
High-Throughput 1H-Nuclear Magnetic Resonance-Based Screening for the Identification and Quantification of Heartwood Diterpenic Acids in Four Black Pine (Pinus nigra Arn.) Marginal Provenances in Greece


Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
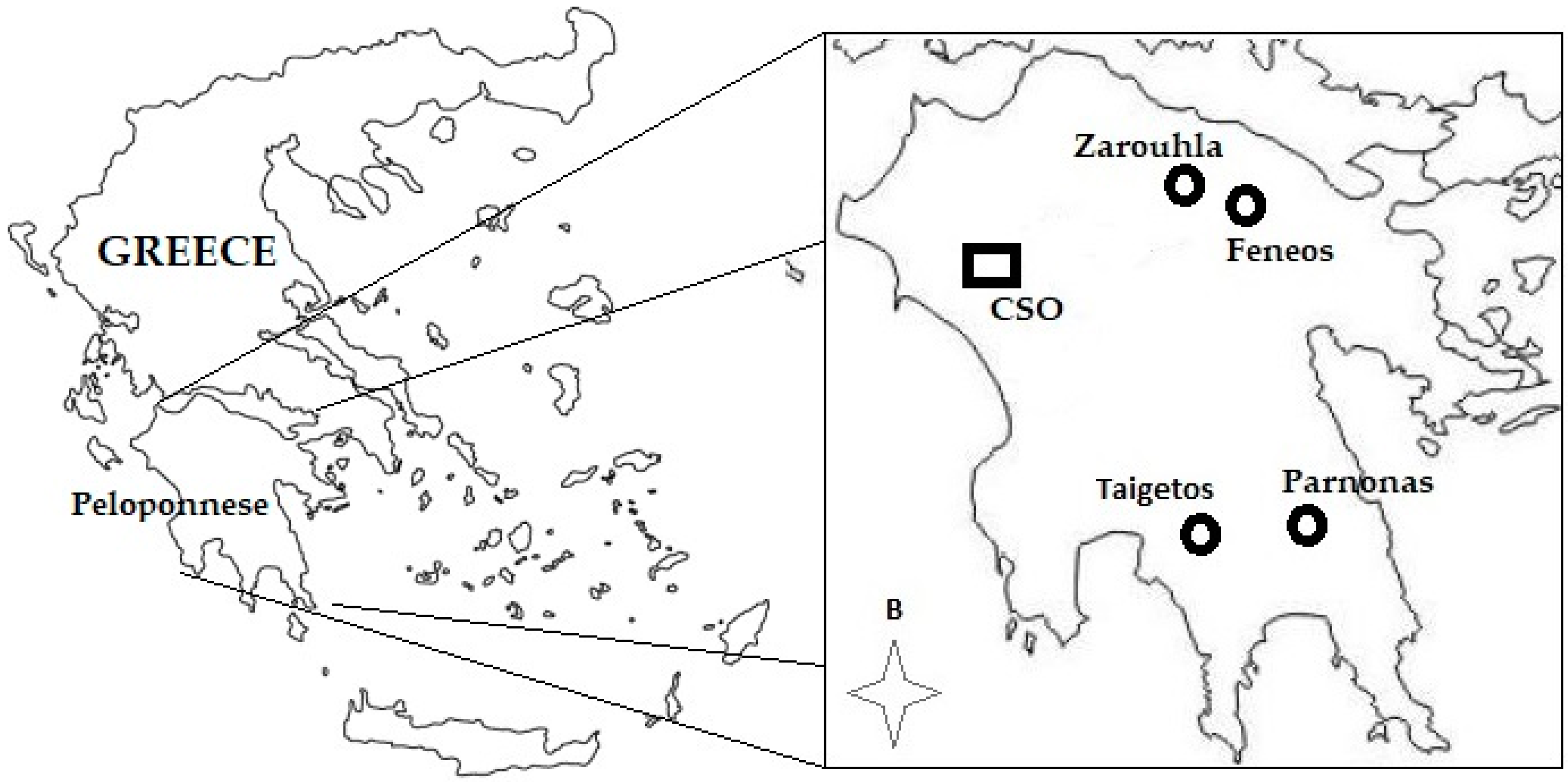
4.1. Plant Material
4.2. Sampling
4.3. Extraction Protocol
4.3.1. H-NMR Spectral Analysis/Quantitation Methodology
4.3.2. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Matziris, D. Variation in growth and branching characters in black pine (Pinus nigra Arnold) of Peloponnesos. Silvae Genet. 1989, 38, 77–81. [Google Scholar]
- Pietarinen, S.P.; Willför, S.M.; Ahotupa, M.O.; Hemming, J.E.; Holmbom, B.R. Knotwood and bark extracts: Strong antioxidants from waste materials. J. Wood Sci. 2006, 52, 436–444. [Google Scholar] [CrossRef]
- Ioannidis, K.; Melliou, E.; Alizoti, P.; Magiatis, P. Identification of black pine (Pinus nigra Arn.) heartwood as a rich source of bioactive stilbenes by qNMR. J. Sci. Food Agric. 2017, 97, 1708–1716. [Google Scholar] [CrossRef] [PubMed]
- Hillis, W.E. Heartwood and Tree Exudates; Springer Verlag: Berlin, Germany, 1987; p. 268. [Google Scholar]
- Porter, L.J. The resin and fatty acid content of living Pinus radiata wood. N. Z. J. For. Sci. 1969, 12, 687–693. [Google Scholar]
- Hemingway, R.W.; Hillis, W. Changes in fats and resins of Pinus radiata associated with heartwood formation. Appita 1971, 24, 439–443. [Google Scholar]
- Lloyd, J.A. Distribution of extractives in Pinus radiata earlywood and latewood. N. Z. J. For. Sci. 1978, 8, 288–294. [Google Scholar]
- Nisula, L. Wood Extractives in Conifers: A Study of Stemwood and Knots of Industrially Important Species; Åbo Akademi University Press: Åbo, Finland, 2018; p. 372. [Google Scholar]
- Joye, N.M.; Lawrence, R.V. Resin acid composition of pine oleoresins. J. Chem. Eng. Data 1967, 12, 279–282. [Google Scholar] [CrossRef]
- Conner, A.H.; Diehl, M.A.; Rowe, J.W. Tall oil precursors in three western pines: Ponderosa, Lodgepole, and Limber pine. Wood Sci. 1980, 12, 183–191. [Google Scholar]
- Conner, A.; Diehl, M.; Rowe, J. Tall oil precursors and turpentine in jack and eastern white pine. Wood Sci. 1980, 12, 194–200. [Google Scholar]
- Lichtenthaler, H.K. The 1-deoxy-d-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef]
