
The Stimulatory Effects of Nanochitin Whisker on Carbon and Nitrogen Metabolism and on the Enhancement of Grain Yield and Crude Protein of Winter Wheat


Abstract
:1. Introduction
2. Results
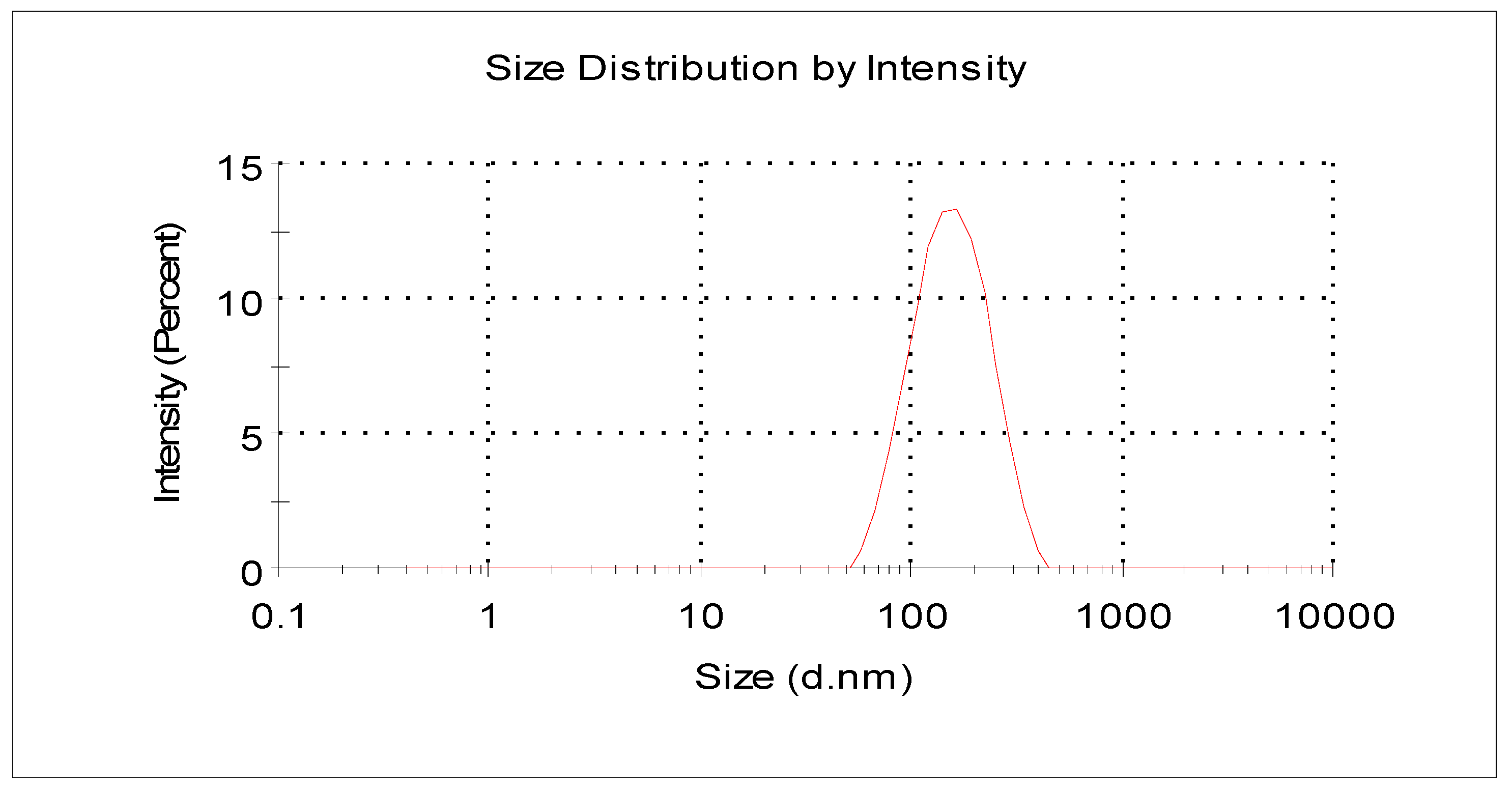
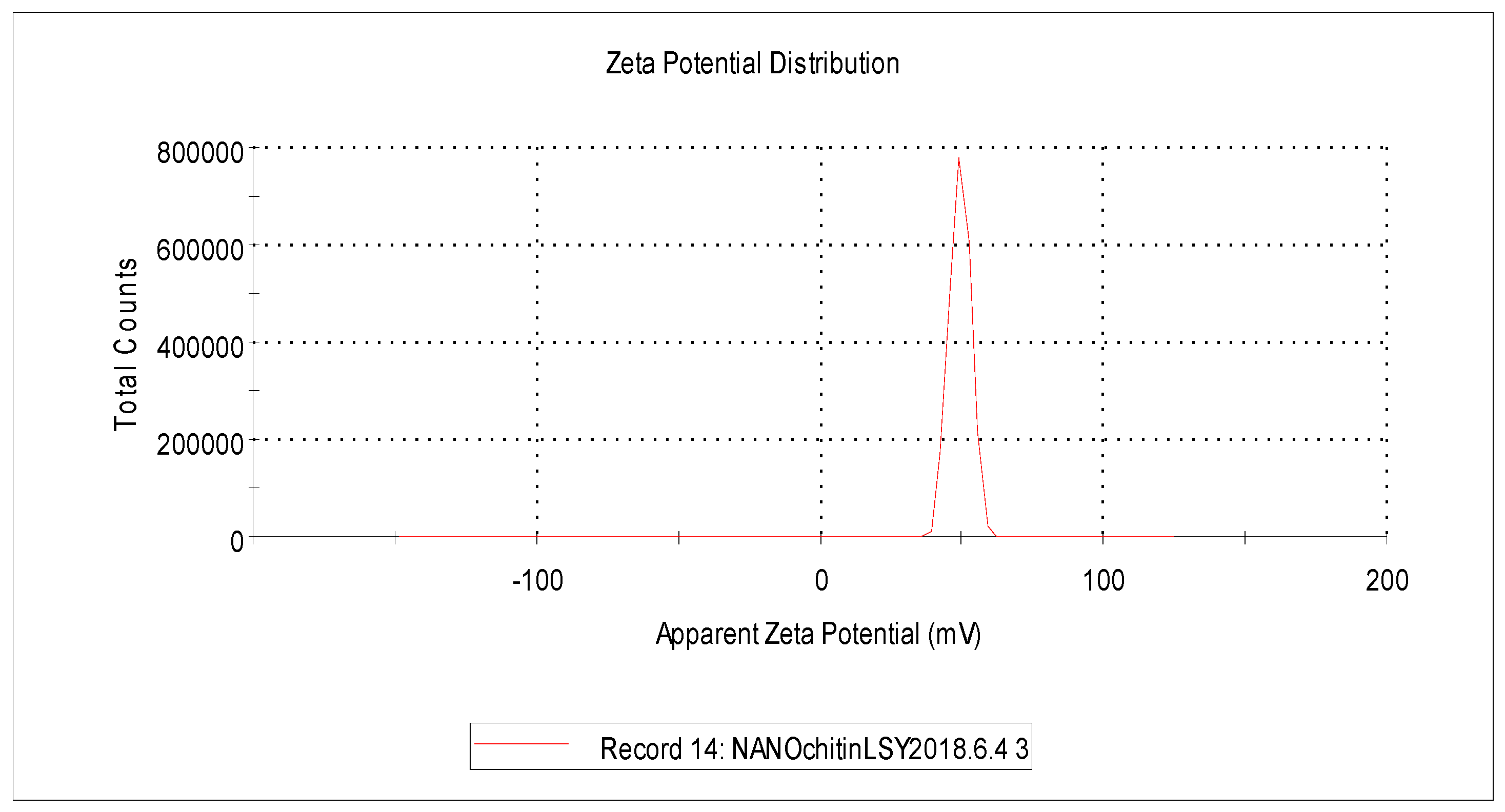
2.1. Characteristics of Nanochitin Particle
2.2. Effects of Nanochitin on Grain Yield and Crude Protein Concentration
2.3. Enhancement of Dry Matter and N Accumulation by Nanochitin
2.4. Improvement in the Retranslocation of Dry Matter and N from Pre-Anthesis Vegetation Organs to Grains
2.5. Effects of Nanochitin on Contributions of Dry Matter and N to Grain
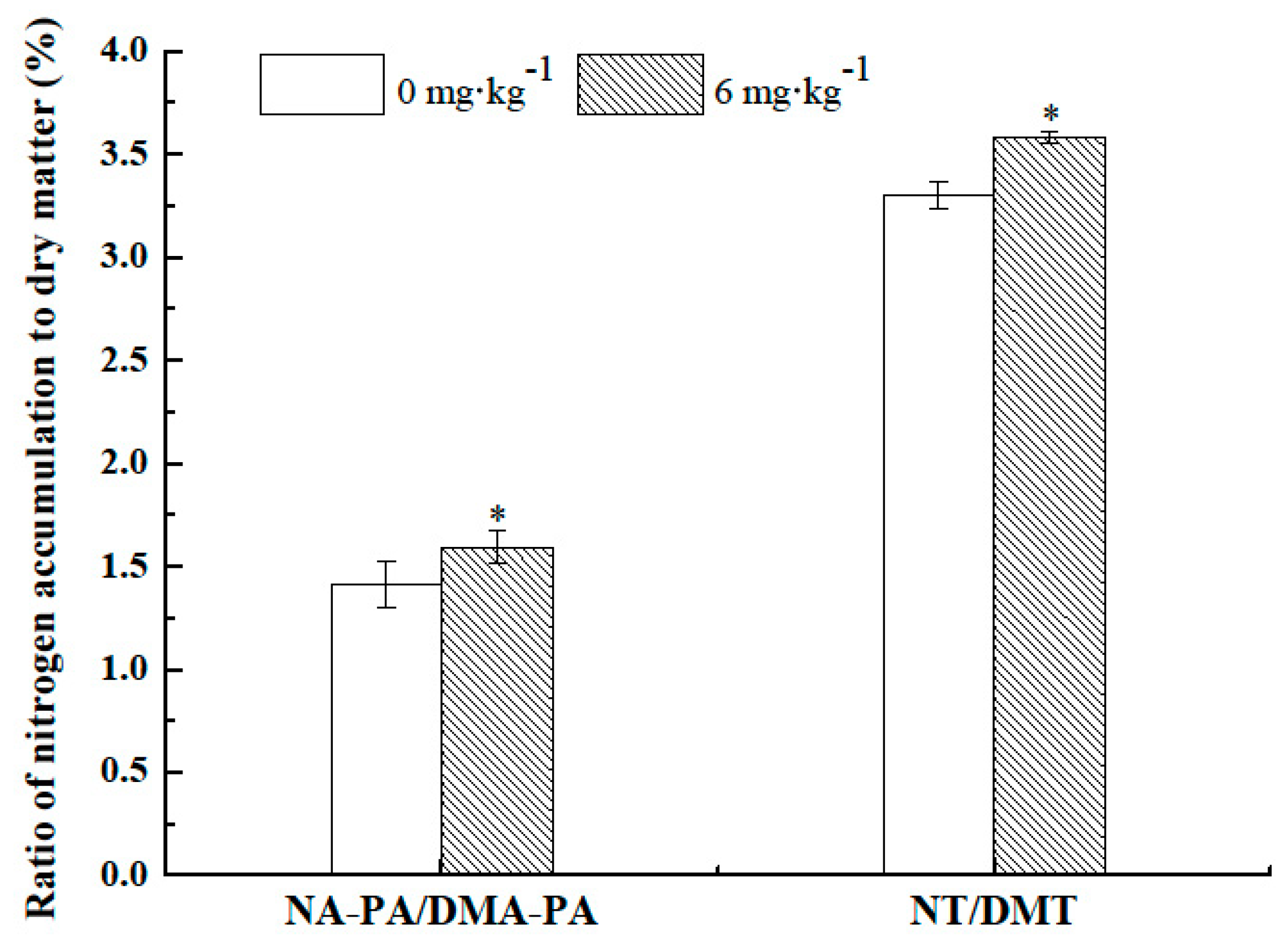
2.6. Effects of Nanochitin on the Ratio of N to Dry Matter Accumulation and Translocation
2.7. Regulation of Carbon and Nitrogen Metabolism
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation and Characterization of Nanochitin
4.3. Plant Materials and Treatments
4.4. Fresh Sample Treatments and Assay of Enzymes
4.5. Calculations of Relevant Indicators
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coruzzi, G.; Bush, D.R. Nitrogen and carbon nutrient and metabolite signaling in plants. Plant Physiol. 2001, 125, 61–64. [Google Scholar] [CrossRef]
- Wang, L.H.; Cao, X.B.; Zuo, S.Y.; Wei, S.; Li, J. Differences in flag leaf nitrogen metabolism and grain protein formation among triticale varieties after anthesis (In Chinese). Plant Physiol. J. 2018, 54, 401–410. [Google Scholar] [CrossRef]
- Tian, Z.W.; Wang, F.R.; Dai, T.B.; Cai, J.; Jiang, D.; Cao, W.X. Characteristics of dry matter accumulation and translocation during the wheat genetic improvement and their relationship to grain yield (In Chinese). Sci. Agric. Sin. 2012, 45, 801–808. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Shi, W.; Kronzucker, H.J. Nitrogen transformations in modern agriculture and the role of biological nitrification inhibition. Nat. Plants 2017, 3, 17074. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhao, C.P.; Cao, C.X.; Jiang, Q.J.; Jiang, H.D. Effects of N dressing ratio on carbon and nitrogen transport and on grain yield of Lolium multiflorum. Acta Prataculturae Sin. 2010, 19, 47–53. [Google Scholar]
