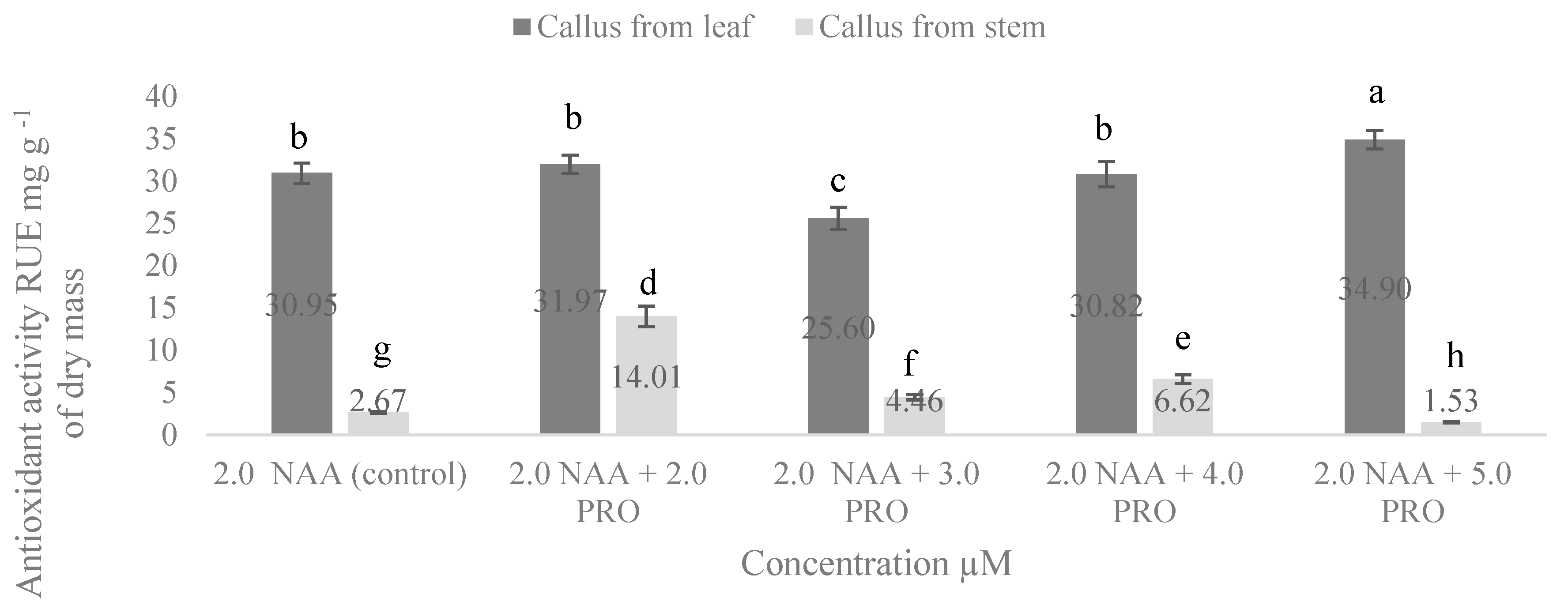
2.5. NAA and PRO Effect on Antioxidant Activity in S. rebaudiana of Cellular Compounds of Callus
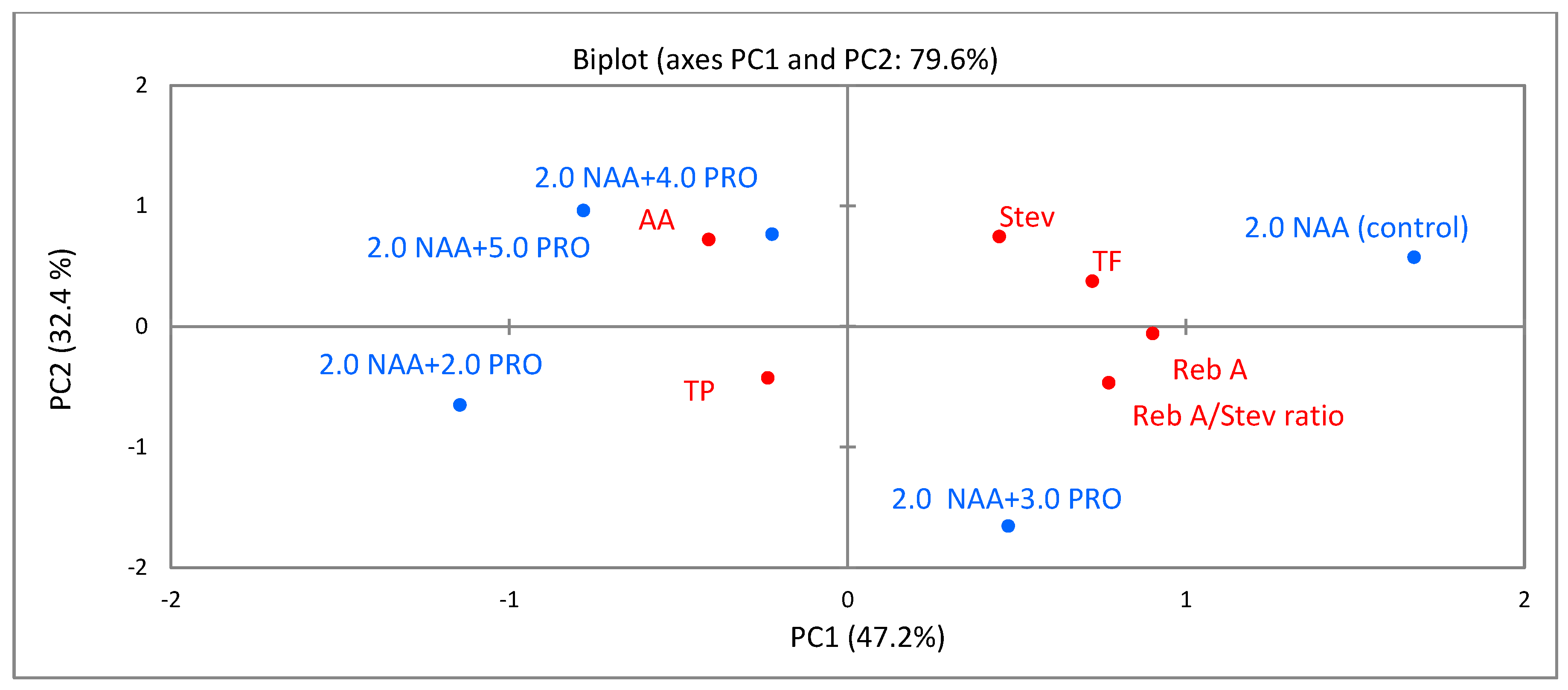
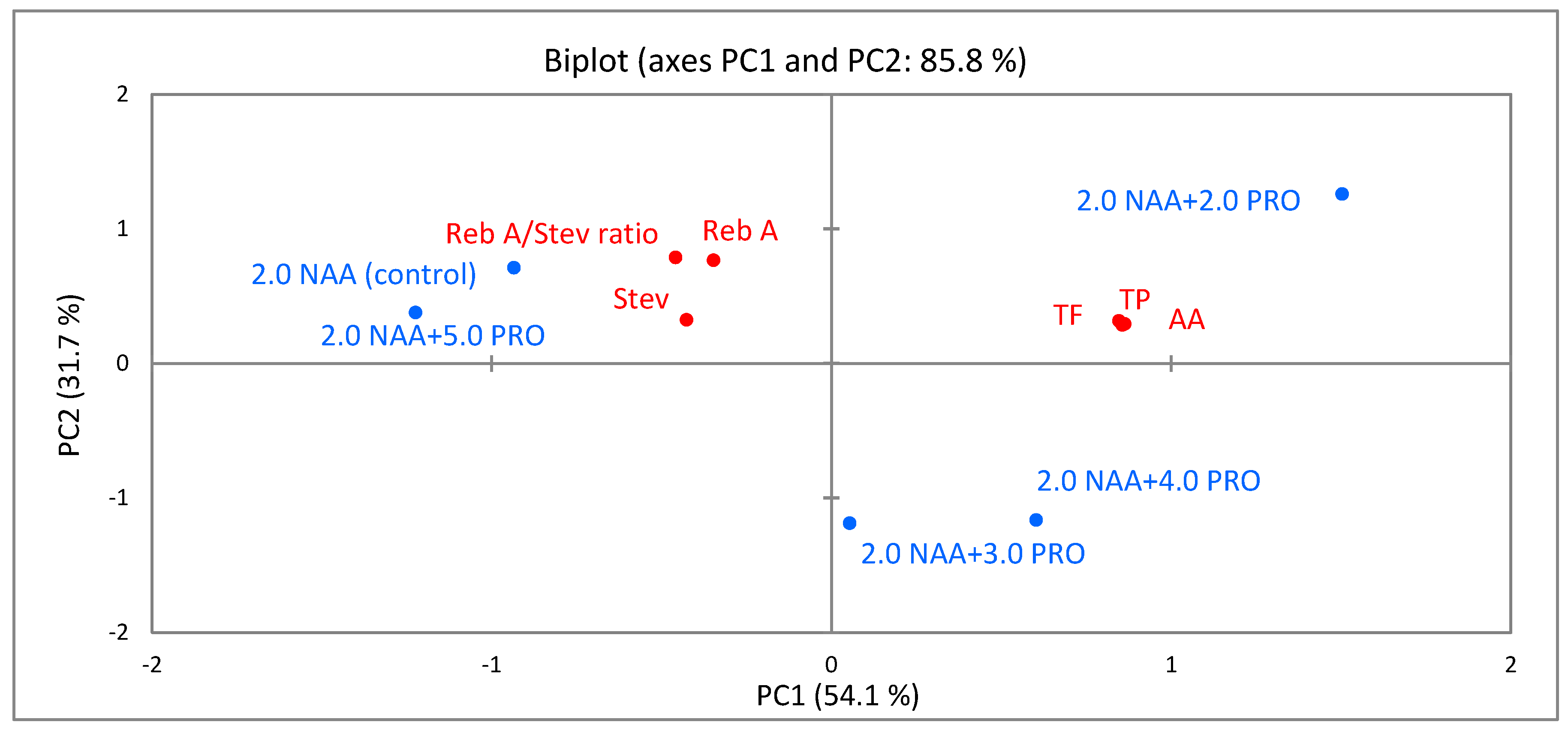
2.6. Principal-Component Analysis (PCA)
3. Materials and Methods
3.1. Plant Material
3.2. In Vitro Preparation of S. rebaudiana Material
3.3. Callus-Formation Determination
3.4. Callus-Mass Determination
3.5. SG Extraction
3.6. HPLC SG Analysis
3.7. Extraction Procedure (Phenolics, Flavonoids, Antioxidant Activity)
3.8. Total-Phenolic-Content Determination
3.9. Total-Flavonoid-Content Determination
3.10. Antioxidant-Activity Determination
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yadav, A.K.; Singh, S.; Dhyani, D.; Ahuja, P.S. A review on the improvement of stevia [Stevia rebaudiana (Bertoni)]. Can. J. Plant Sci. 2011, 91, 1–27. [Google Scholar] [CrossRef]
- Soejarto, D.D.; Compadre, C.M.; Medon, P.J.; Kamath, S.K.; Kinghorn, A.D. Potential sweetening agents of plant origin—II: Field research for sweet-tasting of Stevia spp. Econ. Bot. 1983, 18, 37–41. [Google Scholar]
- Goyal, S.K.; Samsher, G.R.K.; Goyal, R.K. Stevia (Stevia rebaudiana) a bio-sweetener: A review. Int. J. Food Sci. Nutr. 2010, 61, 1–10. [Google Scholar] [CrossRef]
- Tadhani, M.B.; Petel, V.H.; Subhash, R. In vitro antioxidant activities of Stevia rebaudiana leaves and callus. J. Food Compost Anal. 2007, 20, 323–329. [Google Scholar] [CrossRef]
- Kinghorn, A.D.; Compadre, C.M. Less common high-potency sweeteners. In Alternative Sweeteners, 3rd ed.; O’Brien Nabors, L., Ed.; Marcel Dekker Publisher: New York, NY, USA, 2001; pp. 209–233. [Google Scholar]
- Modi, A.; Kumar, N. Conventional and biotechnological approaches to enhance steviol glycosides (SGs) in Stevia rebaudiana Bertoni. Biotechnol. Approaches Med. Aromat. Plants 2018, 53–62. [Google Scholar]
- Kinghorn, A.D.; Soejarto, N.P.D.; Nanayakkara, C.M. A phytochemical screening procedure for sweet entkaurene glycosides in the genus Stevia. J. Nat. Prod. 1984, 47, 439–444. [Google Scholar] [CrossRef]