- Keeling, C.I.; Bohlmann, J. Diterpene resin acids in conifers. Phytochemistry 2006, 67, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Fries, A.; Ericsson, T.; Gref, R. High heritability of wood extractives in Pinus sylvestris progeny tests. Can. J. For. Res. 2000, 30, 1707–1713. [Google Scholar] [CrossRef]
- Ericsson, T.; Fries, A.; Gref, R. Genetic correlations of heartwood extractives in Pinus sylvestris progeny tests. For. Genet. 2001, 8, 73–79. [Google Scholar]
- Squillace, A.; Harrington, T. Olustee’s high-yielder produces 487 bbls pine gum per crop for four straight years. Nav. Stores Rev. 1968, 77, 4–5. [Google Scholar]
- Franklin, E.; Taras, M.; Volkman, D. Genetic gains in yields of oleoresin, wood extractives and tall oil. Tappi 1970, 53, 2302–2304. [Google Scholar]
- Taylor, A.M.; Gartner, B.L.; Morre, J.I. Heartwood formation and natural durability—A review. Wood Fiber Sci. 2002, 34, 587–611. [Google Scholar]
- Langenheim, J.H. Plant Resins: Chemistry, Evolution, Ecology, and Ethnobotany; Timber Press: Portland, OR, USA, 2003; p. 612. ISBN 0-88192-574-8. [Google Scholar]
- Rezzi, S.; Bighelli, A.; Castola, V.; Casanova, J. Composition and chemical variability of the oleoresin of Pinus nigra ssp. laricio from Corsica. Ind. Crops Prod. 2005, 21, 71–79. [Google Scholar] [CrossRef]
- Spessard, G.O.; Matthews, D.R.; Nelson, M.D.; Rajtora, T.C.; Fossum, M.J.; Giannini, J.L. Phytoalexin-like activity of abietic acid and its derivatives. J. Agric. Food Chem. 1995, 43, 1690–1694. [Google Scholar] [CrossRef]
- De Oliveira, A.M.; Tirapelli, C.R.; Ambrosio, S.R.; da Costa, F.B. Diterpenes: A therapeutic promise for cardiovascular diseases. Recent Patents Cardiovasc. Drug Discov. 2008, 3, 1–8. [Google Scholar] [CrossRef]
- Gonzalez, M.A.; Correa-Royero, J.; Agudelo, L.; Mesa, A.; Betancur-Galvis, L. Synthesis and biological evaluation of abietic acid derivatives. Eur. J. Med. Chem. 2009, 44, 2468–2472. [Google Scholar] [CrossRef]
- Ulusu, N.N.; Ercil, D.; Sakar, M.K.; Tezcan, E.F. Abietic acid inhibits lipoxygenase activity. Phytother. Res. 2002, 16, 88–90. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.; Tornos, M.; Garcia, M.; De las Heras, B.; Villar, A.; Saenz, M. Anti-inflammatory activity of abietic acid, a diterpene isolated from Pimenta racemosa var. grissea. J. Pharm. Pharmacol. 2001, 53, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Kawada, T.; Goto, T.; Kim, C.-S.; Taimatsu, A.; Egawa, K.; Yamamoto, T.; Jisaka, M.; Nishimura, K.; Yokota, K. Abietic acid activates peroxisome proliferator-activated receptor-γ (PPARγ) in RAW264. 7 macrophages and 3T3-L1 adipocytes to regulate gene expression involved in inflammation and lipid metabolism. FEBS Lett. 2003, 550, 190–194. [Google Scholar] [CrossRef]