- Zhang, L.; Lü, J.-Y.; Jia, S.-L. Photosynthetic characteristics of spike and distribution of 14C-assimilates accumulated before anthesis in wheat under water deficit condition. Acta Agron. Sin. 2013, 39, 1514–1519. [Google Scholar] [CrossRef]
- Tian, X.-Y.; Liu, X.; Wang, Q.-S.; Jiang, Q.; Feng, J.-X.; Zhang, H.; Ding, Y.-F. Effects of brassinosteroids on photosynthetic matter, nitrogen accumulation and use efficiency during grain filling stage of hybridJaponica(in Chinese). Acta Agron. Sin. 2015, 41, 1844–1857. [Google Scholar] [CrossRef]
- Popko, M.; Michalak, I.; Wilk, R.; Gramza, M.; Chojnacka, K.; Górecki, H. Effect of the new plant growth biostimulants based on amino acids on yield and grain quality of winter wheat. Molecules 2018, 23, 470. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Liu, M.; Liu, Z.; Yang, H.; Sun, X.; Chen, J.; Xiang, S.; Ding, W. MgONPs can boost plant growth: Evidence from increased seedling growth, morpho-physiological activities, and Mg uptake in tobacco (Nicotiana tabacum L.). Molecules 2018, 23, 3375. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, P.; Priyanka, N.; Manikandan, K.; Ganeshbabu, I.; Indiraarulselvi, P.; Geetha, N.; Muralikrishna, K.; Bhattacharya, R.C.; Tiwari, M.; Sharma, N.; et al. Enhanced plant growth promoting role of phycomolecules coated zinc oxide nanoparticles with P supplementation in cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2017, 110, 118–127. [Google Scholar] [CrossRef]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; AlMutairi, K.A.; Siddiqui, Z.H. Role of nanomaterials in plants under challenging environments. Plant Physiol. Biochem. 2017, 110, 194–209. [Google Scholar] [CrossRef]