- Shibata, H.; Sawa, Y.; Oka, T. Steviol and steviol-glycoside: Glucosyltransferase activities in Stevia rebaudiana Bertoni—Purification and partial characterization. Arch. Biochem. Biophys. 1995, 321, 390–396. [Google Scholar] [CrossRef]
- Lailerd, N.; Saengsirisuwan, V.; Sloniger, J.A.; Toskulkao, C.; Henriksen, E.J. Effects of stevioside on glucose transport activity in insulin-sensitive and insulin-528 resistant rat skeletal muscle. Metabolism 2004, 53, 101–107. [Google Scholar] [CrossRef]
- Puri, M.; Sharma, D.; Tiwari, A.K. Downstream processing of stevioside and its 525 potential applications. Biotechnol. Adv. 2011, 29, 781–791. [Google Scholar] [CrossRef]
- Taleie, N.; Hamidoghli, Y.; Rabiei, B.; Hamidoghli, S. Effects of plant density and transplanting date on herbage, stevioside, phenol and flavonoid yield of Stevia rebaudiana Bertoni. Int. J. Agric. Crop Sci. 2012, 4, 298–302. [Google Scholar]
- Kim, D.; Jeond, S.; Lee, C. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Randhir, R.; Lin, Y.-T.; Shetty, K. Phenolics, their antioxidant and antimicrobial activity in dark germinated fenugreek sprouts in response to peptide and phytochemical elicitors. Asia Pac. J. Clin. Nutr. 2004, 13, 295–307. [Google Scholar] [PubMed]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Ghasemzadeh, A.; Ghasemzadeh, N. Flavonoids and phenolic acids: Role and biochemical activity in plants and human. J. Med. Plant. Res. 2011, 5, 6697–6703. [Google Scholar] [CrossRef]
- Gaweł-Bęben, K.; Bujak, T.; Nizioł-Łukaszewska, Z.; Antosiewicz, B.; Jakubczyk, A.; Karaś, M.; Rybczyńska, K. Stevia Rebaudiana Bert. leaf extracts as a multifunctional source of natural antioxidants. Molecules 2015, 20, 5468–5486. [Google Scholar] [CrossRef] [Green Version]
- Rathi, N.; Arya, S. In vitro regeneration through callus culture of medicinally important plant Stevia rebaudiana (Bert.) Bertoni. Int. J. Plant Sci. 2009, 4, 559–563. [Google Scholar]
- Sakaguchi, M.; Kan, T. Japanese researches on Stevia rebaudiana (Bert.) Bertoni and stevioside. Cienc. Cult. 1982, 34, 235–248. [Google Scholar]
