An Investigation on Glucuronidation Metabolite Identification, Isozyme Contribution, and Species Differences of GL-V9 In Vitro and In Vivo

Abstract
:
1. Introduction
2. Results
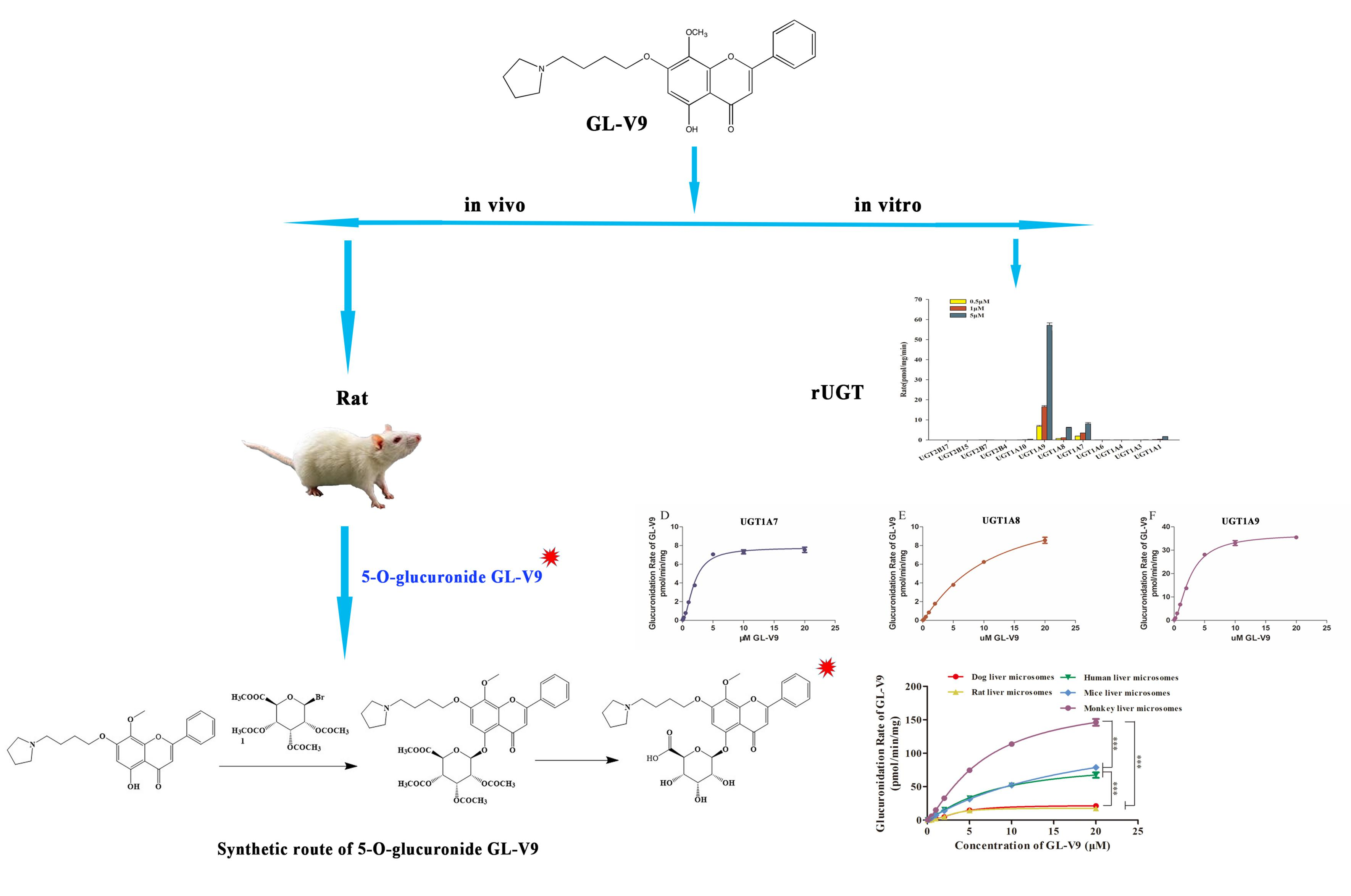
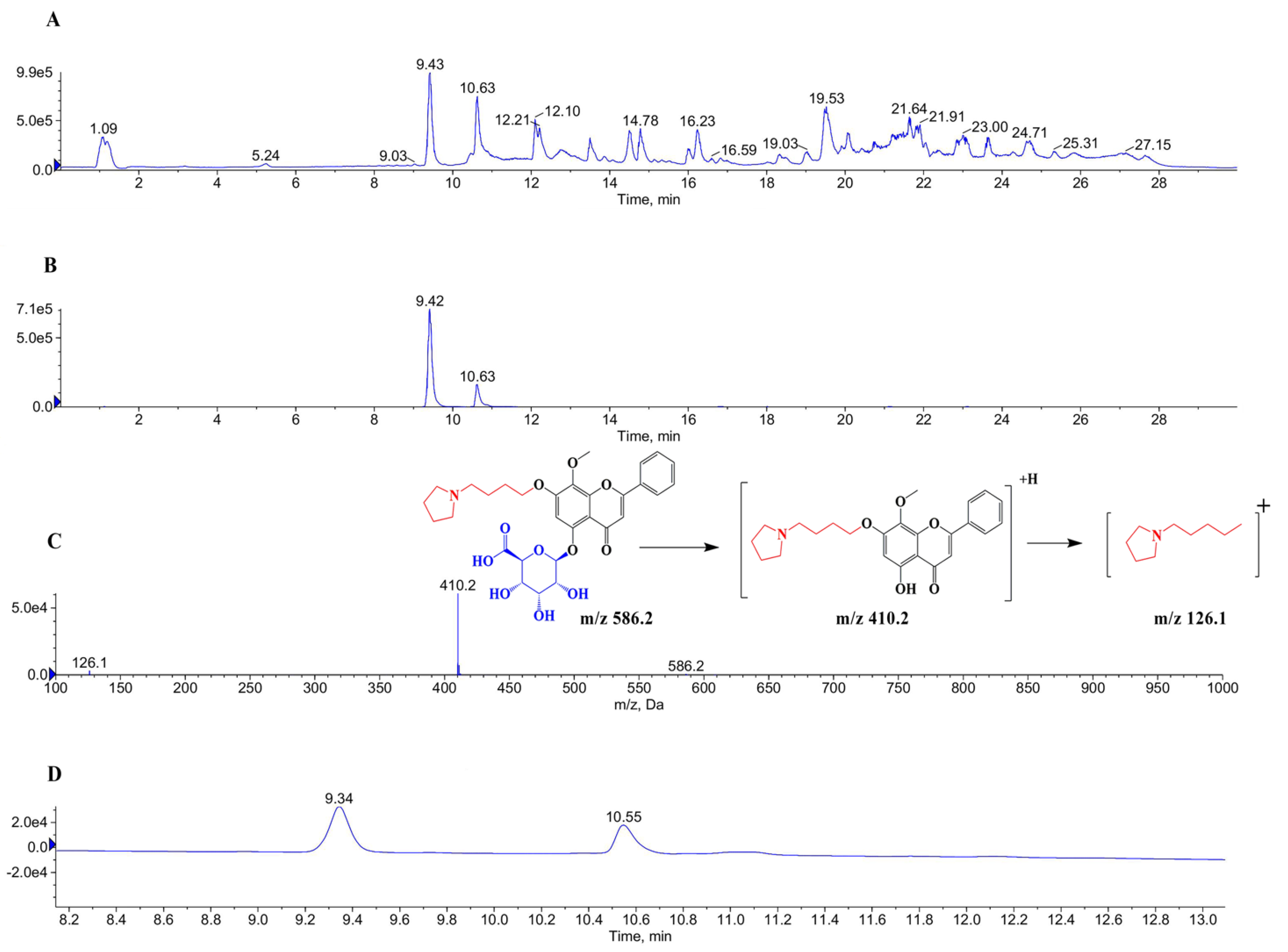
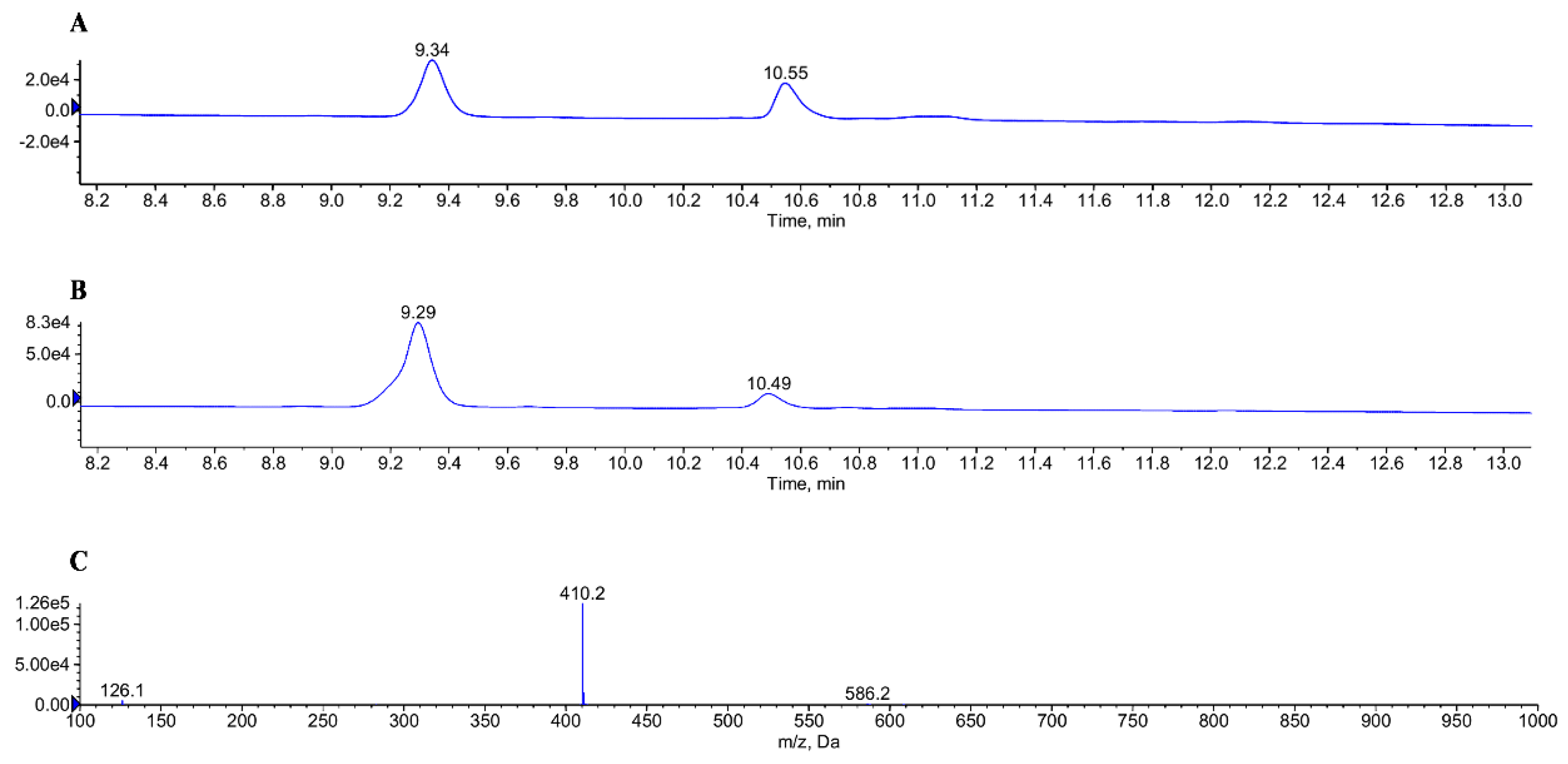
2.1. Identification of Glucuronidation of GL-V9 In Vitro and In Vivo
2.2. Chemical Synthesis of 5-O-Glucuronide GL-V9 and 1H-NMR Analysis
2.3. Main UGT Isoforms Responsible for the Metabolism of GL-V9 In Vitro
2.4. Kinetics of GL-V9 in Liver Microsomes and Recombinant UGTs (1A7, 1A8 and 1A9)
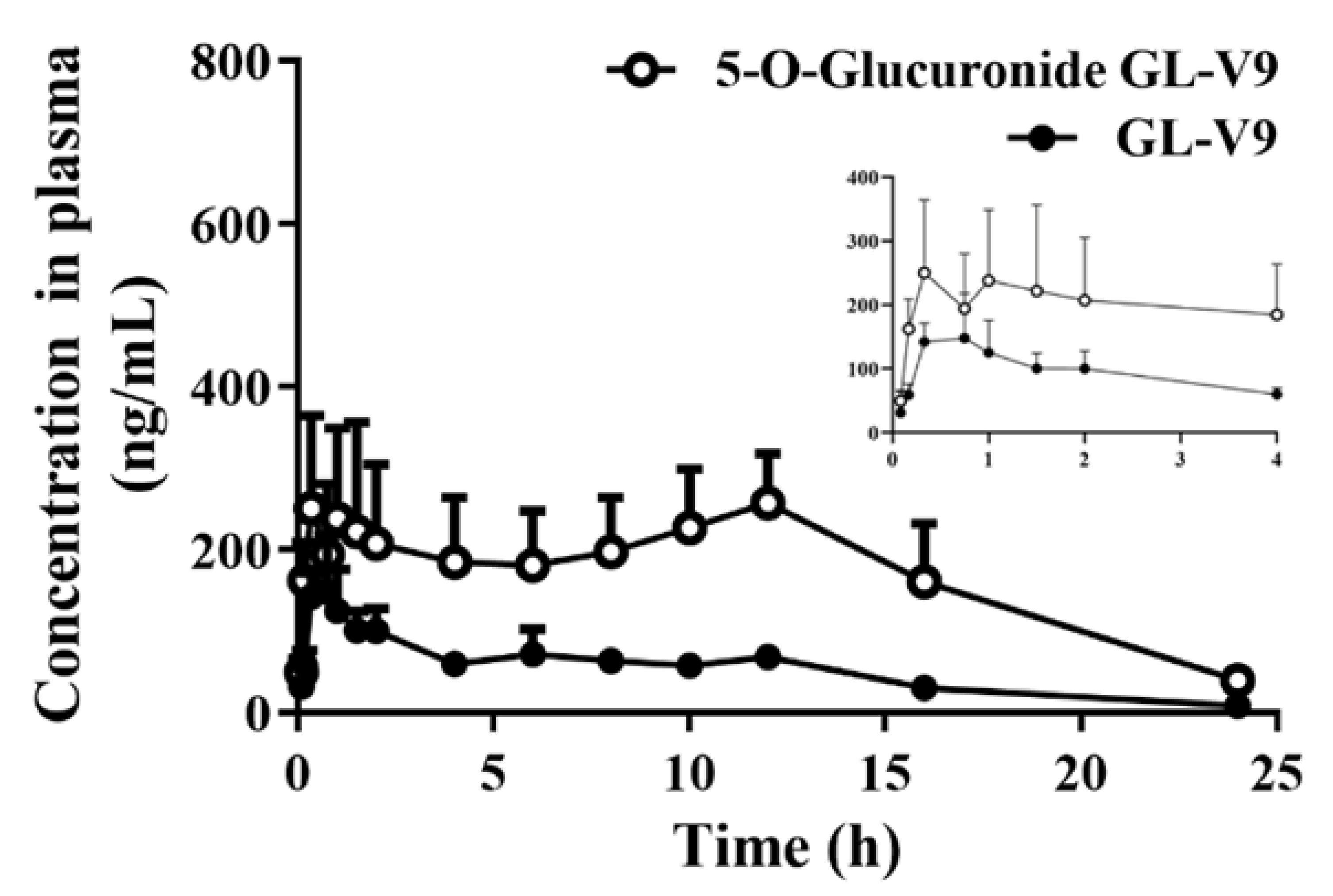
2.5. Quantification of GL-V9 and 5-O-glucuronide GL-V9 In Vivo
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagents
4.2. Animals
4.3. Identification of Glucuronidation of GL-V9 In Vivo and In Vitro
4.4. Chemical Synthesis of 5-O-glucuronide GL-V9 and NMR Analysis
4.5. Glucuronidation of GL-V9 in Monkey, Beagle Dog, Rat, Mouse and Human Liver Microsomes
4.6. Quantification of GL-V9 and Its Metabolites in Rat Plasma
4.7. UGT Phenotyping and Enzymes Kinetic Studies of rUGTs (rUGT1A1, 1A7, 1A8, 1A9)
4.8. Analytical Conditions
4.8.1. HPLC-UV-TripleTOF Methods
4.8.2. UPLC-MS/MS Methods
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiang, N.; Doseff, A.; Grotewold, E. Flavones: From Biosynthesis to Health Benefits. Plants 2016, 5, 27. [Google Scholar] [CrossRef]
- Middleton, E. Effect of Plant Flavonoids on Immune and Inflammatory Cell Function. In Flavonoids in the Living System; Manthey, J.A., Buslig, B.S., Eds.; Springer: Boston, MA, USA, 1998; Volume 439, pp. 175–182. ISBN 978-1-4613-7434-3. [Google Scholar]