- Shinano, T.; Nakajima, K.; Wasaki, J.; Mori, H.; Zheng, T.; Osaki, M. Developmental regulation of photosynthate distribution in leaves of rice. Photosynthetica 2006, 44, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Németh, E.; Nagy, Z.; Pécsváradi, A. Chloroplast glutamine synthetase, the key regulator of nitrogen metabolism in wheat, performs its role by fine regulation of enzyme activity via negative cooperativity of its subunits. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Foyer, C.H.; Ferrario-Méry, S.; Huber, S.C. Regulation of carbon fluxes in the cytosol: Coordination of sucrose synthesis, nitrate reduction and organic acid and amino acid biosynthesis. In Photosynthesis: Physiology and Metabolism; Leegood, R.C., Sharkey, T.D., von Caemmerer, S., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 177–203. [Google Scholar]
- Weiner, H.; McMichael, R.W.; Huber, S.C. Identification of factors regulating the phosphorylation status of sucrose-phosphate synthase in Vivo. Plant Physiol. 1992, 99, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef] [PubMed]
- Rendina, N.; Nuzzaci, M.; Scopa, A.; Cuypers, A.; Sofo, A. Chitosan-elicited defense responses in Cucumber mosaic virus (CMV)-infected tomato plants. J. Plant Physiol. 2019, 234–235, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Li, X.; Yuan, W.; Jin, S.; Hou, S.; Wang, M.; Wang, H. Antifungal activity of nanochitin whisker against crown rot diseases of wheat. J. Agric. Food Chem. 2018, 66, 9907–9913. [Google Scholar] [CrossRef]
- Zhang, X.; Li, K.; Xing, R.; Liu, S.; Li, P. Metabolite profiling of wheat seedlings induced by chitosan: Revelation of the enhanced carbon and nitrogen metabolism. Frontiers plant sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, H.M.M.; Hasaneen, M.N.A.; Omer, A.M. Nano chitosan-NPK fertilizer enhances the growth and productivity of wheat plants grown in sandy soil. Span. J. Agric. Res. 2016, 14, e0902–e0909. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; EI Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Y.; Zhang, X.; Merewitz, E.; Peng, Y.; Ma, X.; Huang, L.; Yan, Y. Metabolic pathways regulated by chitosan contributing to drought resistance in white clover. J. Proteome Res. 2017, 16, 3039–3052. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Luo, X.; Tu, R. Application of bioactive coatings based on chitosan for soybean seed protection. Int. J. Carbohydr. Chem. 2012, 2012. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Escudero, N.; Zavala-Gonzalez, E.A.; Esteve-Bruna, D.; Blázquez, M.A.; Alabadí, D.; Lopez-Llorca, L.V. Induction of auxin biosynthesis and WOX5 repression mediate changes in root development in Arabidopsis exposed to chitosan. Sci. Rep. 2017, 7, 16813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources: Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, K.; Liu, S.; Xing, R.; Yu, H.; Chen, X.; Li, P. Size effects of chitooligomers on the growth and photosynthetic characteristics of wheat seedlings. Carbohydr. Polym. 2016, 138, 27–33. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, S.; Jiao, Y.; Wang, H. Synergistic effects of nanochitin on inhibition of tobacco root rot disease. Int. J. Biol. Macromol. 2017, 99, 205–212. [Google Scholar] [CrossRef]
- Xue, W.; Han, Y.; Tan, J.; Wang, Y.; Wang, G.; Wang, H. Effects of nanochitin on enhancement of grain yield and quality of winter wheat. J. Agric. Food Chem. 2018, 66, 6637–6645. [Google Scholar] [CrossRef]
- Domingos, R.F.; Baalousha, M.A.; Ju-Nam, Y.; Reid, M.M.; Tufenkji, N.; Lead, J.R.; Leppard, G.G.; Wilkinson, K.J. Characterizing Manufactured Nanoparticles in the Environment: Multimethod Determination of Particle Sizes. Environ. Sci. Technol. 2009, 43, 7277–7284. [Google Scholar] [CrossRef]
- Doane, T.L.; Chuang, C.-H.; Hill, R.J.; Burda, C. Nanoparticle ζ -Potentials. Acc. Chem. Res. 2012, 45, 317–326. [Google Scholar] [CrossRef]
- Irving, L.J.; Vaughan, J.K.E.; Ong, G.; Schwier, N.; Hama, T.; Cameron, D.D. Differential carbon allocation to nitrogen-rich patches in Poa annua precedes root proliferation but has no immediate benefit to N uptake. J. Plant Physiol. 2019, 234–235, 54–59. [Google Scholar] [CrossRef]