- Khalil, S.A.; Zamir, R.; Ahmad, N. Selection of suitable propagation method for consistent plantlets production in Stevia rebaudiana (Bertoni). Saudi J. Biol. Sci. 2014, 21, 566–573. [Google Scholar] [CrossRef] [Green Version]
- Depuydt, S.; Hardtke, C.S. Hormone signalling crosstalk in plant growth regulation. Curr. Biol. 2011, 21, R365–R373. [Google Scholar] [CrossRef]
- Evans, D.A.; Sharp, W.R.; Flick, C.E. Growth and behavior of cell cultures: Embryogenesis and organogenesis. In Plant Tissue Culture: Methods and Applications in Agriculture; Thorpe, T.A., Ed.; Academic Press: New York, NY, USA, 1981; pp. 45–113. [Google Scholar]
- Vasil, I.K.; Thorpe, T.A. Plant. Cell and Tissue Culture; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 18, 25, 25, 46, 210, 543. [Google Scholar]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppi, N.H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell Dev. Biol. Plant. 1996, 32, 272–289. [Google Scholar] [CrossRef]
- Javed, R.; Yucesan, B.; Zia, M.; Gurel, E. Elicitation of Secondary Metabolites in Callus Cultures of Stevia rebaudiana Bertoni Grown Under ZnO and CuO Nanoparticles Stress. Sugar Tech. 2017, 20, 194–201. [Google Scholar] [CrossRef]
- Rao, R.S.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [PubMed]
- Xu, J.; Yin, H.X.; Li, X. Protective effects of proline against cadmium toxicity in micropropagated hyperaccumulator Solanum nigrum L. Plant. Cell Report 2009, 28, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Hsing, Y.I.; Su, W.F.; Chang, W.C. Accumulation of stevioside and rebaudioside A in callus culture of Stevia rebaudiana Bertoni. Bot. Bull. Acad. 1983, 24, 115–119. [Google Scholar]
- Swanson, S.M.; Mahady, G.B.; Beecher, C.W.W. Stevioside biosynthesis by callus, root, shoot and rooted-shoot cultures in vitro. Plant. Cell Tissue Organ. Cult. 1992, 28, 151–157. [Google Scholar] [CrossRef]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant. Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Gupta, P.; Sharma, S.; Saxena, S. Biomass yield and steviol glycoside production in callus and suspension culture of Stevia rebaudiana treated with proline and polyethylene glycol. Appl. Biochem. Biotechnol. 2015, 176, 863–874. [Google Scholar] [CrossRef]
