
Metabolic Profiling of Alpinetin in Rat Plasma, Urine, Bile and Feces after Intragastric Administration


Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Drug Administration and Animals
2.3. Instrument and UPLC-Q-TOF-MS Conditions
2.4. Sample Preparation
2.5. Data Analysis
3. Results
3.1. Mass Fragmentation of Alpinetin
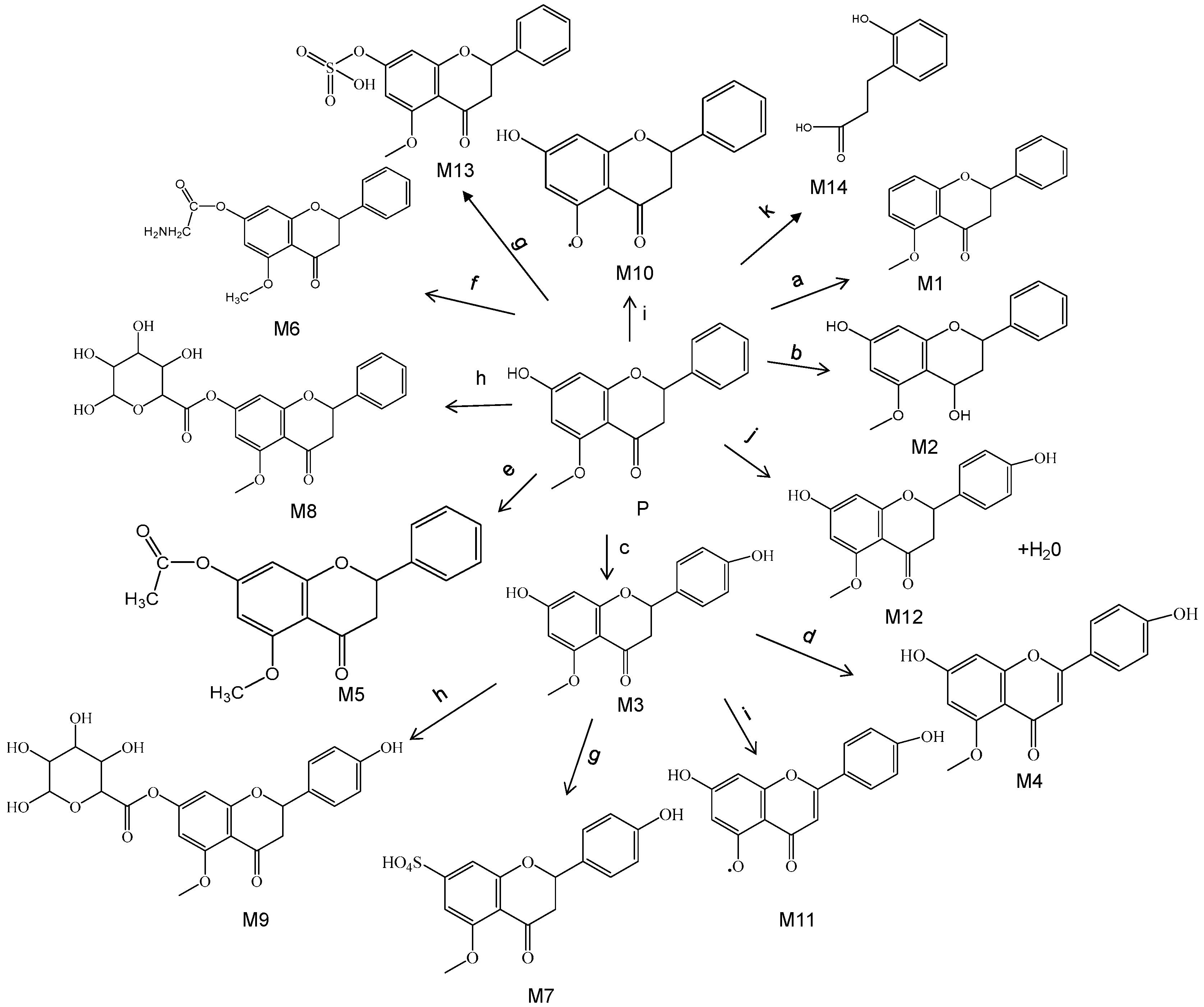
3.2. Identification of Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liu:, L.; Chen, X.; Hu, Z. Separation and determination of alpinetin and cardamonin in Alpinia katsumadai Hayata by flow injection-micellar electrokinetic chromatography. Talanta 2007, 71, 155–159. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, L.-S.; Yan, L.; Ren, J.; Zhou, D.-X.; Li, S.-S. Alpinetin inhibits lipopolysaccharide-induced acute kidney injury in mice. Int. Immunopharmacol. 2015, 28, 1003–1008. [Google Scholar]
- Lv, Q.; Shi, C.; Qiao, S.; Cao, N.; Guan, C.; Dai, Y.; Wei, Z. Alpinetin exerts anti-colitis efficacy by activating AhR, regulating miR-302/DNMT-1/CREB signals, and therefore promoting Treg differentiation. Cell Death Dis. 2018, 9, 890. [Google Scholar] [CrossRef]
- Liang, Y.; Shen, T.; Ming, Q.; Han, G.; Zhang, Y.; Liang, J.; Zhu, D. Alpinetin ameliorates inflammatory response in LPS-induced endometritis in mice. Int. Immunopharmacol. 2018, 62, 309–312. [Google Scholar] [CrossRef]
- Tan, Y.; Zheng, C. Effects of alpinetin on intestinal barrier function, inflammation and oxidative stress in dextran sulfate sodium-induced ulcerative colitis mice. Am. J. Med. Sci. 2018, 355, 377–386. [Google Scholar] [CrossRef]
