
Inhibition of Satellite RNA Associated Cucumber Mosaic Virus Infection by Essential Oil of Micromeria croatica (Pers.) Schott

 , ,
, ,  ,
,  , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
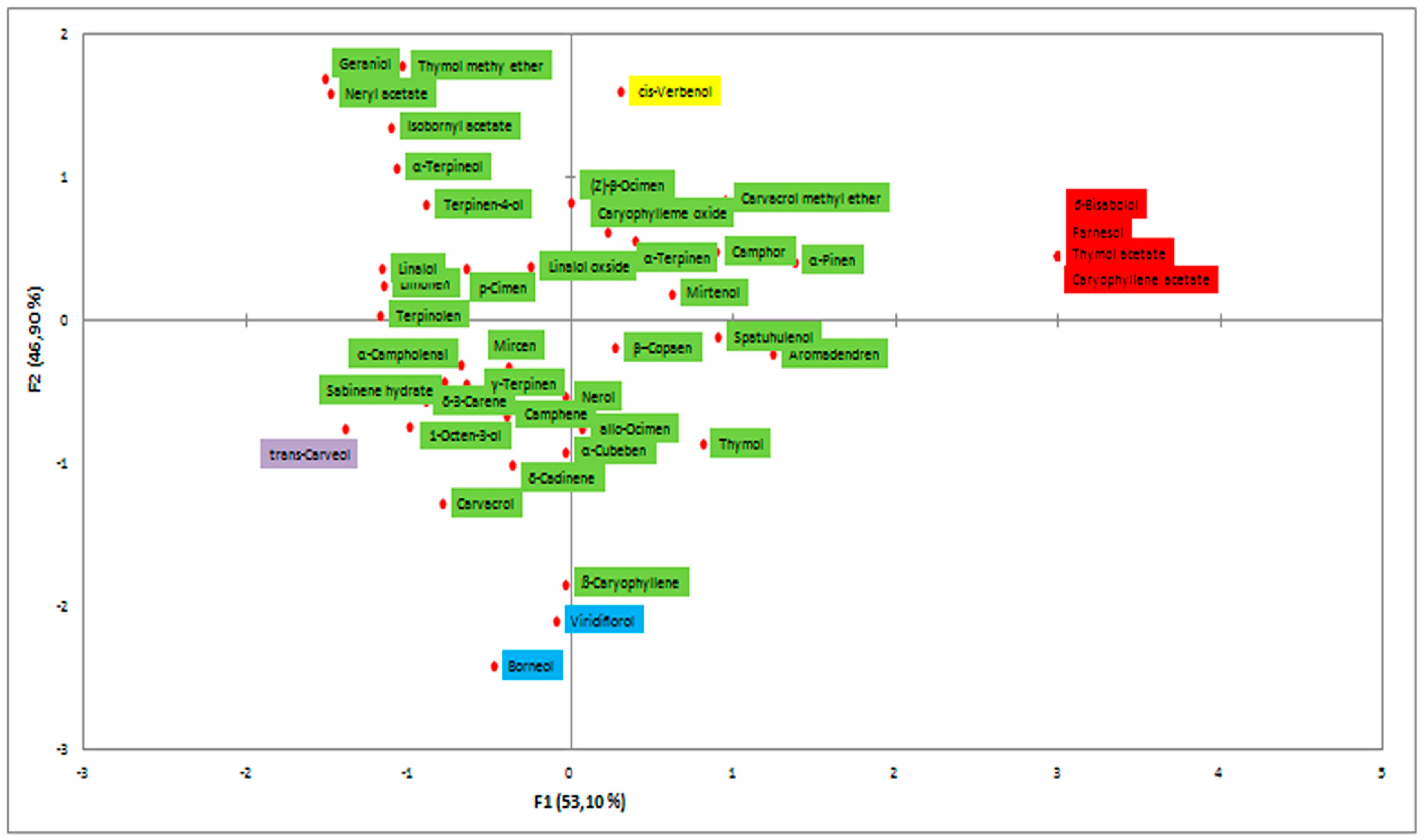
2.1. Essential Oil Composition
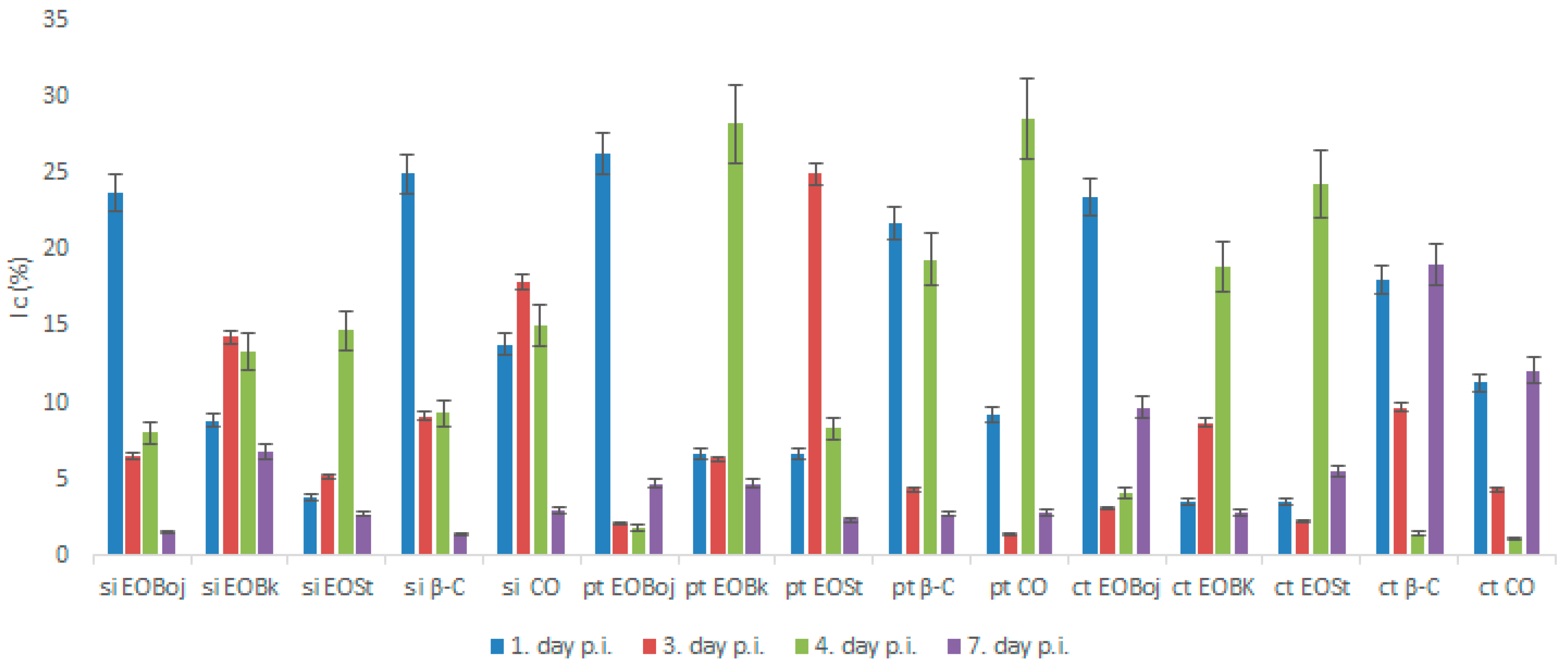
2.2. Antiphytoviral Activity
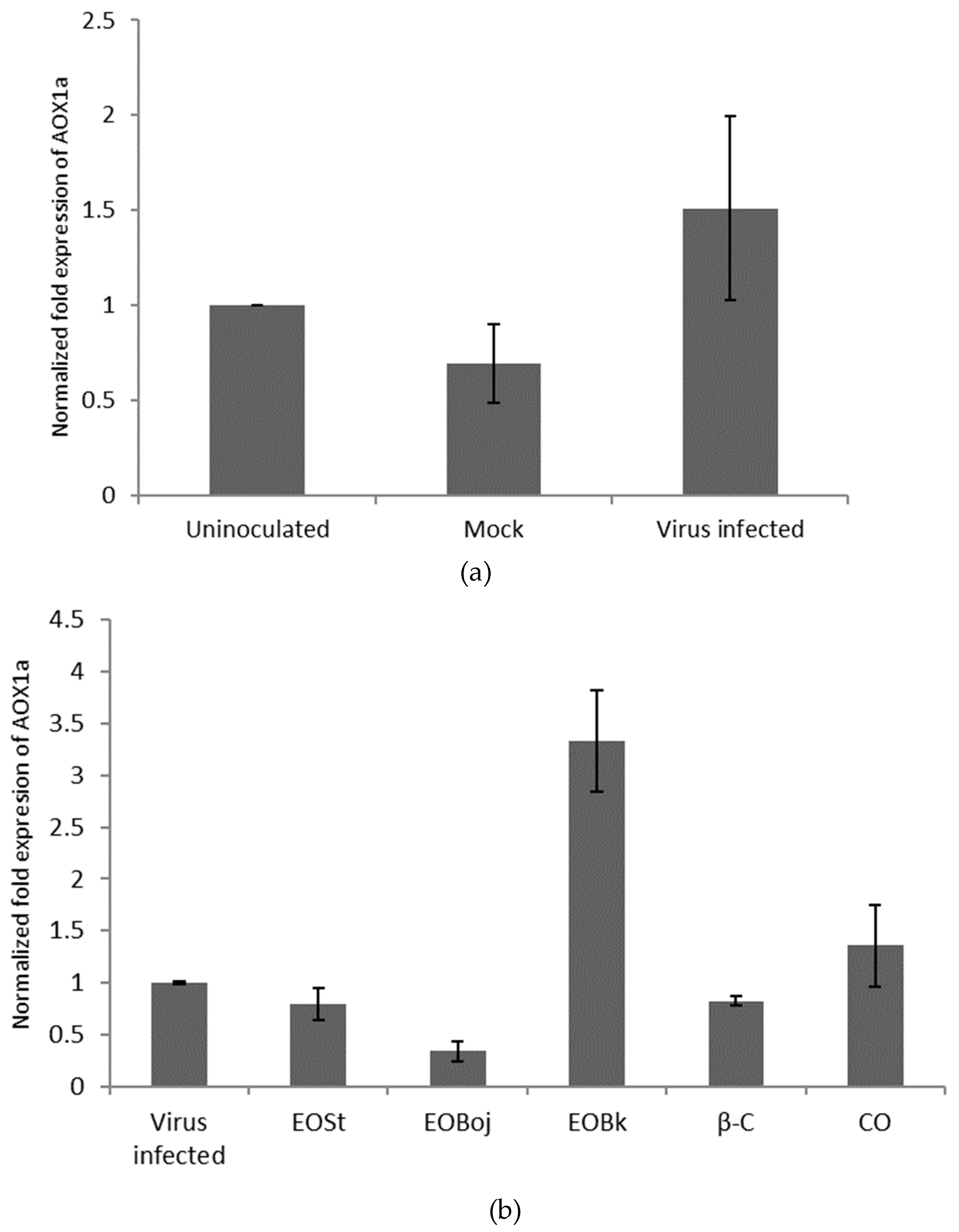
2.3. Aox Gene Expression
3. Materials and Methods
3.1. Micromeria Croatica
3.2. Essential Oil Isolation
3.3. Virus and Plant Hosts
3.4. Antiphytoviral Activity Assay on Local Host Plants
3.4.1. Simultaneous Inoculation with Virus
3.4.2. Treatment Prior to Inoculation
3.5. Antiphytoviral Activity Assay on Systemic Host Plants
3.6. Aox Gene Expression Assays
3.6.1. Treatment of Arabidopsis Thaliana
3.6.2. Quantitative Reverse Transcription-Polymerase Chain Reaction
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils. A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Poma, P.; Labbozzetta, M.; Notarbartolo, M.; Bruno, M.; Maggio, A.; Rosselli, S.; Sajeva, M.; Zito, P. Chemical composition, in vitro antitumor and pro-oxidant activities of Glandora rosmarinifolia (Boraginaceae) essential oil. PLoS ONE 2018, 13, e0196947. [Google Scholar] [CrossRef] [PubMed]
- Padovan, A.; Keszei, A.; Foley, W.J.; Kulheim, C. Differences in gene expression within a striking phenotypic mosaic Eucalyptus tree that varies in susceptibility to herbivory. BMC Plant Biol. 2013, 13, 29. [Google Scholar]
- McLean, S.; Foley, W.J.; Davies, N.W.; Brandon, S.; Duo, L.; Blackman, A.J. Metabolic fate of dietary terpenes from Eucalyptus radiata in common ringtail possum (Pseudocheirus peregrinus). J. Chem. Ecol. 1993, 19, 1625–1643. [Google Scholar] [CrossRef]
- Giamakis, A.; Kretsi, O.; Chinou, I.; Spyropoulos, C.G. Eucalyptus camaldulensis: Volatiles from immature flowers and high production of 1,8-cineole and beta-pinene by in vitro cultures. Phytochemistry 2001, 58, 351–355. [Google Scholar] [CrossRef]