- Talevi, A.; Cravero, M.S.; Castro, E.A.; Bruno-Blanch, L.E. Discovery of anticonvulsant activity of abietic acid through application of linear discriminant analysis. Bioorg. Med. Chem. Lett. 2007, 17, 1684–1690. [Google Scholar] [CrossRef] [PubMed]
- Schmeda-Hirschmann, G.; Astudillo, L.; Rodríguez, J.; Theoduloz, C.; Yáñez, T. Gastroprotective effect of the Mapuche crude drug Araucaria araucana resin and its main constituents. J. Ethnopharmacol. 2005, 101, 271–276. [Google Scholar] [CrossRef]
- Schmeda-Hirschmanna, G.; Astudillo, L.; Sepúlveda, B.; Rodríguez, J.A.; Theoduloz, C.; Yáñez, T.; Palenzuela, J.A. Gastroprotective effect and cytotoxicity of natural and semisynthetic labdane diterpenes from Araucaria araucana resin. Z. Naturforschung C 2005, 60, 511–522. [Google Scholar] [CrossRef]
- San Feliciano, A.; Gordaliza, M.; Salinero, M.A.; del Corral, J.M.M. Abietane acids: Sources, biological activities, and therapeutic uses. Planta Med. 1993, 59, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Reveglia, P.; Cimmino, A.; Masi, M.; Nocera, P.; Berova, N.; Ellestad, G.; Evidente, A. Pimarane diterpenes: Natural source, stereochemical configuration, and biological activity. Chirality 2018, 30, 1115–1134. [Google Scholar] [CrossRef]
- Simmler, C.; Napolitano, J.G.; McAlpine, J.B.; Chen, S.-N.; Pauli, G.F. Universal quantitative NMR analysis of complex natural samples. Curr. Opin. Biotechnol. 2014, 25, 51–59. [Google Scholar] [CrossRef]
- Nikolantonaki, M.; Magiatis, P.; Waterhouse, A.L. Direct analysis of free and sulfite-bound carbonyl compounds in wine by two-dimensional quantitative proton and carbon nuclear magnetic resonance spectroscopy. Anal. Chem. 2015, 87, 10799–10806. [Google Scholar] [CrossRef]
- Cerulli, A.; Masullo, M.; Montoro, P.; Hošek, J.; Pizza, C.; Piacente, S. Metabolite profiling of “green” extracts of Corylus avellana leaves by 1H NMR spectroscopy and multivariate statistical analysis. J. Pharm. Biomed. Anal. 2018, 160, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Popescu, R.; Ionete, R.E.; Botoran, O.R.; Costinel, D.; Bucura, F.; Geana, E.I.; Alabedallat, Y.F.J.; Botu, M. 1H-NMR profiling and carbon isotope discrimination as tools for the comparative assessment of walnut (Juglans regia L.) cultivars with various geographical and genetic origins—A preliminary study. Molecules 2019, 24, 1378. [Google Scholar] [CrossRef] [PubMed]