- He, X.; Wei, Z.; Wang, J.; Kou, J.; Liu, W.; Fu, Y.; Yang, Z. Alpinetin attenuates inflammatory responses by suppressing TLR4 and NLRP3 signaling pathways in DSS-induced acute colitis. Sci. Rep. 2016, 6, 28370. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, X.; Shen, J.; Hua, D. Alpinetin inhibits proliferation and migration of ovarian cancer cells via suppression of STAT3 signaling. Mol. Med. Rep. 2018, 18, 4030–4036. [Google Scholar] [CrossRef] [Green Version]
- Malami, I.; Abdul, A.B.; Abdullah, R.; Kassim, N.K.; Rosli, R.; Yeap, S.K.; Waziri, P.; Etti, I.C.; Bello, M.B. Crude extracts, flavokawain B and alpinetin compounds from the rhizome of alpinia mutica induce cell death via UCK2 enzyme inhibition and in turn reduce 18S rRNA biosynthesis in HT-29 cells. PLoS ONE 2017, 12, e0170233. [Google Scholar]
- Wang, J.; Yan, Z.; Liu, X.; Che, S.; Wang, C.; Yao, W. Alpinetin targets glioma stem cells by suppressing Notch pathway. Tumour Biol. 2016, 37, 9243–9248. [Google Scholar] [CrossRef]
- Wu, L.; Yang, W.; Zhang, S.N.; Lu, J.B. Alpinetin inhibits lung cancer progression and elevates sensitization drug-resistant lung cancer cells to cis-diammined dichloridoplatium. Drug Des. Devel. Ther. 2015, 9, 6119–6127. [Google Scholar] [Green Version]
- Zhou, Y.; Ding, Y.L.; Zhang, J.L.; Zhang, P.; Wang, J.Q.; Li, Z.H. Alpinetin improved high fat diet-induced non-alcoholic fatty liver disease (NAFLD) through improving oxidative stress, inflammatory response and lipid metabolism. Biomed. Pharmacother. 2018, 97, 1397–1408. [Google Scholar] [CrossRef]
- Liu, T.G.; Sha, K.H.; Zhang, L.G.; Liu, X.X.; Yang, F.; Cheng, J.Y. Protective effects of alpinetin on lipopolysaccharide/d-Galactosamine-induced liver injury through inhibiting inflammatory and oxidative responses. Microb. Pathog. 2019, 126, 239–244. [Google Scholar] [CrossRef]
- Liu, E.Y.L.; Xu, M.L.; Xia, Y.; Kong, X.; Wu, Q.; Dong, T.T.X.; Tsim, K.W.K. Activation of G protein-coupled receptor 30 by flavonoids leads to expression of acetylcholinesterase in cultured PC12cells. Chem. Biol. Interact. 2019, 306, 147–151. [Google Scholar] [CrossRef]