- Alves, M.C.S.; Medeiros Filho, S.; Innecco, R.; Torres, S.B. Allelopathy of plant volatile extracts on seed germination and radicle length of lettuce. Pesqui. Agropecu. Bras. 2004, 39, 1083–1086. [Google Scholar] [CrossRef]
- Eyles, A.; Davies, N.W.; Mohammed, C. Novel detection of formylated phloroglucinol compounds (FPCs) in the wound wood of Eucalyptus globulus and E. nitens. J. Chem. Ecol. 2003, 29, 881–898. [Google Scholar] [CrossRef]
- Zulak, K.G.; Bohlmann, J. Terpenoid biosynthesis and specialized vascular cells of conifer defense. J. Integr Plant Biol. 2010, 52, 86–97. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Bezić, N.; Vuko, E.; Dunkić, V.; Ruščić, M.; Blažević, I.; Burčul, F. Antiphytoviral Activity of Sesquiterpene-Rich Essential Oils from Four Croatian Teucrium Species. Molecules 2011, 16, 8119–8129. [Google Scholar] [CrossRef]
- Bishop, C.D. Antiviral activity of the essential oil of Melaleuca alternifolia (Maiden & Betche) cheel (tea tree) against tobacco mosaic virus. J. Essent. Oil Res. 1995, 7, 641–644. [Google Scholar]
- Dunkić, V.; Bezić, N.; Vuko, E.; Cukrov, D. Antiphytoviral Activity of Satureja montana L. ssp. variegata (Host) P. W. Ball Essential Oil and Phenol Compounds on CMV and TMV. Molecules 2010, 15, 6713–6721. [Google Scholar]
- Dunkić, V.; Bezić, N.; Vuko, E. Antiphytoviral Activity of Essential Oil from Endemic Species Teucrium arduini. Nat. Prod. Commun. 2011, 6, 1385–1388. [Google Scholar] [CrossRef] [PubMed]
- Dunkić, V.; Vuko, E.; Bezić, N.; Kremer, D.; Ruščić, M. Composition and Antiviral Activity of the Essential Oils of Eryngium alpinum and E. amethystinum. Chem. Biodivers. 2013, 10, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- Othman, B.A.; Shoman, S.A. Antiphytoviral activity of the Plectranthus tenuiflorus on some important viruses. Int. J. Agric. Biol. 2004, 6, 844–849. [Google Scholar]