- Merkx, D.W.H.; Hong, G.T.S.; Ermacora, A.; van Duynhoven, J.P.M. Rapid quantitative profiling of lipid oxidation products in a food emulsion by 1H NMR. Anal. Chem. 2018, 90, 4863–4870. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zhong, Q.; Fauhl-Hassek, C.; Pfister, M.K.-H.; Horn, B.; Huang, Z. Classification of Chinese wine varieties using 1H NMR spectroscopy combined with multivariate statistical analysis. Food Control 2018, 88, 113–122. [Google Scholar] [CrossRef]
- Barrilero, R.; Gil, M.; Amigó, N.; Dias, C.B.; Wood, L.G.; Garg, M.L.; Ribalta, J.; Heras, M.; Vinaixa, M.; Correig, X. LipSpin: A new bioinformatics tool for quantitative 1H NMR lipid profiling. Anal. Chem. 2018, 90, 2031–2040. [Google Scholar] [CrossRef] [PubMed]
- Suckling, I.D.; Ede, R. A quantitative 13C nuclear magnetic resonance method for the analysis of wood extractives and pitch samples. Appita J. 1990, 43, 77–80. [Google Scholar]
- Rezzi, S.; Bighelli, A.; Castola, V.; Casanova, J. Direct identification and quantitative determination of acidic and neutral diterpenes using 13C-NMR spectroscopy. application to the analysis of oleoresin of Pinus nigra. Appl. Spectrosc. 2002, 56, 312–317. [Google Scholar] [CrossRef]
- Skakovskii, E.D.; Tychinskaya, L.Y.; Gaidukevich, O.A.; Kozlov, N.G.; Klyuev, A.Y.; Lamotkin, S.A.; Shpak, S.I.; Rykov, S.V. NMR determination of the composition of balsams from Scotch pine resin. J. Appl. Spectrosc. 2008, 75, 439–443. [Google Scholar] [CrossRef]
- Rotondo, A.; Mannina, L.; Salvo, A. Multiple assignment recovered analysis (MARA) NMR for a direct food labeling: The case study of olive oils. Food Anal. Methods 2019, 12, 1238–1245. [Google Scholar] [CrossRef]
- Karkoula, E.; Skantzari, A.; Melliou, E.; Magiatis, P. Direct measurement of oleocanthal and oleacein levels in olive oil by quantitative 1H NMR. Establishment of a new index for the characterization of extra virgin olive oils. J. Agric. Food Chem. 2012, 60, 11696–11703. [Google Scholar] [CrossRef]
- Karkoula, E.; Skantzari, A.; Melliou, E.; Magiatis, P. Quantitative measurement of major secoiridoid derivatives in olive oil using qNMR. Proof of the artificial formation of aldehydic oleuropein and ligstroside aglycon isomers. J. Agric. Food Chem. 2014, 62, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Manoukian, P.; Melliou, E.; Liouni, M.; Magiatis, P. Identification and quantitation of benzoxazinoids in wheat malt beer by qNMR and GC–MS. LWT Food Sci. Technol. 2016, 65, 1133–1137. [Google Scholar] [CrossRef]