- Maggio, A.; Miyazaki, S.; Veronese, P.; Fujita, T.; Ibeas, J.I.; Damsz, B.; Narasimhan, M.L.; Hasegawa, P.M.; Joly, R.J.; Bressan, R.A. Does proline accumulation play an active role in stress-induced growth reduction? Plant. J. 2002, 31, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Mandal, N. Enhanced development of embryogenic callus in Stevia rebaudiana Bert. by assitive and amino acids. Biotechnology 2010, 9, 368–372. [Google Scholar]
- Lukatkin, A.S.; Silva, J.A.T. Effects of cultivation parameters of Stevia rebaudiana Bertoni callus culture on callus proliferation. Med. Aromat. Plant. Sci. Biotechnol. 2007, 107–111. [Google Scholar]
- Gauchan, D.P.; Dhakal, A.; Sharma, N.; Bhandari, S.; Maskey, E.; Shrestha, N.; Gautam, R.; Giri, S.; Gurung, S. Regenerative callus induction and biochemical analysis of Stevia rebaudiana Bertoni. J. Adv. Lab. Res. Biol. 2014, 5, 41–45. [Google Scholar]
- Uddin, M.S.; Chodhary, M.S.H.; Khan, M.M.M.H.; Uddin, M.B.; Ahmad, R.; Baten, M.A. In vitro propagation of Stevia rebaudiana Bertoni in Bangladesh. Afr. J. Biotechnol. 2006, 5, 1238–1240. [Google Scholar]
- Mathur, S.; Shekhawat, G.S. Establishment and characterization of Stevia rebaudiana (Bertoni) cell suspension culture: An in vitro approach for production of stevioside. Acta Physiol. Plant. 2013, 35, 1–9. [Google Scholar] [CrossRef]
- Patel, R.M.; Shah, R.R. Regeneration of stevia plant through callus culture. Indian J. Pharm. Sci. 2009, 71, 46–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Sharma, S.; Saxena, S. Callusing in Stevia rebaudiana (Natural Sweetener) for steviol glycoside production. World Acad. Sci. Eng. Technol. Int. J. Biol. Biomol. Agric. Food Biotechnol. Eng. 2010, 4, 893–897. [Google Scholar]
