
Galactosaminoglycans: Medical Applications and Drawbacks
by
 ,
,
Vitor H. Pomin
1,2,* ,
,
William P. Vignovich
1,
Alysia V. Gonzales
1,
Ariana A. Vasconcelos
3 and
Barbara Mulloy
4 1
Department of Biomolecular Sciences, School of Pharmacy, University of Mississippi, Oxford, MS 38677-1848, USA
2
Research Institute of Pharmaceutical Sciences, School of Pharmacy, University of Mississippi, Oxford, MS 38677-1848, USA
3
Institute of Medical Biochemistry Leopoldo de Meis, Federal University of Rio de Janeiro, Rio de Janeiro, RJ 21941-590, Brazil
4
Imperial College, Department of Medicine, Burlington Danes Building, Du Cane Road, London W12 0NN, UK
*
Author to whom correspondence should be addressed.
Molecules 2019, 24(15), 2803; https://doi.org/10.3390/molecules24152803
Submission received: 19 June 2019
/
Revised: 24 July 2019
/
Accepted: 30 July 2019
/
Published: 1 August 2019
(This article belongs to the Special Issue Emerging Frontiers in Glycosaminoglycans and Mimetics)
Abstract
:Galactosaminoglycans (GalAGs) are sulfated glycans composed of alternating N-acetylgalactosamine and uronic acid units. Uronic acid epimerization, sulfation patterns and fucosylation are modifications observed on these molecules. GalAGs have been extensively studied and exploited because of their multiple biomedical functions. Chondroitin sulfates (CSs), the main representative family of GalAGs, have been used in alternative therapy of joint pain/inflammation and osteoarthritis. The relatively novel fucosylated chondroitin sulfate (FCS), commonly found in sea cucumbers, has been screened in multiple systems in addition to its widely studied anticoagulant action. Biomedical properties of GalAGs are directly dependent on the sugar composition, presence or lack of fucose branches, as well as sulfation patterns. Although research interest in GalAGs has increased considerably over the three last decades, perhaps motivated by the parallel progress of glycomics, serious questions concerning the effectiveness and potential side effects of GalAGs have recently been raised. Doubts have centered particularly on the beneficial functions of CS-based therapeutic supplements and the potential harmful effects of FCS as similarly observed for oversulfated chondroitin sulfate, as a contaminant of heparin. Unexpected components were also detected in CS-based pharmaceutical preparations. This review therefore aims to offer a discussion on (1) the current and potential therapeutic applications of GalAGs, including those of unique features extracted from marine sources, and (2) the potential drawbacks of this class of molecules when applied to medicine.
1. Introduction
Galactosaminoglycan (GalAG) is a term specifically used to designate the classes and sub-classes of glycosaminoglycans (GAGs) that contain N-acetylgalactosamine (GalNAc) units as the hexosamine type in their repeating disaccharide building blocks. In addition to the GalNAc unit, the disaccharide blocks of these high molecular weight (MW) polymers are additionally composed of an uronic acid, which can be either β-d-glucuronic acid (GlcA) or its C5 epimer, α-l-iduronic acid (IdoA) [1]. Although the GalAGs are homogeneously composed of alternating 3-linked GalNAc and 4-linked IdoA/GlcA units, the backbones of these glycans are extensively modified by different levels of C5 epimerization [2], sulfation contents and patterns [3,4], and in certain cases, fucosylation [5,6] (Figure 1). As detailed below, the fucose decoration on the GalAG backbone can be further sulfated at different positions and degrees [6]. The principal representatives of GalAGs are the chondroitin sulfates (CSs), dermatan sulfate (DS) and fucosylated chondroitin sulfate (FucCS) (Figure 1). Like all GAGs, GalAGs are key structural players of living organisms and are found more commonly in the extracellular matrix (ECM) environment or attached to the cell surface than inside the cells.
The structural diversity of GalAGs directly reflects their great number of biological roles and medical uses. Examples of this wide spectrum of biological roles are cell growth and differentiation [8,9], morphogenesis [10,11], cell migration [12,13] and microbial infection [14]. Examples of medical effects in which these molecules participate are anticoagulation [15,16,17,18,19], antithrombosis [20,21], anti-inflammation [22,23,24], anticancer [24,25,26], antiviral [27,28,29,30], anti-angiogenesis [31,32] and antimalaria [33], in addition to some beneficial effects in events of hemodialysis [34], cellular growth modulation [35], fibrosis [36], hyperglycemia [37] and diabetes-related nephropathy [38]. Due to the many biological and medical functions of GalAGs, several scientific groups in the world are now conducting research on these molecules. Consequently, the number of publications regarding GalAGs has increased considerably in the last three decades. This growing interest in research about GalAGs has been accompanied by the boom of glycomics [39]. The main research goals of the GalAG-related works have been the isolation and structural characterization of novel compounds, screening of new biological and medical functions, advanced structural analysis, development of novel and more efficient sequencing and analytical methods, and glycoengineering processes for partial or full chemical synthesis of GalAG oligosaccharides in the laboratory. The achievements made in these works have enabled great progress concerning the details underlying the biomedical functions and physicochemical properties of GalAGs and placed these glycans in a special position in medicinal glycomics.
However, as opposed to the many articles that have been published so far reporting the beneficial effects of GalAGs in healthcare, a few, but key, studies are now exposing the lack or potential negative effects of these molecules. In this review, we not only offer an up-to-date overview regarding the main chemical, biological and clinical aspects of GalAGs from both common and marine sources, but also highlight the downsides of these glycans regarding their current and potential medical uses. This subject is worthy of special attention from the scientific community. Hence, it is worth discussing here the facts associated with the issues related to possible impurities during manufacturing, actual effectiveness and safety during the application of GalAGs as pharmaceutical ingredients, and their potential harmful effects already pointed out in some basic studies.
2. CS
2.1. Overview
CSs are a group of GalAGs composed of alternating β-d-GlcA and β-d-GalNAc units (Figure 1A) polymerized within relatively large (50–100 kDa) and heterogeneous chains consisting of a non-uniform distribution of different disaccharide units [3]. Figure 2 illustrates the 3D structure of this disaccharide building block of the CS backbone without characteristic sulfation. The disaccharide units of CSs are further sulfated and the consequent classification of the different CS types relies primarily on the variation of the different resulting sulfation patterns. Sulfation occurs on the hydroxyl groups on the C3 and C2 positions of the β-d-GlcA and C4 and C6 of the β-d-GalNAc (Figure 1A). The type of the CS polymer is defined by the major composing disaccharide building block although hybrid compositions are more common.
The different CS polymers are found in a species and tissue-specific manner and are correlated with their biological functions once sulfation patterns are known to impact directly on activity [41]. CSs are ubiquitously expressed in vertebrates but can also be found in invertebrates [42,43,44,45], especially marine species, which express CSs with unusual sulfation patterns [46,47] (Table 1). A case of a CS with a rare sulfation pattern not included in Table 1 is the CS found in the head of the shrimp Litopenaeus vannamei. This CS is disulfated at C2 and C3 positions of the GlcA unit [48]. As a main component of the ECM, CSs possess a diverse set of physiological functions, especially those necessary for tissue development and repair [49,50], inhibitory and stimulant controlling effects over the central nervous system [51,52,53], and maturation of the organisms [8,9]. Due to these general properties, CS has been widely exploited as an active ingredient in over-the-counter healthcare supplements, mainly for easing issues associated with osteoarthritis [54], especially joint (knee) pain and inflammation [55,56,57].
A common and rich biological source of CS is the cartilaginous tissues. CS-based dietary supplements are basically derived from terrestrial sources, which are easy to obtain as a byproduct of commercial agriculture from both bovine trachea and porcine (ear and nose) tissues, but shark fin, ray, crocodile and chicken keel are not entirely uncommon [63]. This, however, is a limiting approach to the utilization of these compounds as therapeutic drugs because of the lack of diversity within structures of the terrestrial sources which often contain mostly CS-A with lesser amounts of CS-C subunits [63]. In addition to the lack of chemical diversity, there is also a concern with safety associated with some oversulfated derivatives of these more traditional sources after the “Heparin crisis” of 2018 [64]. In fact, as discussed in more detail below, impurities, although harmless, have been reported in CS-based pharmaceutical preparations destined for oral administration [65]. All these factors have contributed to encouraging research for other CS sources of more diverse structural expression as well as safer manufacturing processes. Marine animals are exceptionally promising sources of more novel CS structures with unique sulfation patterns that are either not present or present in irrelevant amounts in terrestrial organisms. In addition to that, the marine sources have provided a robust safety profile, economic utilization and more ecological use of bycatch [46,47].
Although CS-A and CS-C (Figure 3) are still expressed in marine organisms, they do not possess the same observed and potential therapeutic applications outside of more commonplace use in osteoarthritis as the other CS sub-types (Figure 1A). CS-D, CS-E, and CS-K (Figure 3), and very rarely CS-L and CS-M (Table 1) have been shown to exhibit an array of other biomedical activities related to anti-inflammation [66,67], cancer and metastasis [25,68,69], neurology [52], and antipathogenesis [28]. These CS types are less commonly found than CS-A and CS-C in mammals, and more common in marine organisms (Table 1), but indications exist of their presence, although in minor portions, in tissues of terrestrial mammals [54,70,71].
The functions promoted by CS, regardless of the CS type, are ultimately related to the activity of functional GAG binding proteins (GBPs) [72]. The biological functions of these proteins are modulated by interactions with GAGs (GalAGs), which must contain the necessary structural moieties for binding and consequential regulation [73,74], mainly sulfation patterns. The optimal systemic concentration of the active carbohydrate is another crucial requirement [75,76].
2.2. Biomedical Properties
2.2.1. Inflammation
CS’s effects on inflammation have mostly been described in its usages in patients with osteoarthritis [22,23,77]. However, anti-inflammatory actions in multiple organs or systems, including the skin, liver, and digestive systems, have also been reported [78,79]. These effects are highly mediated by CS’s interactions with pro-inflammatory enzymes and transcription factors related to attenuation of inflammatory response [78]. Jeong-Sook Noh et al. demonstrated that in hepatic tissues CS-E produced noticeable protection against inflammatory and oxidative damage by lipopolysaccharides through down-regulation in TNF-α, IL-1β, COX-2, and iNOS hepatic inflammatory factors [66].
H. Kawashima et al. determined that CS-E has a strong binding affinity to both L- and P- selectins, as well as some chemokines [67]. The resultant CS interactions effectively produce inhibition of the activity of these proteins in recruitment of leukocytes to the inflamed sites during events of infection and inflammation. While this is an important immune response in infection prevention and in certain controlled inflammatory processes, it is also a key contributor in the pathology and progress of multiple autoimmune diseases like psoriasis and bowel disease [80]. The balanced activity of the inflammatory events is required to restore systemic homeostasis and this stage is very commonly not achieved after some auto-immune responses or pathological inflammatory processes. In these uncontrolled events of immune and/or inflammatory responses, medical intervention is needed to down-regulate the processes. CSs of various sulfation patterns have been a key agent in these complications [22,23,66,77,78,79].
Identification of the mechanisms by which the active carbohydrate-protein complexes are regulated and stabilized, and consequently how they function during the different stages of disease, is crucial to understanding the underlying biochemistry and pharmacology of GalAGs, and to fully exploring their medical properties, especially the CSs with diverse sulfation patterns (Figure 3, Table 1). In this regard, Xu Wang et al. characterized by advanced nuclear magnetic resonance (NMR) methods the binding properties of two CS hexasaccharides: one entirely 4-sulfated at the three composing disaccharides (CS444), and one with 6-sulfation only at the non-reducing end disaccharide (CS644), in complex with the pro-inflammatory CCL5/RANTES chemokine [81]. This slight change of sulfation patterns was enough to cause big changes in the interactions. The CS444 was shown to be the best ligand for interaction and consequential structural mapping of the CCL5:CS hexasaccharide complex.
2.2.2. Cancer and Metastasis
CS has, however, been shown to interact with pathways that enhance cancer growth and metastasis. Native CS-A has been shown to participate in proliferation and spread of some cancers [25]. CS-E, and likely other highly sulfated derivatives of CS, can, for example, negatively regulate the pro-tumorigenic Wnt/beta-catenin-Collagen 1 axis in breast cancer cells [82]. Inhibition of L- and P-selectins may have potential applications in treatments of some cancers where carcinoma cells bind platelets through recognition of the selectins and chemokines. Binding of platelets is an aspect of cancer metastasis that sequesters cancer cells from the blood to tissues [83,84]. Inhibition of selectins and chemokines can reduce lethality associated with certain cancers as these factors directly contribute to both the proliferation of tumor cells (cancer growth) and spread (metastasis). The use of exogenous CSs shows the ability to compete with native CSs that are involved in the development of cancer in different stages of progress of this devastating disease [25,68,69,82].
2.2.3. Neural Growth Stimulation and Inhibition
CS plays a key role in regulating processes during the development of the central nervous system [51]. CS’s activity is spatiotemporally dependent, acting as either stimulator in certain cases [85,86] or as inhibitor in others [86,87]. CS, together with keratan sulfate (KS), is necessary in the high-MW molecular assembly aggrecan to inhibit neurite outgrowth [88,89]. Curiously, of particular biological importance is the potential exogenous use of highly sulfated CS-E and CS-D to bind and inhibit neural growth factors that elicit outgrowth of hippocampal neurites [52]. The more sulfated CSs, like CS-D and CS-E, show mechanisms that are likely mediated through interactions between CS and midkine, a GBP that promotes growth, survival, and other activities in target cells [90]. Ueoka et al. showed that midkine binding of CS-E is comparable to heparin, as well as to CS-K [90]. The reasons for the similar binding properties of the different highly sulfated GAGs was further clarified. In the work of Maeda et al. [91], it was shown that the binding of CS to pleiotrophin, a GBP highly homologous to midkine, is regulated by highly sulfated sequences of approximately 20 monosaccharides in length. Pleiotrophin binds with good affinity to 6B4-PG/phosphacan, a highly expressed CS proteoglycan that is present in the brain [92,93]. Exogenous CS-E can compete with the 6B4-PG/phosphacan-CS binding [94]. This highlights the role of more sulfated CS units in the binding process and regulatory events. In certain cases, the inhibitory activity of CS in neural repair can be also attenuated with the use of digestive enzymes capable to cleave CS, such as chondroitinase [95]. Careful design of this proposed enzymatic therapy will be necessary to avoid potential disadvantages, as non-selective CS lyase activity will be unable to distinguish between CS chains which play an inhibitory role in the neural repair and those involved in homeostatic functions of the neural system.
2.2.4. Antiviral and Antibacterial Activity
The antimicrobial activity of GalAGs has been explored extensively, and for CS, results are promising regarding both antiviral and antibacterial applications. CS-E has displayed significant antiviral activities in all serotypes of Dengue fever [27]. CS-E has reduced infectivity of this virus through a similar action to heparin via interactions with the virus’ envelope proteins. It was observed that CS-D does not lead to the same quality of interactions, showing that in this case the position of sulfation is more important than the overall degree of sulfation [27]. It has also been shown that squid CS-E can produce antiviral activity for herpes simplex virus 1 and 2 with much higher success than heparin. The mechanism of action that is proposed is through a competition between CS-E and the attachment of the virus on the host cell’s GAG receptors [28]. Due to the relatively high MW of squid CS-E, this GalAG was not able to reduce the cell-to-cell spread within the monolayer of densely growing cells [28]. CS was also proposed as a new therapeutic target for modulating infection by Borrelia burgdorferi, the bacterium causative of lyme disease. This pathogen has a GAG binding site that is crucial for its spread through the tissues and consequential states of the disease related to the development of lyme arthritis [96].
3. DS
3.1. Overview
DS, also denominated CS-B, was first isolated in 1941 from porcine skin [97,98]. DS is found relatively easily but is not as common as CS-A and CS-C in animals. It is an important component of the ECM of the connective tissues. Like the CS backbone, DS is structurally composed of disaccharide building blocks, in this case of 4-linked IdoA and 3-linked GalNAc units, although GlcA units may also occur. The presence of the C5 epimerized IdoA in DS differentiates it from CS [99] (Figure 1). The 3D model of this building block is shown in Figure 4.
As for CS, the composing monosaccharides of DSs are differentially sulfated to varying degrees and positions depending on the tissues [100], cells [99] and pathophysiological conditions from which they are extracted [101]. Table 2 shows a few examples of DS sub-classes. For example, GalNAc units are mostly sulfated at the C4 position (~95%), but small amounts of sulfation (~15%) can also occur at different positions such as the C6 position of GalNAc and (~10%) at the C2 position of the IdoA [102,103]. The marine sources of DS, like for CSs, also offer unique structures to this GalAG. Interesting representatives are the DSs isolated from different species of ascidians (Urochordata) presenting distinct patterns and proportions of sulfation [99,104,105,106].
Like the diverse set of CSs with different sulfation patterns (Table 1), the structural heterogeneity of the DS molecules (Table 2) also promotes diversified biological functions [72,99], and contributes to the variety of biomedical roles played by DS. The ability of this GalAG to bind to and regulate multiple ECM proteins that are functional in numerous pathophysiological systems is also directly impacted by sulfation content and pattern [107]. Below we discuss some of the major biomedical roles played by DS.
3.2. Biomedical Properties
3.2.1. Coagulation and Thrombosis
Among the various biological properties attributed to DS, the main one is the ability to bind to and regulate heparin cofactor II (HCII), therefore catalyzing its inhibitory effect on thrombin (IIa) [15,113]. This activity promoted by DS has been intensely investigated in terms of its use as a new therapeutic agent in coagulation and thrombosis [106,114,115]. Studies in the early 2000’s reported on the potential anticoagulant/antithrombotic use of intimatan (dermatan 4,6-O-di-sulfated) [116,117,118,119,120,121]. The influence of disulfated disaccharide clusters on the HCII-mediated IIa inhibition by DS of different origins has been also evaluated [122]. Quantitation of disulfated disaccharide units, including active binding sites for in HCII interaction, in DS has been performed via NMR [123] Research has also been carried out on the potential anticoagulant properties of DS-derived oligosaccharides of well-defined chemical structures [124,125], and low-MW derivatives [126], as well as on molecules with unique structures derived from marine organisms [18,104].
About this latter case for instance, while the DS isolated from Ascidia nigra is fully sulfated at the C6 position of the GalNAc unit (100%) and mostly at the C2 position of the IdoA (80%) (Figure 5), the Styela plicata and Halocynthia pyriformis DSs are less sulfated at the C2 position of IdoA (65 and 70%, respectively) and widely sulfated at the C4 position of GalNAc (95 and 100%, respectively) [102,104] (Figure 5). This distinct pattern of sulfation directly impacts on the anticoagulant outcome of the ascidian DS [18,104]. While the DS from A. nigra has no significant anticoagulant activity, the DSs from S. plicata and H pyriformis show 4 and 5–6 times greater anticoagulant activities than mammalian DS comparatively in terms of their specific activities (units/mg) obtained by the activated partial thromboplastin time assay [104]. Regarding their potencies to catalyze the HCII inhibition of IIa, the DS from A. nigra shows negligible activity, while the DSs from S. plicata and H pyriformis have ∼10-fold and ∼6-fold higher activity than the native and oversulfated mammalian DS, respectively [104]. This is a clear example of sulfation pattern of GalAGs impacting biological activity. The difference in the sulfation sites of GalNAc units, 4-O- versus 6-O-sulfation, although very close in terms of content, is enough to lead to a complete different anticoagulant outcome.