- Pyrzynska, K.; Biesaga, M. Analysis of phenolic acids and flavonoids in honey. TrAC Trends Anal. Chem. 2009, 28, 893–902. [Google Scholar] [CrossRef]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Singh, P.; Jain, S.; Bhala, M.; Goyal, R.B.; Jayaprakash, D.; Lohiya, N.K. Wogonin, 5,7-dihydroxy-8-methoxyflavone as oestrogenic and anti-implantational agent in the rat. Phytother. Res. 1990, 4, 86–89. [Google Scholar] [CrossRef]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Rice-evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The Relative Antioxidant Activities of Plant-Derived Polyphenolic Flavonoids. Free Radic. Res. 1995, 22, 375–383. [Google Scholar] [CrossRef]
- Chan, S.-C.; Chang, Y.-S.; Wang, J.-P.; Chen, S.-C.; Kuo, S.-C. Three New Flavonoids and Antiallergic, Anti-Inflammatory Constituents from the Heartwood of Dalbergia odorifera. Planta Med. 1998, 64, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Sugimoto, Y.; Masuda, H.; Kamei, C. Antiallergic Effect of Flavonoid Glycosides Obtained from Mentha piperita L. Biol. Pharm. Bull. 2002, 25, 256–259. [Google Scholar] [CrossRef]
- Nishihori, Y.; Kato, K.; Tanaka, M.; Okamoto, T.; Hagiwara, S.; Araki, N.; Kogawa, K.; Kuribayashi, K.; Nakamura, K.; Niitsu, Y. Anti-tumour effects of nobiletin, a citrus flavonoid, on gastric cancer include: Antiproliferative effects, induction of apoptosis and cell cycle deregulation. Aliment. Pharmacol. Ther. 2004, 20, 95–101. [Google Scholar]