- Hendawey, M.H.; Abo, E.I.F.R.R. Biochemical studies on the production of active. GJBB 2014, 9, 76–93. [Google Scholar]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids. 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Rajasekaran, T.; Giridhar, P.; Ravishankar, G.A. Production of steviosides in ex vitro and in vitro grown Stevia rebaudiana Bertoni. J. Sci. Food Agric. 2007, 87, 420–424. [Google Scholar] [CrossRef]
- Massoud, M.I.; Khattab, S.N.; Jad, Y.E.-S.; Bekhit, A.A.; Faham, A.E. Production and physicochemical assessment of new stevia amino acid sweeteners from the natural stevioside. Food Chem. 2014, 173, 979–985. [Google Scholar]
- Chranioti, C.; Chanioti, S.; Tzia, C. Comparison of spray, freeze and oven drying as a means of reducing bitter aftertaste of steviol glycosides (derived from Stevia rebaudiana Bertoni plant)–Evaluation of the final products. Food Chem. 2016, 190, 1151–1158. [Google Scholar] [CrossRef]
- Taware, A.S.; Mukadam, D.S.; Chavan, A.M.; Taware, S.D. Comparative studies of in vitro and in vivo grown plants and callus of Stevia rebaudiana (Bertoni). Int. J. Integr. Biol. 2010, 9, 10–15. [Google Scholar]
- Kahkonen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Muanda, F.; Soulimani, R.; Diop, B.; Dicko, A. Study on chemical composition and biological activities of essential oil and extracts from Stevia rebaudiana Bertoni leaves. Lwt-Food Sci. Technol. 2011, 44, 1865–1872. [Google Scholar] [CrossRef]
- Periche, A.; Koutsidis, G.; Escriche, I. Composition of antioxidants and amino acids in Stevia leaf infusions. Plant. Foods Hum. Nutr. 2014, 69, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Periche, A.; Castelló, M.L.; Heredia, A.; Escriche, I. Influence of drying method on steviol glycosides and antioxidants in Stevia rebaudiana leaves. Food Chem. 2015, 172, 1–6. [Google Scholar] [CrossRef]
- Kwok, D.; Shetty, K. Effects of proline and proline analogs on total phenolic and rosmarinic acid levels in shoot cclones of thyme (Thymus vulgaris L.). J. Food Biochem. 1997, 22, 37–51. [Google Scholar] [CrossRef]