- Vuko, E.; Dunkić, V.; Bezić, N.; Ruščić, M.; Kremer, D. Chemical Composition and Antiphytoviral Activity of Essential Oil of Micromeria graeca. Nat. Prod. Commun. 2012, 7, 1227–1230. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.J.; Xue, X.Y.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [PubMed]
- Kremer, D.; Stabentheiner, E.; Dunkić, V.; Müller, I.D.; Vujić, L.; Kosalec, I.; Ballian, D.; Bogunić, F.; Bezić, N. Micromorphological and chemotaxonomical traits of Micromeria croatica (Pers.) Schott. Chem. Biodivers. 2012, 4, 755–768. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J.C. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Rodrigues, L.; Póvoa, O.; Teixeira, T.; Figueiredo, A.C.; Moldão, M.; Monteiro, A. Trichomes micromorphology and essential oil variation at different developmental stages of cultivated and wild growing Mentha pulegium L. populations from Portugal. Ind. Crop. Prod. 2013, 43, 692–700. [Google Scholar] [CrossRef]
- Rodrigues, L.; Póvoa, O.; van den Berg, C.; Figueiredo, A.C.; Moldão, M.; Monteiro, A. Genetic diversity in Mentha cervina based on morphological traits, essential oils profile and ISSRs markers. Biochem. Syst. Ecol. 2013, 51, 50–59. [Google Scholar] [CrossRef]
- Palukaitis, P.; García-Arenal, F. Cucumoviruses. Adv. Virus Res. 2003, 62, 241–323. [Google Scholar] [CrossRef] [PubMed]
- Sleat, D.E.; Zhang, L.; Palukaitis, P. Mapping determinants within cucumber mosaic virus and its satellite RNA for the induction of necrosis in tomato plants. Mol. Plant Microbe Interact. 1994, 7, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.M.; Gilliland, A.; York, C.J.; Hyman, B.; Carr, J.P. High-level expression of alternative oxidase protein sequences enhances the spread of viral vectors in resistant and susceptible plants. J. Gen. Virol. 2004, 85, 3777–3786. [Google Scholar] [CrossRef] [Green Version]