- Sharma, N.; Dobhal, M.; Joshi, Y.; Chahar, M. Flavonoids: A versatile source of anticancer drugs. Pharmacogn. Rev. 2011, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Desideri, G.; Croce, G.; Tiberti, S.; Aggio, A.; Ferri, C. Flavonoids, Vascular Function and Cardiovascular Protection. Curr. Pharm. Des. 2009, 15, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Folts, J.D. Potential Health Benefits from the Flavonoids in Grape Products on Vascular Disease. In Flavonoids in Cell Function; Buslig, B.S., Manthey, J.A., Eds.; Springer: Boston, MA, USA, 2002; Volume 505, pp. 95–111. ISBN 978-1-4419-3383-6. [Google Scholar]
- Talbi, A.; Zhao, D.; Liu, Q.; Li, J.; Fan, A.; Yang, W.; Han, X.; Chen, X. Pharmacokinetics, Tissue Distribution, Excretion and Plasma Protein Binding Studies of Wogonin in Rats. Molecules 2014, 19, 5538–5549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Lu, N.; Dai, Q.; Wei, L.; Zhao, Q.; Li, Z.; He, Q.; Dai, Y.; Guo, Q. GL-V9, a newly synthetic flavonoid derivative, induces mitochondrial-mediated apoptosis and G2/M cell cycle arrest in human hepatocellular carcinoma HepG2 cells. Eur. J. Pharmacol. 2011, 670, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Li, W.; Miao, H.; Yao, J.; Li, Z.; Wei, L.; Zhao, L.; Guo, Q. LW-214, a newly synthesized flavonoid, induces intrinsic apoptosis pathway by down-regulating Trx-1 in MCF-7 human breast cells. Biochem. Pharmacol. 2014, 87, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Liu, W.; Nie, F.; Zhao, Q.; Rong, J.; Wang, J.; Tao, L.; Qi, Q.; Lu, N.; Li, Z.; et al. LYG-202, a new flavonoid with a piperazine substitution, shows antitumor effects in vivo and in vitro. Biochem. Biophys. Res. Commun. 2009, 385, 551–556. [Google Scholar] [CrossRef]