- Samadi, S.; Ghasemnezhad, A.; Alizade, M. Fresh weigh, total phenol, total flavonoids, antioxidant and pal enzyme activity of stevia callus variation affected by salicylic acid and methyl jasmonate. J. Plant. Proc. Func. 2019, 8, 325–337. [Google Scholar]
- Radić, S.; Vujčić, V.; Glogoški, M.; Radić-Stojković, M. Influence of pH and plant growth regulators on secondary metabolite production and antioxidant activity of Stevia rebaudiana (Bert). Period. Biol. 2016, 118, 9–19. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and biossays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bergs, D.; Burghoff, B.; Joehnck, M.; Martin, G.; Schembecker, G. Fast and isocratic HPLC-method for steviol glycosides analysis from Stevia rebaudiana leaves. J. Verbrauch Lebensm. 2012, 7, 147–154. [Google Scholar] [CrossRef]
- Butkutė, B.; Benetis, R.; Padarauskas, A.; Cesevičienė, J.; Dagilytė, A.; Taujenis, L.; Rodovičius, H.; Lemežienė, N. Young herbaceous legumes – a natural reserve of bioactive compounds and antioxidants for healthy food and supplements. J. Appl. Bot. Food Qual. 2017, 90, 346–353. [Google Scholar]
- Kaškonienė, V.; Ruočkuvienė, G.; Kaškonas, P.; Akuneca, I.; Maruška, A. Chemometric analysis of bee pollen based on volatile and phenolic compound compositions and antioxidant properties. Food Anal. Methods 2015, 8, 1150–1163. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Callus Formation Frequency % | |
|---|---|---|
| After One Week | After Two Weeks | |
| Callus from Leaf | ||
| MS (−) | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 1.0 µM NAA | 69.4 ± 6.3 b | 100.0 ± 0.0 a |
| 2.0 µM NAA | 83.3 ± 0.0 a | 100.0 ± 0.0 a |
| 3.0 µM NAA | 69.4 ± 4.4 b | 87.7 ± 0.8 cb |
| 1.0 µM NAA + 1.0 µM BAP | 58.3 ± 0.0 bcd | 100.0 ± 0.0 a |
| 2.0 µM NAA + 1.0 µM BAP | 58.3 ± 0.0 bcd | 100.0 ± 0.0 a |
| 3.0 µM NAA + 1.0 µM BAP | 58.3 ± 0.0 bcd | 100.0 ± 0.0 a |
| Average | 66.17 | 97.95 |
| Callus from stem | ||
| MS (−) | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 1.0 µM NAA | 47.2 ± 6.3 d | 80.3 ± 5.2 c |
| 2.0 µM NAA | 61.6 ± 0.0 bc | 91.0 ± 9.2 ab |
| 3.0 µM NAA | 63.8 ± 4.8 b | 69.5 ± 0.1 d |