- Vanlerberghe, G.C.; McIntosh, L. Alternative oxidase: From gene to function. Annu. Rev. Plant Biol. 1997, 48, 703–734. [Google Scholar] [CrossRef] [PubMed]
- Simons, B.H.; Millenaar, F.F.; Mulder, L.; Van Loon, L.C.; Lambers, H. Enhanced expression and activation of the alternative oxidase during infection of Arabidopsis with Pseudomonas syringae pv. tomato. Plant Physiol. 1999, 120, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, D.P.; Wang, Y.; Mcintosh, L. The alternative oxidase lowers mitochondrial reactive oxygen production in plant cells. Proc. Natl. Acad. Sci. USA 1999, 96, 8271–8276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorani, F.; Umbach, A.L.; Siedow, J.N. The alternative oxidase of plant mitochondria is involved in the acclimation of shoot growth at low temperature. A study of Arabidopsis AOX1a transgenic plants. Plant Physiol. 2005, 139, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Clifton, R.; Millar, A.H.; Whelan, J. Alternative oxidases in Arabidopsis: A comparative analysis of differential expression in the gene family provides new insights into function of non-phosphorylating bypasses. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 730–741. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/mass Spectrometry; Allured Publishing: Carol Stream, IL, USA, 2017. [Google Scholar]
- Juszczuk, I.M.; Rychter, A.M. Alternative oxidase in higer plants. Acta Biochim. Pol. 2003, 50, 1257–1271. [Google Scholar]
- Ordog, S.H.; Higgins, V.J.; Vanlerberghe, G.C. Mitochondrial alternative oxidase is not a critical component of plant viral resistance but may play a role in the hypersensitive response. Plant Physiol. 2002, 129, 1858–1865. [Google Scholar] [CrossRef]
