1. Introduction
In settings of agriculture, botany, politics, and medicine, much controversy has emerged on the overuse and risks of herbicides in agricultural production, and discussions have recently focused on the development of biological controls using an allelopathy approach. Allelopathy is a biological phenomenon by which one species produces phytochemicals that affect the growth and development of other nearby plant species [
1]. These phytochemicals, also called allelochemicals, are produced principally as secondary metabolites of plants and microorganisms [
2]. As natural toxins, allelochemicals affect a target plant by morphological, cytological, physiological, and biochemical changes in plant aerial parts and roots [
3]. They inhibit cellular processes in target plants such as cell division, membrane permeability, stomatal closure, absorption of nutrients, photosynthesis, ATP synthesis, metabolic processes and gene expression [
4].
Some secondary metabolites produced by fungi are also allelochemicals [
5,
6,
7,
8]. The phytotoxicity of fungi is dependent on the quantity, strength and variety of allelochemicals [
9]. Utilization of phytochemicals from fungi for weed control in agriculture has received increasing attention [
10]. Fungi containing phytotoxic compounds might be a promising natural tool to manage weeds and pathogenic infestation [
9]. Therefore, isolation and identification of the active herbicidal substances from fungal metabolites warrant further efforts.
Cordyceps militaris (L.) Link is a fungus belonging to the class Ascomycetes [
11]. This fungus exhibits a wide spectrum of pharmacological activities, including anti-stress, antifatigue [
12], antioxidant [
13], antifungal, and anticancer [
14]. The fruiting body of
C. militaris contains various active components such as cordycepin, adenosine, polysaccharides, fatty acids, amino acids, and other chemicals [
15]. Among of them, cordycepin, a nucleoside analog (3′-deoxyadenosine), exhibits the most biological and pharmacological functions such as antineoplastic, antioxidant, and anti-inflammatory activities, tumor cell apoptosis and a decrease in tumor cell proliferation [
16,
17,
18].
To compare the phytotoxic power of isolated natural phytotoxins, benzoic acid (BA) is commonly used as a standard control [
19]. BA is applied as a commercial herbicide [
20] and is reported to modify in indicator plants physiological processes such as the net photosynthetic rate, stomatal conductance, nutrient uptake, and resulted in growth inhibition [
21,
22,
23]. Recently, Andriana et al. [
24] demonstrated that BA inhibited the germination of radish at a concentration of 1.0 mg/mL in bioassays.
Our present study evaluates the phytotoxic potential of C. militaris on R. sativus (radish) compared with BA in a search for nature-based alternatives for disputed herbicides, such as glyphosate. Results of isolation, application of different extraction methods to obtain a maximum yield of cordycepin, and bioassay-guided phytotoxicity evaluations have been provided. More specifically, the inhibition of cordycepin on physiological and biochemical responses of radish seedlings including total phenolic contents (TPC), total flavonoid contents (TFC), chlorophyll and carotenoid contents, electrolyte leakage, lipid peroxidation, and proline contents were also investigated.
3. Discussion
The present study evaluated plant-derived alternatives for globally used agrochemicals, including herbicides such as glyphosate, which has recently been debated and removed from the market in some countries [
26]. The focus is on cordycepin, and details were presented on a variety of isolation methods and herbicide characteristics (
Figure 1,
Figure 2,
Figure 3,
Figure 4,
Figure 5,
Figure 6,
Figure 7,
Figure 8 and
Figure 9,
Table 1,
Table 2,
Table 3,
Table 4 and
Table 5). This study thereby expands previous studies and experience in the area of botanical science including herbicide properties of plants [
1,
9,
26]. Consensus exists among agricultural scientists that chemical herbicides should help provide the world population with enough crops that are free of toxic residues and well tolerated by consumers without causing any health problems. Under these premises, allelopathy can become a promising tool for the sustainable development of agricultural production due to the inhibition of weed growth associated with the prevention of weed resistance to toxic chemicals [
27,
28]. Released from allelopathic plants, allelochemicals such as phenolics, momilactones, alkaloids, carbohydrates, purines, nucleosides, and amino acids are important sources for natural herbicide development [
25,
28,
29]. Thus, natural products from herbal plants may help decrease the use of synthetic herbicides for weed management, reduce pollution, and provide safer agricultural products of high quality [
30].