- Cannac, M.; Barboni, T.; Ferrat, L.; Bighelli, A.; Castola, V.; Costa, J.; Trecul, D.; Morandini, F.; Pasqualini, V. Oleoresin flow and chemical composition of Corsican pine (Pinus nigra subsp. laricio) in response to prescribed burnings. For. Ecol. Manag. 2009, 257, 1247–1254. [Google Scholar] [CrossRef]
- Zobel, B.; Jett, J. Genetics of Wood Production; Springer-Verlag: Berlin, Germany, 1995; 337p. [Google Scholar]
- Venäläinen, M.; Harju, A.M.; Kainulainen, P.; Viitanen, H.; Nikulainen, H. Variation in the decay resistance and its relationship with other wood characteristics in old Scots pines. Ann. For. Sci. 2003, 60, 409–417. [Google Scholar] [CrossRef]
- Matziris, D. Genetic variation in morphological and anatomical needle characteristics in the Black pine of Peloponnesos. Silvae Genet. 1984, 33, 164–169. [Google Scholar]
- Ioannidis, K. Genetic Improvement of Multiple Traits in Black Pine (Pinus nigra Arn.). Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2018. [Google Scholar]
- Willför, S.; Hemming, J.; Reunanen, M.; Holmbom, B. Phenolic and lipophilic extractives in Scots pine knots and stemwood. Holzforschung 2003, 57, 359–372. [Google Scholar] [CrossRef]
- Song, Z. Characteristics of Oleoresin and Classification of Pinus in China; China Forestry Publishing House: Beijing, China, 1998; 206p. [Google Scholar]
- Martínez-Inigo, M.J.; Immerzeel, P.; Gutierrez, A.; del Río, J.C.; Sierra-Alvarez, R. Biodegradability of extractives in sapwood and heartwood from Scots pine by sapstain and white-rot fungi. Holzforschung 1999, 53, 247–252. [Google Scholar] [CrossRef]
- Belt, T.; Keplinger, T.; Hänninen, T.; Rautkari, L. Cellular level distributions of Scots pine heartwood and knot heartwood extractives revealed by Raman spectroscopy imaging. Ind. Crops Prod. 2017, 108, 327–335. [Google Scholar] [CrossRef]
- Hafizoglu, H. Wood extractives of Pinus sylvestris L., Pinus nigra Arn. and Pinus brutia Ten. with special reference to nonpolar components. Holzforschung 1983, 37, 321–326. [Google Scholar]
- Uprichard, J.M.; Lloyd, J.A. Influence of tree age on the chemical composition of Radiata pine. N. Z. J. For. Sci. 1980, 10, 551–557. [Google Scholar]
- Yildirim, H.; Holmbom, B. Investigations on the wood extractives of pine species from Turkey: III Non-volatile, nonpolar components in Pinus brutia. Acta Acad. Abo. 1977, B37, 1–9. [Google Scholar]
