Enhanced Anti-Allergic Activity of Milk Casein Phosphopeptide by Additional Phosphorylation in Ovalbumin-Sensitized Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Phosphorylation and Dephosphorylation of CPP
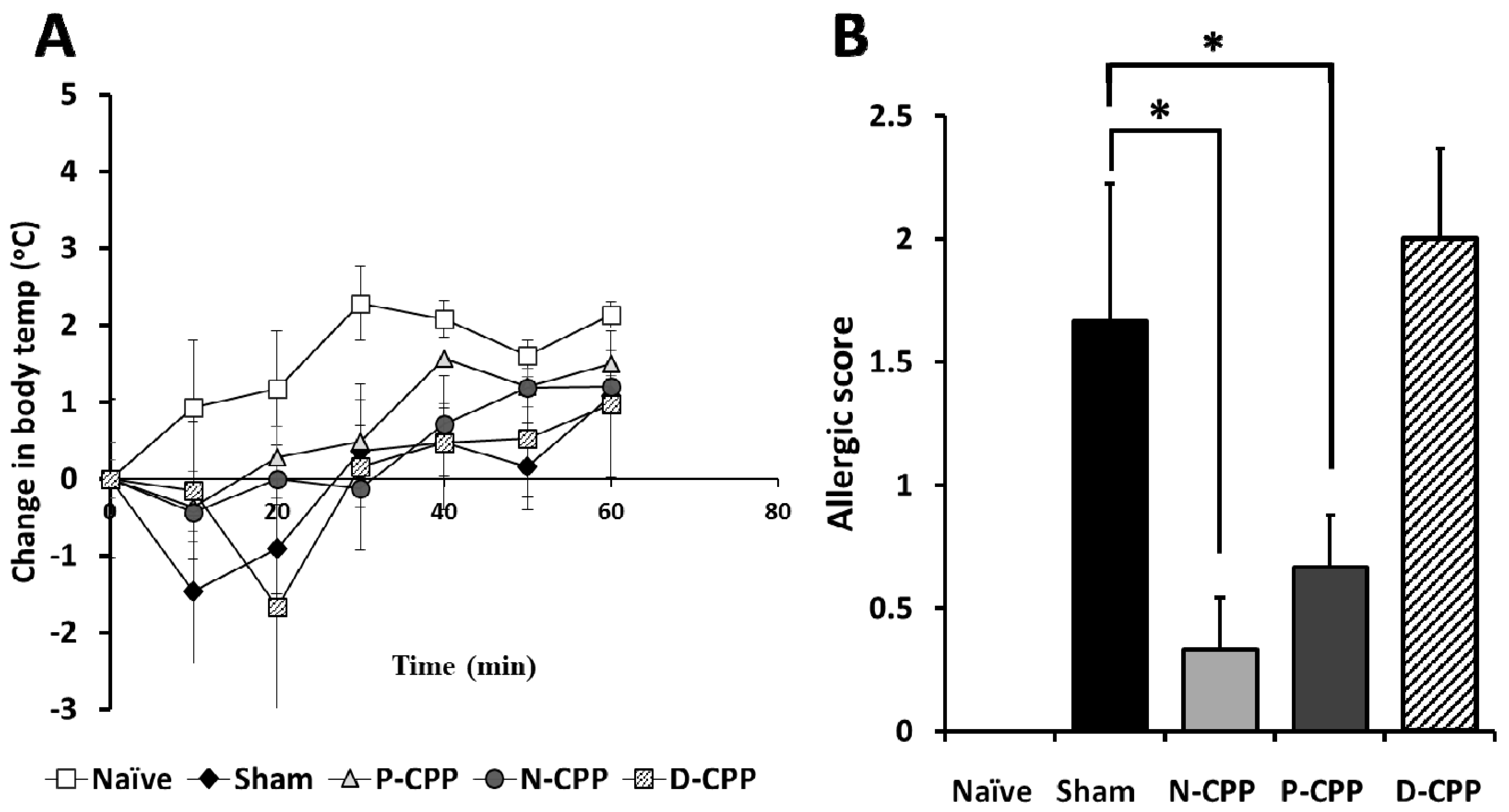
2.2. Effects of Oral Administration of P-CPP and D-CPP against OVA-Induced IgE and Allergic Reactions
2.3. Effects of Orally Fed Phosphorylated and Dephosphorylated CPP-III on OVA-Induced Cytokine Production In Vitro
2.4. Effects of Orally Fed Phosphorylated CPP-III on Differentiation and Population Changes of Regulatory T (Treg) Cells and T Follicular Helper (Tfh) Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Phosphorylated and Dephosphorylated CPP-III
4.3. Mice
4.4. OVA-Sensitized Allergic Model Mice
4.5. Allergic Score
4.6. ELISA Analysis for OVA-Specific IgE, IgA, Total IgE, IgA, and OVA-Specific IgG1 and IgG2a
4.7. ELISA Quantification of Cytokine Levels in Splenocytes
4.8. GATA-3 and IL-4 Gene EXPRESSION Analysis by Quantitative PCR (qPCR)
4.9. Flow Cytometric Analysis of Treg and Tfh Cell Populations
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sicherer, S.H.; Sampson, H.A. Food allergy. J. Allergy Clin. Immunol. 2006, 117, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Noval Rivas, M.; Chatila, T.A. Regulatory T cells in allergic diseases. J. Allergy Clin. Immunol. 2016, 138, 639–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagan, R.S. Food allergy: An overview. Environ. Health Perspect. 2003, 111, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Katayama, S.; Mine, Y. Quillaja saponin can modulate ovalbumin-induced IgE allergic responses through regulation of Th1/Th2 balance in a murine model. J. Agric. Food Chem. 2006, 54, 3271–3276. [Google Scholar] [CrossRef] [PubMed]
- Théolier, J.; Fliss, I.; Jean, J.; Hammami, R. Antimicrobial Peptides of Dairy Proteins: From Fundamental to Applications. Food Rev. Int. 2014, 30, 134–154. [Google Scholar] [CrossRef]
- Meisel, H. Multifunctional peptides encrypted in milk proteins. Biofactors 2004, 21, 55–61. [Google Scholar] [CrossRef]
- Meisel, H. Biochemical properties of peptides encrypted in bovine milk proteins. Curr. Med. Chem. 2005, 12, 1905–1919. [Google Scholar] [CrossRef]
- Park, Y.W.; Nam, M.S. Bioactive Peptides in Milk and Dairy Products: A Review. Korean J. Food Sci. Anim. Resour. 2015, 35, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Tobita, K.; Kawahara, T.; Otani, H. Bovine β-casein (1-28), a casein phosphopeptide, enhances proliferation and IL-6 expression of mouse CD19+ cells via toll-like receptor 4. J. Agric. Food Chem. 2006, 54, 8013–8017. [Google Scholar] [CrossRef]