- Tao, L.; Fu, R.; Wang, X.; Yao, J.; Zhou, Y.; Dai, Q.; Li, Z.; Lu, N.; Wang, W. LL-202, a newly synthesized flavonoid, inhibits tumor growth via inducing G2/M phase arrest and cell apoptosis in MCF-7 human breast cancer cells in vitro and in vivo. Toxicol. Lett. 2014, 228, 1–12. [Google Scholar] [CrossRef]
- Zhao, K.; Li, G.; Yao, Y.; Zhou, Y.; Li, Z.; Guo, Q.; Lu, N. Activation of phospholipase C-γ1 and translocation of phosphatidylinositol-3,4,5-trisphosphate 3-phosphatase contribute to GL-V9-induced apoptosis in human gastric cancer cells. Exp. Cell Res. 2017, 8–19. [Google Scholar] [CrossRef]
- Li, L.; Chen, P.; Ling, Y.; Song, X.; Lu, Z.; He, Q.; Li, Z.; Lu, N.; Guo, Q. Inhibitory effects of GL-V9 on the invasion of human breast carcinoma cells by downregulating the expression and activity of matrix metalloproteinase-2/9. Eur. J. Pharm. Sci. 2011, 43, 393–399. [Google Scholar] [CrossRef]
- Walle, T.; Otake, Y.; Brubaker, J.A.; Walle, U.K.; Halushka, P.V. Disposition and metabolism of the flavonoid chrysin in normal volunteers. Br. J. Clin. Pharmacol. 2001, 51, 143–146. [Google Scholar] [CrossRef] [Green Version]
- Claudine, M.; Donovan, J.L. Pharmacokinetics and metabolism of dietary flavonoids in humans. Free Radic. Res. 2004, 38, 771–785. [Google Scholar]
- Walle, T. Absorption and metabolism of flavonoids. Free Radic. Biol. Med. 2004, 36, 829–837. [Google Scholar] [CrossRef]
- Chow, H.-H.S.; Cai, Y.; Alberts, D.S.; Hakim, I.; Dorr, R.; Shahi, F.; Crowell, J.A.; Yang, C.S.; Hara, Y. Phase I pharmacokinetic study of teapolyphenols following single-dose administration of epigallocatechin gallate and polyphenol E. Cancer Causes Control 2001, 10, 53–58. [Google Scholar]