| 1.0 µM NAA + 1.0 µM BAP | 59.5 ± 1.3 bcd | 79.3 ± 0.5 c |
| 2.0 µM NAA + 1.0 µM BAP | 58.3 ± 6.9 bcd | 83.2 ± 4.7 cb |
| 3.0 µM NAA + 1.0 µM BAP | 49.9 ± 6.2 cd | 81.8 ± 2.0 cb |
| Average | 56.72 | 80.85 |
| p-Value (growth regulator x explant type) | <0.00001 | n.s |
| Treatments | Callus Mass Increase mg | |
|---|---|---|
| After one Month | After two Months | |
| Callus from Leaf | ||
| MS (−) | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 1.0 µM NAA | 10.0 ± 0.9 d | 19.3 ± 1.9 d |
| 2.0 µM NAA | 25.7 ± 1.2 a | 39.3 ± 0.1 a |
| 3.0 µM NAA | 9.4 ± 0.1 d | 36.5 ± 1.0 b |
| 1.0 µM NAA + 1.0 µM BAP | 16.1 ± 0.3 c | 26.5 ± 1.8 c |
| 2.0 µM NAA + 1.0 µM BAP | 18.4 ± 1.0 b | 29.7 ± 1.9 bc |
| 3.0 µM NAA + 1.0 µM BAP | 15.8 ± 0.8 c | 32.1 ± 2.2 b |
| Average | 15.90 | 30.5 |
| Callus from stem | ||
| MS (−) | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 1.0 µM NAA | 2.8 ± 0.3 ef | 6.3 ± 0.5 f |
| 2.0 µM NAA | 3.7 ± 0.3 e | 8.3 ± 0.1 e |
| 3.0 µM NAA | 0.4 ± 0.05 g | 2.5 ± 0.1 g |
| 1.0 µM NAA + 1.0 µM BAP | 0.6 ± 0.0 g | 5.0 ± 0.8 f |
| 2.0 µM NAA +1.0 µM BAP | 1.0 ± 0.3 g | 6.7 ± 0.3 f |
| 3.0 µM NAA + 1.0 µM BAP | 1.8 ± 0.2 fg | 6.3 ± 0.2 f |
| Average | 1.71 | 5.85 |
| p-Value (growth regulator x explant type) | <0.00100 | <0.00001 |
| Treatments | Stevioside | Rebaudioside A | RebA/Stev Ratio |
|---|---|---|---|
| mg g−1 | |||
| Callus from Leaf | |||
| 2.0 µM NAA (control) | 0.40 ± 0.05 a | 0.09 ± 0.001 a | 0.23 ± 0.02 b |
| 2.0 µM NAA + 2.0 µM PRO | 0.20 ± 0.09 c | 0.009 ± 0.0001 f | 0.04 ± 0.0001 d |
| 2.0 µM NAA + 3.0 µM PRO | 0.20 ± 0.05 c | 0.05 ± 0.005 b | 0.25 ± 0.10 b |
| 2.0 µM NAA + 4.0 µM PRO | 0.39 ± 0.01 ab | 0.02 ± 0.003 de | 0.05 ± 0.05 d |
| 2.0 µM NAA + 5.0 µM PRO | 0.30 ± 0.05 b | 0.01 ± 0.004 ef | 0.03 ± 0.02 d |
| Average | 0.329 | 0.036 | 0.12 |
| Callus from stem | |||
| 2.0 µM NAA (control) | 0.06 ± 0.005 d | 0.04 ± 0.003 bc | 0.66 ± 0.11 b |
| 2.0 µM NAA + 2.0 µM PRO | 0.04 ± 0.003 e | 0.03 ± 0.005 cd | 0.75 ± 0.07 b |
| 2.0 µM NAA + 3.0 µM PRO | 0.04 ± 0.008 e | 0.009 ± 0.0001 f | 0.22 ± 0.01 c |