Experiments of the current study revealed that EtOAc extract of
C. militaris had the maximum inhibitory effects on the germination, shoot height and root length of radish, as compared to hexane, chloroform, and aqueous residues (
Table 1). This suggests that EtOAc extract of fruiting bodies of
C. militaris might contain principal allelochemicals. In fact, extraction with various suitable solvents can also provide high yields of potent allelochemicals [
29]. Among fractions separated by column chromatography eluted by chloroform and methanol (10:0 to 0:10
v/
v), CM4 was the most active fraction to inhibit the germination and growth of radish (
Table 2). The analyses of GC-MS and LC-ESI-MS confirmed that cordycepin was the major compound in CM4 (55.38%), followed by pentadecanal (19.8%) (
Table 4). Immersion of methanol for two weeks provided more cordycepin than immersion in hot water (70–100 °C) (
Table 5), and it was found that repeat boiling of the
C. militaris fruiting bodies apparently provided greater yields of cordycepin than a single boil. Commonly, cordycepin is isolated from a liquid medium of
C. militaris. In the present study, we isolated cordycepin from the fruiting body of
C. militaris. Compared with the previous study that isolated cordycepin from the fruiting body of
C. militaris [
31] by high-speed countercurrent chromatography, our method is more simple and provides a higher yield.
Several factors, such as the type of solvents, temperature, sonication, and extraction time affect the yield of bioactive compounds from plant tissues [
32]. In this study, solvent polarity and extraction time may affect the yield of cordycepin in fruiting body of
C. militaris. Additional solvents combined with different extracting techniques should be tested to receive maximum yields of cordycepin from
C. militaris.Allelochemicals inhibit plants directly by affecting their morphology, physiology, and biochemistry [
4]. In this study, BA, synthetic cordycepin, and the CM4 fraction containing cordycepin significantly reduced the amount of chlorophyll (a, b, total chlorophylls) and carotenoid of radish as compared with control (
Figure 2). Kaya et al. [
33] reported that BA reduced pigment contents and decreased the photosynthesis rate. In this study, synthetic cordycepin decreased 2.0- to 9.5-fold the pigment contents of radish compared to BA (
Figure 2). These findings suggest that cordycepin possesses stronger phytotoxic activity than BA. Similar to other allelochemicals, both BA and cordycepin may affect porphyrin, a precursor for chlorophyll biosynthesis [
34]. Reduction of chlorophylls under allelochemical stress may be caused by impaired chlorophyll biosynthesis, the stimulation of pigment degradation, or both [
35].
There are many physiological and biochemical indicators that can be employed to understand how allelochemicals inhibit the receiver plant. Jaballah et al. [
36] mentioned that electrolyte leakage, lipid peroxidation, pigment, proline, total phenolic and flavonoid contents are common indicators in response to allelochemical stress. Electrolyte leakage (EL) is one of the strongest indicators of membrane damage in plants effected by allelochemical stress [
19]. Increasing membrane permeability could be due to peroxidation of polyunsaturated fatty acids in the bio-membranes, leading to a variety of products including malondialdehyde (MDA) [
36]. In this study, MDA accumulation and electrolyte leakage of radish were increased in both the roots and aerial parts when treated with the CM4 fraction containing cordycepin, as well as BA. Chen et al. [
37] reported that BA destructs the cell membrane’s integrity due to the formation of free radicals. In this study, cordycepin increased the leakage percentage and MDA accumulation of radish more than BA (
Figure 3 and
Figure 4).