- Walle, T. High Absorption but Very Low Bioavailability of Oral Resveratrol in Humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef]
- Otake, Y. Glucuronidation versus Oxidation of the Flavonoid Galangin by Human Liver Microsomes and Hepatocytes. Drug Metab. Dispos. 2002, 30, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Spencer, J.P.E.; Abd El Mohsen, M.M.; Rice-Evans, C. Cellular uptake and metabolism of flavonoids and their metabolites: Implications for their bioactivity. Arch. Biochem. Biophys. 2004, 423, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tu, M.; Cheng, C.; Tian, J.; Zhang, F.; Deng, Z.; Li, X.; Li, Z.; Liu, Y.; Lei, G. Wogonoside induces apoptosis in Bel-7402, a hepatocellular carcinoma cell line, by regulating Bax/Bcl-2. Oncol. Lett. 2015, 10, 1831–1835. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Xing, G.; Zhang, M.; Zhong, M.; Han, Z.; He, C.; Liu, X. Wogonoside inhibits cell growth and induces mitochondrial-mediated autophagy-related apoptosis in human colon cancer cells through the PI3K/AKT/mTOR/p70S6K signaling pathway. Oncol. Lett. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, H.; Cong, Z.; Xu, J.; Zhu, J.; Ji, X.; Ding, K. Wogonoside induces autophagy-related apoptosis in human glioblastoma cells. Oncol. Rep. 2014, 32, 1179–1187. [Google Scholar] [CrossRef]
- Tukey, R.H.; Strassburg, C.P. Human UDP-Glucuronosyltransferases: Metabolism, Expression, and Disease. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 581–616. [Google Scholar] [CrossRef] [PubMed]