- Britto, D.T.; Wilhelm, C.; Kronzucker, H.J. From biochemical pathways to the agro-ecological scale: Carbon capture in a changing climate. J. Plant Physiol. 2016, 203, 1–2. [Google Scholar] [CrossRef]
- Sharp, R.G. A review of the applications of chitin and Its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 2013, 3, 757–793. [Google Scholar] [CrossRef]
- Gooday, G.W. The ecology of chitin degradation. In Advances in Microbial Ecology; Marshall, K.C., Ed.; Springer: Boston, MA, USA, 1990. [Google Scholar]
- Wieczorek, A.S.; Hetz, S.A.; Kolb, S. Microbial responses to chitin and chitosan in oxic and anoxic agricultural soil slurries. Biogeosciences 2014, 11, 3339–3352. [Google Scholar] [CrossRef]
- Winkler, A.J.; Dominguez-Nuñez, J.A.; Aranaz, I.; Poza-Carrión, C.; Ramonell, K.; Somerville, S.; Berrocal-Lobo, M. Short-chain chitin oligomers: Promoters of plant growth. Mar. Drugs 2017, 15. [Google Scholar] [CrossRef]
- Fan, W.; Yan, W.; Xu, Z.; Ni, H. Formation mechanism of monodisperse, low molecular weight chitosan nanoparticles by ionic gelation technique. Colloids and Surfaces B: Biointerfaces 2012, 90, 21–27. [Google Scholar] [CrossRef]
- Iavicoli, I.; Leso, V.; Fontana, L.; Calabrese, E. Nanoparticle exposure and hormetic dose–responses: An update. Int. J. Mol. Sci. 2018, 19, 805. [Google Scholar] [CrossRef]
- Spiegel, Y.; Kafkafi, U.; Pressman, E. Evaluation of a protein-chitin derivative of crustacean shells as a slow-release nitrogen fertilizer on Chinese cabbage. J. Hortic. Sci. 1988, 63, 621–627. [Google Scholar] [CrossRef]
- Roberts, P.; Jones, D.L. Microbial and plant uptake of free amino sugars in grassland soils. Soil Biol. Biochem. 2012, 49, 139–149. [Google Scholar] [CrossRef]
- Li, X.; Zhou, L.; Liu, F.; Zhou, Q.; Cai, J.; Wang, X.; Dai, T.; Cao, W.; Jiang, D. Variations in protein concentration and nitrogen sources in different positions of grain in wheat. Frontiers Plant Sci. 2016, 7, 00942. [Google Scholar] [CrossRef]