- Otani, H.; Nakano, K.; Kawahara, T. Stimulatory effect of a dietary casein phosphopeptide preparation on the mucosal IgA response of mice to orally ingested lipopolysaccharide from Salmonella typhimurium. Biosci. Biotechnol. Biochem. 2003, 67, 729–735. [Google Scholar] [CrossRef]
- Otani, H.; Kihara, Y.; Park, M. The immunoenhancing property of a dietary casein phosphopeptide preparation in mice. Food Agric. Immunol. 2000, 12, 165–173. [Google Scholar] [CrossRef]
- Danquah, M.K.; Agyei, D. Pharmaceutical applications of bioactive peptides. OA Biotechnol. 2012, 2, 5. [Google Scholar]
- Caubet, J.C.; Wang, J. Current Understanding of Egg Allergy. Pediatr. Clin. North Am. 2011, 58, 427–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamdad, F.; Shin, S.H.; Suh, J.-W.; Nimalaratne, C.; Sunwoo, H. Anti-Inflammatory and Antioxidant Properties of Casein Hydrolysate Produced Using High Hydrostatic Pressure Combined with Proteolytic Enzymes. Molecules 2017, 22, 609. [Google Scholar] [CrossRef] [PubMed]
- Makabe-Kobayashi, Y.; Hori, Y.; Adachi, T.; Ishigaki-Suzuki, S.; Kikuchi, Y.; Kagaya, Y.; Shirato, K.; Nagy, A.; Ujike, A.; Takai, T.; et al. The control effect of histamine on body temperature and respiratory function in IgE-dependent systemic anaphylaxis. J. Allergy Clin. Immunol. 2002, 110, 298–303. [Google Scholar] [CrossRef]
- Ariza, A.; Fernandez, T.D.; Doña, I.; Aranda, A.; Blanca-Lopez, N.; Melendez, L.; Canto, G.; Blanca, M.; Torres, M.J.; Mayorga, C. Basophil activation after nonsteroidal anti-inflammatory drugs stimulation in patients with immediate hypersensitivity reactions to these drugs. Cytom. Part A 2014, 85, 400–407. [Google Scholar] [CrossRef] [Green Version]
- Oettgen, H.C.; Burton, O.T. IgE receptor signaling in food allergy pathogenesis. Curr. Opin. Immunol. 2015, 36, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Leong, K.W.; Ding, J.L. The Unexplored Roles of Human Serum IgA. DNA Cell Biol. 2014, 33, 823–829. [Google Scholar] [CrossRef] [Green Version]
- Kiewiet, M.B.G.; Faas, M.M.; de Vos, P. Immunomodulatory protein hydrolysates and their application. Nutrients 2018, 10, 904. [Google Scholar] [CrossRef]
- Lebetwa, N.; Mitani, T.; Nakamura, S.; Katayama, S. Role of phosphate groups on antiviral activity of casein phosphopeptide against feline calicivirus as a surrogate for norovirus. J. Sci. Food Agric. 2017, 97, 1939–1944. [Google Scholar] [CrossRef]