Because DS is an effective anticoagulant and antithrombotic agent, this GalAG has been suggested as an emerging alternative for clinical use [72]. However, little was known about its hemorrhagic effect. It is important to highlight that heparin, a GAG of great clinical importance as an anticoagulant agent, presents risks of hemorrhage which varies from patient to patient [127,128,129,130]. In view of this fact, investigations into the hemorrhagic effect of DS were fundamental. Thus, Fernandez et al. compared the antithrombotic, hemorrhagic and anticoagulant effects of heparin and DS in rabbits. The results showed that DS is less hemorrhagic than heparin at equivalent antithrombotic doses [114]. Due to its reduced hemorrhagic potential, DS has been proposed as a safer alternative than heparin for use during hemodialysis in patients with renal impairment [131].
The main structural requirements of DS for its anti-IIa activity mediated by HCII, such as levels of sulfation, content of IdoA units and the minimal chain length for DS binding to HCII and formation of the DS:HCII:IIa ternary complex, were already uncovered by studies in the groups of Linhardt and Tollefsen [124,132,133,134]. Fractions of DS with higher content of IdoA are more active as antithrombotics [135], probably by enhanced flexibility of the IdoA monosaccharide residues. To elucidate the structural and conformational features involved in the formation of the active intermolecular complexes, mainly with HCII, underlying anticoagulant and antithrombotic mechanisms of action of DS [113], studies based on computational methods were recently performed to comprehend the advanced details behind the allosteric mechanisms of the DS-HCII interaction [17].
A screening of a virtual library gave rise to several topologies that were predicted to bind in the heparin-binding site of HCII at a ∼60° angle to helix D, a novel binding mode observed so far for the DS:HCII complex. This new binding geometry supports ternary DS-HCII-IIa complexation through a template mechanism [17]. This proposal stands in contrast to the previously accepted allosteric mechanism in which DS binds only to HCII, displacing the amino-terminal acidic domain of HCII which then interacts with the anion-binding exosite I of IIa and accelerates the formation of the HCII-IIa complex [136].
3.2.2. Wound Healing
DS is predominantly expressed in the skin and is released at high concentrations during wound repair to act as a key player during healing events [137,138]. A study by Penc et al. showed that DS released after injury is a potent promoter of fibroblast growth factor-2 (FGF-2) function [138]. In subsequent research, authors have shown that DS as modulator of the functions of FGF-7 and FGF-10 has the capacity to accelerate wound healing by stimulating proliferation and migration of keratinocytes [139,140]. In fact, DS is able to interact with several GBPs that can be involved in events related to wound repair, including but not limited to IIa [113], activated protein C [141], collagen [142], fibronectin [143], α2β1-integrin [144], defensins [145], interferon-γ [146] and transforming growth factor beta (TGF-β) [147]. All these interactions suggest possible mechanisms by which DS plays its role in healing. However, in addition to these distinct interactions, DS also participates in indirect interactions with multiple other plasma proteins related to tissue repair and wound healing [137].
3.2.3. Inflammation
Another potential medical application of DS relies on its anti-inflammatory effect. Belmiro et al. reported that subcutaneous administration of DS can inhibit colon inflammation in rats and promote reduction of macrophage recruitment, as well as T cell and macrophage activation [148]. In another study of the same group, the authors investigated the effect of DS extracted from the swine intestinal mucosa on inflammation and fibrosis in kidneys of mice after unilateral ureteral obstruction [149]. DS treatments were observed to reduce collagen, monocyte chemoattractive protein-1 (MCP-1) and TGF-β chemotactic protein content, as well as myofibroblasts and accumulation of macrophages in the kidney. The results indicated a protective effect of DS against the progression of renal inflammation and of fibrosis in kidneys [149].
The anti-inflammatory role of DS was also investigated by Gruber et al. [150]. In this study, they described the mucoprotective effect of systemic DS treatment in a preclinical model of oral mucositis (a potential complication of radiotherapy treatment of head and neck cancers). DS was in this model a potent agent capable of modulating radiation-induced oral mucositis. Further analysis was particularly made of protein expression associated with mucositis and the signaling changes involved in this complication. In all, epithelial morphology, proliferation and signaling were evaluated over two weeks of fractional irradiation with and without systemic administration of DS. The radioprotective effect of DS appears to be mainly based on an increase in epithelial junctions, thus strengthening epithelial integrity and reducing cellular loss. The authors pointed out that reduction of inflammation promoted by DS administration and local hypoxia may have contributed to mucoprotective activity [151].
3.2.4. Cancer and Metastasis
Because DS is a key player in several evolutionary stages of cancer (tumorigenesis, growth and spread) [26,152], exogenous DS was studied regarding its potential property as anticancer agent, particularly to inhibit metastatic processes [153]. In the work of Kozlowski et al. [153], the ascidia DSs from S. plicata (Figure 5A) and Phallusia nigra (with similar structure to A. nigra, Figure 5B) were investigated regarding their ability to bind to selectins, blocking therefore the functions of these co-receptors during attachment of cancer cells in events of tumor spread and invasion. The assays were performed in wild type mice and with P-selectin deficient animals. What the study has shown was that DS molecules extracted from both tunicate species attenuated the metastasis in wild-type mice; however, in the tests performed with P-selectin deficient mice they did not show a significant effect. This suggests that the specificity of P-selectin inhibition is the primary mechanism by which DS plays in its anti-metastatic activity [153]. The key roles of GAGs in cancer process as well as potential targets for drug development have been extensively reviewed by Afratis et al. [152]. This reference is a useful source of information on the molecular basis of cancer-related properties of DS.
3.2.5. Antiviral Activity
Besides the above-reported activities, the GalAG DS has also shown antiviral activities. In the work of Di Caro et al. [154], fractions of DS and CS were chemically oversulfated and tested regarding their properties against human immunodeficiency virus type 1 (HIV-1), herpes simplex virus type 1 (HSV-1) and human cytomegalovirus (HCMV). The assays performed revealed that while a 23.4 kDa oversulfated DS exhibits an EC50 (EC50 = concentration of drug that induces half of the maximum effect) of 0.04 μg/mLin HIV-1 inhibition, a second oversulfated DS of MW of 25 kDa showed activity against HSV-1 with EC50 of 0.01 μg/mL. The oversulfated CS of MW 17.3 kDa was the strongest anti-HCMV agent, showing its EC50 of 0.4 μg/mL [154].
4. FucCS
4.1. Overview
FucCS is a unique GalAG found in marine invertebrates such as sea cucumbers [6], crab [4] and octopus [61]. It plays a role as a structural component of the holothurian body wall and cartilage tissues of crab and octopus. In terms of structure, FucCS is composed of the same CS structure as vertebrates (Figure 1A, Figure 2 and Figure 6), although with additional branches of sulfated fucopyranose (Fucp) units (Figure 1B). FucCS is then chemically described as {→4)-[α-L-Fucp-(1→3)]-β-d-GlcA-(1→3)-β-d-GalNAc-(1→}n (Figure 6). The Fucp unit may bear O-sulfation at positions C2 and C4, according to the holothurian species (Figure 1B and Table 3). Although sulfation patterns of the CS backbones may vary among different FucCSs, the E type sulfation (4,6-di-sulfation) seems to dominate [6,155,156]. Other unusual types of branches can also occur in CS. These rarer branches may include fructose in the case of the k-4 uropathogenic Escherichia coli [157], glucose in squid CS [158], a KS disaccharide [159], and the sulfated carbohydrate-protein linkage region in inter-α-trypsin inhibitor in human plasma, bikunin [160,161].
Structure determination of many holothurian FucCS via NMR spectroscopy has shown the presence of 4,6-di-sulfated, 4-mono-sulftated, 6-mono-sulfated and non-sulfated GalNAc units in the CS backbone [162]. In addition, a minor degree of 3-O-sulfation of GlcA can occur [163]. These proportions may vary with the species of holothurians [6,164]. Hence, structural heterogeneity of CS backbones can be observed among FucCS molecules extracted from the same species or among different species. The variation of sulfation patterns on the branching Fucp units seems to be more relevant in terms of biological activity than the variation of sulfation on the main backbone. This is because the presence of the branching units on FucCS molecules provides this invertebrate molecule with the capacity to modulate numerous physiological systems. The levels of activity in these systems are not exclusively dependent on fucosylation but also on sulfation patterns of the branching units (Table 3). Below, we discuss in detail the potential biomedical functions exerted by FucCS in multiple pathophysiological systems. The structural motifs responsible for the best activity are also discussed when known. These biologically active structural motifs are very frequently the sulfation patterns of the branching Fucp unit.
4.2. Biomedical Properties
4.2.1. Coagulation and Thrombosis
CS is not active as an anticoagulant, but fucosylation renders it active depending on the pattern of sulfation on the fucosyl branch (Table 3) [164]. Among the multiple potential biomedical applications of FucCSs, their potential anticoagulant properties are the most desirable and best studied. Previously, the anticoagulant/antithrombotic mechanism of action of FucCS had been believed to be centered primarily on being a catalyst for the inhibitory functions of the serpins antithrombin (AT) and HCII, and for the coagulation serine-proteases, IIa and Xa [19]. The ability of FucCS to potentiate IIa inhibition is mainly characterized by the activity of HCII [185], and the sulfation degrees have been shown to play a crucial role in this serpin-dependent activity [186]. More recently, however, a serpin-independent anticoagulant/antithrombotic activity has been assigned to holothurian FucCS [187,188,189,190,191], and the mechanism of action behind this unique property has been reported to rely on the capacity of the FucCS to impair factor Xa generation by inhibiting the formation of the intrinsic tenase complex, which is in turn made by factors VIIIa, IXa and X [156,187,191,192]. This serpin-independent anticoagulant activity of FucCS has been reported to dominate the process, and the serpin-dependent activity seems to be just residual in the systemic point-of-view [191].
Since non-fucosylated CS is inactive in anticoagulation, the removal of the Fucp branching unit from the FucCS is expectedly detrimental to anticoagulant and antithrombotic activities [19,21]. In addition to the presence of the sulfated fucosyl branch, the sulfation pattern of this branch was also reported to be a contributing factor [6,164,186]. A Fucp unit with the 2,4-disulfation pattern was reported to be the anticoagulant/antithrombotic motif [193]. The concentration of 2,4-O-disulfated Fucp units in the FucCSs of the four holothurian species Isostichopus badionotus, Thelenota ananas, Stichopus japonicus, and Apostichopus japonicus are higher when compared to the molecules of the other species (Table 3). Although this concept of the 2,4-di-sulfation is highly supported based on several studies [165,166,186,193], a new proposition defends the conception that different structures (different sulfation patterns) of the branching α-L-Fucp units do not have an impact on the anticoagulant outcome [173]. Procoagulant effects from low doses of the holothurian FucCS, as measured by thrombin generation, have also been reported [194], adding to the range of contradictions. Dual and antagonic biomedical effects of sulfated glycans, including those of marine source, are not unexpected [195].
4.2.2. Hemodialysis
Minamiguchi et al. explored the anticoagulant property of the FucCS from S. japonicus (MW around 14 kDa) in hemodialysis [34,196]. As opposed to unfractionated heparin (UFH) and low molecular weight heparin (LMWH), which showed significant bleeding time, nafamostatmesilate (FUT) and the FucCS have just scarcely prolonged bleeding time. As opposed to FUT, which was able to increase only the activated partial thromboplastin time (aPTT), the FucCS was able to increase both aPTT and thrombin clotting time. This indicates that the studied FucCS can be used as an anticoagulant agent in procedures of hemodialysis accompanied by low risks of hemorrhage [34]. FucCS has also shown an AT-independent anticoagulant mechanism of action in the hemodialysis experiment [196]. This mechanism helps to explain the low hemorrhagic risks of the low MW FucCS tested in the in vivo experimental models of hemodialysis as compared to the other standards, including UFH and LMWH [196].
4.2.3. Atherosclerosis
FucCS from sea cucumber species Ludwigothurea grisea has shown great affinity for lipoproteins, especially apo B-containing lipoproteins like low-density lipoprotein (LDL) and very low-density lipoprotein [197]. The binding affinity for LDL increases proportionally to MW of L. grisea FucCS and is lost upon desulfation and defucosylation. Since LDL is largely responsible for the development of atherosclerosis or any other disease related with the obstructions of the coronaries, and because FucCS has great affinity to LDL, application of FucCS seems promising in capture and decrease of LDL plasma content [197]. The reasonable lifetime of FucCS in the plasma when orally administered supports the medical use of holothurian FucCS through the oral route [198]. In another study, FucCS from S. japonicus has also indicated the potential medical benefits of this invertebrate GalAG in atherosclerosis [199]. The authors postulated that FucCS has the capacity to inhibit neointimal thickening induced by balloon catheterization. The mechanism of action proposed for FucCS in the event of atherosclerosis relies on its capacity to inactivate growth of aberrant smooth muscle cells (SMC) in the system.
Sulfation patterns of fucose branches were proved to affect the anti-hyperlipidemic activity of FucCS [200]. The anti-hyperlipidemic effects were compared using a high-fat high-fructose diet-fed C57BL/6J mice model. Both FucCS from I. badionotus and Pearsonothuria graeffei showed significant effects on lipid profile improvement, liver protection, blood glucose diminution and hepatic glycogen synthesis. Specifically, FucCS from P. graffei with 3,4-O-disulfation fucose branches (Table 3) was more effective in reduction of blood cholesterol, LDL and atherogenic index. These results clearly indicate that FucCS from P. graffei could be used as a potential anti-hyperlipidemic agent.
4.2.4. Cellular Growth
The effects of FucCS from L. grisea on vascular SMC from rat thoracic aorta and on human umbilical vein endothelial cell (HUVEC) in culture, in the presence or absence of FGF-1 and FGF-2, were examined [35]. While FucCS showed an efficient inhibitory activity on SMC proliferation, no effects have been observed on HUVEC proliferation and migration assays without FGF exogenously added to the culture media. The presence of sulfated fucose branches on FucCS was indicated to be an essential determinant to the activity of the holothurian GalAG on cell proliferation and migration; upon defucosylation, the effects of FucCS on cells are abolished [35].
4.2.5. Angiogenesis
The background concerning the effects of FucCS on cellular growth was re-examined by Tapon-Bretaudière et al. with respect to the effects of the holothurian GalAG in angiogenesis [32]. The effects of the FucCS from L. grisea were investigated on endothelial cells through an in vitro angiogenesis-related experimental model utilizing Matrigel. In the presence of FGF-2, FucCS showed the capacity to induce formation of vascular tubes by endothelial cells in Matrigel. The vascular tubes were observed distributed as very organized capillary-like networks within typical closed structures. Heparin did not promote vascular tube formation in this experimental model of angiogenesis indicating therefore that the structural features of the sulfated glycans play a role in the outcome. As in most of the other biomedically relevant actions, desulfation and defucosylation abolish the pro-angiogenic effects of FucCS [32].
4.2.6. Fibrosis
Melo-Filho et al. have studied the properties of FucCS from L. grisea in respect to its capacity of retarding or preventing the pathological formation of renal fibrosis. The method utilized for this was an in vivo model of rats submitted to unilateral ureteral obstruction (UUO) [36]. Compared with control animals (FucCS-untreated rats), collagen deposition decreased in animals treated with FucCS. The cell amounts of myofibroblasts and macrophages were also reduced. Another beneficial factor of FucCS was its capacity to diminish the production of transforming growth factor (TGF)-β. As observed in P-selectin-deficient mice submitted to UUO, renal fibrosis was observed to be greatly attenuated. Interestingly, these deficient mice showed also little response to the L. grisea FucCS. As conclusion, FucCS is capable of attenuating renal fibrosis on UUO-submitted animals, and this effect seems to occur through a P-selectin-related mechanism of action [36].
4.2.7. Cancer and Inflammation
Like heparin, which has the capacity to inhibit P- and L-selectin binding to their ligand, sialylLewisX, resulting consequently in antitumor and anti-inflammatory outcomes, the FucCS from L. grisea was analyzed regarding its antiselectin property, also in the context of presenting potential antitumor and anti-inflammatory activity [24]. Indeed, like heparin, FucCS was observed to exert a potent inhibition effect of both P- and L-selectin bindings to immobilized sialylLewisX and attachment of carcinoma cell (LS180) to immobilized P- and L-selectins. The FucCS inhibition process occurs in a dose-dependent manner. FucCS showed also the capacity of inhibiting adenocarcinoma MC-38 cells to colonize lungs in an experimental model of metastasis in mice [24]. In addition to this activity, the authors also demonstrated through two different models of inflammation (thioglycollate-induced peritonitis and lipopolysaccharide-induced lung inflammation) that L. grisea FucCS showed the capacity to inhibit neutrophil recruitment. Both inhibitory properties are dose-dependent, and the effective doses gave no alterations in the plasma aPTT. This indicates that FucCS showed its antitumoral or anti-inflammatory response without presenting hemorrhagic risks [24]. Panagos et al. also investigated this antiselectin capacity of FucCS from Holothuria forskali by screening FucCS-derived oligosaccharides via microarray techniques [182]. In this study, the authors have proposed the structural conformation of the FucCS building blocks and the relevance of sulfation patterns of the Fucp units in modulating the overall 3D structure of this GalAG in solution.
Zhao et al. observed by both in vivo and in vitro assays that FucCS can concomitantly reduce levels of both metastasis using B16F10 cells of mouse melanoma, and tendency to thromboembolism when FucCS is administered. It has been shown that under the holothurian GalAG administration, down-regulation of both transcription and protein expression of tissue factor can be achieved, and fibrin formation is inhibited primarily by attenuating the generation of activated factor Xa. In addition to these activities, FucCS also has the capacity to suppress activation of p38MAPK and ERK1/2 signaling pathways. These signaling pathways are considered the central routes for the expression of metastasis-related matrix metalloproteinases [201]. These findings suggest that a single GalAG molecule can exert dual medical properties, as both anticancer and antithrombotic agent. In a separate publication by He et al., the investigators found that the FucCS from I. badionotus could markedly inhibited tumor growth by down-regulating expressions of Hif-1α (hypoxia-inducible factor 1-alpha), heparanase, and vascular endothelial growth factor in vivo [202].
4.2.8. Microbial Infections
Viral Infections
The anti-HIV activity of the FucCS from T. ananas has been studied [29,30]. Data suggested that this FucCS can effectively and selectively block both X4- and R5 × 4-tropic HIV-1 infection of C8166 cells, as well as peripheral blood mononuclear cells with little cytotoxicity against these cells. Results indicated that this FucCS can bind to the HIV membrane glycoprotein 120 (gp120) at nanomolar binding affinity via molecular interaction with the CD4i epitope. As expected, based on all actions of FucCS examined so far, the sulfated fucosyl branches were indicated to be structural motifs responsible for anti-HIV-1 activity. In addition, molecular size was also observed to participate given the fact that several FucCS-derived low MW fragments have exerted higher anti-HIV-1 and lower anticoagulant activities than heparin and the native FucCS used as standards [29]. The primary mechanism of action underlying the anti-HIV outcome of FucCS was reported to be on blocking the virus entry into the host cells. This mechanism of action is in good accordance with the general antiviral mechanism of action proposed for sulfated polysaccharides [203].
Malaria-Related Protozoan Infection
FucCS also modulates infections caused by the protozoan parasite Plasmodium falciparum [33]. Sequestration of P. falciparum-infected erythrocytes (Pf-iEs) in the microvasculature of vital organs is an essential event during P. falciparum-caused pathogenesis, such as cerebral malaria and malaria in pregnancy. These events are driven by cytoadhesion of Pf-iEs onto GAG receptors displayed at the surface of host endothelial cells, of non-infected erythrocytes, and on placenta trophoblasts. Based on the general background concerning the antimicrobial activity of sulfated glycans [203], exogenous GAGs can be used to interfere with the host GAG-Pf-iEs interactions as a therapeutic strategy against the malaria complications. On this basis, the use of FucCS isolated from L. grisea has shown the ability to inhibit Pf-iEscytoadhesion [33]. The holothurian GalAG is also capable of blocking parasite development by interfering with the merozoite invasion of the P. falciparum. The inhibition of protozoan infection is abrogated when the Fucp branches are removed [33].