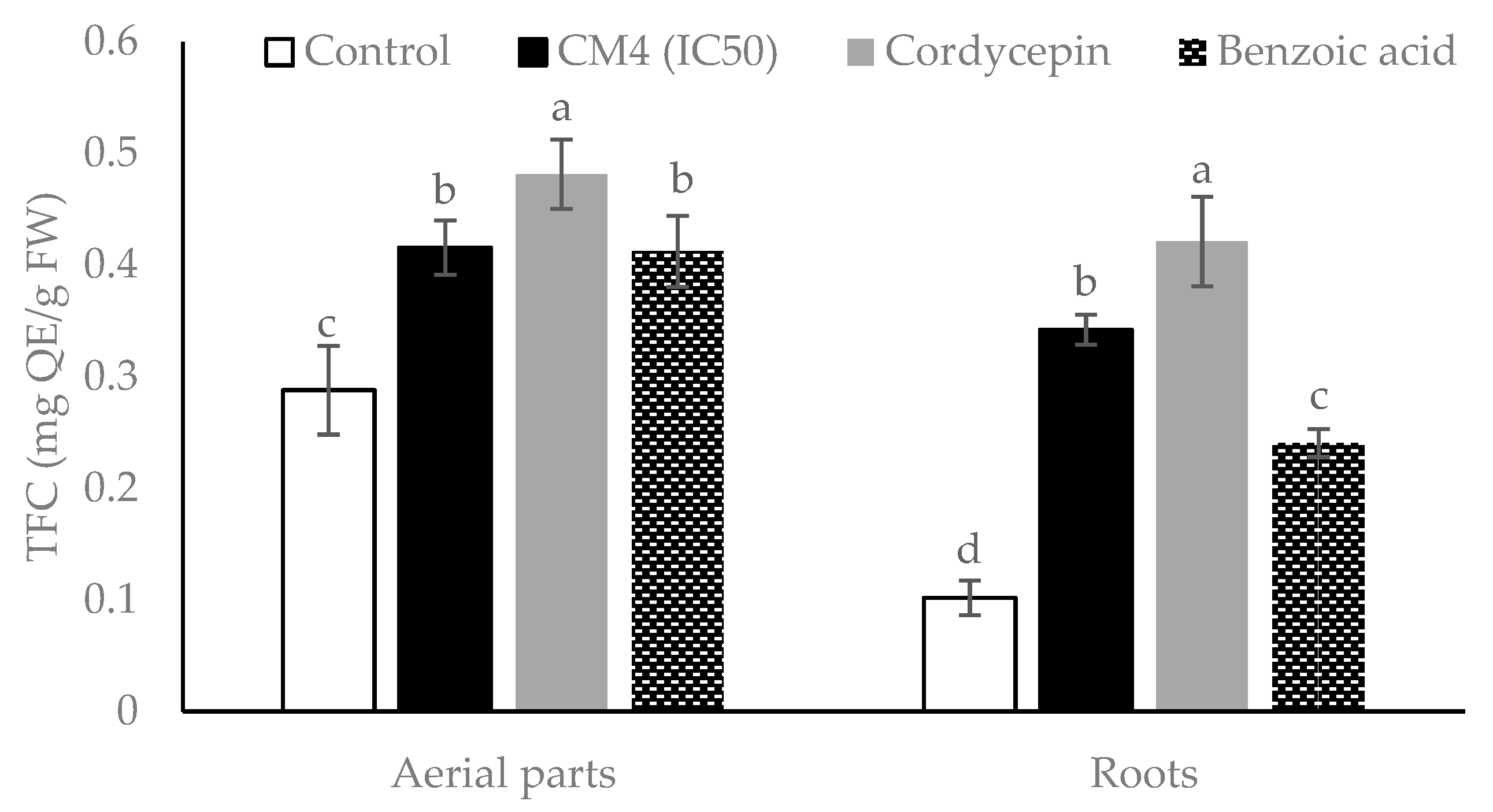
Total phenolic and total flavonoid contents as well as the proline accumulation in radish, was significantly increased as compared to the control by BA, CM4, and synthetic cordycepin (
Figure 5,
Figure 6 and
Figure 7). Secondary metabolites from plants, such as total phenolics and flavonoids, are structural components of cell walls and participate in defense mechanisms of plants against abiotic and biotic stressors [
38]. Ladhari et al. [
3] reported that allelochemicals from aqueous and methanolic extracts increase the accumulation of proline in the roots and leaves of lettuce. Flavonoids are the predominant phenolic compounds with important roles as potential inhibitors of the lipoxygenase enzyme, which converts polyunsaturated fatty acid to oxygen-containing derivatives. They accumulate in organs of plants and may help inhibit the process of lipid peroxidation in plants under stress [
38]. Similarly, the aerial parts of radish accumulated more proline than the roots. These results are in line with previous research, which reported that the aqueous extract of corn leaves augmented the proline content in wheat leaves [
39]. In addition, BA increased the proline content in leaves of tomato seedlings [
19] and wheat seedlings [
40]. Similarly, in this study synthetic cordycepin induced proline accumulation more than BA in both the roots and aerial parts (
Figure 7). Proline synthesis is regulated by various types of stress to allow for the accumulation of proline, a common solute compatible with protective properties [
41]. Therefore, increasing the proline level in
R. sativus may be due to the phytotoxic effects of cordycepin, as well as BA (
Figure 7).