- Meulenbroek, L.A.P.M.; van Esch, B.C.A.M.; Hofman, G.A.; den Hartog Jager, C.F.; Nauta, A.J.; Willemsen, L.E.M.; Bruijnzeel-Koomen, C.A.F.M.; Garssen, J.; van Hoffen, E.; Knippels, L.M.J. Oral treatment with β-lactoglobulin peptides prevents clinical symptoms in a mouse model for cow’s milk allergy. Pediatr. Allergy Immunol. 2013, 24, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; van Esch, B.C.A.M.; Garssen, J.; Faas, M.M.; de Vos, P. Partially hydrolyzed whey proteins prevent clinical symptoms in a cow’s milk allergy mouse model and enhance regulatory T and B cell frequencies. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Van Esch, B.C.A.M.; Schouten, B.; de Kivit, S.; Hofman, G.A.; Knippels, L.M.J.; Willemsen, L.E.M.; Garssen, J. Oral tolerance induction by partially hydrolyzed whey protein in mice is associated with enhanced numbers of Foxp3+regulatory T-cells in the mesenteric lymph nodes. Pediatr. Allergy Immunol. 2011, 22, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.D.; Wu, Z.; Liu, J.; Cao, X.Y.; Zeng, X.Q. Immunomodulatory and hypoallergenic properties of milk protein hydrolysates in ICR mice. J. Dairy Sci. 2013, 96, 4958–4964. [Google Scholar] [CrossRef] [PubMed]
- Corry, D.B.; Kheradmand, F. Induction and regulation of the IgE response. Nature 1999, 402, B18–B23. [Google Scholar] [CrossRef] [PubMed]
- Bellanti, J.A. Cytokines and allergic diseases: Clinical aspects. Allergy Asthma Proc. 1998, 19, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.Y. GATA3: A master of many trades in immune regulation. Trends Immunol. 2014, 35, 233–242. [Google Scholar] [CrossRef]
- Zheng, W.P.; Flavell, R.A. The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. Cell 1997, 89, 587–596. [Google Scholar] [CrossRef]
- O’Garra, A.; Gabryšová, L. Transcription Factors Directing Th2 Differentiation: Gata-3 Plays a Dominant Role. J. Immunol. 2016, 196, 4423–4425. [Google Scholar] [CrossRef] [Green Version]
- Ferber, I.A.; Lee, H.J.; Zonin, F.; Heath, V.; Mui, A.; Arai, N.; O’Garra, A. GATA-3 significantly downregulates IFN-γ production from developing Th1 cells in addition to inducing IL-4 and IL-5 levels. Clin. Immunol. 1999, 91, 134–144. [Google Scholar] [CrossRef]
- Finotto, S.; De Sanctis, G.T.; Lehr, H.A.; Herz, U.; Buerke, M.; Schipp, M.; Bartsch, B.; Atreya, R.; Schmitt, E.; Galle, P.R.; et al. Treatment of allergic airway inflammation and hyperresponsiveness by antisense-induced local blockade of GATA-3 expression. J. Exp. Med. 2001, 193, 1247–1260. [Google Scholar] [CrossRef] [PubMed]
- Cao, A.T.; Yao, S.; Gong, B.; Nurieva, R.I.; Elson, C.O.; Cong, Y. Interleukin (IL)-21 promotes intestinal IgA response to microbiota. Mucosal. Immunol. 2015, 8, 1072–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamagawa-Mineoka, R.; Kishida, T.; Mazda, O.; Katoh, N. IL-21 reduces immediate hypersensitivity reactions in mouse skin by suppressing mast cell degranulation and IgE production. J. Dermatol. Sci. 2010, 131, 1513–1520. [Google Scholar] [CrossRef]