- Hovelstad, H.; Leirset, I.; Oyaas, K.; Fiksdahl, A. Screening analyses of pinosylvin stilbenes, resin acids and lignans in Norwegian conifers. Molecules 2006, 11, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Harju, A.M.; Venäläinen, M.; Anttonen, S.; Viitanen, H.; Kainulainen, P.; Saranpää, P.; Vapaavuori, E. Chemical factors affecting the brown-rot decay resistance of Scots pine heartwood. Trees 2003, 17, 263–268. [Google Scholar]
- Arshadi, M.; Backlund, I.; Geladi, P.; Bergsten, U. Comparison of fatty and resin acid composition in boreal Lodgepole pine and Scots pine for biorefinery applications. Ind. Crops Prod. 2013, 49, 535–541. [Google Scholar] [CrossRef]
- Yildirim, H.; Holmbom, B. Investigations on the wood extractives of pine species from Turkey: II Composition of fatty and resin acids in Pinus sylvestris and Pinus nigra. Acta Acad. Abo. 1977, B37, 1–6. [Google Scholar]
- Willför, S.; Hafizo-Glu, H.; Tümen, I.; Yazici, H.; Arfan, M.; Ali, M.; Holmbom, B. Extractives of Turkish and Pakistani tree species. Holz Roh. Werkst. 2007, 65, 215–221. [Google Scholar] [CrossRef]
- Lewinsohn, E.; Savage, T.J.; Gijzen, M.; Croteau, R. Simultaneous analysis of monoterpenes and diterpenoids of conifer oleoresin. Phytochem. Anal. 1993, 4, 220–225. [Google Scholar] [CrossRef]
- Benouadah, N.; Pranovich, A.; Aliouche, D.; Hemming, J.; Smeds, A.; Willför, S. Analysis of extractives from Pinus halepensis and Eucalyptus camaldulensis as predominant trees in Algeria. Holzforschung 2018, 72, 97–104. [Google Scholar] [CrossRef]
- Anderson, A.B.; Riffer, R.; Wong, A. Monoterpenes, fatty and resin acids of Pinus contorta and Pinus attenuata. Phytochemistry 1969, 8, 2401–2403. [Google Scholar] [CrossRef]
- Uçar, G.; Fengel, D. Variation in composition of extractives from wood of Pinus nigra varieties. Phytochemistry 1995, 38, 877–880. [Google Scholar] [CrossRef]
- Uçar, G.; Balaban, M. Cyclohexane extracts of black pine wood naturally grown in eastern Thrace. Holz Roh. Werkst. 2002, 60, 34–40. [Google Scholar] [CrossRef]
- Harju, A.M.; Kainulainen, P.; Venäläinen, M.; Tiitta, M.; Viitanen, H. Differences in resin acid concentration between brown-rot resistant and susceptible Scots pine heartwood. Holzforschung 2002, 56, 479–486. [Google Scholar] [CrossRef]