4.2.9. Hyperglycemia Diabetes-Related Processes
Diabetic Nephropathy
GAG-based drugs such as LMWH can be renoprotective in diabetic nephropathy. In this regard, FucCS from L. grisea has been tested for its clinical potential to prevent or diminish the pathological consequences related to diabetic nephropathy [38]. Diabetes mellitus was experimentally induced by streptozotocin in male Wistar rats to perform the tests. FucCS and LMWH were both administered subcutaneously. Blood glucose, blood pressure, albuminuria, and renal function were monitored after 12-day treatment. Kidneys were evaluated for mesangial expansion and collagen content. Immunohistochemical quantifications of macrophages, TGF-β, nestin and immunofluorescence analysis of heparanase-1 and glomerular basement membrane, heparan sulfate content were also examined. Gene expression of proteoglycan core proteins and enzymes involved in GAG assembly/degradation were analyzed by TaqMan real-time PCR [38]. Treatment with GAGs prevented albuminuria but did not change the glucose levels or other functional aspects. The negative group exhibited increased mesangial matrix deposition and tubulointerstitial expansion, but these aspects were prevented by administration of both GAG types. TGF-β expression and macrophage infiltration were also prevented by both GAGs, and podocyte damage was halted. The diabetic milieu resulted in the downregulation of agrin, perlecan and collagen XVIII mRNAs, along with the expression of enzymes involved in GAG biosynthesis. It was observed that FucCS and LMWH treatments could positively modulate such changes. Heparanase-1 expression was significantly reduced after GAG treatment without affecting the globular basement membrane and heparan sulfate content. These findings suggest that FucCS could act as a protective agent against the aggravating consequences of nephropathy during diabetes at similar levels of response seen for LMWH [38].
Diabetes-Related Apoptosis of Islets of Langerhans
Apoptosis of pancreatic islets of Langerhans is a common characteristic of diabetes. High-fat high-sucrose diet-induced insulin-resistant mice were studied for 19 weeks with and without FucCS treatment [204]. It was shown that FucCS treatment significantly decreased fasting blood glucose, insulin, and TNF-α levels. FucCS treatment also increased the serum levels of adiponectin, improved insulin resistance, and repaired HFSD-injured islets of Langerhans. FucCS supplementation showed the ability to inhibit pancreas cell apoptosis via reduction of cytochrome c, caspase 9, and caspase 3 mRNA expressions, cytochrome c release into the cytoplasm, and caspase 9 and cleaved caspase 3 protein expressions. These effects were associated with up-regulation of Bcl-2 and Bcl-xL mRNA and protein expressions, and down-regulation of Bax and t-Bid mRNA and protein expressions. Treatments achieved by FucCS combined with rosiglitazone showed a synergistic protection of pancreatic islets. Overall, these data show that improvement in insulin resistance and protection of islets of Langerhans from apoptosis can be achieved by FucCS via inactivation of the intrinsic mitochondrial pathway [204].
4.2.10. Tissue Damage
Monteiro-Machado et al. showed that FucCS from L. grisea can present potential therapeutic properties against myotoxicity induced by snake venom from Bothrops jararacussu [205]. Myotoxicity was induced in muscle tissue in vitro by crude venom, by bothropstoxin-I alone or by bothropstoxin-I and bothropstoxin-II combined. The myotoxicity was abrogated in all cases in the presence of FucCS. It has also shown that FucCS can inhibit edematogenic activity and partially prevent the reduction of total leukocytes in blood when pre-incubated with crude venom. Taking together all data presented by reference [205], it has been shown that FucCS is a reliable inhibitor of myotoxicity and inflammation induced by B. jararacussu venom or the related toxins.
5. Drawbacks
5.1. The Dilemma Regarding Effectiveness and Safety of CS as a Dietary Supplement for Treatments of Osteoarthritis
CS has been recommended for use in symptomatic relief and possible treatment for osteoarthritis, often in combination with glucosamine, since the mid 1900s [206]. It has been hypothesized that increasing concentrations of CS will allow for regeneration of the cartilaginous tissues. However, a defined mechanism of action has not been described [206]. Because of this lack of clear efficacy, CS has been under scrutiny for whether it is in fact beneficial in treatment of osteoarthritis. Guidelines were set by the American College of Rheumatology for knee osteoarthritis [207] and by the Osteoarthritis Research Society International [208]. Neither entity explicitly condones the use of CS. Much of the experimental data collected in vitro that identifies bioactivity of CS relevant to efficacy in treatment of osteoarthritis are based on CS concentrations far higher than are achievable in the circulation [206]. Understanding the structural requirements for bioactivity might lead eventually to structurally better-defined and less heterogenous CS-based medicinal natural products.
In addition to some of these issues, there are also some considerations to make concerning patients who are taking CS for relief of osteoarthritis, especially in comparison to those who are administered more well-defined medicines like celecoxib. In comparison with placebo, for example, patients with osteoarthritis taking CS reported a reduction in pain [207,208]. However, other surveys of patients have found the opposite, showing no advantage over placebo [209]. This indicates that CS may be either effective or ineffective in the relief of pain and osteoarthritis. This variation in effectiveness may be a result of patient to patient variation, and/or to the different levels of pain and osteoarthritis. What is certainly clear is that more in-depth studies must be conducted to determine the safety and extent to which the effects are clinically achieved and observed and, at the end, statistically registered.
Another point to be clarified is the efficiency of glucosamine in combined administration with CS. Although evidence for the symptomatic efficacy of glucosamine added to CS preparations for treatments of knee osteoarthritis have been seen, for example, in the study of Zeng et al. [207], pain relief following ingestion of CS has been reported to be independent of simultaneous or prior intake of glucosamine, as stated in the work of Jackson et al. [210]. In this study, the authors stated that although combined dosing of glucosamine with CS was found to reduce the plasma levels of glucosamine, any improved pain relief by combined administration of the two components cannot be explained by higher circulating concentrations of glucosamine.
5.2. The Presence of Unexpected Components in CS-Based Pharmaceutical Formulations for Oral Administration
Of great concern with the application of CS are potential issues associated with the presence of non-desirable structures or components during the manufacturing processes of CS as this GalAG is extracted from a wide range of sources [211]. Because CS is expressed and isolated as long chains, with high structural diversity with respect to sulfation patterns containing just a single chain, it is important to highlight the off-target effects that may be associated with the structural heterogeneity. Further unexpected effects may arise from the presence of another GAG type present in the preparations, such as KS [212]. KS is a structurally complex GAG that can potentially give rise to unexpected effects when administered accidentally during common usage of CS [213] although any deleterious effects have not thus far been demonstrated [65].
In an industrial setting, it is not uncommon for these impurities to be found in the final product due to similarities within both isolation and purification procedures of CS and KS [211]. KS contamination can therefore be expected in CS formulations for multiple usages, ranging from laboratory reagents to its use as an alternative medicine [212]. In some cases, this contamination of KS may be remarkably high, with the total amount being up to 16% in the final product [65]. This may result both in possible negative attributes within the formulation and label claims, as well as possible dishonest representation of the product on the market [65]. This contamination, however, is not limited to KS, as other GAG types, protein and nucleic acids may also be found in some of the CS formulations [211]. In addition to the presence of non-desirable components, there is also the possibility of occurrence of disulfated residues in final formulations [214]. This is particularly likely to occur when derivatives from marine sources are used. This whole issue clearly highlights the importance of source material versus the final product composition that reaches the consumer [214]. This issue becomes especially important in the cases of nutraceutical preparations of CS that are not subject to the same quality standards as pharmaceutical formulations of drugs, which must follow rigorous processes of quality assurance and control. In some cases, the stated purity of CS is not even found or offered [54,214].
Nutraceutical formulations of CS could also be subject to intentional adulteration to reduce the initial cost involved in the production of the active ingredient. This practice is thought to have played a major role in the 2008 “Heparin crisis” and resulted in potent adverse effects to the end consumers [211]. The discrepancy between pharmaceutical and nutraceutical preparations can also go as far as a noticeable difference in expected therapeutic effects. The efficacy of pharmaceutical-grade CS preparations should not be extrapolated to nutraceutical formulations [215] for which controls are usually less stringent, depending on the local jurisdiction.
5.3. Potential Antigenic and Autoimmune Side-Effects of CS
The structural heterogeneity of GAGs and likely GalAGs gives a diversity of sequences for interaction with GBPs, and consequent regulation of multiple GBPs. This consequently enlarges the spectrum of biological actions of these sulfated glycans and makes them important structural players in perhaps all physiological systems of the body. Conversely, this same structural heterogeneity can give rise to issues related to possible antigenic effects. Binding epitopes that have been associated with serious autoimmune disorders may be involved in interaction with GAGs (and likely GalAGs). One of interest is the human natural killer-1 (HNK-1) epitope, composed of the 3-sulfated GlcA [216], which is characteristic of CS-K, CS-L and CS-M (Table 1). The HNK-1 epitope is found within the myelin sheaths and is linked to autoimmune disease in the peripheral nervous system. It is recognized by its conjugate monoclonal antibody anti-HNK-1 [217]. Interesting, the rare 3-sulfated GlcA unit in intact CS chains is also recognized by anti-HNK-1 [218]. Hence, this raises the question of potential triggering of autoimmune complications by CS-K, CS-L, CS-M and some FucCS.
Regarding this same issue regarding the possible antigenicity of CS, it is well-known that platelet factor 4 (PF4)/heparin complexes can trigger the immunological response that will cause heparin-induced thrombocytopenia (HIT) [219,220]. In this event, the intermolecular complexes aggregate and will trigger the production of antibodies associated with the disease state. Furthermore, PF4 has shown binding properties to CS [221,222] and its oversulfated version [223], therefore leading to high-level production of HIT-related antibodies in patients undergoing CS-based treatment [224]. For these reasons, these GalAGs may also be capable of inducing thrombocytopenia in a similar manner as heparin does in HIT. This side-effect will, of course, be dependent on the concentration of the epitope. Thus, sulfation patterns of CS, chain heterogeneity and concentration are all factors that play a role in the antigenic events of CSs.
5.4. Highly Sulfated CS can Activate the Kinin-Kallikrein System
As in the “Heparin crisis” of 2008 [64,225,226,227,228,229], highly sulfated CS from either from natural sources like CS-D, CS-D, CS-K, CS-L, and CS-M (Table 1) or from chemical reactions [230] are likely to be capable of triggering the same side effects as oversulfated chondroitin sulfate (OSCS) in the activation of factor XII and kinin-kallikrein system, which will result in bradykinin formation and severe hypotension. The ability of highly sulfated CS to activate the kinin-kallikrein system is likely to depend on both the extent and the pattern of sulfation [230].
5.5. Potential Problems for the Medical Use of DS and other GalAGs
The potential downsides of the medical use of DS are like those of CS; it is appropriate to discuss here some aspects common to all the GalAGs. The use of tissues from cows and pigs as main sources of raw materials for DS and other GalAGs can, in principle, lead to the potential contamination of potentially harmful and undesirable molecules if the original source is not processed well. This is not a current risk, but problems could arise mainly from suboptimal techniques in the purification processes of GalAGs as a whole, as these glycans are usually extracted as byproducts from the large-scale agricultural exploration of farmed animals. There are three ways to specifically overcome this issue. First would be to ensure that the manufacture of medical grade GalAGs meets the same exacting standards as current production methods for heparin [231]. The second alternative could be the use of other sources of DS, which may not contain, or contain in much lesser amounts, the contaminants of the mammalian tissues. These new sources are the marine organisms [107]. The third and seemingly last alternative would be the use of chemically synthesized DS from the laboratory [107,232].
The second concern in medically exploring DS might be the undesirable side-effects of this molecule when exogenously administered. It is known that DS plays a role in multiple physiological systems, and the administration of DS in pharmacologically active doses for a certain type of health complication could potentially also lead undesirably to detrimental impairments of other key functions of DS in other systems. Comparisons with the widespread use of the multivalent GAG heparin as an antithrombotic agent are reassuring but should not lead to complacency. A rational way to overcome this problem would be the recognition of specific sulfation patterns that are selectively important for only one or a few complications and the use of exogenous DS with a particular sulfation pattern, so that a selective drug could be developed, and the specific type of treatment could be reached.
The third concern about the medical use of exogenous DS would be the potentially dangerous harmful effects of certain DS derivatives, especially the oversulfated versions tested for antiviral activity [154]. As documented above, oversulfated DS is likely to trigger the same side effects of highly sulfated CSs and OSCS regarding the activation of factor XII and kini-kallikrein system-dependent hypotension. As opposed to CS in which experiments were performed to prove the side-effects of highly sulfated CS [230], no tests have been performed to prove such a hypothesis for highly sulfated DS. If it is confirmed that these undesirable actions can be induced by oversulfated DS, these GalAG derivatives are very unlikely to be employed as future drugs for any of their biomedical functions because they will not fulfill the minimal requirements for safety.
The fourth concern regarding the medical application of some DS structures is related to the autoimmune response related to PF4, which may lead to thrombocytopenia or HIT [233]. Because naturally or artificially oversulfated DS structure resembles the one of OSCS, DS can also trigger HIT, although the regular DS (CS-B) structure, as used as active ingredient of danaparoid, is used in treatments of patients prone to HIT [234,235]. As for CS, this side effect might be dependent on concentration and structural heterogeneity. On the other hand, DS will probably not be as antigenic as CS in the case of HNK-1 because, as opposed to the latter, which can contain the antigen 3-sulfated GlcA epitope, DS does not present this motif (Table 2). Nonetheless, while not confirming the hypotensive side-effect, there are studies that indicate that oversulfated DS can cause kallikrein activation [236,237].
5.6. Potential Harmful Effects of FucCS
FucCS is the GalAG that exerts the highest number of biomedical functions. These activities include anticoagulation and antithrombosis; beneficial effects in hemodialysis, atherosclerosis, cellular growth, and angiogenesis; in fights against fibrosis, tumor growth, inflammation, viral and protozoan infections; beneficial effects in hyperglycemia, in diabetes-related pathological events and in snake venom-caused tissue damage. FucCS would comprise a potent and promising medical agent to be explored in the future. However, obstacles such as potentially harmful side-effects and unclear routes of synthesis make its real implementation as a drug candidate difficult, therefore hindering its use in studies of clinical trials and the future application of this marine GalAG in medicine.
Although FucCS shows good effectiveness as opposed to CS’s use in osteoarthritis, and relative safety since it is extracted from a non-harmful sea organism, the greatest obstacle of all would be the similar harmful effects reported for OSCS. In this case, the harmful effect resides in the capacity of highly sulfated glycans such as FucCS to activate coagulation blood factor XII [238,239] and consequently induce kinin-related severe hypotension caused by kallikrein, a potent regulator of blood pressure via activation of bradykinin, besides inducing platelet aggregation [186] and spontaneous bleeding in humans [188]. Unfortunately, the real use of FucCS in the pharmaceutical industries is still some years away. Nevertheless, an enormous amount of recent research concerns the potential medicinal properties of this molecule [6,164,240].
6. Concluding Remarks
Medical interest in GalAGs has expanded worldwide, especially with the boom in projects related to medical glycomics. The principal types of GalAGs are CS, DS and FucCS. Sub-types also exist and are primarily characterized by different patterns of sulfation. Screenings have shown positive effects of GalAGs in multiple biomedical areas, including but not limited to thrombosis, cancer, inflammation, neurological disorders and microbial infections. However, the applications of GalAGs in medicine have been questioned. Doubts center primarily on their real effectiveness and potential harmful effects. There are contradictory opinions regarding the benefits of CS-based pharmaceutical formulations used for arthritis as well as the safety conditions in manufacturing processes of GalAGs, as GAGs, during the steps of isolation and purification for the global market. Impurities such as the KS content have also been reported in CS-based pharmaceutical preparations. More important are the harmful side effects of FucCS, including OSCS in activation of factor XII and kinin-kallikrein system-dependent hypotension. Despite the great number of potential biomedical applications and the diversity of chemical structures, the downsides of GalAGs considerably restrict the other medical exploitations of GalAGs, in addition to the use of CS as dietary supplements destined for aiding complications related to joint pain/inflammation and osteoarthritis. As clear synthetic routes of biomedically active GalAGs’ oligosaccharide sequences are not yet fully practical, and the mechanisms of action concerning their intermolecular complexes with GBPs are not completely understood, the current and future medical applications of GalAGs will continue to be limited. Protocols employed during large-scale manufacture must be also optimized. We believe all these areas related to the science and medical exploration of GalAGs can significantly develop in the next few years, especially accompanied by the incentive and parallel progress of medical glycomics worldwide.
Author Contributions
W.P.V. wrote the first draft of the section about CS, made the figures and improved the first draft of the graphical abstract. A.A.V. wrote the first draft of the section about DS and worked on references and citation. A.V.G. wrote the first draft of the section about FucCS and worked on the references. V.H.P. integrated, completed, and edited the entire manuscript, and drafted and edited the graphical abstract. B.M. edited and placed comments/additions on the draft. V.H.P. and B.M. envisioned the article. All authors searched the literature.
Funding
This research was funded by the University of Mississippi, and American Association of Colleges of Pharmacy (AACP). The APC (10%) was funded by Imperial College London.