- Shi, Z.; Li, D.; Jing, Q.; Cai, J.; Jiang, D.; Cao, W.; Dai, T. Effects of nitrogen applications on soil nitrogen balance and nitrogen utilization of winter wheat in a rice–wheat rotation. Field Crops Res. 2012, 127, 241–247. [Google Scholar] [CrossRef]
- Kong, L.; Xie, Y.; Hu, L.; Feng, B.; Li, S. Remobilization of vegetative nitrogen to developing grain in wheat (Triticum aestivum L.). Field Crops Res. 2016, 196, 134–144. [Google Scholar] [CrossRef]
- Dang, H.-k.; Li, R.-q.; Li, Y.-m.; Sun, Y.-h.; Zhang, X.-w.; Liu, M.-x. Absorption, accumulation and distribution of nitrogen in super-highly yielding winter wheat (in Chinese). J. Plant Nutr. Fertilizer 2013, 19, 1037–1047. [Google Scholar]
- Li, S.; Huang, Y.; An, Z.; Ye, Y. Effect of different cultivation modes on accumulation and translocation of dry matter and nitrogen after anthesis of winter wheat (in Chinese). J. Triticeae Crops 2017, 5, 687–693. [Google Scholar]
- Simmonds, N.W. The relation between yield and protein in cereal grain. J. Sci. Food Agric. 1995, 67, 309–315. [Google Scholar] [CrossRef]
- Kang, J.; Turano, F.J. The putative glutamate receptor 1.1 (AtGLR1.1) functions as a regulator of carbon and nitrogen metabolism in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6872–6877. [Google Scholar] [CrossRef]
- Bao, A.; Zhao, Z.; Ding, G.; Shi, L.; Xu, F.; Cai, H. The stable level of Glutamine synthetase 2 plays an important role in rice growth and in carbon-nitrogen metabolic balance. Int. J. Mol. Sci. 2015, 16, 12713–12736. [Google Scholar] [CrossRef]
- Yu, J.-l.; Song, H.-x.; Xie, G.-x.; Zhang, Z.-h.; Liao, Q.; Guan, C.-y. Different activities of SPS and PEPC in oilseed rape with different nitrogen use efficiency and their influence on seed yield and oil-content (in Chinese). J. Plant Nutr. Fertilizer 2016, 22, 618–625. [Google Scholar]