- Kiang, T.; Ensom, M.; Chang, T. UDP-glucuronosyltransferases and clinical drug-drug interactions. Pharmacol. Ther. 2005, 106, 97–132. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Xie, R.; Ma, T.; Yan, W.; Pang, L. Probe substrate and enzyme source-dependent inhibition of UDP-glucuronosyltransferase (UGT) 1A9 by wogonin. Afr. Health Sci. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Singh, R.; Liu, Z.; Hu, M. Structure and Concentration Changes Affect Characterization of UGT Isoform-Specific Metabolism of Isoflavones. Mol. Pharm. 2009, 6, 1466–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, H.; Ren, C.; Kong, Y.; Ning, C.; Kong, D.; Zhang, Y.; Zhao, D.; Li, N.; Wang, Z.; Chen, X.; et al. Mechanistic study of absorption and first-pass metabolism of GL-V9, a derivative of wogonin. Biopharm. Drug Dispos. 2019. [Google Scholar] [CrossRef]
- Nakamura, A.; Nakajima, M.; Yamanaka, H.; Fujiwara, R.; Yokoi, T. Expression of UGT1A and UGT2B mRNA in Human Normal Tissues and Various Cell Lines. Drug Metab. Dispos. 2008, 36, 1461–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, S.; Nakajin, S. Determination of mRNA Expression of Human UDP-Glucuronosyltransferases and Application for Localization in Various Human Tissues by Real-Time Reverse Transcriptase-Polymerase Chain Reaction. Drug Metab. Dispos. 2009, 37, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Ren, C.; Kong, Y.; Ni, Q.; Wang, Z.; Zhao, D.; Li, N.; Chen, X.; Lu, Y. Determination of GL-V9, a derivative of wogonin, in rat plasma by UPLC-MS/MS and its application to a pharmacokinetic study after oral and pulmonary administration. Biomed. Chromatogr. 2019. [Google Scholar] [CrossRef]