- Love, A.J.; Yun, B.W.; Laval, V.; Loake, G.J.; Milner, J.J. Cauliflower mosaic virus, a compatible pathogen of Arabidopsis, engages three distinct defense-signaling pathways and activates rapid systemic generation of reactive oxygen species. Plant Physiol. 2005, 139, 935–948. [Google Scholar] [CrossRef] [PubMed]
- Likić, S. Phenolics in Arabidopsis thaliana (L.) Heynh. infected with Satellite RNA Associated Cucumber Mosaic Virus. Ph.D. Thesis, Faculty of Science, Univesity of Zagreb, Zagreb, Croatia, 2010. [Google Scholar]
- Mayers, C.N.; Lee, K.C.; Moore, C.A.; Wong, S.-M.; Carr, J.P. Salicylic acid-induced resistance to cucumber mosaic virus in squash and Arabidopsis thaliana: Contrasting mechanisms of induction and antiviral action. Mol. Plant Microbe Interact. 2005, 18, 428–434. [Google Scholar] [CrossRef]
- Lacomme, C.; Roby, D. Identification of new early markers of the hypersensitive response in Arabidopsis thaliana. FEBS Lett. 1999, 459, 149–153. [Google Scholar] [CrossRef]
- Škorić, D.M.; Krajačić, M.; Barbarossa, L.; Cillo, F.; Grieco, F.; Šarić, A.; Gallitelli, D. Occurrence of Cucumber Mosaic Cucumovirus with Satellite RNA in Lethal Necrosis Affected Tomatoes in Croatia. J. Phytopathol. 1996, 144, 543–549. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Quantification strategies in real-time PCR. In A-Z of Quantitative PCR; Bustin, S.A., Ed.; International University Line (IUL): La Jolla, CA, USA, 2004; pp. 87–112. [Google Scholar]
Sample Availability: Samples of the essential oils are available from the author V.D. and E.V. |
 EO components identified in M. croatica from Boj and Bk;
EO components identified in M. croatica from Boj and Bk;  EO components identified in M. croatica from Boj and St;
EO components identified in M. croatica from Boj and St;  EO components identified in M. croatica from St;
EO components identified in M. croatica from St;  EO components identified in M. croatica from St and Bk.