- Wang, X.; Huang, J. Principles and Techniques of Plant Physiology and Biochemistry Experiment, 3rd ed.; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Chen, M.; Gao, Z.; Sun, M.; Ren, A.; Deng, Y.; University, S.A. Effect of phosphorus application under tillage in fallow period on nitrogen absorption and translocation in dryland wheat (in Chinese). J. Triticeae Crops 2015, 35, 1569–1575. [Google Scholar]
Sample Availability: Samples of the nanochitin whisker are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CNanochitin* (mg·kg−1) | Number of Spike | Grains/Spike | Kernel Weight | Yield | Crude Protein Content (g·kg−1) |
|---|---|---|---|---|---|
| (number·pot−1) | (number·ear−1) | (g·1000 grain−1) | (g·pot−1) | ||
| 0 | 31 ± 1.53b** | 30 ± 0.81b | 34.35 ± 0.22c | 27.43 ± 0.57c | 142.18 ± 6.44b |
| 2 | 34 ± 0.58a | 31 ± 0.31a | 36.29 ± 0.59b | 32.68 ± 0.83b | 156.28 ± 4.44a |
| 6 | 35 ± 1.00a | 32 ± 0.72a | 37.32 ± 0.15a | 34.99 ± 0.08a | 161.03 ± 3.15a |
| 20 | 30 ± 1.00b | 29 ± 0.67b | 34.88 ± 0.51c | 26.26 ± 0.68d | 138.46 ± 7.75b |
| CNanochitin* (mg·kg−1) | DMA-A (g·pot−1) | DMA-M (g·pot−1) | DMA-PA (g·pot−1) | NA-A (g·pot−1) | NA-M (g·pot−1) | NA-PA (g·pot−1) |
|---|---|---|---|---|---|---|
| 0 | 77.05 ± 2.30c** | 91.00 ± 2.58c | 13.94 ± 0.47c | 1.03 ± 0.07c | 1.23 ± 0.09c | 0.20 ± 0.02b |
| 2 | 85.70 ± 2.04b | 100.71 ± 1.97b | 15.01 ± 0.44b | 1.30 ± 0.06b | 1.54 ± 0.07b | 0.24 ± 0.02a |
| 6 | 89.49 ± 2.57a | 105.57 ± 2.45a | 16.08 ± 0.23a | 1.43 ± 0.06a | 1.69 ± 0.07a | 0.26 ± 0.01a |
| 20 | 72.47 ± 1.96c | 85.18 ± 1.89d | 12.71 ± 0.29d | 0.92 ± 0.05c | 1.09 ± 0.07c | 0.17 ± 0.02c |
| CNanochitin* (mg·kg−1) | DMA-VO-M (g·pot−1) | DMT (g·pot−1) | DMTP (%) | NA-VO-M (g·pot−1) | NT (g·pot−1) | NTP (%) |
|---|---|---|---|---|---|---|
| 0 | 60.31 ± 0.95c** | 16.74 ± 1.37c | 21.70 ± 1.15b | 0.48 ± 0.01c | 0.55 ± 0.05b | 53.70 ± 1.82b |
| 2 | 63.75 ± 1.02b | 21.95 ± 1.21b | 25.60 ± 0.86a | 0.55 ± 0.02b | 0.75 ± 0.04a | 57.77 ± 0.68a |
| 6 | 66.33 ± 1.50a | 23.16 ± 1.08a | 25.87 ± 0.47a | 0.60 ± 0.02a | 0.83 ± 0.04a | 57.87 ± 0.37a |
| 20 | 58.26 ± 1.12c | 14.21 ± 0.93c | 19.60 ± 0.81c | 0.45 ± 0.01c | 0.47 ± 0.03c | 50.58 ± 1.18c |