- Hu, K.; Liu, L.; Qian, H.; Zhou, T.; Li, Y.; Yu, J.; Tan, B. Alpinetin promotes the binding of PPAR and methyltransferase. Xi bao yu fen zi mian yi xue za zhi = Chin. J. Cell. Mol. Immunol. 2017, 33, 1610–1614. [Google Scholar]
- Bi, S.; Sun, X.; Wang, Y.; Wu, J.; Zhou, H. A sensitive resonance Rayleigh light scattering method for alpinetin using gold nanorods probes. Luminescence 2018, 33, 1164–1170. [Google Scholar] [CrossRef]
- Wang, Z.R.; Li, Y.; Tang, B. Protection Mechanism of Alpinetin on Pulmonary Microvascular Endothelial Cells Injury. Zhongguo Zhong Xi Yi Jie He Za Zhi 2017, 37, 476–479. [Google Scholar]
- Ming-Jiang, W.U.; Xiao-Lei, W.U.; Zhang, D.Q.; Ding, L.Q.; Qiu, F.; Hao-Ling, M.A.; Chen, X.Z. Identification of in vivo substances of rats after oral administration of mulberry leaves flavonoids by UPLC-Q-TOF/MS. Chin. Tradit. Herb. Drugs 2017. [Google Scholar]
- Qi, C.; Fu, J.; Zhao, H.; Xing, H.; Dong, D.; Wu, B. Identification of UGTs and BCRP as potential pharmacokinetic determinants of the natural flavonoid alpinetin. Xenobiotica 2019, 49, 276–283. [Google Scholar] [CrossRef]
- Ye, W.; Lin, X.; Zhang, Y.; Xu, Y.; Sun, R.; Wen, C.; Wang, X.; Bao, S.; Chen, R. Quantification and pharmacokinetics of alpinetin in rat plasma by UHPLC-MS/MS using protein precipitation coupled with dilution approach to eliminate matrix effects. J. Pharm. Biomed. Anal. 2018, 152, 242–247. [Google Scholar] [CrossRef]
- Jin, X.L.; Fang, Z.Z.; Yan-Qing, Q.U.; Tang, B.; Yang, L.; Wang, L.M. Study on the glucuronidation of alpinetin in human liver microsomes. Chin. J. Clin. Pharmacol. 2011. [Google Scholar]