- Kemeny, D. The role of the T follicular helper cells in allergic disease. Cell. Mol. Immunol. 2012, 9, 386–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiromura, Y.; Kishida, T.; Nakano, H.; Hama, T.; Imanishi, J.; Hisa, Y.; Mazda, O. IL-21 Administration into the Nostril Alleviates Murine Allergic Rhinitis. J. Immunol. 2007, 179, 7157–7165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhardt, R.L.; Liang, H.E.; Locksley, R.M. Cytokine-secreting follicular T cells shape the antibody repertoire. Nat. Immunol. 2009, 10, 385–393. [Google Scholar] [CrossRef] [Green Version]
- King, I.L.; Mohrs, M. IL-4–producing CD4+ T cells in reactive lymph nodes during helminth infection are T follicular helper cells. J. Exp. Med. 2009, 206, 1001–1007. [Google Scholar] [CrossRef]
- Kobayashi, T.; Iijima, K.; Dent, A.L.; Kita, H. Follicular helper T cells mediate IgE antibody response to airborne allergens. J. Allergy Clin. Immunol. 2017, 139, 300–313.e7. [Google Scholar] [CrossRef] [Green Version]
- Dolence, J.J.; Kobayashi, T.; Iijima, K.; Krempski, J.; Drake, L.Y.; Dent, A.L.; Kita, H. Airway exposure initiates peanut allergy by involving the IL-1 pathway and T follicular helper cells in mice. J. Allergy Clin. Immunol. 2018, 142, 1144–1158.e8. [Google Scholar] [CrossRef]
- Meli, A.P.; Fontés, G.; Leung Soo, C.; King, I.L. T Follicular Helper Cell–Derived IL-4 Is Required for IgE Production during Intestinal Helminth Infection. J. Immunol. 2017. [Google Scholar] [CrossRef]
- Coste, M.; Rochet, V.; Léonil, J.; Mollé, D.; Bouhallab, S.; Tomé, D. Identification of C-terminal peptides of bovine β-casein that enhance proliferation of rat lymphocytes. Immunol. Lett. 1992, 33, 41–46. [Google Scholar] [CrossRef]
- Kasai, T.; Iwasaki, R.; Tanaka, M.; Kiriyama, S. Caseinphosphopeptides (cpp) in feces and contents in digestive tract of rats fed casein and cpp preparations. Biosci. Biotechnol. Biochem. 1995, 59, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Meisel, H.; Frister, H. Chemical characterization of bioactive peptides from in vivo digests of casein. J. Dairy Res. 1989, 56, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.M.; Kato, T.; Ohnishi, H.; Kawamoto, N.; Kato, Z.; Kaneko, H.; Kondo, N.; Nakano, T. Hypoallergenic casein hydrolysate for peptide-based oral immunotherapy in cow’s milk allergy. J. Allergy Clin. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Kim, H.S.; Kim, D.K.; Nam, S.T.; Kim, H.W.; Park, Y.H.; Lee, D.; Lee, M.B.; Lee, J.H.; Kim, B.; et al. Mesenteric IL-10-producing CD5+ regulatory B cells suppress cow’s milk casein-induced allergic responses in mice. Sci. Rep. 2016, 6, 19685. [Google Scholar] [CrossRef] [Green Version]