- Tang, L.; Zhou, J.; Yang, C.-H.; Xia, B.-J.; Hu, M.; Liu, Z.-Q. Systematic Studies of Sulfation and Glucuronidation of 12 Flavonoids in the Mouse Liver S9 Fraction Reveal both Unique and Shared Positional Preferences. J. Agric. Food Chem. 2012, 60, 3223–3233. [Google Scholar] [CrossRef]
- Wong, Y.C.; Zhang, L.; Lin, G.; Zuo, Z. Structure–activity relationships of the glucuronidation of flavonoids by human glucuronosyltransferases. Expert Opin. Drug Metab. Toxicol. 2009, 5, 1399–1419. [Google Scholar] [CrossRef]
- Day, A.J.; Bao, Y.; Morgan, M.R.A.; Williamson, G. Conjugation position of quercetin glucuronides and effect on biological activity. Free Radic. Biol. Med. 2000, 29, 1234–1243. [Google Scholar] [CrossRef]
- Zhang, L.; Zuo, Z.; Lin, G. Intestinal and Hepatic Glucuronidation of Flavonoids. Mol. Pharm. 2007, 4, 833–845. [Google Scholar] [CrossRef]
- Li, C.; Zhang, L.; Lin, G.; Zuo, Z. Identification and quantification of baicalein, wogonin, oroxylin A and their major glucuronide conjugated metabolites in rat plasma after oral administration of Radix scutellariae product. J. Pharm. Biomed. Anal. 2011, 54, 750–758. [Google Scholar] [CrossRef]
- Yang, N.; Sun, R.; Liao, X.; Aa, J.; Wang, G. UDP-glucuronosyltransferases (UGTs) and their related metabolic cross-talk with internal homeostasis: A systematic review of UGT isoforms for precision medicine. Pharmacol. Res. 2017, 121, 169–183. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds GL-V9 and 5-O-glucuronide GL-V9 are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Monkey | Dog | Rat | Mice | Human |
|---|---|---|---|---|---|