| CNanochitin* (mg·kg−1) | CR-DMT | CR-DMA | CR-NT | CR-NA |
|---|---|---|---|---|
| (%) | (%) | (%) | (%) | |
| 0 | 54.51 ± 1.88b** | 45.49 ± 1.88a | 73.73 ± 1.20b | 26.27 ± 1.20a |
| 2 | 59.36 ± 1.82a | 40.64 ± 1.82b | 75.89 ± 1.00a | 24.11 ± 1.00b |
| 6 | 59.00 ± 1.37a | 41.00 ± 1.37b | 76.45 ± 0.72a | 23.55 ± 0.72b |
| 20 | 52.76 ± 2.06b | 47.24 ± 2.06a | 72.84 ± 0.84b | 27.16 ± 0.84a |
| Organ | CNanochitin* (mg·kg−1) | Anthesis Stage | 15-Days After Anthesis | ||||
|---|---|---|---|---|---|---|---|
| SPS | PEPC | SPS/PEPC | SPS | PEPC | SPS/PEPC | ||
| (mg/g·h) | (nmol/g·min) | (mg/g·h) | (nmol/g·min) | ||||
| Flag Leaf | 0 | 7.27 ± 0.58b** | 10.16 ± 1.09b | 0.72 ± 0.04a | 4.61 ± 0.92b | 5.99 ± 0.83b | 0.77 ± 0.05a |
| 6 | 10.16 ± 0.89a | 15.96 ± 0.74a | 0.64 ± 0.03b | 5.91 ± 0.43a | 8.69 ± 1.12a | 0.68 ± 0.05b | |
| Spike | 0 | 5.81 ± 1.05b | 12.23 ± 1.29b | 0.48 ± 0.02a | 2.96 ± 0.19b | 4.14 ± 0.81b | 0.71 ± 0.04a |
| 6 | 7.90 ± 1.09a | 19.42 ± 1.93a | 0.41 ± 0.01b | 4.21 ± 0.35a | 6.48 ± 0.61a | 0.65 ± 0.06b | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.; Wang, Y.; Han, Y.; Li, D.; Zhang, Z.; Zhu, X.; Tan, J.; Wang, H. The Stimulatory Effects of Nanochitin Whisker on Carbon and Nitrogen Metabolism and on the Enhancement of Grain Yield and Crude Protein of Winter Wheat. Molecules 2019, 24, 1752. https://doi.org/10.3390/molecules24091752
Cheng Y, Wang Y, Han Y, Li D, Zhang Z, Zhu X, Tan J, Wang H. The Stimulatory Effects of Nanochitin Whisker on Carbon and Nitrogen Metabolism and on the Enhancement of Grain Yield and Crude Protein of Winter Wheat. Molecules. 2019; 24(9):1752. https://doi.org/10.3390/molecules24091752
Chicago/Turabian StyleCheng, Yingying, Yi Wang, Yanlai Han, Dongya Li, Zhongkui Zhang, Xueqiang Zhu, Jinfang Tan, and Hezhong Wang. 2019. "The Stimulatory Effects of Nanochitin Whisker on Carbon and Nitrogen Metabolism and on the Enhancement of Grain Yield and Crude Protein of Winter Wheat" Molecules 24, no. 9: 1752. https://doi.org/10.3390/molecules24091752