EO components identified in M. croatica from Boj and Bk; EO components identified in M. croatica from Boj and St; EO components identified in M. croatica from St; EO components identified in M. croatica from St and Bk.
EO components identified in M. croatica from St and Bk.
EO components identified in M. croatica from Boj and Bk; EO components identified in M. croatica from Boj and St; EO components identified in M. croatica from St; EO components identified in M. croatica from St and Bk.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample (Yield in %) c | |||||
|---|---|---|---|---|---|
| Component | RI a | RI b | Micromeria croatica | ||
| Boj ± SD (1.3) | Bk ± SD (0.7) | St ± SD (0.6) | |||
| Monoterpene hydrocarbons | 21.8 | 24.5 | 14.0 | ||
| α-Thujene | 924 | 1035 | - | 0.9 ± 0.03 | - |
| α-Pinene ** | 938 | 1032 | 0.5 ± 0,15 | 1.1 ± 0.01 | 1.0 ± 0.01 |
| Camphene ** | 962 | 1059 | 0.9 ± 0.01 | 0.4 ± 0.01 | 0.6 ± 0.01 |
| 1-Octen-3-ol | 974 | 1452 | 1.2 ± 0.05 | 1.1 ± 0.01 | 0.3 ± 0.01 |
| Myrcene | 992 | 1174 | 2.4 ± 0.01 | 1.4 ± 0.01 | 0.8 ± 0.01 |
| δ-3-Carene | 1008 | 1153 | 2.5 ± 0.01 | 1.8 ± 0.01 | 0.6 ± 0.1 |
| α-Terpinene | 1016 | 1179 | 2.7 ± 0.01 | 4.2 ± 0.05 | 5.1 ± 0.01 |
| p-Cymene | 1021 | 1268 | 1.5 ± 0.05 | 3.3 ± 0.01 | 1.3 ± 0.06 |
| Limonene | 1032 | 1204 | 3.9 ± 0.06 | 5.9 ± 0.01 | 0.8 ± 0.01 |
| (Z)-β-Ocimene ** | 1032 | 1218 | 0.4 ± 0.01 | 1.6 ± 0.01 | 0.9 ± 0.01 |
| γ-Terpinene | 1057 | 1255 | 3.5 ± 0.03 | 2.6 ± 0.01 | 1.6 ± 0.01 |
| allo-Ocimene | 1128 | 1351 | 2.3 ± 0.01 | 0.9 ± 0.01 | 1.0 ± 0.03 |
| Oxygenated monoterpenes | 25.8 | 41.3 | 27.6 | ||
| Linalool oxide ** | 991 | - | 0.9 ± 0.01 | 0.8 ± 0.01 | 0.5 ± 0.01 |
| Sabinene hydrate | 1065 | 1474 | 1.1 ± 0.04 | 0.7 ± 0.01 | 0.4 ± 0.01 |
| Terpinolene | 1085 | 1286 | 1.6 ± 0.03 | 2.4 ± 0.1 | 0.4 ± 0.07 |
| Linalool | 1097 | 1548 | 3.5 ± 0.03 | 6.4 ± 0.01 | 1.1 ± 0.01 |
| α-Campholenal | 1122 | - | 1.8 ± 0.01 | 1.2 ± 0.01 | 0.7 ± 0.02 |
| cis-Verbenol | 1137 | - | 0.2 ± 0.01 | 0.7 ± 0.03 | 0.5 ± 0.01 |
| Camphor | 1143 | 1499 | 1.9 ± 0.01 | 3.1 ± 0.01 | 6.8 ± 0.01 |
| Borneol | 1165 | 1719 | 2.2 ± 0.03 | - | 0.2 ± 0.01 |
| Terpinen-4-ol | 1174 | 1611 | 1 ± 0.01 | 2.5 ± 0.01 | 0.7 ± 0.01 |
| α-Terpineol | 1186 | 1646 | 1.1 ± 0.01 | 2.1 ± 0.01 | 0.5 ± 0.1 |
| Myrtenol | 1194 | 1804 | 1.4 ± 0.03 | 2.5 ± 0.01 | 3.6 ± 0.03 |
| trans-Carveol | 1216 | - | 1 ± 0.01 | 1.3 ± 0.01 | - |