| Concentration of protein (mg/mL) | 0.1 | 0.1 | 0.1 | 0.2 | 0.075 |
| Km (μM) | 11.19 ± 0.47 | 12.48 ± 1.43 | 6.54 ± 0.33 | 20.12 ± 1.04 * | 11.15 ± 0.66 |
| Vmax (pmol/min/mg) | 182.5 ± 4.92 * | 22.03 ± 0.42 | 32.72 ± 1.45 | 153.2 ± 10.44 | 88.49 ± 4.21 |
| Clint (Vmax/Km, μL/min/mg) | 16.31 * | 1.76 | 5.00 | 7.61 | 7.94 |
| Species | rUGT1A7 | rUGT1A8 | rUGT1A9 |
|---|---|---|---|
| Concentration of protein (mg/mL) | 0.05 | 0.06 | 0.06 |
| Km (μM) | 3.25 ± 0.29 | 13.92 ± 1.05 * | 4.72 ± 0.28 |
| Vmax (pmol/min/mg) | 31.15 ± 0.65 | 50.63 ± 4.09 | 148.1 ± 2.56 * |
| Clint (Vmax/Km, μL/min/mg) | 9.58 | 3.64 | 31.38 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, H.; Kong, D.; Ning, C.; Kong, Y.; Ren, C.; Cheng, Y.; Cai, H.; Wang, J.; Zhao, D.; Li, N.; et al. An Investigation on Glucuronidation Metabolite Identification, Isozyme Contribution, and Species Differences of GL-V9 In Vitro and In Vivo. Molecules 2019, 24, 1576. https://doi.org/10.3390/molecules24081576
Xing H, Kong D, Ning C, Kong Y, Ren C, Cheng Y, Cai H, Wang J, Zhao D, Li N, et al. An Investigation on Glucuronidation Metabolite Identification, Isozyme Contribution, and Species Differences of GL-V9 In Vitro and In Vivo. Molecules. 2019; 24(8):1576. https://doi.org/10.3390/molecules24081576
Chicago/Turabian StyleXing, Han, Dexuan Kong, Chen Ning, Ying Kong, Chang Ren, Yujie Cheng, Hui Cai, Jubo Wang, Di Zhao, Ning Li, and et al. 2019. "An Investigation on Glucuronidation Metabolite Identification, Isozyme Contribution, and Species Differences of GL-V9 In Vitro and In Vivo" Molecules 24, no. 8: 1576. https://doi.org/10.3390/molecules24081576