- Liang, S.C.; Ge, G.B.; Liu, H.X.; Shang, H.T.; Wei, H.; Fang, Z.Z.; Zhu, L.L.; Mao, Y.X.; Yang, L. Determination of propofol UDP-glucuronosyltransferase (UGT) activities in hepatic microsomes from different species by UFLC-ESI-MS. J. Pharm. Biomed. Anal. 2011, 54, 236–241. [Google Scholar] [CrossRef]
- Jaeger, B.N.; Parylak, S.L.; Gage, F.H. Mechanisms of dietary flavonoid action in neuronal function and neuroinflammation. Mol. Aspects Med. 2018, 61, 50–62. [Google Scholar] [CrossRef]
- Yin, J.; Ma, Y.; Liang, C.; Gao, J.; Wang, H.; Zhang, L. A systematic study of the metabolites of dietary acacetin in vivo and in vitro based on UHPLC-Q-TOF-MS/MS analysis. J. Agric. Food Chem. 2019, 67, 5530–5543. [Google Scholar] [CrossRef]
- Tanaka, S.; Trakooncharoenvit, A.; Nishikawa, M.; Ikushiro, S.; Hara, H. Comprehensive analyses of quercetin conjugates by LC/MS/MS revealed that isorhamnetin-7-O-glucuronide-4′-O-sulfate is a major metabolite in plasma of rats fed with quercetin glucosides. J. Agric. Food Chem. 2019, 67, 4240–4249. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
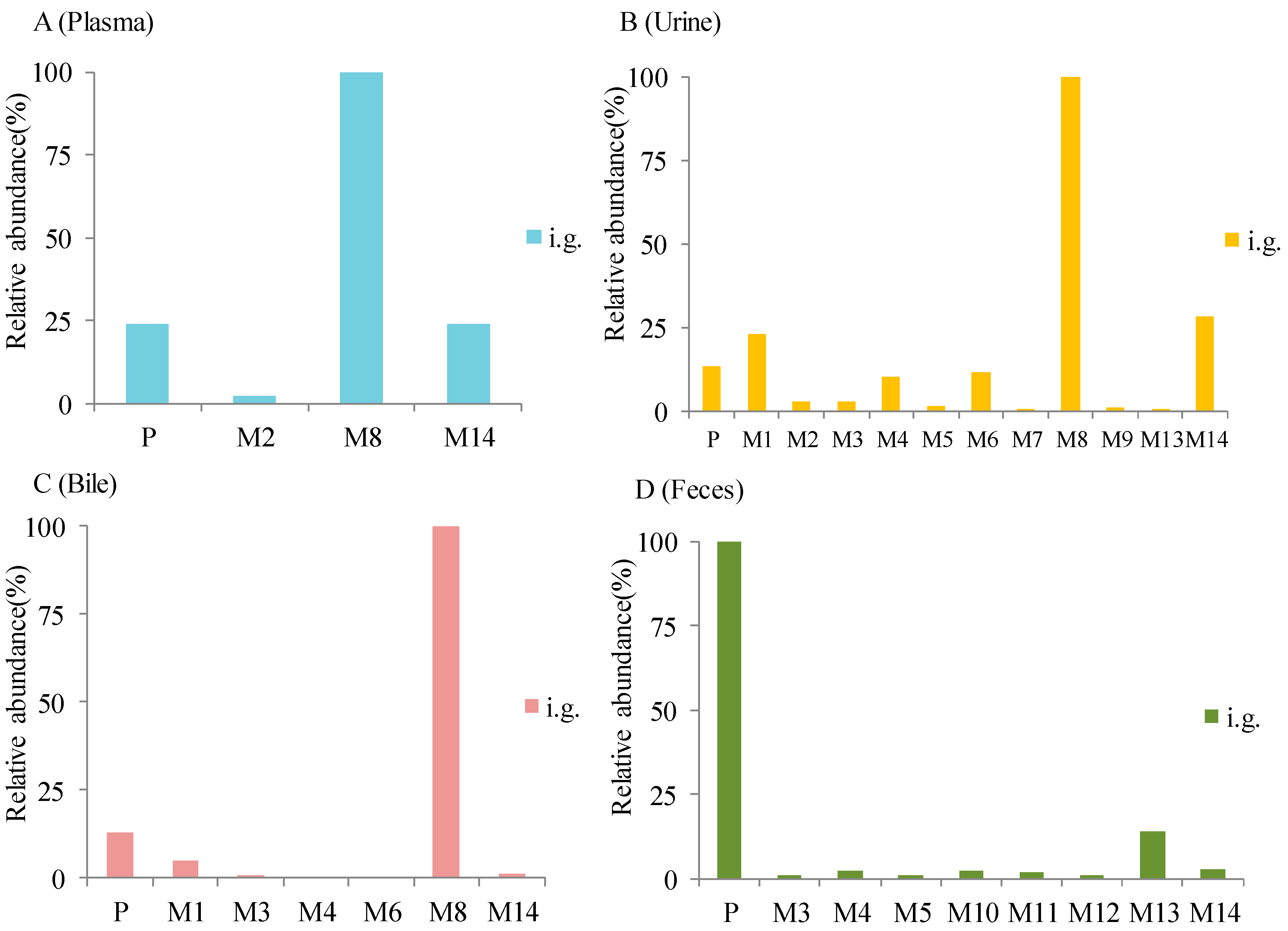{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | RT | m/z | Formula | Polarity | Error (×10−6) | MS2 | Metabolism |
|---|---|---|---|---|---|---|---|
| P | 16.52 | 271.0948 | C16H15O4 | POS | −3.27 | 257,167,104 | Prototype |
| M1 | 14.98 | 255.1017 | C16H15O3 | POS | 0.39 | 151,136,104 | Deoxydation |
| M2 | 13.41 | 273.1131 | C16H17O4 | POS | 3.30 | 241,225,169,104 | Reduction |
| M3 | 13.28 | 287.0929 | C16H15O5 | POS | 5.23 | 271,255,104,167 | Oxidation |
| M4 | 15.29 | 285.0735 | C16H1305 | POS | −4.07 | 269,253,167,118 | Oxidation + desaturation |
| M5 | 15.47 | 313.1051 | C18H1705 | POS | −0.64 | 271,209,104,167 | Acelylation |
| M6 | 15.69 | 328.1169 | C18H1805N | POS | −3.35 | 271,257,104,167 | Glycine conjugation |