Acknowledgments
The authors are grateful to the guest editors of the Special Issue “Emerging Frontiers in Glycosaminoglycans and Mimetics” Giangiacomo Torri, Jawed Fareed and Job Harenberg for the kind invitation to contribute a paper from our authorship to their Special Issue.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pomin, V.H.; Mulloy, B. Glycosaminoglycans and Proteoglycans. Pharmaceuticals 2018, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Silbert, J.E.; Sugumaran, G. Biosynthesis of Chondroitin/Dermatan Sulfate. IUBMB Life 2002, 54, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, K.; Mikami, T.; Uyama, T.; Mizuguchi, S.; Nomura, K.; Kitagawa, H. Recent advances in the structural biology of chondroitin sulfate and dermatan sulfate. Curr. Opin. Struct. Biol. 2003, 13, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, H.; Tanaka, Y.; Yamada, S.; Seno, N.; Haslam, S.M.; Morris, H.R.; Dell, A.; Sugahara, K. A Novel Pentasaccharide Sequence GlcA(3-sulfate)(β1-3)GalNAc(4-sulfate)(β1-4)(FucR1-3)GlcA(β1-3)GalNAc(4-sulfate) in the Oligosaccharides Isolated from King Crab Cartilage Chondroitin Sulfate K and Its Differential Susceptibility to Chondroitinases and Hyaluronidase. Biochemistry 1997, 2960, 3998–4008. [Google Scholar] [CrossRef]
- Vieira, R.P.; Mourão, P.A. Occurrence of a unique fucose-branched chondroitin sulfate in the body wall of a sea cucumber. J. Biol. Chem. 1988, 263, 18176–18183. [Google Scholar]
- Pomin, V.H. Holothurian Fucosylated Chondroitin Sulfate. Mar. Drugs 2014, 12, 232–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomin, V.H.; Sharp, J.S.; Li, X.; Wang, L.; Prestegard, J.H. Characterization of Glycosaminoglycans by 15 N NMR Spectroscopy and in Vivo Isotopic Labeling. Anal Chem. 2010, 82, 4078–4088. [Google Scholar] [CrossRef]
- Sirko, S.; Von Holst, A.; Wizenmann, A.; Götz, M.; Faissner, A. Chondroitin sulfate glycosaminoglycans control proliferation, radial glia cell differentiation and neurogenesis in neural stem/progenitor cells. Development 2007, 2738, 2727–2738. [Google Scholar] [CrossRef]
- Gu, W.; Fu, S.; Wang, Y.; Li, Y.; Lü, H.; Xu, M.; Lu, P. Chondroitin sulfate proteoglycans regulate the growth, differentiation and migration of multipotent neural precursor cells through the integrin signaling pathway. BMC Neurosci. 2009, 15. [Google Scholar] [CrossRef]
- Tanaka, M.; Maeda, N.; Noda, M.; Marunouchi, T. A Chondroitin Sulfate Proteoglycan PTP ζ/RPTPβ Regulates the Morphogenesis of Purkinje Cell Dendrites in the Developing Cerebellum. J. Neurosci. 2003, 23, 2804–2814. [Google Scholar] [CrossRef]
- Shannon, J.M.; Mccormick-Shannon, K.; Burhans, M.S.; Shangguan, X.; Srivastava, K.; Hyatt, B.A. Chondroitin sulfate proteoglycans are required for lung growth and morphogenesis in vitro. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 285, L1323–L1336. [Google Scholar] [CrossRef] [Green Version]
- Lane, M.C.; Solursh, M. Primary mesenchyme cell migration requires a chondroitin sulfate/dermatan sulfate proteoglycan. Dev. Biol. 1991, 143, 389–397. [Google Scholar] [CrossRef]
- Villena, J.; Brandan, E. Dermatan Sulfate Exerts an Enhanced Growth Factor Response on Skeletal Muscle Satellite Cell Proliferation and Migration. J. Cell. Physiol. 2004, 178, 169–178. [Google Scholar] [CrossRef]
- Higgins, M.K. The Structure of a Chondroitin Sulfate-binding Domain Important in Placental Malaria. J. Biol. Chem. 2008, 283, 21842–21846. [Google Scholar] [CrossRef] [Green Version]
- Tollefsen, D.M. Activation of heparin cofactor II by heparin and dermatan sulfate. Nouv. Rev. Fr. Hematol. 1984, 26, 233–237. [Google Scholar] [CrossRef]
- Tollefsen, D.M. Vascular Dermatan Sulfate and Heparin Cofactor II. In Progress in Molecular Biology and Translational Science; Academic Press: Cambridge, MA, USA, 2010; pp. 351–372. [Google Scholar]
- Raghuraman, A.; Mosier, P.D.; Desai, U.R. Understanding Dermatan Sulfate-Heparin Cofactor II Interaction through Virtual Library Screening. ACS Med. Chem. Lett. 2010, 1, 281–285. [Google Scholar] [CrossRef]
- Vicente, C.P.; Zancan, P.; Peixoto, L.L.; Alves-Sá, R.; Araújo, F.S.; Mourão, P.A.S.; Pavão, M.S.G. Unbalanced effects of dermatan sulfates with different sulfation patterns on coagulation, thrombosis and bleeding. Thromb. Haemost. 2001, 86, 1215–1220. [Google Scholar] [CrossRef]
- Mourão, P.A.S.; Pavão, M.S.G.; Mulloy, B.; Tollefsen, D.M.; Mowinckel, M.; Abildgaard, U. Structure and Anticoagulant Activity of a Fucosylated Chondroitin Sulfate from Echinoderm. Sulfated fucose branches on the polysaccharide account for its high anticoagulant action. J. Biol. Chem. 1996, 271, 23973–23984. [Google Scholar] [CrossRef]
- Prandoni, P.; Meduri, F.; Cuppini, S.; Toniato, A.; Zangrandi, F. Dermatan sulphate: A safe approach to prevention of postoperative deep vein thrombosis. Br. J. Surg. 1992, 79, 505–509. [Google Scholar] [CrossRef]
- Mourão, P.A.S.; Guimarães, M.A.M.; Mulloy, B.; Thomas, S.; Gray, E. Antithrombotic activity of a fucosylated chondroitin sulphate from echinoderm: Sulphated fucose branches on the polysaccharide account for its antithrombotic action. Br. J. Haematol. 1998, 101, 647–652. [Google Scholar] [CrossRef]
- Iovu, M.M.D.; Dumais, G.B.S.; Souich, P.M.D. Anti-inflammatory activity of chondroitin sulfate. Osteoarthr. Cartil. 2008, 16, 14–18. [Google Scholar] [CrossRef]
- Volpi, N. Anti-inflammatory activity of chondroitin sulphate: New functions from an old natural macromolecule. Inflammopharmacology 2011, 19, 299–306. [Google Scholar] [CrossRef]
- Borsig, L.; Wang, L.; Cavalcante, M.C.M.; Cardilo-Reis, L.; Ferreira, P.L.; Mourão, P.A.S.; Esko, J.D.; Pavão, M.S.G. Selectin Blocking Activity of a Fucosylated Chondroitin Sulfate Glycosaminoglycan from Sea Cucumber. J. Biol. Chem. 2007, 282, 14984–14991. [Google Scholar] [CrossRef] [Green Version]
- Asimakopoulou, A.P.; Theocharis, A.D.; Tzanakakis, G.N.; Karamanos, N.K. The Biological Role of Chondroitin Sulfate in Cancer and Chondroitin-based Anticancer Agents. In Vivo 2008, 390, 385–389. [Google Scholar]
- Thelin, M.A.; Svensson, K.J.; Shi, X.; Bagher, M.; Axelsson, J.; Isinger-ekstrand, A.; Van Kuppevelt, T.H.; Johansson, J.; Nilbert, M.; Zaia, J.; et al. Dermatan Sulfate Is Involved in the Tumorigenic Properties of Esophagus Squamous Cell Carcinoma. Cancer Res. 2012, 72, 1943–1953. [Google Scholar] [CrossRef]
- Kato, D.; Era, S.; Watanabe, I.; Arihara, M.; Sugiura, N.; Kimata, K.; Suzuki, Y.; Morita, K.; Hidari, K.I.P.J.; Suzuki, T. Antiviral activity of chondroitin sulphate E targeting dengue virus envelope protein. Antivir. Res. 2010, 88, 236–243. [Google Scholar] [CrossRef]
- Bergefall, K.; Trybala, E.; Johansson, M.; Uyama, T.; Naito, S.; Yamada, S.; Kitagawa, H.; Sugahara, K.; Bergstro, T. Chondroitin Sulfate Characterized by the E-disaccharide Unit Is a Potent Inhibitor of Herpes Simplex Virus Infectivity and Provides the Virus Binding Sites on gro2C Cells. J. Biol. Chem. 2005, 280, 32193–32199. [Google Scholar] [CrossRef] [Green Version]
- Lian, W.; Wu, M.; Huang, N.; Gao, N.; Xiao, C.; Li, Z.; Zhang, Z.; Zheng, Y.; Peng, W.; Zhao, J. Anti-HIV-1 activity and structure-activity-relationship study of a fucosylated glycosaminoglycan from an echinoderm by targeting the conserved CD4 induced epitope. Biochim. Biophys. Acta. 2013, 1830, 4681–4691. [Google Scholar] [CrossRef]
- Huang, N.; Wu, M.; Zheng, C.; Zhu, L.; Zhao, J.; Zheng, Y. The depolymerized fucosylated chondroitin sulfate from sea cucumber potently inhibits HIV replication via interfering with virus entry. Carbohyd. Res. 2013, 380, 64–69. [Google Scholar] [CrossRef]
- Calamia, V.; Mateos, J.; Lourido, L.; Rocha, B.; Montell, E.; Vergés, J.; Blanco, F.J. Anti-angiogenic action of chondroitin sulfate: A novel target for osteoarthritis therapy. Abstr. Osteoarthr. Cartil. 2013, 21, S271. [Google Scholar] [CrossRef]
- Tapon-Bretaudière, J.; Chabut, D.; Zierer, M.; Matou, S.; Helley, D.; Bros, A.; Mourão, P.A.S.; Fischer, A. A Fucosylated Chondroitin Sulfate from Echinoderm Modulates in Vitro Fibroblast Growth Factor 2—Dependent Angiogenesis. Mol. Cancer Res. 2002, 1, 96–102. [Google Scholar]
- Bastos, M.F.; Albrecht, L.; Kozlowski, E.O.; Lopes, S.C.; Blanco, Y.C.; Carlos, B.C.; Castiñeiras, C.; Vicente, C.P.; Werneck, C.C.; Wunderlich, G.; et al. Fucosylated chondroitin sulfate inhibits Plasmodium falciparum cytoadhesion and merozoite invasion. Antimicrob Agents Chemother. 2014, 58, 1862–1871. [Google Scholar] [CrossRef]
- Minamiguchi, K.G.; Kitazato, K.T.; Sasaki, E.; Nagase, H.; Kitazato, K. The Anticoagulant and Hemorrhagic Effects of DHG, a New Depolymerized Holothurian Glycosaminoglycan, on Experimental Hemodialysis in Dogs. Thromb. Haemost. 1997, 77, 1148–1153. [Google Scholar] [CrossRef]
- Tapon-Bretaudière, J.; Drouet, B.; Matou, S.; Mourão, A.S.; Bros, A.; Letourneur, D.; Fischer, A.M. Modulation of Vascular Human Endothelial and Rat Smooth Muscle Cell Growth by a Fucosylated Chondroitin Sulfate from Echinoderm. Thromb. Haemost. 2000, 84, 332–337. [Google Scholar] [CrossRef]
- Melo-filho, N.M.; Belmiro, C.L.; Gonçalves, R.G.; Takiya, C.M.; Leite, M.; Pavão, M.S.G.; Mourão, P.A.S. Fucosylated chondroitin sulfate attenuates renal fibrosis in animals submitted to unilateral ureteral obstruction: A P-selectin-mediated even? Am. J. Physiol. Renal Physiol. 2010, 299, F1299–F1307. [Google Scholar] [CrossRef]
- Hu, S.; Chang, Y.; Wang, J.; Xue, C.; Shi, D.; Xu, H.; Wang, Y. Fucosylated chondroitin sulfate from Acaudina molpadioides improves hyperglycemia via activation of PKB/GLUT4 signaling in skeletal muscle of insulin resistant mice. Food Funct. 2013, 4, 1639–1646. [Google Scholar] [CrossRef]
- Gomes, C.L.; Leão, C.L.; Venturotti, C.; Barreira, A.L.; Guimarães, G.; Fonseca, R.J.; Fortunato, R.S.; Mourão, P.A.; Delgado, A.G.; Takiya, C.M.; et al. The Protective Role of Fucosylated Chondroitin Sulfate, a Distinct Glycosaminoglycan, in a Murine Model of Streptozotocin-Induced Diabetic Nephropathy. PLoS ONE 2014, 9, e106929. [Google Scholar] [CrossRef]
- Hart, G.W.; Copeland, R.J. Glycomics Hits the Big Time. Cell 2010, 143, 672–676. [Google Scholar] [CrossRef] [Green Version]
- Woods Group GLYCAM Web. Complex Carbohydrate Research Center, University of Georgia, Athens, GA. Available online: http://glycam.org (accessed on 30 May 2019).
- Nandini, C.D.; Sugahara, K. Role of the Sulfation Pattern of Chondroitin Sulfate in its Biological Activities and in the Binding of Growth Factors. Adv. Pharmacol. 2006, 53, 253–279. [Google Scholar] [CrossRef]
- Yamada, S.; Sugahara, K.; Özbek, S. Evolution of glycosaminoglycans: Comparative biochemical study. Commun. Integr. Biol. 2011, 4, 150–158. [Google Scholar] [CrossRef]
- Cássaro, C.M.; Dietrich, C.P. Distribution of sulfated mucopolysaccharides in invertebrates. J. Biol. Chem. 1977, 252, 2254–2261. [Google Scholar]
- Medeiros, G.F.; Mendes, A.; Castro, R.A.; Baú, E.C.; Nader, H.B.; Dietrich, C.P. Distribution of sulfated glycosaminoglycans in the animal kingdom: Widespread occurrence of heparin-like compounds in invertebrates. Biochim. Biophys. Acta 2000, 1475, 287–294. [Google Scholar] [CrossRef]
- Sampaio, L.O.; Nader, H.B. Emergence and structural characteristics of chondroitin sulfates in the animal kingdom. Adv. Pharmacol. 2006, 53, 233–251. [Google Scholar] [CrossRef]
- Kinoshita, A.; Yamada, S.; Haslam, S.M.; Morris, H.R.; Dell, A.; Sugahara, K. Isolation and Structural Determination of Novel Sulfated Hexasaccharides from Squid Cartilage Chondroitin Sulfate E That Exhibits Neuroregulatory Activities. Biochemistry 2001, 40, 12654–12665. [Google Scholar] [CrossRef]
- Vázquez, J.; Rodríguez-Amado, I.; Montemayor, M.I.; Fraguas, J.; González, M.; Murado Anxo, M. Chondroitin Sulfate, Hyaluronic Acid and Chitin/Chitosan Production Using Marine Waste Sources: Characteristics, Applications and Eco-Friendly Processes: A Review. Mar. Drugs. 2013, 11, 747–774. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, R.S.; Brito, A.S.; Palhares, L.C.G.F.; Lima, M.A.; Cavalheiro, R.P.; Nader, H.B.; Sassaki, G.L.; Chavante, S.F. 2,3-Di-O-sulfo glucuronic acid: An unmodified and unusual residue in a highly sulfated chondroitin sulfate from Litopenaeus vannamei. Carbohydr. Polym. 2018, 183, 192–200. [Google Scholar] [CrossRef]
- Caterson, B.; Mahmoodian, F.; Sorrell, J.M.; Tim, E.; Bayliss, M.T.; Carney, S.L.; Ratcliffe, A.; Muir, H. Modulation of native chondroitin sulphate structure in tissue development and in disease. J. Cell Sci. 1990, 97 Pt 3, 411–417. [Google Scholar]
- Farrugia, B.L.; Lord, M.S.; Whitelock, J.M.; Melrose, J. Harnessing chondroitin sulphate in composite scaffolds to direct progenitor and stem cell function for tissue repair. Biomate. Sci. 2018, 6, 947–957. [Google Scholar] [CrossRef]
- Sugahara, K.; Mikami, T. Chondroitin/dermatan sulfate in the central nervous system. Curr. Opin. Struct. Biol. 2007, 17, 536–545. [Google Scholar] [CrossRef]
- Nadanaka, S.; Clement, A.; Masayama, K.; Faissner, A.; Sugahara, K.; Biol, M.J.C. Characteristic Hexasaccharide Sequences in Octasaccharides Derived from Shark Cartilage Chondroitin Sulfate D with a Neurite Outgrowth Promoting Activity. J. Biol. Chem. 1998, 273, 3296–3307. [Google Scholar] [CrossRef] [Green Version]
- Shida, M.; Mikami, T.; Tamura, J.; Kitagawa, H. A characteristic chondroitin sulfate trisaccharide unit with a sulfated fucose branch exhibits neurite outgrowth-promoting activity: Novel biological roles of fucosylated chondroitin sulfates isolated from the sea cucumber Apostichopus JPN. Biochem. Biophys. Res. Commun. 2017, 487, 1–6. [Google Scholar] [CrossRef]
- Martel-Pelletier, J.; Farran, A.; Montell, E.; Vergés, J.; Pelletier, J. Discrepancies in Composition and Biological Effects of Different Formulations of Chondroitin Sulfate. Molecules 2015, 20, 4277–4289. [Google Scholar] [CrossRef] [Green Version]
- Bishnoi, M.; Jain, A.; Hurkat, P.; Jain, S.K. Chondroitin sulphate: A focus on osteoarthritis. Glycoconj. J. 2016, 52, 693–705. [Google Scholar] [CrossRef]
- Hochberg, M.C.; Clegg, D.O. Potential effects of chondroitin sulfate on joint swelling: A GAIT report. Osteoarthr. Cartil. 2008, 16, 22–24. [Google Scholar] [CrossRef]
- Hochberg, M.C.; Martel-Pelletier, J.; Monfort, J.; Möller, I.; Castillo, J.R.; Arden, N.; Berenbaum, F.; Blanco, F.J.; Conaghan, P.G.; Doménech, G.; et al. Combined chondroitin sulfate and glucosamine for painful knee osteoarthritis: A multicentre randomised, double-blind, non-inferiority trial, versus. Celecoxib 2016, 75, 37–44. [Google Scholar] [CrossRef]
- Población, C.A.; Michelacci, Y.M. Structural differences of dermatan sulfates from different origins. Carbohydr. Res. 1986, 147, 87–100. [Google Scholar] [CrossRef]
- Sim, J.; Im, A.; Mock, S.; Jin, H.; Ho, J.; Shik, Y. Evaluation of chondroitin sulfate in shark cartilage powder as a dietary supplement: Raw materials and finished products. Food Chem. 2007, 101, 532–539. [Google Scholar] [CrossRef]
- Falshaw, R.; Hubl, U.; Ofman, D.; Slim, G.C.; Tariq, M.A.; Watt, D.K.; Yorke, S.C. Comparison of the glycosaminoglycans isolated from the skin and head cartilage of Gould’s arrow squid (Nototodarus gouldi). Carbohydr. Polym. 2000, 41, 357–364. [Google Scholar] [CrossRef]
- Higashi, K.; Okamoto, Y.; Mukuno, A.; Wakai, J.; Hosoyama, S.; Linhardt, R.J.; Toida, T. Functional chondroitin sulfate from Enteroctopus dofleini containing a 3-O-sulfo glucuronic acid residue. Carbohydr. Polym. 2015, 134, 557–565. [Google Scholar] [CrossRef]
- Sugahara, K.; Tanaka, Y.; Yamada, S.; Seno, N.; Kitagawa, H.; Haslam, S.M.; Morris, H.R.; Dell, A. Novel Sulfated Oligosaccharides Containing 3-O-Sulfated Glucuronic Acid from King Crab Cartilage Chondroitin Sulfate K. Unexpected degradation by chondroitinase ABC. J. Biol. Chem. 1996, 271, 26745–26754. [Google Scholar] [CrossRef]
- Garnjanagoonchorn, W.; Wongekalak, L.; Engkagul, A. Determination of chondroitin sulfate from different sources of cartilage. Chem. Eng. Process. Process. Intensif. 2007, 46, 465–471. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Z.; Linhardt, R.J.; Liu, H.; Linhardt, R.J. Lessons learned from the contamination of heparin. Nat. Prod. Rep. 2009, 26, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Pomin, V.H.; Piquet, A.A.; Pereira, M.S.; Mourão, P.A.S. Residual keratan sulfate in chondroitin sulfate formulations for oral administration. Carbohydr. Polym. 2012, 90, 839–846. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.O.; Kim, M.; Woo, M.; Baek, J.; Kang, K.; Kim, S.H.; Roh, S.S.; Park, C.H.; Jeong, K.S.; Noh, J.S. Chondroitin Sulfate-Rich Extract of Skate Cartilage Attenuates Lipopolysaccharide-Induced Liver Damage in Mice. Mar. Drugs 2017, 15, 178. [Google Scholar] [CrossRef]