| Nerol | 1227 | 1808 | 2 ± 0.04 | 0.6 ± 0.01 | 1.2 ± 0.01 |
| Thymol methyl ether | 1230 | 1604 | 0.5 ± 0.01 | 3.3 ± 0.01 | 0.8 ± 0.01 |
| Carvacol methyl ether | 1241 | 1614 | 0.2 ± 0.01 | 1.1 ± 0.01 | 2.1 ± 0.01 |
| Geraniol | 1249 | 1857 | 0.8 ± 0.01 | 5.1 ± 0.01 | 0.6 ± 0.01 |
| Isobornyl acetate | 1283 | - | 0.4 ± 0.01 | 1.7 ± 0.01 | 0.2 ± 0.04 |
| Thymol | 1290 | - | 1.2 ± 0.01 | 0.2 ± 0.01 | 1.1 ± 0.01 |
| Carvacrol | 1298 | 2239 | 2.2 ± 0.01 | 0.7 ± 0.01 | 0.4 ± 0.03 |
| Thymol acetate | 1349 | - | - | - | 5.4 ± 0.01 |
| Neryl acetate | 1358 | 1692 | 0.8 ± 0.1 | 4.9 ± 0.01 | 0.4 ± 0.03 |
| Sesquiterpene hydrocarbons | 31.4 | 5.0 | 16.3 | ||
| α-Cubebene | 1345 | - | 3.5 ± 0.07 | 1.4 ± 0.01 | 2.3 ± 0.01 |
| β-Caryophyllene ** | 1417 | 1585 | 25.2 ± 0.03 | 1.8 ± 0.01 | 10.2 ± 0.01 |
| β-Copaene | 1430 | 1457 | 1 ± 0.01 | 0.7 ± 0.01 | 1.4 ± 0.01 |
| Aromadendrene | 1439 | - | 0.7 ± 0.02 | 0.3 ± 0.01 | 1.9 ± 0.1 |
| δ-Cadinene | 1517 | 1745 | 1 ± 0.01 | 0.8 ± 0.01 | 0.5 ± 0.01 |
| Oxygenated sesquiterpenes | 15.0 | 21.6 | 32.9 | ||
| Sphatulenol | 1578 | 2144 | 1.6 ± 0.04 | 0.5 ± 0.01 | 2.2 ± 0.01 |
| Caryophyllene oxide | 1582 | 1927 | 10.1 ± 0.08 | 21.1 ± 0.01 | 20.2 ± 0.01 |
| Viridiflorol | 1592 | - | 3.3 ± 0.01 | - | 1.1 ± 0.01 |
| β-Bisabolol | 1674 | 2166 | - | - | 3.0 ± 0.01 |
| Caryophyllene acetate ** | 1701 | - | - | - | 4.9 ± 0.1 |
| Farnesol | 1742 | - | - | - | 1.5 ± 0.01 |
| Total identified (%) | 94.0 | 92.4 | 90.8 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vuko, E.; Rusak, G.; Dunkić, V.; Kremer, D.; Kosalec, I.; Rađa, B.; Bezić, N. Inhibition of Satellite RNA Associated Cucumber Mosaic Virus Infection by Essential Oil of Micromeria croatica (Pers.) Schott. Molecules 2019, 24, 1342. https://doi.org/10.3390/molecules24071342
Vuko E, Rusak G, Dunkić V, Kremer D, Kosalec I, Rađa B, Bezić N. Inhibition of Satellite RNA Associated Cucumber Mosaic Virus Infection by Essential Oil of Micromeria croatica (Pers.) Schott. Molecules. 2019; 24(7):1342. https://doi.org/10.3390/molecules24071342
Chicago/Turabian StyleVuko, Elma, Gordana Rusak, Valerija Dunkić, Dario Kremer, Ivan Kosalec, Biljana Rađa, and Nada Bezić. 2019. "Inhibition of Satellite RNA Associated Cucumber Mosaic Virus Infection by Essential Oil of Micromeria croatica (Pers.) Schott" Molecules 24, no. 7: 1342. https://doi.org/10.3390/molecules24071342