| M7 | 16.65 | 367.0467 | C16H15O8S | POS | −4.08 | 353,287,271,104,120,167 | Oxidation + sulfation |
| M8 | 13.94 | 447.1283 | C22H23O10 | POS | −0.67 | 271,104,167 | Glucuronide conjugation |
| M9 | 15.39 | 463.1230 | C22H23O11 | POS | −1.08 | 287,271,104,167 | Oxidation + glucuronide conjugation |
| M10 | 11.79 | 257.0806 | C15H13O4 | POS | −0.78 | 228,104,153 | Demethylation |
| M11 | 15.96 | 273.0741 | C15H13O5 | POS | −4.22 | 255,241,152 | Oxidation + demethylation |
| M12 | 11.21 | 289.1072 | C16H17O5 | POS | 0.34 | 271,255,104,167 | Hydration |
| M13 | 15.55 | 351.0520 | C16H15O7S | POS | 1.34 | 333,255 | Sulfation |
| M14 | 2.36 | 166.0468 | C9H10O3 | POS | −2.56 | 148,132,104 | Phenolic acids |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, J.; Wu, H.; Feng, F.; He, X.; Wang, C.; Chu, S.; Xiang, Z. Metabolic Profiling of Alpinetin in Rat Plasma, Urine, Bile and Feces after Intragastric Administration. Molecules 2019, 24, 3458. https://doi.org/10.3390/molecules24193458
Qiu J, Wu H, Feng F, He X, Wang C, Chu S, Xiang Z. Metabolic Profiling of Alpinetin in Rat Plasma, Urine, Bile and Feces after Intragastric Administration. Molecules. 2019; 24(19):3458. https://doi.org/10.3390/molecules24193458
Chicago/Turabian StyleQiu, Jieying, Hongyu Wu, Feng Feng, Xiaoying He, Caihong Wang, Shenghui Chu, and Zheng Xiang. 2019. "Metabolic Profiling of Alpinetin in Rat Plasma, Urine, Bile and Feces after Intragastric Administration" Molecules 24, no. 19: 3458. https://doi.org/10.3390/molecules24193458
APA StyleQiu, J., Wu, H., Feng, F., He, X., Wang, C., Chu, S., & Xiang, Z. (2019). Metabolic Profiling of Alpinetin in Rat Plasma, Urine, Bile and Feces after Intragastric Administration. Molecules, 24(19), 3458. https://doi.org/10.3390/molecules24193458