- Kawashima, H.; Atarashi, K.; Hirose, M.; Hirose, J.; Yamada, S.; Sugahara, K.; Miyasaka, M. Oversulfated chondroitin/dermatan sulfates containing GlcAβ1/IdoAα1-3GalNAc(4,6-O-disulfate) interact with L- and P-selectin and chemokines. J. Biol. Chem. 2002, 277, 12921–12930. [Google Scholar] [CrossRef]
- Yip, G.W.; Smollich, M.; Go, M. Therapeutic value of glycosaminoglycans in cancer. Mol. Cancer Ther. 2006, 5, 2139–2149. [Google Scholar] [CrossRef]
- Cooney, C.A.; Jousheghany, F.; Yao-borengasser, A.; Phanavanh, B.; Gomes, T.; Kieber-emmons, A.M.; Siegel, E.R.; Suva, L.J.; Ferrone, S.; Kieber-emmons, T. Chondroitin sulfates play a major role in breast cancer metastasis: A role for CSPG4 and CHST11 gene expression in forming surface P-selectin ligands in aggressive breast cancer cells. Breast Cancer Res. 2011, 13, R58. [Google Scholar] [CrossRef]
- Razin, E.; Stevens, R.L.; Akiyama, F.; Schmid, K.; Austen, K.F. Culture from Mouse Bone Marrow of a Subclass of Mast Cells Possessing a Distinct Chondroitin Sulfate Proteoglycan with Glycosaminoglycans Rich in in N-acetylgalactosamine-4,6-disulfate. J. Biol. Chem. 1982, 257, 7229–7236. [Google Scholar]
- Stevens, R.L.; Fox, C.C.; Lichtenstein, L.M.; Austen, K.F. Identification of chondroitin sulfate E proteoglycans and heparin proteoglycans in the secretory granules of human lung mast cells. Proc. Natl. Acad. Sci. USA 1988, 85, 2284–2287. [Google Scholar] [CrossRef]
- Yamada, S.; Sugahara, K. Potential Therapeutic Application of Chondroitin Sulfate/Dermatan Sulfate. Curr. Drug Discov. Technol. 2008, 5, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Hileman, R.E.; Fromm, J.R.; Weiler, J.M.; Linhardt, R.J. Glycosaminoglycan-protein interactions: Definition of consensus sites in glycosaminoglycan binding proteins. BioEssays 1998, 20, 156–167. [Google Scholar] [CrossRef]
- Kjellén, L.; Lindahl, U. Specificity of glycosaminoglycan—Protein interactions. Curr. Opin. Struct. Biol. 2018, 50, 101–108. [Google Scholar] [CrossRef]
- Yamaguchi, I.; Suda, H.; Tsuzuike, N.; Seto, K.; Seki, M.; Yamaguchi, Y.; Hasegawa, K.; Takahashi, N.; Yamamoto, S.; Gejyo, F.; et al. Glycosaminoglycan and proteoglycan inhibit the depolymerization of β2-microglobulin amyloid fibrils in vitro. Kindey Int. 2003, 64, 1080–1088. [Google Scholar] [CrossRef]
- Kimura, K.; Matsubara, H.; Sogoh, S.; Kita, Y.; Sakata, T.; Nishitani, Y.; Watanabe, S.; Hamaoka, T.; Fujiwara, H. Role of glycosaminoglycans in the regulation of T cell proliferation induced by thymic stroma-derived T cell growth factor. J. Immunol. 1991, 146, 2618–2624. [Google Scholar]
- Vallières, M.; Souich, P. Modulation of inflammation by chondroitin sulfate. Osteoarthr. Cartil. 2010, 18, 18–23. [Google Scholar] [CrossRef]
- Du Souich, P.; García, A.G.; Vergés, J.; Montell, E. Immunomodulatory and anti-inflammatory effects of chondroitin sulphate. J. Cell. Mol. Med. 2009, 13, 1451–1463. [Google Scholar] [CrossRef]
- Linares, P.M.; Chaparro, M.; Algaba, A.; Román, M.; Moreno Arza, I.; Abad Santos, F.; Ochoa, D.; Guerra, I.; Bermejo, F.; Gisbert, J.P. Effect of Chondroitin Sulphate on Pro-Inflammatory Mediators and Disease Activity in Patients with Inflammatory Bowel Disease. Digestion 2015, 92, 203–210. [Google Scholar] [CrossRef]
- Schön, M.P. Inhibitors of selectin functions in the treatment of inflammatory skin disorders. Ther. Clin. Risk Manag. 2005, 1, 201–208. [Google Scholar]
- Deshauer, C.; Morgan, A.M.; Prestegard, J.H.; Wang, X.; Deshauer, C.; Morgan, A.M.; Ryan, E.O.; Handel, T.M.; Prestegard, J.H.; Wang, X. Interactions of the Chemokine CCL5/RANTES with Medium-Sized Chondroitin Sulfate Ligands. Cell 2015, 23, 1066–1077. [Google Scholar] [CrossRef] [Green Version]
- Willis, C.M.; Klu, M. Chondroitin Sulfate-E Is a Negative Regulator of a Pro- Tumorigenic Wnt/Beta-Catenin-Collagen 1 Axis in Breast Cancer Cells. PLoS ONE 2014, 9, 1–10. [Google Scholar] [CrossRef]
- Barthel, S.R.; Gavino, J.D.; Descheny, L.; Dimitroff, C.J. Targeting selectins and selectin ligands in inflammation and cancer. J. Expert Opin. Ther. Targets. 2007, 11, 1473–1491. [Google Scholar] [CrossRef]
- Kim, Y.J.; Borsig, L.; Han, H.; Varki, N.M.; Varki, A. Distinct Selectin Ligands on Colon Carcinoma Mucins Can Mediate Pathological Interactions among Platelets, Leukocytes, and Endothelium. Am. J. Pathol. 1999, 155, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Laabs, T.; Carulli, D.; Geller, H.M.; Fawcett, J.W. Chondroitin sulfate proteoglycans in neural development and regeneration. Curr. Opin. Neurobiol. 2005, 15, 116–120. [Google Scholar] [CrossRef]
- Rauvala, H.; Paveliev, M.; Kuja-panula, J.; Kulesskaya, N. Inhibition and enhancement of neural regeneration by chondroitin sulfate proteoglycans. Neural Regen. Res. 2017, 12, 687–692. [Google Scholar] [CrossRef]
- Siebert, J.R.; Steencken, A.C.; Osterhout, D.J. Chondroitin Sulfate Proteoglycans in the Nervous System: Inhibitors to Repair. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Ito, Z.; Sakamoto, K.; Imagama, S.; Matsuyama, Y.; Zhang, H.; Hirano, K.; Ando, K.; Yamashita, T.; Ishiguro, N.; Kadomatsu, K. N -Acetylglucosamine 6- O -Sulfotransferase-1-Deficient Mice Show Better Functional Recovery after Spinal Cord Injury. J. Neurosci. 2010, 30, 5937–5947. [Google Scholar] [CrossRef]
- Imagama, S.; Sakamoto, K.; Tauchi, R.; Shinjo, R.; Ohgomori, T.; Ito, Z.; Zhang, H.; Nishida, Y.; Asami, N.; Takeshita, S.; et al. Keratan Sulfate Restricts Neural Plasticity after Spinal Cord Injury. J. Neurosci. 2011, 31, 17091–17102. [Google Scholar] [CrossRef] [Green Version]
- Ueoka, C.; Kaneda, N.; Okazaki, I.; Nadanaka, S.; Muramatsu, T.; Sugahara, K. Neuronal Cell Adhesion, mediated by the Heparin-binding Neuroregulatory Factor Midkine, Is Specifically Inhibited by Chondroitin Sulfate E. J. Biol. Chem. 2000, 275, 37407–37413. [Google Scholar] [CrossRef]
- Maeda, N.; Fukazawa, N.; Hata, T. The Binding of Chondroitin Sulfate to Pleiotrophin/Heparin- binding Growth-associated Molecule Is Regulated by Chain Length and Oversulfated Structures. J. Biol. Chem. 2005, 281, 4894–4902. [Google Scholar] [CrossRef]
- Maeda, N.; He, J.; Yajima, Y.; Mikami, T.; Sugahara, K.; Yabe, T. Heterogeneity of the Chondroitin Sulfate Portion of Phosphacan/6B4 Proteoglycan Regulates Its Binding Affinity for Pleiotrophin/Heparin Binding Growth-associated Molecule. J. Biol. Chem. 2003, 278, 35805–35811. [Google Scholar] [CrossRef] [Green Version]
- Maeda, N.; Nishiwaki, T.; Shintani, T.; Hamanaka, H.; Noda, M. 6B4 Proteoglycan/Phosphacan, an Extracellular Variant of Receptor-like Protein-tyrosine Phosphatase ζ/RPTP β, Binds Pleiotrophin/Heparin-binding Growth-associated Molecule. J. Biol. Chem. 1996, 271, 21446–21452. [Google Scholar] [CrossRef]
- Deepa, S.S.; Umehara, Y.; Higashiyama, S.; Itoh, N.; Sugahara, K. Specific Molecular Interactions of Oversulfated Chondroitin Sulfate E with Various Heparin-binding Growth Factors. J. Biol. Chem. 2002, 277, 43707–43716. [Google Scholar] [CrossRef] [Green Version]
- Rani, A.; Patel, S.; Goyal, A. Chondroitin Sulfate (CS) Lyases: Structure, Function and Application in Therapeutics. Curr. Protein Pept. Sci. 2018, 19, 22–33. [Google Scholar] [CrossRef]
- Lin, Y.; Li, L.; Zhang, F.; Linhardt, R.J. Borrelia burgdorferi glycosaminoglycan-binding proteins: A potential target for new therapeutics against Lyme disease. Microbiology 2017, 163, 1759–1766. [Google Scholar] [CrossRef]
- Chaffee, K.M.A.E. Mucopolysaccharides of Skin. J. Biol. Chem. 1941, 138, 491–499. [Google Scholar]
- Singla, S.K. Dermatan sulphate structure biosynthesis and functions. Biochem. Educ. 1988, 16, 156–157. [Google Scholar] [CrossRef]
- Trowbridge, J.M.; Gallo, R.L. Dermatan sulfate: New functions from an old glycosaminoglycan. Glycobiology 2002, 12, 117R–125R. [Google Scholar] [CrossRef]
- Osborne, S.A.; Daniel, R.A.; Desilva, K.; Seymour, R.B. Antithrombin activity and disaccharide composition of dermatan sulfate from different bovine tissues. Glycobiology 2008, 18, 225–234. [Google Scholar] [CrossRef]
- Malavaki, C.; Mizumoto, S.; Karamanos, N.; Sugahara, K. Recent advances in the structural study of functional chondroitin sulfate and dermatan sulfate in health and disease. Connect. Tissue Res. 2008, 49, 133–139. [Google Scholar] [CrossRef]
- Vasconcelos, A.; Pomin, V. The Sea as a Rich Source of Structurally Unique Glycosaminoglycans and Mimetics. Microorganisms 2017, 5, 51. [Google Scholar] [CrossRef]
- Bossennec, V.; Petitou, M.; Perly, B. 1H-n.m.r. investigation of naturally occurring and chemically oversulphated dermatan sulphates. Identification of minor monosaccharide residues. Biochem. J. 1990, 267, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Pavão, M.S.; Aiello, K.R.; Werneck, C.C.; Silva, L.C.F.; Valente, A.P.; Mulloy, B.; Colwell, N.S.; Tollefsen, D.M.; Mourão, P.A.S. Highly sulfated dermatan sulfates from Ascidians. Structure versus anticoagulant activity of these glycosaminoglycans. J. Biol. Chem. 1998, 273, 27848–27857. [Google Scholar] [CrossRef]
- Pavão, M.; Mourão, P.; Mulloy, B.; Tollefsen, D. A unique dermatan sulfate-like glycosaminoglycan from Ascidian: Its structure and the effect of its unusual sulfation pattern on anticoagulant activity. J. Biol. Chem. 1995, 270, 31027–31036. [Google Scholar] [CrossRef]
- Pavão, M.S.G. Structure and anticoagulant properties of sulfated glycosaminoglycans from primitive Chordates. An. Acad. Bras. Cienc. 2002, 74, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Pomin, V.H. A Dilemma in the Glycosaminoglycan-Based Therapy: Synthetic or Naturally Unique Molecules? Med. Res. Rev. 2015, 35, 1195–1219. [Google Scholar] [CrossRef]
- Sanderson, P.N.; Huckerby, T.N.; Nieduszynski, I.A. Chondroitinase ABC digestion of dermatan sulphate. N.m.r. spectroscopic characterization of the oligo- and poly-saccharides. Biochem. J. 1989, 257, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Pavão, M.S.G.; Rodrigues, M.A.; Mourão, P.A.S. Acidic polysaccharides of the ascidian Styela plicata. Biosynthetic studies on the sulfated l-galactans of the tunic, and preliminary characterization of a dermatan sulfate-like polymer in body tissues. BBA Gen. Subj. 1994, 1199, 229–237. [Google Scholar] [CrossRef]
- Anno, K.; Seno, N.; Mathews, M.B.; Yamagata, T.; Suzukic, S. A new dermatan polysulfate, chondroitin sulfate H, from hagfish notochord. BBA Gen. Subj. 1971, 237, 173–177. [Google Scholar] [CrossRef]
- Seno, N.; Fumiko, A.; Anno, K. A novel dermatan polysulfate from hagfish skin, containing trisulfated disaccharide residues. BBA Gen. Subj. 1972, 264, 229–233. [Google Scholar] [CrossRef]
- Sugahara, K.; Yamada, S. Structure and Function Unique Sulfation of Oversulfated Chondroitin Patterns and Neuroregulatory Sulfate Variants. Trends Glycosci. Glyc. 2000, 12, 321–349. [Google Scholar] [CrossRef]
- Casu, B.; Guerrini, M.; Torri, G. Structural and conformational aspects of the anticoagulant and anti-thrombotic activity of heparin and dermatan sulfate. Curr. Pharm. Des. 2004, 10, 939–949. [Google Scholar] [CrossRef]
- Fernandez, F.; Van Ryn, J.; Ofosu, F.A.; Hirsh, J.; Buchanan, M.R. The haemorrhagic and antithrombotic effects of dermatan sulphate. Br. J. Haematol. 1986, 64, 309–317. [Google Scholar] [CrossRef]
- Linhardt, R.J.; Hileman, R.E. Dermatan sulfate as a potential therapeutic agent. Gen. Pharmacol. 1995, 26, 443–451. [Google Scholar] [CrossRef]
- Buchanan, M.R.; Brister, S.J. Anticoagulant and antithrombin effects of intimatan, a heparin cofactor II agonist. Thromb. Res. 2000, 99, 603–612. [Google Scholar] [CrossRef]
- Buchanan, M.R.; Maclean, G.A.; Brister, S.J. Selective and sustained inhibition of surface-bound thrombin activity by intimatan/heparin cofactor II and its relevance to assessing systemic anticoagulation in vivo, ex vivo and in vitro. Thromb. Haemost. 2001, 86, 909–913. [Google Scholar]
- Hennan, J.K.; Hong, T.T.; Shergill, A.K.; Driscoll, E.M.; Cardin, A.D.; Lucchesi, B.R. Intimatan prevents arterial and venous thrombosis in a canine model of deep vessel wall injury. J. Pharmacol. Exp. Ther. 2002, 301, 1151–1156. [Google Scholar] [CrossRef]
- Hong, T.T.; White, A.J.; Lucchesi, B.R. Dermatan disulfate (Intimatan) prevents complement-mediated myocardial injury in the human-plasma-perfused rabbit heart. Int. Immunopharmacol. 2005, 5, 381–391. [Google Scholar] [CrossRef]
- Hong, T.T.; Van Gorp, C.L.; Cardin, A.D.; Lucchesi, B.R. Intimatan (dermatan 4,6-O-disulfate) prevents rethrombosis after successful thrombolysis in the canine model of deep vessel wall injury. Thromb. Res. 2006, 117, 333–342. [Google Scholar] [CrossRef]
- Tanaka, K.A.; Szlam, F.; Vinten-Johansen, J.; Cardin, A.D.; Levy, J.H. Effects of antithrombin and heparin cofactor II levels on anticoagulation with Intimatan. Thromb. Haemost. 2005, 94, 808–813. [Google Scholar] [CrossRef]
- Mascellani, G.; Liverani, L.; Prete, A.; Guppola, P.A.; Bergonzini, G.; Bianchini, P. Relative influence of different disulphate disaccharide clusters on the HCII-mediated inhibition of thrombin by dermatan sulphates of different origins. Thromb. Res. 1994, 74, 605–615. [Google Scholar] [CrossRef]
- Mascellani, G.; Liverani, L.; Prete, A.; Bergonzini, G.; Bianchini, P.; Torri, G.; Bisio, A.; Guerrini, M.; Casu, B. Quantitation of dermatan sulfate active site for heparin cofactor II by 1H nuclear magnetic resonance spectroscopy. Anal. Biochem. 1994, 223, 135–141. [Google Scholar] [CrossRef]
- Maimone, M.M.; Tollefsen, D.M. Structure of a dermatan sulfate hexasaccharide that binds to heparin cofactor II with high affinity. J. Biol. Chem. 1990, 265, 18263–18271. [Google Scholar]
- Basten, J.E.M.; Van Boeckel, C.A.A.; Jaurand, G.; Petitou, M.; Soijker, N.M.; Westerduin, P. Synthesis of a dermatan sulphate-like hexasaccharide with a “non-glycosamino” glycan structure. Bioorg. Med. Chem. Lett. 1994, 4, 893–898. [Google Scholar] [CrossRef]
- Yang, H.O.; Gunay, N.S.; Toida, T.; Kuberan, B.; Yu, G.; Kim, Y.S.; Linhardt, R.J. Preparation and structural determination of dermatan sulfate-derived oligosaccharides. Glycobiology 2000, 10, 1033–1039. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Lassepas, M.; Quinones, M.R. Heparin therapy for stroke Hemorrhagic complications and risk factors for intracerebral hemorrhage. Neurology 1984, 34, 114. [Google Scholar] [CrossRef]
- Hirsh, J. Heparin induced bleeding. Nouv. Rev. Fr. Hematol. 1984, 26, 261–266. [Google Scholar]
- Qin, X.; Zhang, B. Cerebral Hemorrhage Complicating Heparin—Induced Thrombocytopenia after Percutaneous Coronary Intervention: A Clinical Dilemma of Medical Treatment. Chin. Med. J. 2015, 128, 1561–1562. [Google Scholar] [CrossRef]
- Babikian, V.L.; Kase, C.S.; Pessin, M.S.; Norrving, B.; Gorelick, P.B. Intracerebral Hemorrhage in Stroke Patients Anticoagulated with Heparin. Stroke 1989, 20, 1500–1503. [Google Scholar] [CrossRef]
- Gianese, F.; Nurmohamed, M.; Imbimbo, B.; Buller, H.; Berckmans, R.; Ten Cate, J. The pharmacokinetics and pharmacodynamics of dermatan sulphate MF701 during haemodialysis for chronic renal failure. Br. J. Clin. Pharmacol. 1993, 35, 335–339. [Google Scholar] [CrossRef]
- Tollefsens, D.M.; Peacock, M.E.; Monafo, W.J. Molecular Size of Dermatan Sulfate Oligosaccharides Required to Bind and Activate Heparin Cofactor II. J. Biol. Chem. 1986, 261, 8854–8858. [Google Scholar]
- Vicente, C.P.; He, L.; Pavão, M.S.G.; Tollefsen, D.M. Antithrombotic activity of dermatan sulfate in heparin cofactor II-deficient mice. Blood 2004, 104, 3965–3970. [Google Scholar] [CrossRef]
- Linhardt, R.J.; Desai, U.R.; Liu, J.; Pervin, A.; Hoppensteadt, D.; Fareed, J. Low molecular weight dermatan sulfate as an antithrombotic agent structure-activity relationship studies. Biochem. Pharmacol. 1994, 47, 1241–1252. [Google Scholar] [CrossRef]
- Linhardt, R.J.; Al-Hakim, A.; Liu, J.; Hoppensteadt, D.; Mascellani, G.; Bianchini, P.; Fareed, J. Structural features of dermatan sulfates and their relationship to anticoagulant and antithrombotic activities. Biochem. Pharmacol. 1991, 42, 1609–1619. [Google Scholar] [CrossRef]
- Sheehan, J.P.; Tollefsen, D.M.; Sadler, J.E. Heparin cofactor II is regulated allosterically and not primarily by template effects. Studies with mutant thrombins and glycosaminoglycans. J. Biol. Chem. 1994, 269, 32747–32751. [Google Scholar]
- Plichta, J.K.; Radek, K.A. Sugar-coating wound repair: A review of FGF-10 and dermatan sulfate in wound healing and their potential application in burn wounds. J. Burn Care Res. 2012, 33, 299–310. [Google Scholar] [CrossRef]