- Ekeberg, D.; Flæte, P.-O.; Eikenes, M.; Fongen, M.; Naess-Andresen, C.F. Qualitative and quantitative determination of extractives in heartwood of Scots pine (Pinus sylvestris L.) by gas chromatography. J. Chromatogr. A 2006, 1109, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Papajannopoulos, A.D.; Song, Z.Q.; Liang, Z.Q.; Spanos, J.A. GC-MS analysis of oleoresin of three Greek pine species. Holz Roh. Werkst. 2001, 59, 443–446. [Google Scholar] [CrossRef]
- Lange, W.; Stevanović Janežić, T. Chemical composition of some Pinus sylvestris L. oleoresins from southern Serbia, Bosnia and Makedonia: Two entities of Scotch Pine present on the Balkan peninsula. Holzforschung 1993, 47, 207–212. [Google Scholar] [CrossRef]
- Matziris, D. Variation in cone production in a clonal seed orchard of black pine. Silvae Genet. 1993, 42, 136–141. [Google Scholar]
- Landucci, L.L.; Zinkel, D.F. The 1H and 13C-NMR spectra of the abietadienoic resin acids. Holzforschung 1991, 45, 341–346. [Google Scholar] [CrossRef]
- Muto, N.; Tomokuni, T.; Haramoto, M.; Tatemoto, H.; Nakanishi, T.; Inatomi, Y.; Murata, H.; Inada, A. Isolation of apoptosis- and differentiation-inducing substances toward human promyelocytic leukemia HL-60 cells from leaves of Juniperus taxifolia. Biosci. Biotechnol. Biochem. 2008, 72, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Olate, V.R.; Usandizaga, O.G.; Schmeda-Hirschmann, G. Resin diterpenes from Austrocedrus chilensis. Molecules 2011, 16, 10653–10667. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the extracts are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
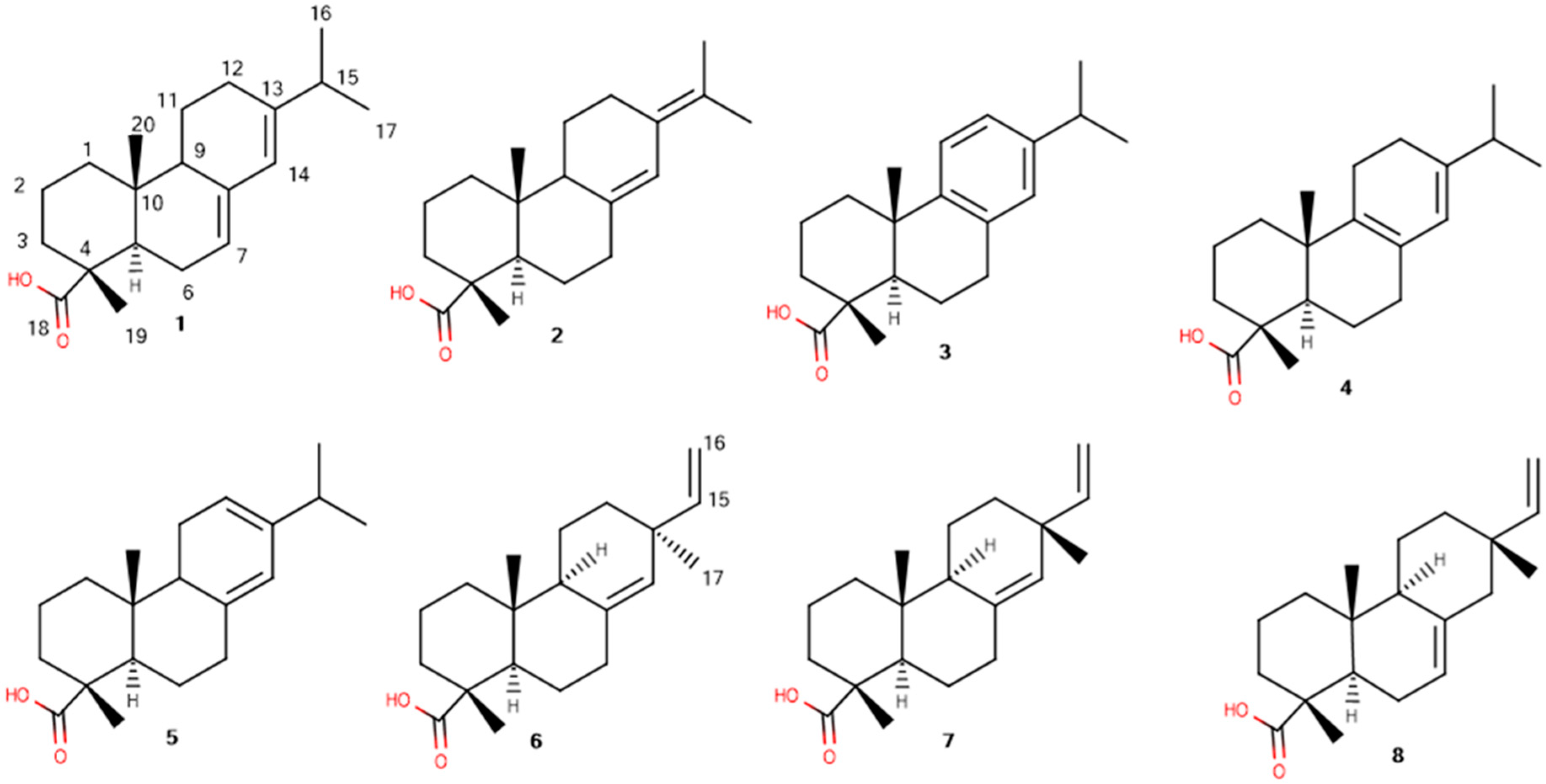
| Resin Acid | Abietane Type | Pimarane Type | ||||||
|---|---|---|---|---|---|---|---|---|
| Proton | Abietic Acid (1) | Neoabietic Acid (2) | Dehydroabietic Acid (3) | Palustric Acid (4) | Levopimaric Acid (5) | Pimaric Acid (6) | Sandaracopimaric Acid (7) | Isopimaric Acid (8) |
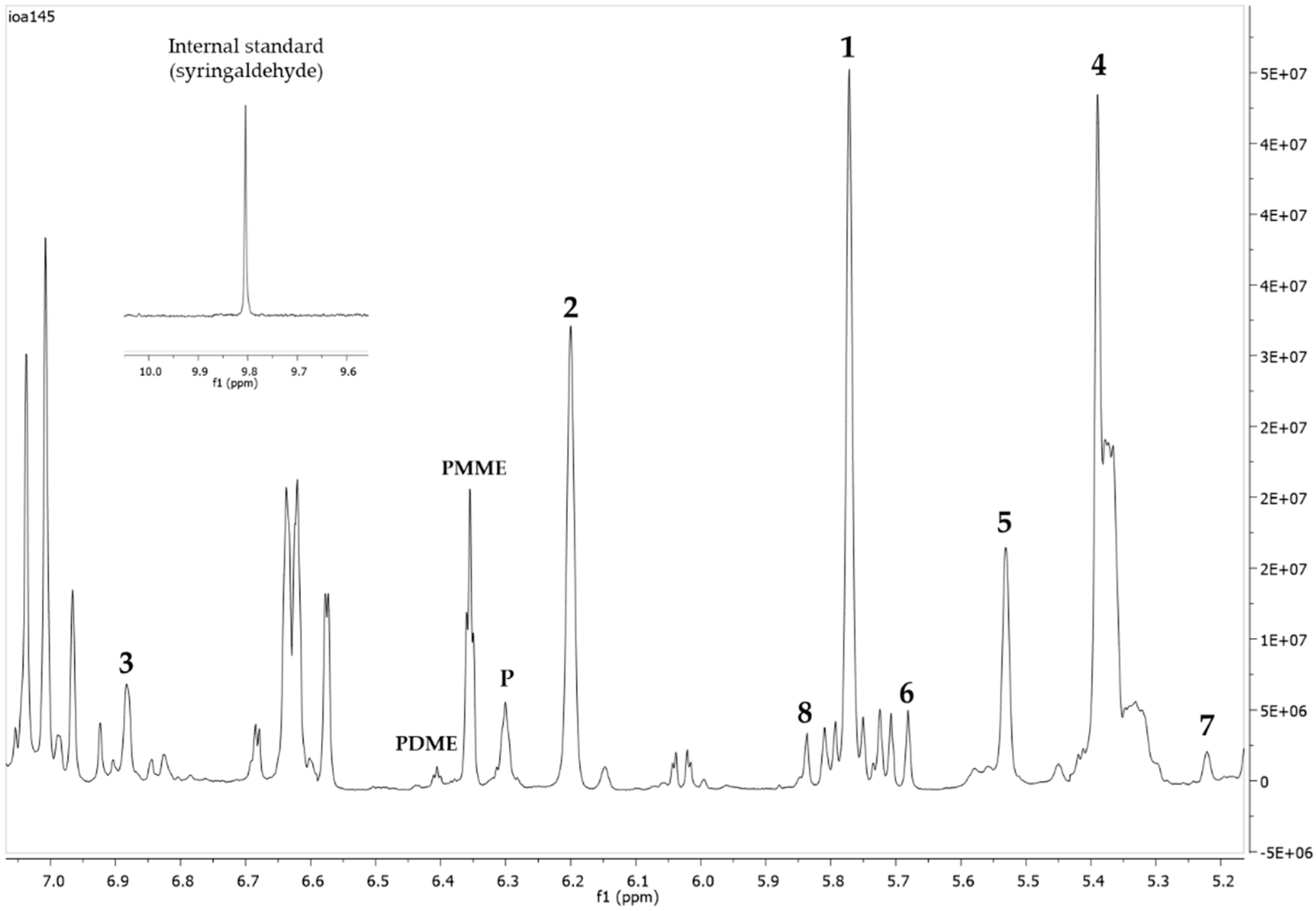
| C(H)-15 | - | - | - | - | - | 5.67 | - | 5.84 |
| C(H)-14 | 5.77 | 6.20 | 6.88 | 5.39 | 5.53 | - | 5.22 | - |
| Abietic Acid (1) | Neoabietic Acid (2) | Dehydroabietic Acid (3) | Palustric Acid (4) | Levopimaric Acid (5) | Pimaric Acid (6) | Sandaracopimaric Acid (7) | Isopimaric Acid (8) | |
|---|---|---|---|---|---|---|---|---|
| Recovery (%) | 90.26 | 93.52 | 88.78 | 90.50 | 90.73 | 92.75 | 92.07 | 87.37 |
| Coefficient of Variance (%) | 1.06 | 1.82 | 0.85 | 2.39 | 8.40 | 1.93 | 3.82 | 1.48 |
| ID | Abietic Acid (1) | Neoabietic Acid (2) | Dehydroabietic Acid (3) | Palustric Acid (4) | Levopimaric Acid (5) | Pimaric Acid (6) | Sandaracopimaric Acid (7) | Isopimaric Acid (8) |
|---|---|---|---|---|---|---|---|---|