Phytotoxic capacity and physiological and biochemical responses of
R. sativus and some indicator plants to glyphosate have been reported. In a study [
42] treating radish plants at 2 mM doses for four days exposure, glyphosate stimulated the shoot height over the control by 20%, but when the dose of glyphosate was increased, this herbicide had a strong inhibitory effect. Compared to our results, synthetic cordycepin inhibited 50% of the shoot height of radish at 0.188 mM dose. Furthermore, in the greenhouse experiment, glyphosate needed more than 600 g/ha dose to decrease chlorophyll a and b of radish from 81.6% to 86.28%, compared with control [
43]. In other indicator plants and at a 10 mM dose, glyphosate had a negligible effect on proline accumulation in maize, but EL and MDA increased notably [
44]. Similarly, 0.5% (
v/
v) glyphosate concentration showed ineffective inhibition of cogongrass [
45]. Therefore, cordycepin is more phytotoxic and has a greater effect on the physiological and biochemical processes of receiver plants. With respect to the mode of action, cordycepin functions as an allelochemical compound by inhibiting the germination and growth of radish, reducing pigment synthesis (chlorophylls and carotenoids), stimulating electrolyte leakage, lipid peroxidation, and proline and total phenolic and flavonoid accumulation compared to BA (
Table 2;
Figure 3,
Figure 4,
Figure 5,
Figure 6 and
Figure 7). However, evaluating additional activities of cordycepin, including its functional groups -OH, -NH
2, and the presence of N on the C4, 7, and 9 of its chemical structure (
Figure 8) is required, and respective studies are in progress.
Cordyceps militaris is a highly valued edible fungus. It is in use as a dietary supplement in the US and elsewhere, belongs to the large group of herbal TCM (traditional Chinese medicine), and has likely been used safely for centuries in Eastern countries, particularly in China and South East Asia, to treat various diseases [
46]. Cordycepin has been well characterized in animal experiments regarding an in vivo subacute toxicity test [
47] and Ames test [
47], and it is characterized also by rapid decomposition in humans through adenosine deaminase, as evidenced by a short half-life of about 1 min [
47]. This likely reduces the possible risk of toxicity and tumor initiation. The chemical structure of cordycepin is very similar to adenosine, with exception of a missing hydroxyl group on carbon number 3 [
47]. Therefore, cordycepin can easily be synthetized from adenosine, a natural chemical present in all human cells [
48]. The isolation of cordycepin has been described in the literature using various methods [
11,
16,
17,
47,
48]. The present study improved the isolation procedure, provided a rapid isolation method, and conducted bioassay-guided isolation and identification of plant growth inhibitors from
C. militaris (
Figure 1 and
Figure 8;
Table 2), resulting in a detailed description of the inhibitory activity of this fungus with its active cordycepin (
Table 4). Due to its high water solubility, cordycepin may be problematic for its potential use as a herbicide if it contaminates the ground water. Therefore, respective studies are needed to clarify this issue; it may also be solved by synthesis of a less water-soluble derivate that is still rapidly decomposable in the soil. The overall question remains whether cordycepin can outperform currently used herbicides, which are presently under worldwide dispute despite an impressive list of study results contained in a technical fact sheet on glyphosate [
26], but further investigations on cordycepin are also required to exclude any health risk of humans and to carefully evaluate the benefit to risk ratio of cordycepin use. If by error entering the human body, cordycepin will be decomposed within a minute.
This study shows that Cordyceps militaris, with cordycepin as its active component, effectively inhibits plant growth and may be a promising natural source to develop plant-based herbicides. Presently, it is unclear whether cordycepin can outperform glyphosate. Further examinations on the effects of cordycepin on the growth of different and principal weeds in agricultural production, such as Echinochloa crus-galli and Bidens pilosa are needed. Cordycepin shows at 0.04 mg/mL greater inhibitory efficacy as compared with BA and is thereby basically a promising phytochemical. To clarify the mode of action of this compound, different concentrations of cordycepin as well as its synthesized derivatives, should be examined to detail the physiological and biochemical responses of indicator plants as well as agricultural weeds. The correlation between the phytotoxicity of cordycepin on radish with the corresponding levels of proline, chlorophylls and carotenoids, electrolyte leakage, lipid peroxidation, and total phenolics and flavonoids should be addressed. This would help clarify under what conditions cordycepin could safely and effectively be used as a herbicide.


 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}