| 2.0 µM NAA + 4.0 µM PRO | 0.04 ± 0.003 e | 0.005 ± 0.0009 f | 0.12 ± 0.01 cd |
| 2.0 µM NAA + 5.0 µM PRO | 0.04 ± 0.003 e | 0.04 ± 0.002 bc | 1.00 ± 0.04 a |
| Average | 0.044 | 0.025 | 0.55 |
| p-Value (proline concentration x explant type) | <0.00542 | <0.00001 | <0.00001 |
| Callus from Leaf. | r2 | Coefficient of Correlation % | p-Value |
|---|---|---|---|
| 2.0 µM NAA (control) | 0.93 | 96.80 | 0.0320 |
| 2.0 µM NAA + 2.0 µM PRO | 0.47 | 68.32 | n.s.1 |
| 2.0 µM NAA + 3.0 µM PRO | 0.96 | 98.03 | 0.0196 |
| 2.0 µM NAA + 4.0 µM PRO | 0.71 | 84.47 | 0.0167 |
| 2.0 µM NAA + 5.0 µM PRO | 0.89 | 94.66 | 0.0146 |
| Callus from stem | |||
| 2.0 µM NAA (control) | 0.96 | 97.79 | 0.0020 |
| 2.0 µM NAA + 2.0 µM PRO | 0.56 | 75.08 | n.s. |
| 2.0 µM NAA + 3.0 µM PRO | 0.85 | 92.50 | 0.0444 |
| 2.0 µM NAA + 4.0 µM PRO | 0.91 | 95.81 | 0.0419 |
| 2.0 µM NAA + 5.0 µM PRO | 0.43 | 65.44 | n.s. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blinstrubienė, A.; Burbulis, N.; Juškevičiūtė, N.; Vaitkevičienė, N.; Žūkienė, R. Effect of Growth Regulators on Stevia rebaudiana Bertoni Callus Genesis and Influence of Auxin and Proline to Steviol Glycosides, Phenols, Flavonoids Accumulation, and Antioxidant Activity In Vitro. Molecules 2020, 25, 2759. https://doi.org/10.3390/molecules25122759
Blinstrubienė A, Burbulis N, Juškevičiūtė N, Vaitkevičienė N, Žūkienė R. Effect of Growth Regulators on Stevia rebaudiana Bertoni Callus Genesis and Influence of Auxin and Proline to Steviol Glycosides, Phenols, Flavonoids Accumulation, and Antioxidant Activity In Vitro. Molecules. 2020; 25(12):2759. https://doi.org/10.3390/molecules25122759
Chicago/Turabian StyleBlinstrubienė, Aušra, Natalija Burbulis, Neringa Juškevičiūtė, Nijolė Vaitkevičienė, and Rasa Žūkienė. 2020. "Effect of Growth Regulators on Stevia rebaudiana Bertoni Callus Genesis and Influence of Auxin and Proline to Steviol Glycosides, Phenols, Flavonoids Accumulation, and Antioxidant Activity In Vitro" Molecules 25, no. 12: 2759. https://doi.org/10.3390/molecules25122759