| Sample 1 | 5.55 | 6.29 | 8.84 | 2.17 | 9.07 | 5.09 | 9.82 | 9.07 |
| Sample 2 | 6.51 | 7.38 | 3.49 | 5.18 | 4.85 | 7.55 | 2.88 | 3.31 |
| Sample 3 | 3.68 | 5.40 | 2.48 | 9.92 | 8.81 | 7.66 | 2.98 | 7.57 |
| Average | 5.25 | 6.36 | 4.93 | 5.76 | 7.58 | 6.77 | 5.22 | 6.65 |
| Abietic Acid (1) | Neoabietic Acid (2) | Dehydroabietic Acid (3) | Palustric Acid (4) | Levopimaric Acid (5) | Pimaric Acid (6) | Sandaracopimaric Acid (7) | Isopimaric Acid (8) | Total Resin Acids | |
|---|---|---|---|---|---|---|---|---|---|
| Average mg/gdhw | 76.77 | 39.34 | 11.69 | 47.94 | 8.07 | 22.54 | 2.72 | 10.91 | 219.98 |
| Min. mg/gdhw | 7.00 | 2.91 | 2.56 | 9.76 | 0.08 | 2.20 | 0.16 | 0.50 | 30.05 |
| Max. mg/gdhw | 181.75 | 101.82 | 38.59 | 105.22 | 64.91 | 59.42 | 6.67 | 34.09 | 424.70 |
| Provenance | Abietic Acid (1) | Neoabietic Acid (2) | Dehydroabietic Acid (3) | Palustric Acid (4) | Levopimaric Acid (5) | Pimaric Acid (6) | Sandaracopimaric Acid (7) | Isopimaric Acid (8) | Total Resin Acids |
|---|---|---|---|---|---|---|---|---|---|
| Zarouhla | 81.99 a | 40.06 a | 11.10 a | 48.83 a | 7.41 a | 21.60 ab | 2.61 a | 11.45 a | 224.46 a |
| Feneos | 78.64 a | 37.84 a | 12.29 a | 46.43 a | 8.91 a | 25.09 a | 2.74 a | 11.03 a | 222.768 a |
| Parnonas | 79.99 ab | 41.87 a | 11.74 a | 49.86 a | 6.88 a | 23.57 ab | 2.92 a | 11.20 a | 227.83 a |
| Taigetos | 66.05 b | 36.36 a | 11.82 a | 45.61 a | 9.69 a | 20.35 b | 2.54 a | 9.89 a | 201.25 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ioannidis, K.; Melliou, E.; Magiatis, P. High-Throughput 1H-Nuclear Magnetic Resonance-Based Screening for the Identification and Quantification of Heartwood Diterpenic Acids in Four Black Pine (Pinus nigra Arn.) Marginal Provenances in Greece. Molecules 2019, 24, 3603. https://doi.org/10.3390/molecules24193603
Ioannidis K, Melliou E, Magiatis P. High-Throughput 1H-Nuclear Magnetic Resonance-Based Screening for the Identification and Quantification of Heartwood Diterpenic Acids in Four Black Pine (Pinus nigra Arn.) Marginal Provenances in Greece. Molecules. 2019; 24(19):3603. https://doi.org/10.3390/molecules24193603
Chicago/Turabian StyleIoannidis, Kostas, Eleni Melliou, and Prokopios Magiatis. 2019. "High-Throughput 1H-Nuclear Magnetic Resonance-Based Screening for the Identification and Quantification of Heartwood Diterpenic Acids in Four Black Pine (Pinus nigra Arn.) Marginal Provenances in Greece" Molecules 24, no. 19: 3603. https://doi.org/10.3390/molecules24193603