- Penc, S.F.; Pomahac, B.; Winkler, T.; Dorschner, R.A.; Eriksson, E.; Herndon, M.; Gallo, R.L. Dermatan sulfate released after injury is a potent promoter of fibroblast growth factor-2 function. J. Biol. Chem. 1998, 273, 28116–28121. [Google Scholar] [CrossRef]
- Radek, K.A.; Taylor, K.R.; Gallo, R.L. FGF-10 and specific structural elements of dermatan sulfate size and sulfation promote maximal keratinocyte migration and cellular proliferation. Wound Repair Regen. 2009, 17, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Trowbridge, J.M.; Rudisill, J.A.; Ron, D.; Gallo, R.L. Dermatan sulfate binds and potentiates activity of keratinocyte growth factor (FGF-7). J. Biol. Chem. 2002, 277, 42815–42820. [Google Scholar] [CrossRef]
- Priglinger, U.; Geiger, M.; Bielek, E.; Vanyek, E.; Binder, B.R. Binding of urinary protein C inhibitor to cultured human epithelial kidney tumor cells (TCL-598). The role of glycosaminoglycans present on the luminal cell surface. J. Biol. Chem. 1994, 269, 14705–14710. [Google Scholar]
- Iozzo, R.V. The family of the small leucine-rich proteoglycans: Key regulators of matrix assembly and cellular growth. Crit. Rev. Biochem. Mol. Biol. 1997, 32, 141–174. [Google Scholar] [CrossRef]
- Tumova, S.; Woods, A.; Couchman, J.R. Heparan sulfate chains from glypican and syndecans bind the Hep II domain of fibronectin similarly despite minor structural differences. J. Biol. Chem. 2000, 275, 9410–9417. [Google Scholar] [CrossRef]
- Jungmann, O.; Nikolovska, K.; Stock, C.; Schulz, J.; Eckes, B.; Owens, R.T.; Iozzo, R.V.; Seidler, D.G.; Riethmu, C. The dermatan sulfate proteoglycan decorin modulates α2β1 integrin and the vimentin intermediate filament system during collagen synthesis. PLoS ONE 2012, 7, e50809. [Google Scholar] [CrossRef]
- Seo, E.S.; Blaum, S.; Vargues, T.; De Cecco, M.; Deakin, J.A.; Lyon, M.; Barran, P.E.; Campopiano, D.J.; Uhrín, D. Interaction of Human β -Defensin 2 (HBD2) with Glycosaminoglycans. Biochemistry 2010, 2, 10486–10495. [Google Scholar] [CrossRef]
- Brooks, B.; Briggs, D.M.; Eastmond, N.C.; Fernig, D.G.; Coleman, J.W. Presentation of IFN-γ to Nitric Oxide-Producing Cells: A Novel Function for Mast Cells. J. Immunol. 2000, 164, 573–579. [Google Scholar] [CrossRef]
- Hildebrand, A.; Romarís, M.; Rasmussen, L.M.; Heinegård, D.; Twardzik, D.R.; Border, W.A.; Ruoslahti, E. Interaction of the small interstitial proteoglycans biglycan, decorin and fibromodulin with transforming growth factor beta. Biochem. J. 1994, 302, 527–534. [Google Scholar] [CrossRef]
- Belmiro, C.L.R.; Souza, H.S.P.; Elia, C.C.S.; Castelo-Branco, M.T.L.; Silva, F.R.; Machado, R.L.; Pavão, M.S.G. Biochemical and immunohistochemical analysis of glycosaminoglycans in inflamed and non-inflamed intestinal mucosa of patients with Crohn’s disease. Int. J. Colorectal Dis. 2005, 20, 295–304. [Google Scholar] [CrossRef]
- Belmiro, C.L.R.; Gonçalves, R.G.; Kozlowski, E.O.; Werneck, A.F.; Takyia, C.M.; Leite, M., Jr.; Pavão, M.S.G. Dermatan sulfate reduces monocyte chemoattractant protein 1 and TGF-β production, as well as macrophage recruitment and myofibroblast accumulation in mice with unilateral ureteral obstruction. Brazilian J. Med. Biol. Res. 2011, 44, 624–633. [Google Scholar] [CrossRef]
- Gruber, S.; Frings, K.; Kuess, P.; Dörr, W. Protective effects of systemic dermatan sulfate treatment in apreclinical model of radiation-induced oral mucositis. Strahlenthe. Onkol. 2018, 194, 675–685. [Google Scholar] [CrossRef]
- Gruber, S.; Arnold, M.; Cini, N.; Gernedl, V.; Hetzendorfer, S.; Kowald, L.M.; Kuess, P.; Mayer, J.; Morava, S.; Pfaffinger, S.; et al. Radioprotective effects of dermatan sulfate in a preclinical model of oral mucositis—targeting inflammation, hypoxia and junction proteins without stimulating proliferation. Int. J. Mol. Sci. 2018, 19, 1684. [Google Scholar] [CrossRef]
- Afratis, N.; Gialeli, C.; Nikitovic, D.; Tsegenidis, T.; Karousou, E.; Tzanakakis, G.N.; Karamanos, N.K. Glycosaminoglycans: Key players in cancer cell biology and treatment. FEBS J. 2012, 279, 1177–1197. [Google Scholar] [CrossRef]
- Kozlowski, E.O.; Pavao, M.S.G.; Borsig, L. Ascidian dermatan sulfates attenuate metastasis, inflammation and thrombosis by inhibition of P-selectin. J. Thromb. Haemost. 2011, 9, 1807–1815. [Google Scholar] [CrossRef] [Green Version]
- Di Caro, A.; Perola, E.; Bartoloni, B.; Marzano, M.; Liverani, L.; Mascellani, G.; Benedetto, A.; Cellai, L. Fractions of chemically oversulphated galactosaminoglycan sulphates inhibit three enveloped viruses: Human immunodeficiency virus type 1, herpes simplex virus type 1 and human cytomegalovirus. Antivir. Chem. Chemother. 1999, 10, 33–38. [Google Scholar] [CrossRef]
- Nagase, B.H.; Enjyoji, K.; Minamiguchi, K.; Kitazato, K.T.; Kitazato, K.; Saito, H. Depolymerized holothurian glycosaminoglycan with novel anticoagulant actions: Antithrombin III- and heparin cofactor II-independent inhibition of factor X activation by factor IXa-factor VIIIa complex and heparin cofactor II-dependent inhibition of thrombin. Blood 1995, 85, 1527–1534. [Google Scholar]
- Wu, M.; Wen, D.; Gao, N.; Xiao, C.; Yang, L.; Xu, L.; Lian, W.; Peng, W.; Jiang, J.; Zhao, J. Anticoagulant and antithrombotic evaluation of native fucosylated chondroitin sulfates and their derivatives as selective inhibitors of intrinsic factor Xase. Eur. J. Med. Chem. 2015, 92, 257–269. [Google Scholar] [CrossRef]
- Lidholt, K.; Fjelstad, M. Biosynthesis of the Escherichia coli K4 Capsule Polysaccharide A parallel system for studies of glycosyltransferases in chondroitin formation. J. Biol. Chem. 1997, 272, 2682–2687. [Google Scholar] [CrossRef]
- Habuchi, O.; Sugiura, K.; Kawai, N.; Suzuki, S. Glucose Branches in Chondroitin Sulfates from Squid Cartilage. J. Biol. Chem. 1977, 252, 4570–4576. [Google Scholar]
- Higashi, K.; Takeda, K.; Mukuno, A.; Okamoto, Y.; Masuko, S.; Linhardt, R.J.; Toida, T. Identification of keratan sulfate disaccharide at C-3 position of glucuronate of chondroitin sulfate from Mactra chinensis. Biochem. J. 2016, 473, 4145–4158. [Google Scholar] [CrossRef]
- Lord, M.S.; Day, A.J.; Youssef, P.; Zhuo, L.; Watanabe, H.; Caterson, B.; Whitelock, J.M. Sulfation of the bikunin chondroitin sulfate chain determines heavy chain·hyaluronan complex formation. J. Biol. Chem. 2013, 288, 22930–22941. [Google Scholar] [CrossRef]
- Zhuo, L.; Hascall, V.C.; Kimata, K. Inter-alpha-trypsin inhibitor, a covalent protein-glycosaminoglycan-protein complex. J. Biol. Chem. 2004, 279, 38079–38082. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Nifantiev, N.E.; Usov, A.I. New insight on the structural diversity of holothurian fucosylated chondroitin sulfates. Pure Appl. Chem. 2019, 91, 1–7. [Google Scholar] [CrossRef]
- Vieira, R.P.; Mulloy, B.; Mourão, P.A. Structure of a fucose-branched chondroitin sulfate from sea cucumber. Evidence for the presence of 3-O-sulfo-beta-d-glucuronosyl residues. J. Biol. Chem. 1991, 266, 13530–13536. [Google Scholar]
- Pomin, V.H. Medical Gains of Chondroitin Sulfate Upon Fucosylation. Curr. Med. Chem. 2015, 22, 4166–4176. [Google Scholar] [CrossRef]
- Fonseca, R.J.C.; Santos, G.R.C.; Mourão, P.A.S. Effects of polysaccharides enriched in 2,4-disulfated fucose units on coagulation, thrombosis and bleeding Practical and conceptual implications. Thromb. Haemost. 2009, 102, 829–836. [Google Scholar] [CrossRef]
- Chen, S.; Xue, C.; Tang, Q.; Yu, G.; Chai, W. Comparison of structures and anticoagulant activities of fucosylated chondroitin sulfates from different sea cucumbers. Carbohydr. Polym. 2011, 83, 688–696. [Google Scholar] [CrossRef]
- Wu, M.; Xu, S.; Zhao, J.; Kang, H.; Ding, H. Physicochemical characteristics and anticoagulant activities of low molecular weight fractions by free-radical depolymerization of a fucosylated chondroitin sulphate from sea cucumber Thelenata Ananas. Food Chem. 2010, 122, 716–723. [Google Scholar] [CrossRef]
- Wu, M.; Huang, R.; Wen, D.; Gao, N.; He, J.; Li, Z.; Zhao, J. Structure and effect of sulfated fucose branches on anticoagulant activity of the fucosylated chondroitin sulfate from sea cucumber Thelenata ananas. Carbohydr. Polym. 2012, 87, 862–868. [Google Scholar] [CrossRef]
- Kariya, Y.; Watabe, S.; Kyogashima, M.; Ishihara, M.; Ishii, T. Structure of fucose branches in the glycosaminoglycan from the body wall of the sea cucumber Stichopus japonicus. Carbohydr. Res. 1997, 297, 273–279. [Google Scholar] [CrossRef]
- Luo, L.; Wu, M.; Xu, L.; Lian, W.; Xiang, J.; Lu, F.; Gao, N.; Xiao, C.; Wang, S.; Zhao, J. Comparison of Physicochemical Characteristics and Anticoagulant Activities of Polysaccharides from Three Sea Cucumbers. Mar. Drugs 2013, 11, 399–417. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Xu, L.; Li, J. Preparation and anticoagulant activity of a fucosylated polysaccharide sulfate from a sea cucumber Acaudina molpadioidea. Carbohydr. Polym. 2012, 87, 2052–2057. [Google Scholar] [CrossRef]
- Matsuhiro, B.; Osorio-román, I.O.; Torres, R. Vibrational spectroscopy characterization and anticoagulant activity of a sulfated polysaccharide from sea cucumber Athyonidium chilensis. Carbohydr. Polym. 2012, 88, 959–965. [Google Scholar] [CrossRef]
- Santos, G.R.C.; Glauser, B.F.; Parreiras, L.A.; Vilanova, E. Distinct structures of the α -fucose branches in fucosylated chondroitin sulfates do not affect their anticoagulant activity. Glycobiology 2015, 25, 1–32. [Google Scholar] [CrossRef]
- Ustyuzhanina, A.N.E.; Bilan, M.I.; Dmitrenok, S.; Borodina, E.Y.; Stonik, V.A.; Nifantiev, N.E.; Usov, A.I. A highly regular fucosylated chondroitin sulfate from the sea cucumber Massinium magnum: Structure and effects on coagulation. Carbohydr. Polym. 2017, 167, 20–26. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Tsvetkova, E.A.; Shashkov, A.S.; Stonik, V.A.; Nifantiev, N.E.; Usov, A.I. Structural characterization of fucosylated chondroitin sulfates from sea cucumbers Apostichopus japonicus and Actinopyga mauritiana. Carbohydr. Polym. 2016, 153, 399–405. [Google Scholar] [CrossRef]
- Li, Q.; Cai, C.; Chang, Y.; Zhang, F.; Linhardt, R.J.; Xue, C.; Li, G.; Yu, G. A novel structural fucosylated chondroitin sulfate from Holothuria Mexicana and its effects on growth factors binding and anticoagulation. Carbohydr. Polym. 2017, 181, 1160–1168. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Panina, E.G.; Sanamyan, N.P.; Dmitrenok, A.S.; Tsvetkova, E.A.; Ushakova, N.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure and Anti-Inflammatory Activity of a New Unusual Fucosylated Chondroitin Sulfate from Cucumaria djakonovi. Mar. Drugs 2018, 16, 389. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Nifantiev, N.E.; Usov, A.I. Two fucosylated chondroitin sulfates from the sea cucumber Eupentacta fraudatrix. Carbohydr. Polym. 2017, 164, 8–12. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Shashkov, A.S. Two structurally similar fucosylated chondroitin sulfates from the holothurian species Stichopus chloronotus and Stichopus horrens. Carbohydr. Polym. 2018, 189, 10–14. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Yang, S.; Lv, Z. Separation, purification, structures and anticoagulant activities of fucosylated chondroitin sulfates from Holothuria scabra. Int. J. Biol. Macromol. 2017, 108, 710–718. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Nifantiev, N.E. Fucosylated chondroitin sulfates from the sea cucumbers Holothuria tubulosa and Holothuria stellati. Carbohydr. Polym. 2018, 200, 1–5. [Google Scholar] [CrossRef]
- Panagos, C.G.; Thomson, D.S.; Moss, C.; Hughes, A.D.; Kelly, M.S.; Liu, Y.; Chai, W.; Venkatasamy, R.; Spina, D.; Page, C.P.; et al. Fucosylated Chondroitin Sulfates from the Body Wall of the Sea Cucumber Holothuria forskali. J. Biol. Chem. 2014, 289, 28284–28298. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. The structure of a fucosylated chondroitin sulfate from the sea cucumber Cucumaria frondosa. Carbohydr. Polym. 2017, 165, 7–12. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Shashkov, A.S.; Kusaykin, M.I.; Stonik, V.A.; Nifantiev, N.E.; Usov, A.I. Structure and biological activity of a fucosylated chondroitin sulfate from the sea cucumber Cucumaria japonica. Glycobiology 2015, 26, 449–459. [Google Scholar] [CrossRef]
- Boisson-vidal, C.; Tapon-bretaudie, J.; Mourão, P.A.S. Inactivation of Thrombin by a Fucosylated Chondroitin Sulfate from Echinoderm. Thromb. Res. 2001, 102, 167–176. [Google Scholar] [CrossRef]
- Chen, S.; Li, G.; Wu, N.; Guo, X.; Liao, N.; Ye, X.; Liu, D.; Xue, C.; Chai, W. Sulfation pattern of the fucose branch is important for the anticoagulant and antithrombotic activities of fucosylated chondroitin sulfates. BBA Gen. Subj. 2013, 1830, 3054–3066. [Google Scholar] [CrossRef]
- Li, J.; Li, S.; Yan, L.; Ding, T.; Linhardt, R.J.; Yu, Y.; Liu, X.; Liu, D.; Ye, X.; Chen, S. Fucosylated chondroitin sulfate oligosaccharides exert anticoagulant activity by targeting at intrinsic tenase complex with low FXII activation: Importance of sulfation pattern and molecular size. Eur. J. Med. Chem. 2017, 139, 191–200. [Google Scholar] [CrossRef]
- Buyue, Y.; Sheehan, J.P. Fucosylated Chondroitin Sulfate Inhibits Plasma Thrombin Generation via Targeting of the Factor IXa Heparin-Binding Exosite. Blood 2009, 114, 3092–3100. [Google Scholar] [CrossRef]
- Zhao, L.; Wu, M.; Xiao, C.; Yang, L.; Zhou, L.; Gao, N.; Li, Z.; Chen, J. Discovery of an intrinsic tenase complex inhibitor: Pure nonasaccharide from fucosylated glycosaminoglycan. Proc. Natl. Acad. Sci. USA 2015, 112, 8284–8289. [Google Scholar] [CrossRef] [Green Version]
- Glauser, B.F.; Pereira, M.S.; Monteiro, R.Q.; Mourão, P.A.S. Serpin-independent anticoagulant activity of a fucosylated chondroitin sulfate. Thromb. Haemost. 2008, 420–428. [Google Scholar] [CrossRef]
- Glauser, B.F.; Mourão, P.A.S.; Pomin, V.H. Marine Sulfated Glycans with Serpin-Unrelated Anticoagulant Properties, 1st ed.; Elsevier Inc.: San Diego, CA, USA, 2013; Volume 62. [Google Scholar]
- Sheehan, J.P.; Walke, E.N. Depolymerized holothurian glycosaminoglycan and heparin inhibit the intrinsic tenase complex by a common antithrombin-independent mechanism. Blood 2019, 107, 3876–3883. [Google Scholar] [CrossRef]
- Pomin, V.H. Anticoagulant motifs of marine sulfated glycans. Glycoconj. J. 2014, 31, 341–344. [Google Scholar] [CrossRef]
- Ben Mansour, M.; Balti, R.; Ollivier, V.; Ben Jannet, H.; Chaubet, F.; Maaroufi, R.M. Characterization and anticoagulant activity of a fucosylated chondroitin sulfate with unusually procoagulant effect from sea cucumber. Carbohydr. Polym. 2017, 174, 760–771. [Google Scholar] [CrossRef]
- Pomin, V.H. Dual and antagonic therapeutic effects of sulfated glycans. Bioorg. Med. Chem. 2016, 24, 3965–3971. [Google Scholar] [CrossRef]
- Minamiguchi, K.; Kitazato, K.T.; Nagase, H.; Sasaki, E.; Ohwada, K.; Kitazato, K. Depolymerized holothurian glycosaminoglycan (DHG), a novel alternative anticoagulant for hemodialysis, is safe and effective in a dog renal failure model. Kidney Int. 2003, 63, 1548–1555. [Google Scholar] [CrossRef] [Green Version]
- Tovar, A.M.F.; Mourão, P.A.S. High affinity of a fucosylated chondroitin sulfate for plasma low density lipoprotein. Atherosclerosis 1996, 126, 185–195. [Google Scholar] [CrossRef]
- Fonseca, R.J.C.; Mourão, P.A.S. Fucosylated chondroitin sulfate as a new oral antithrombotic agent. Thromb. Haemost. 2006, 96, 822–829. [Google Scholar] [CrossRef]
- Igarashi, M.; Takeda, Y.; Mori, S.; Takahashi, K.; Fuse, T. Depolymerized holothurian glycosaminoglycan (DHG) prevents neointimal formation in balloon-injured rat carotid artery. Atherosclerosis 1997, 129, 27–31. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, Y.; Ye, X.; Hu, Y.; Ding, T.; Chen, S. Sulfation pattern of fucose branches affects the anti-hyperlipidemic activities of fucosylated chondroitin sulfate. Carbohydr. Polym. 2016, 147, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, D.; Wang, S.; Tao, L.; Wang, A.; Chen, W.; Zhu, Z.; Zheng, S.; Gao, X.; Lu, Y. Holothurian Glycosaminoglycan Inhibits Metastasis and Thrombosis via Targeting of Nuclear Factor- k B/Tissue Factor/Factor Xa Pathway in Melanoma B16F10 Cells. PLoS ONE 2013, 8, e56557. [Google Scholar] [CrossRef]
- He, M.; Wang, J.; Hu, S.; Wang, Y.; Xue, C.; Li, H. The Effects of Fucosylated Chondroitin Sulfate Isolated from Isostichopus badionotus on Antimetastatic Activity via Down- regulation of Hif-1 α and Hpa. Food Sci. Biotechnol. 2014, 23, 1643–1651. [Google Scholar] [CrossRef]
- Pomin, V.H. Antimicrobial Sulfated Glycans: Structure and Function. Curr. Top. Med. Chem. 2017, 17, 319–330. [Google Scholar] [CrossRef]