- Reddi, S.; Kapila, R.; Dang, A.K.; Kapila, S. Evaluation of allergenic response of milk bioactive peptides using mouse mast cell. Milchwissenschaft-Milk Sci. Int. 2012. [Google Scholar] [CrossRef]
- Bernard, H.; Meisel, H.; Creminon, C.; Wal, J.M. Post-translational phosphorylation affects the IgE binding capacity of caseins. FEBS Lett. 2000. [Google Scholar] [CrossRef]
- Oliver, C.M.; Melton, L.D.; Stanley, R.A. Creating proteins with novel functionality via the maillard reaction: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 337–350. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kassai, M.; Hirose, T.; Katayama, S.; Nakamura, K.; Akiyama, H.; Teshima, R.; Nakamura, S. Modulation of immunoresponse in BALB/c mice by oral administration of Fag e 1-glucomannan conjugate. J. Agric. Food Chem. 2009, 57, 9787–9792. [Google Scholar] [CrossRef]
- Darewicz, M.; Dziuba, J.; Caessens, P.W.; Gruppen, H. Dephosphorylation-induced structural changes in beta-casein and its amphiphilic fragment in relation to emulsion properties. Biochimie 2000, 82, 191–195. [Google Scholar] [CrossRef]
- Onishi, N.; Kawamoto, S.; Ueda, K.; Yamanaka, Y.; Katayama, A.; Suzuki, H.; Aki, T.; Hashimoto, K.; Hide, M.; Ono, K. Dietary Pulverized Konjac Glucomannan Prevents the Development of Allergic Rhinitis-Like Symptoms and IgE Response in Mice. Biosci. Biotechnol. Biochem. 2007, 71, 2551–2556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyahara, S.; Miyahara, N.; Takeda, K.; Joetham, A.; Gelfand, E.W. Physiologic assessment of allergic rhinitis in mice: Role of the high-affinity IgE receptor (FcεRI). J. Allergy Clin. Immunol. 2005, 116, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schofield, B.H.; Huang, C.-K.; Kleiner, G.I.; Sampson, H.A. A murine model of IgE-mediated cow’s milk hypersensitivity. J. Allergy Clin. Immunol. 1999, 103, 206–214. [Google Scholar] [CrossRef]
Sample Availability: Not available. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebetwa, N.; Suzuki, Y.; Tanaka, S.; Nakamura, S.; Katayama, S. Enhanced Anti-Allergic Activity of Milk Casein Phosphopeptide by Additional Phosphorylation in Ovalbumin-Sensitized Mice. Molecules 2019, 24, 738. https://doi.org/10.3390/molecules24040738
Lebetwa N, Suzuki Y, Tanaka S, Nakamura S, Katayama S. Enhanced Anti-Allergic Activity of Milk Casein Phosphopeptide by Additional Phosphorylation in Ovalbumin-Sensitized Mice. Molecules. 2019; 24(4):738. https://doi.org/10.3390/molecules24040738
Chicago/Turabian StyleLebetwa, Ntshepisa, Yuta Suzuki, Sachi Tanaka, Soichiro Nakamura, and Shigeru Katayama. 2019. "Enhanced Anti-Allergic Activity of Milk Casein Phosphopeptide by Additional Phosphorylation in Ovalbumin-Sensitized Mice" Molecules 24, no. 4: 738. https://doi.org/10.3390/molecules24040738