- Jiang, W.; Li, S.; Hu, S.; Wang, J.; Song, W.; Ji, L.; Wang, Y. Fucosylated Chondroitin Sulfate from Sea Cucumber Inhibited Islets of Langerhans Apoptosis via Inactivation of the Mitochondrial Pathway in Insulin Resistant Mice. Food Sci. Biotechnol. 2015, 24, 1105–1106. [Google Scholar] [CrossRef]
- Monteiro-Machado, M.; Tomaz, M.A.; Fonseca, R.J.C.; Strauch, M.A.; Cons, B.L.; Borges, P.A.; Patr, F.C.; Tavares-Henriques, M.S.; Teixeira-Cruz, J.M.; Calil-Elias, S.; et al. Occurrence of sulfated fucose branches in fucosylated chondroitin sulfate are essential for the polysaccharide effect preventing muscle damage induced by toxins and crude venom from Bothrops jararacussu snake. Toxicon 2015, 98, 20–33. [Google Scholar] [CrossRef]
- Henrotin, Y.; Mathy, M.; Sanchez, C.; Lambert, C. Chondroitin sulfate in the treatment of osteoarthritis: From in vitro studies to clinical recommendations. Ther. Adv. Musculoskelet. Dis. 2010, 2, 335–348. [Google Scholar] [CrossRef]
- Zeng, C.; Wei, J.; Li, H.; Wang, Y.; Xie, D.; Yang, T. Effectiveness and safety of Glucosamine, chondroitin, the two in combination, or celecoxib in the treatment of osteoarthritis of the knee. Sci. Rep. 2015, 5, 16827. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Sang, L.; Wu, D.; Rong, J.; Jiang, L. Effectiveness and safety of glucosamine and chondroitin for the treatment of osteoarthritis: A meta-analysis of randomized controlled trials. J. Orthop. Surg. Res. 2018, 13, 1–9. [Google Scholar] [CrossRef]
- Roman-Blas, J.A.; Santos, C.; Sanchez-Pernaute, O.; Largo, R.; Herrero-Beaumont, G. CS/GS Combined Therapy Study Group. Combined Treatment with Chondroitin Sulfate and Glucosamine Sulfate Shows No Superiority Over Placebo for Reduction of Joint Pain and Functional Impairment in Patients with Knee Osteoarthritis. Arthritis Rheumatol. 2017, 69, 77–85. [Google Scholar] [CrossRef]
- Jackson, C.G.; Plaas, A.H.; Sandy, J.D.; Hua, C.; Kim-rolands, S.; Barnhill, J.G.; Harris, C.L.; Clegg, D.O. The human pharmacokinetics of oral ingestion of glucosamine and chondroitin sulfate taken separately or in combination. Osteoarthr. Cartil. 2010, 18, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Volpi, N. Chondroitin Sulfate Safety and Quality. Molecules 2019, 24, 1447. [Google Scholar] [CrossRef]
- Nakano, T.; Ozimek, L. Detection of keratan sulfate by immunological methods in commercial chondroitin sulfate preparations. Carbohydr. Polym. 2014, 99, 547–552. [Google Scholar] [CrossRef]
- Caterson, B.; Melrose, L. Keratan Sulphate, a complex Glycosaminoglycan with Unique Functional Capability. Glycobiology 2018, 28, 182–206. [Google Scholar] [CrossRef]
- Volpi, N. Quality of different chondroitin sulfate preparations in relation to their therapeutic activity. J. Pharm. Pharmacol. 2009, 61, 1271–1280. [Google Scholar] [CrossRef]
- Chevalier, X.; Conrozier, T. Access to highly purified chondroitin sulfate for appropriate treatment of osteoarthritis: A review. Med. Access Point Care 2017, 1. [Google Scholar] [CrossRef]
- Morise, J.; Takematsu, H.; Oka, S. The role of human natural killer-1 (HNK-1) carbohydrate in neuronal plasticity and disease. BBA Gen. Subj. 2017, 1861, 2455–2461. [Google Scholar] [CrossRef]
- Ariga, B.T. Review The role of sulfoglucuronosyl glycosphingolipids in the pathogenesis of monoclonal IgM paraproteinemia and peripheral neuropathy. Proc. Jpn. Acad. Ser. B 2011, 87, 386–404. [Google Scholar] [CrossRef]
- Karamanos, N.K.; Aletras, A.J.; Antonopoulos, C.A.; Hjerpe, A. Determination of the HNK-1 epitope (3-sulphated glucuronic acid) in intact chondroitin sulphates by ELISA. Application to squid skin proteoglycans and their oversulphated carbohydrate structures. Biochimie 1994, 4, 79–82. [Google Scholar] [CrossRef]
- Greinacher, A.; Selleng, K.; Warkentin, T.E. Autoimmune heparin-induced thrombocytopenia. J. Thromb. Haemost. 2017, 15, 2099–2114. [Google Scholar] [CrossRef] [Green Version]
- Blank, M.; Cines, D.B.; Arepally, G.; Eldor, A.; Afek, A. Pathogenicity of human anti-platelet factor 4 (PF4)/heparin in vivo: Generation of mouse anti-PF4/heparin and induction of thrombocytopenia by heparin. Clin. Exp. Immunol. 1997, 108, 333–339. [Google Scholar] [CrossRef]
- Petersen, F.; Brandt, E.; Lindahl, U.; Spillmann, D. Characterization of a Neutrophil Cell Surface Glycosaminoglycan That Mediates Binding of Platelet Factor 4. J. Biol. Chem. 1999, 274, 12376–12382. [Google Scholar] [CrossRef] [Green Version]
- Lord, M.S.; Cheng, B.; Farrugia, B.L.; Mccarthy, S.; Whitelock, J.M. Platelet Factor 4 Binds to Vascular Proteoglycans and Controls Both Growth Factor Activities and Platelet Activation. J. Biol. Chem. 2017, 292, 4054–4063. [Google Scholar] [CrossRef] [Green Version]
- Ramacciotti, E.; Clark, M.; Sadeghi, N.; Hoppensteadt, D.; Thethi, I. Contaminants in Heparin: Review of the Literature, Molecular Profiling, and Clinical Implications. Clin. Appl. Thromb. Hemost. 2011, 17, 126–135. [Google Scholar] [CrossRef]
- Adiguzel, C.; Bansal, V.; Litinas, E.; Cunanan, J.; Iqbal, O.; Nelson, K.; Kannan, M.; Hoppensteadt, D. Increased Prevalence of Antiheparin Platelet Factor 4 Antibodies in Patients May Be Due to Contaminated Heparin. Clin. Appl. Thromb. Hemost. 2008, 15, 145–151. [Google Scholar] [CrossRef]
- Chess, E.K.; Bairstow, S.; Donovan, S.; Havel, K.; Hu, P.; Johnson, R.J.; Lee, S.; Mckee, J.; Miller, R.; Moore, E.; et al. Case Study: Contamination of Heparin with Oversulfated Chondroitin Sulfate. Handb. Exp. Pharmacol. 2012, 207, 99–125. [Google Scholar] [CrossRef]
- Guerrini, M.; Beccati, D.; Shriver, Z.; Naggi, A.; Viswanathan, K.; Bisio, A.; Capila, I.; Lansing, J.C.; Guglieri, S.; Fraser, B.; et al. Oversulfated chondroitin sulfate is a contaminant in heparin associated with adverse clinical events. Nat. Biotechnol. 2008, 26, 669–675. [Google Scholar] [CrossRef]
- Kishimoto, T.K.; Viswanathan, K.; Ganguly, T.; Elankumaran, S.; Smith, S.; Pelzer, K. Al-Hakim, A. Contaminated Heparin Associated with Adverse Clinical Events and Activation of the Contact System. N. Engl. J. Med. 2008, 358, 2457–2467. [Google Scholar] [CrossRef]
- Ramacciotti, E.; Wahi, R.; Messmore, H.L. Contaminated Heparin Preparations, Severe Adverse Events and the Contact System. Clin. Appl. Thromb. Hemost. 2008, 14, 489–491. [Google Scholar] [CrossRef]
- Rosania, L. Heparin crisis 2008: A tipping point for increased FDA enforcement in the pharma sector? Food Drug Law J. 2010, 65, 489–501. [Google Scholar]
- Hogwood, J.; Naggi, A.; Torri, G.; Page, C.; Rigsby, P.; Mulloy, B.; Gray, E. The effect of increasing the sulfation level of chondroitin sulfate on anticoagulant specific activity and activation of the kinin system. PLoS ONE 2018, 13, 1–13. [Google Scholar] [CrossRef]
- Van der Meer, J.Y.; Kellenbach, E.; Van den Bos, L.J. From Farm to Pharma: An Overview of Industrial Heparin Manufacturing Methods. Molecules 2017, 22, 1025. [Google Scholar] [CrossRef]
- Pomin, V.H.; Wang, X. Synthetic Oligosaccharide Libraries and Microarray Technology: A Powerful Combination for the Success of Current Glycosaminoglycan Interactomics. ChemMedChem 2017, 13, 648–661. [Google Scholar] [CrossRef] [Green Version]
- Cella, G.; Boeri, G.; Saggiorato, G.; Paolini, R.; Luzzatto, G.; Terribile, V.I. Interaction between Histidine-Rich Glycoprotein and Platelet Factor 4 with Dermatan Sulfate and Low-Molecular-Weight Dermatan Sulfate. Angiology 1992, 43, 59–62. [Google Scholar] [CrossRef]
- Vitale, C.; Berutti, S.; Bagnis, C. Dermatan sulfate: An alternative to unfractionated heparin for anticoagulation in hemodialysis patients. J. Nephrol. 2013, 26, 158–163. [Google Scholar] [CrossRef]
- Greinacher, M.D.A. Clinical Practice. Heparin-Induced Thrombocytopenia. N. Engl. J. Med. 2015, 373, 252–261. [Google Scholar] [CrossRef]
- Pan, J.; Qian, Y.; Zhou, X.; Lu, H.; Ramacciotti, E.; Zhang, L. Chemically Oversulfated Glycosaminoglycans Are Potent Modulators of Contact System Activation and Different Cell Signaling Pathways. J. Biol. Chem. 2010, 285, 22966–22975. [Google Scholar] [CrossRef] [Green Version]
- Sommers, C.D.; Montpas, N.; Adam, A.; Keire, D.A. Characterization of currently marketed heparin products: Adverse event relevant bioassays. J. Pharm. Biomed. Anal. 2012, 67, 28–35. [Google Scholar] [CrossRef]
- Fonseca, R.J.C.; Oliveira, S.M.C.G.; Pomin, V.H.; Araujo, I.G.; Mourão, P.A. Effects of oversulfated and fucosylated chondroitin sulfates on coagulation Challenges for the study of anticoagulant polysaccharides. Thromb. Haemost. 2010, 103, 994–1004. [Google Scholar] [CrossRef]
- Lin, L.; Xu, L.; Xiao, C.; Zhou, L.; Gao, N.; Wu, M.; Zhao, J. Plasma contact activation by a fucosylated chondroitin sulfate and its structure-activity-relationship study. Glycobiology 2018, 28, 754–764. [Google Scholar] [CrossRef]
- Pang, M.; Siddiquee, S.; Al Azad, S. Fucosylated chondroitin sulfate diversity in sea cucumbers: A review. Carbohydr. Polym. 2014, 112, 173–178. [Google Scholar] [CrossRef]
Figure 1.
(A) Representative repeating disaccharide unit of chondroitin sulfate (CS) structures. CSs are composed of alternating 4-linked β-d-glucuronic acid (GlcA) and 3-linked β-d-N-acetylgalactosamine (GalNAc) units. While CS-A is mostly 4-sulfated at the GalNAc units, CS-C is predominantly 6-sulfated [7]. While CS-D is composed of 2-sulfated GlcA and 6-sulfated GalNAc, CS-E is composed of 4,6-di-sulfated GalNAc. CS-K, CS-L and CS-M are all sulfated at C3 position of GlcA, and respectively sulfated at positions C4, C6, and simultaneously at C4 and C6. Oversulfated chondroitin sulfate (OSCS) is fully sulfated. Dermatan sulfate (DS) also known as CS-B is composed of alternating 4-linked α-L-iduronic acid (IdoA) and 3-linked β-d-GalNAc units. (B) Representative repeating trisaccharide unit of the fucosylated chondroitin sulfate (FucCS). Besides consisting of the regular CS disaccharide unit as shown at panel A, the FucCS also exhibits branching units of α-l-fucopyranose (Fucp) 3-linked to the GlcA unit as highlighted with the box in dashed lines. The detailed structure of the Fucp unit varies according to the holothurian species. In all panels the glycosidic bonds are indicated in ellipses whereas monosaccharide types are indicated in continuous black rectangles. For fast notation of the sulfation patterns, sulfation sites are highlighted by light-grey rectangles.
Figure 1.
(A) Representative repeating disaccharide unit of chondroitin sulfate (CS) structures. CSs are composed of alternating 4-linked β-d-glucuronic acid (GlcA) and 3-linked β-d-N-acetylgalactosamine (GalNAc) units. While CS-A is mostly 4-sulfated at the GalNAc units, CS-C is predominantly 6-sulfated [7]. While CS-D is composed of 2-sulfated GlcA and 6-sulfated GalNAc, CS-E is composed of 4,6-di-sulfated GalNAc. CS-K, CS-L and CS-M are all sulfated at C3 position of GlcA, and respectively sulfated at positions C4, C6, and simultaneously at C4 and C6. Oversulfated chondroitin sulfate (OSCS) is fully sulfated. Dermatan sulfate (DS) also known as CS-B is composed of alternating 4-linked α-L-iduronic acid (IdoA) and 3-linked β-d-GalNAc units. (B) Representative repeating trisaccharide unit of the fucosylated chondroitin sulfate (FucCS). Besides consisting of the regular CS disaccharide unit as shown at panel A, the FucCS also exhibits branching units of α-l-fucopyranose (Fucp) 3-linked to the GlcA unit as highlighted with the box in dashed lines. The detailed structure of the Fucp unit varies according to the holothurian species. In all panels the glycosidic bonds are indicated in ellipses whereas monosaccharide types are indicated in continuous black rectangles. For fast notation of the sulfation patterns, sulfation sites are highlighted by light-grey rectangles.

Figure 2.
Representative disaccharide unit, GlcA(β1−3)GalNAc, of CS backbones in the 3D stick model. Hydrogens, carbons, oxygens and nitrogens are respectively represented in light grey, dark grey, red and blue. Carbon atoms are numbered according to their positions in the ring. Monosaccharides are indicated accordingly. Model created by the online AMBER/GLYCAM-based carbohydrate builder [40].
Figure 2.
Representative disaccharide unit, GlcA(β1−3)GalNAc, of CS backbones in the 3D stick model. Hydrogens, carbons, oxygens and nitrogens are respectively represented in light grey, dark grey, red and blue. Carbon atoms are numbered according to their positions in the ring. Monosaccharides are indicated accordingly. Model created by the online AMBER/GLYCAM-based carbohydrate builder [40].

Figure 3.
Representative disaccharide units of CS types, (A) CS-A, (B) CS-C, (C) CS-D, (D) CS-E, and (E) CS-K in their 3D stick models. Table 1 displays the structural description of these disaccharides. Hydrogens, carbons, oxygens and nitrogens are respectively represented in light grey, dark grey, red and blue. The carbon numbering and monosaccharide units are the same as shown in Figure 2, since all structures are the GlcA(β1−3)GalNAc disaccharide backbone. Models created by the online AMBER/GLYCAM-based carbohydrate builder [40].
Figure 3.
Representative disaccharide units of CS types, (A) CS-A, (B) CS-C, (C) CS-D, (D) CS-E, and (E) CS-K in their 3D stick models. Table 1 displays the structural description of these disaccharides. Hydrogens, carbons, oxygens and nitrogens are respectively represented in light grey, dark grey, red and blue. The carbon numbering and monosaccharide units are the same as shown in Figure 2, since all structures are the GlcA(β1−3)GalNAc disaccharide backbone. Models created by the online AMBER/GLYCAM-based carbohydrate builder [40].

Figure 4.
Representative disaccharide unit, IdoA(α1−3)GalNAc, of DS backbones in the 3D stick model. Hydrogens, carbons, oxygens and nitrogens are respectively represented in light grey, dark grey, red and blue. Carbon atoms are numbered according to their positions in the ring. Monosaccharides are indicated accordingly. Model created by the online AMBER/GLYCAM-based carbohydrate builder [40].
Figure 4.
Representative disaccharide unit, IdoA(α1−3)GalNAc, of DS backbones in the 3D stick model. Hydrogens, carbons, oxygens and nitrogens are respectively represented in light grey, dark grey, red and blue. Carbon atoms are numbered according to their positions in the ring. Monosaccharides are indicated accordingly. Model created by the online AMBER/GLYCAM-based carbohydrate builder [40].

Figure 5.
Representative disaccharide units, (A) IdoA2S(α1−3)GalNAc4S and (B) IdoA2S(α1−3)GalNAc6S, of the DS backbones from three ascidian species in their 3D stick models. These ascidian DS are highly sulfated at the C2 position of the α-L-IdoA units but differ in the pattern of sulfation at the β-d-GalNAc units. (A) In the species S. plicata and H. pyriformis the GalNAc moieties are heavily 4-O-sulfated, (B) whereas in A. nigra, they are 6-O-sulfated. Percentage of sulfation/site/species is shown in the table. Hydrogens, carbons, oxygens and nitrogens are respectively represented in light grey, dark grey, red and blue. Carbon atoms are numbered according to their positions in the ring. Monosaccharides are indicated accordingly. Model created by the online AMBER/GLYCAM-based carbohydrate builder [40].
Figure 5.
Representative disaccharide units, (A) IdoA2S(α1−3)GalNAc4S and (B) IdoA2S(α1−3)GalNAc6S, of the DS backbones from three ascidian species in their 3D stick models. These ascidian DS are highly sulfated at the C2 position of the α-L-IdoA units but differ in the pattern of sulfation at the β-d-GalNAc units. (A) In the species S. plicata and H. pyriformis the GalNAc moieties are heavily 4-O-sulfated, (B) whereas in A. nigra, they are 6-O-sulfated. Percentage of sulfation/site/species is shown in the table. Hydrogens, carbons, oxygens and nitrogens are respectively represented in light grey, dark grey, red and blue. Carbon atoms are numbered according to their positions in the ring. Monosaccharides are indicated accordingly. Model created by the online AMBER/GLYCAM-based carbohydrate builder [40].

Figure 6.
Representative trisaccharide unit {→4)-[α-L-Fucp-(1→3)]-β-d-GlcA-(1→3)-β-d-GalNAc-(1→} of the FucCS backbone from sea cucumber in its 3D stick models. The sites of possible substitution by sulfate groups are C2, C3 and C4 of Fucp unit and C4 and C6 of GalNAc as indicated in the structure. Hydrogens, carbons, oxygens and nitrogens are represented in light grey, dark grey, red and blue, respectively. Carbon atoms are numbered according to their positions in the ring. Monosaccharides are indicated accordingly. Model created by the online AMBER/GLYCAM-based carbohydrate builder [40].
Figure 6.
Representative trisaccharide unit {→4)-[α-L-Fucp-(1→3)]-β-d-GlcA-(1→3)-β-d-GalNAc-(1→} of the FucCS backbone from sea cucumber in its 3D stick models. The sites of possible substitution by sulfate groups are C2, C3 and C4 of Fucp unit and C4 and C6 of GalNAc as indicated in the structure. Hydrogens, carbons, oxygens and nitrogens are represented in light grey, dark grey, red and blue, respectively. Carbon atoms are numbered according to their positions in the ring. Monosaccharides are indicated accordingly. Model created by the online AMBER/GLYCAM-based carbohydrate builder [40].


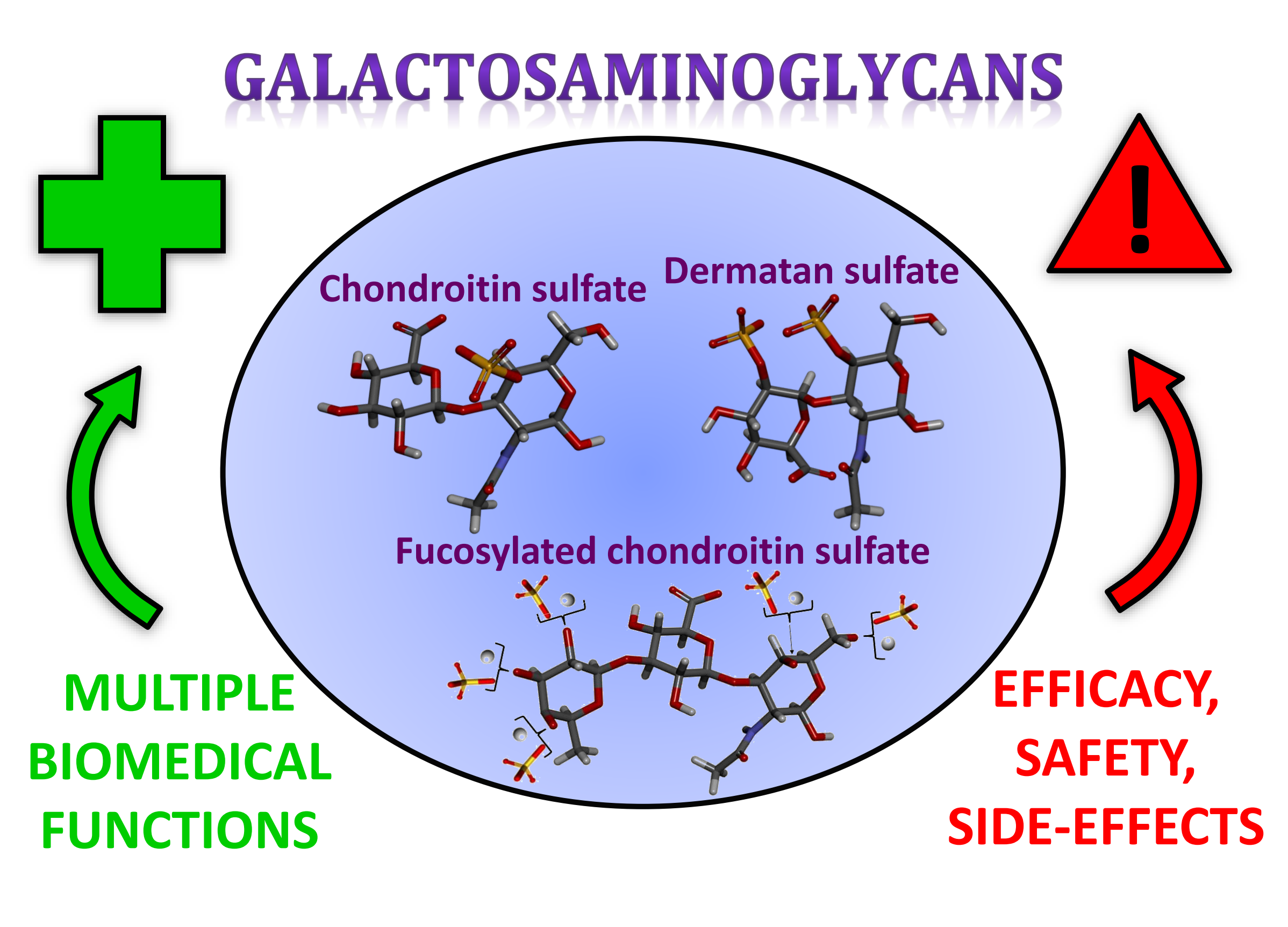{kind=link}
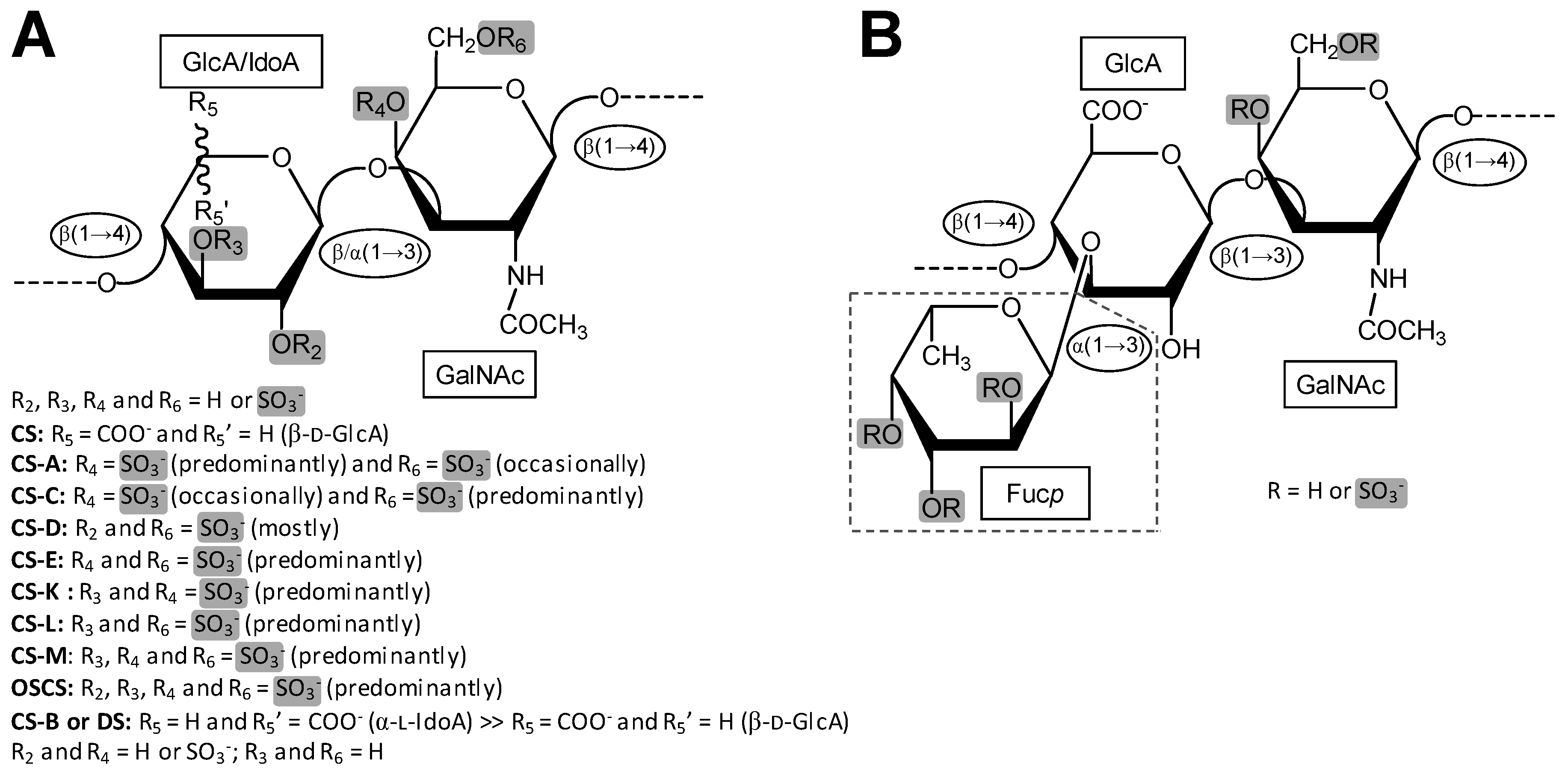{kind=link}
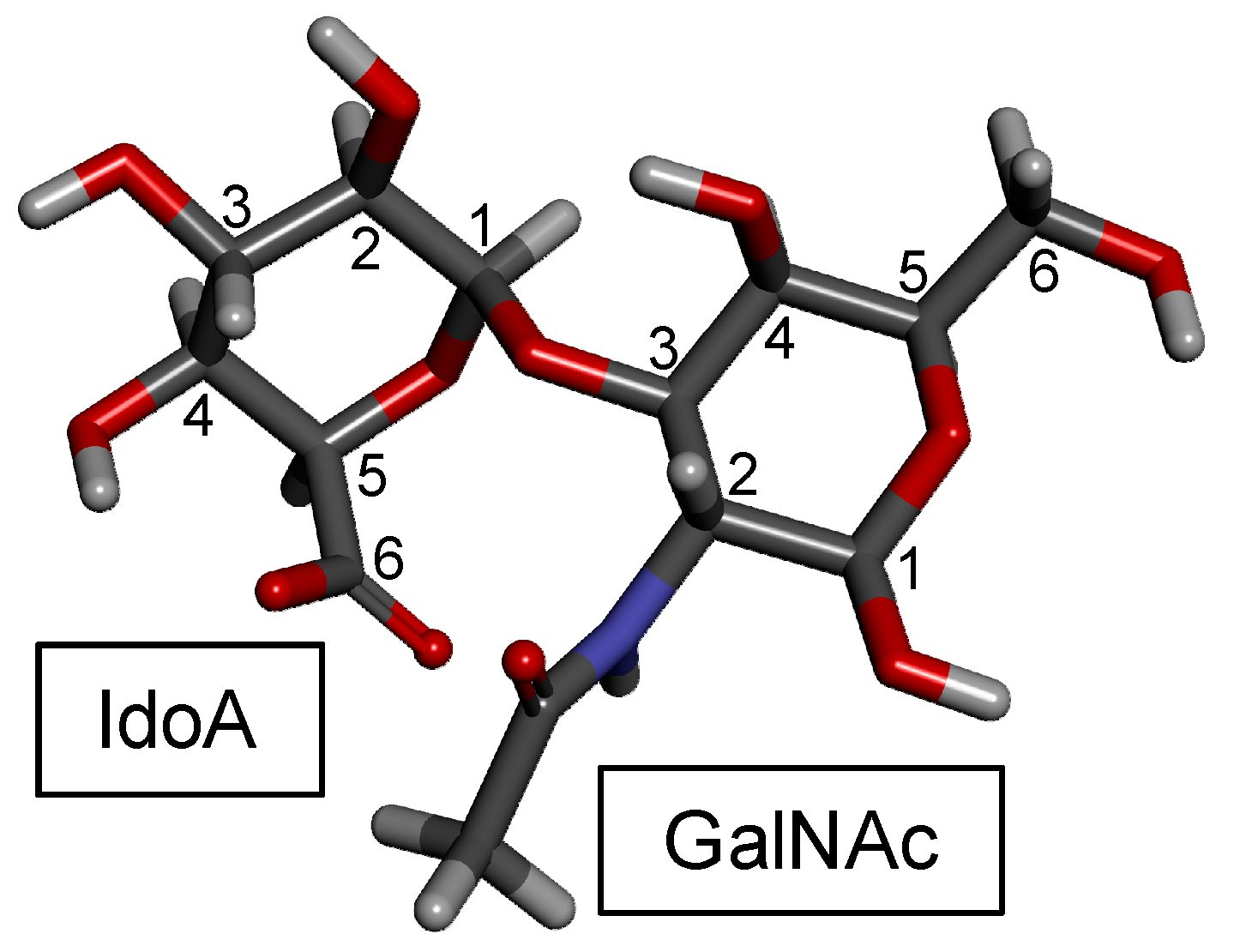{kind=link}
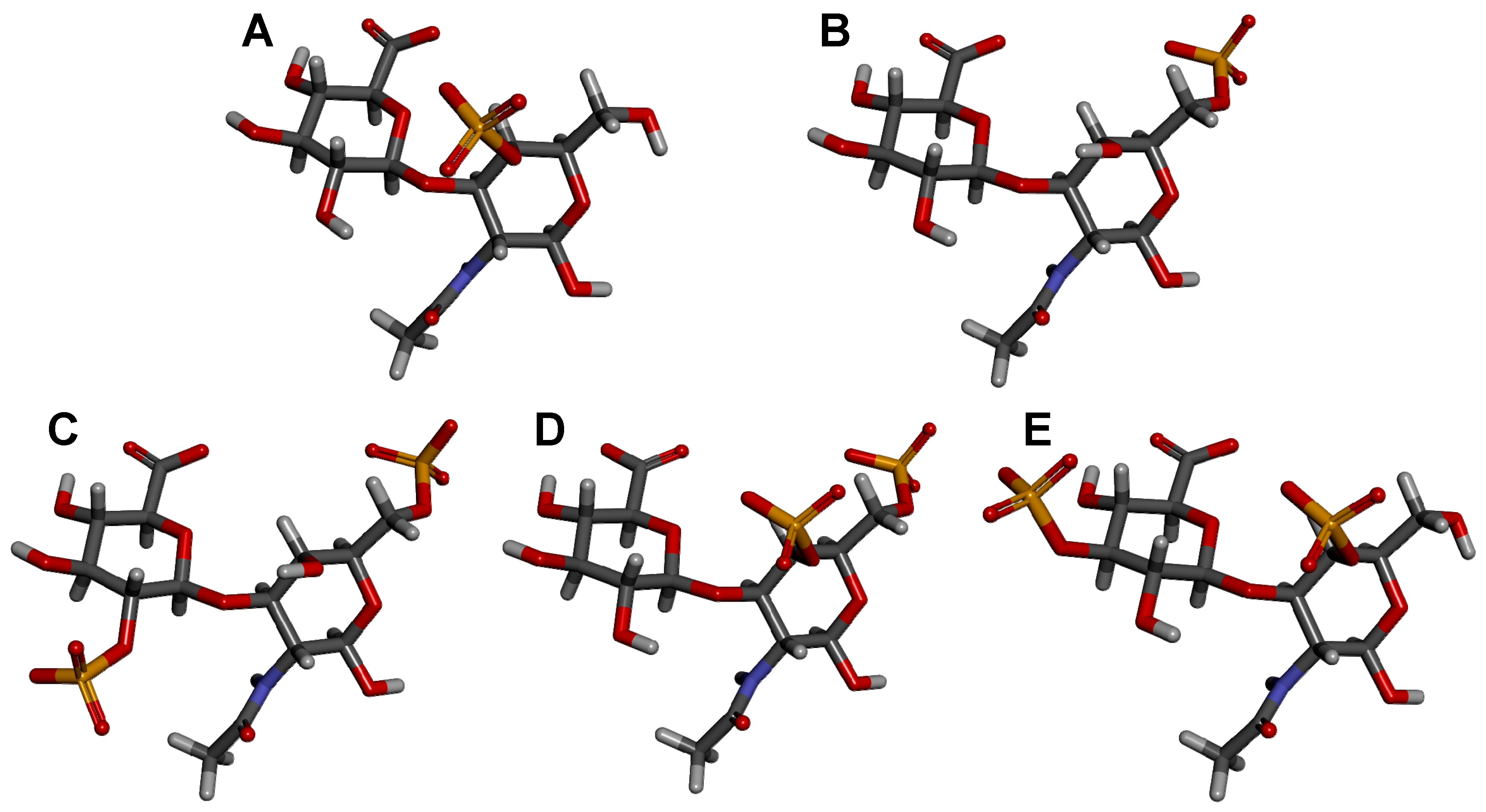{kind=link}
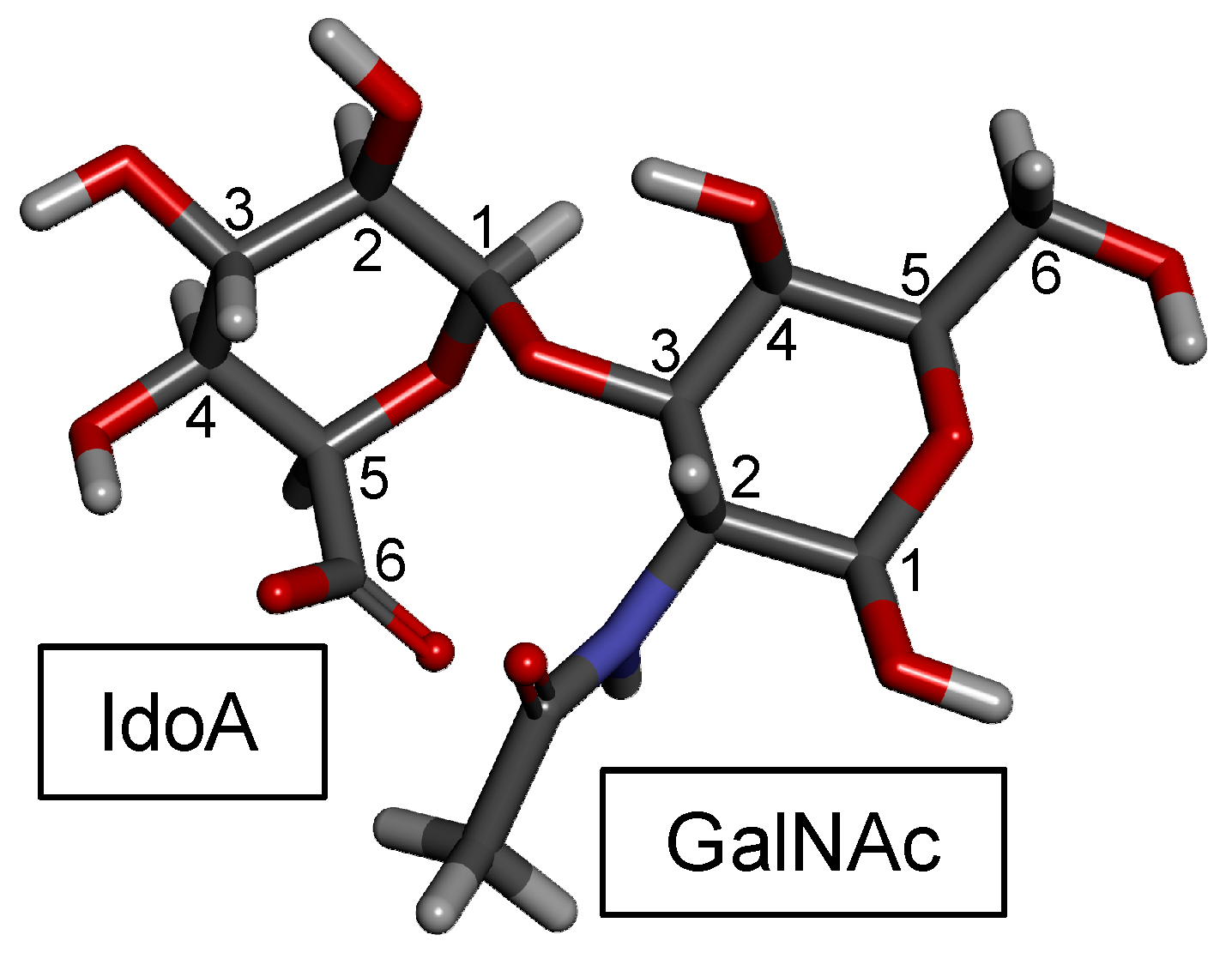{kind=link}
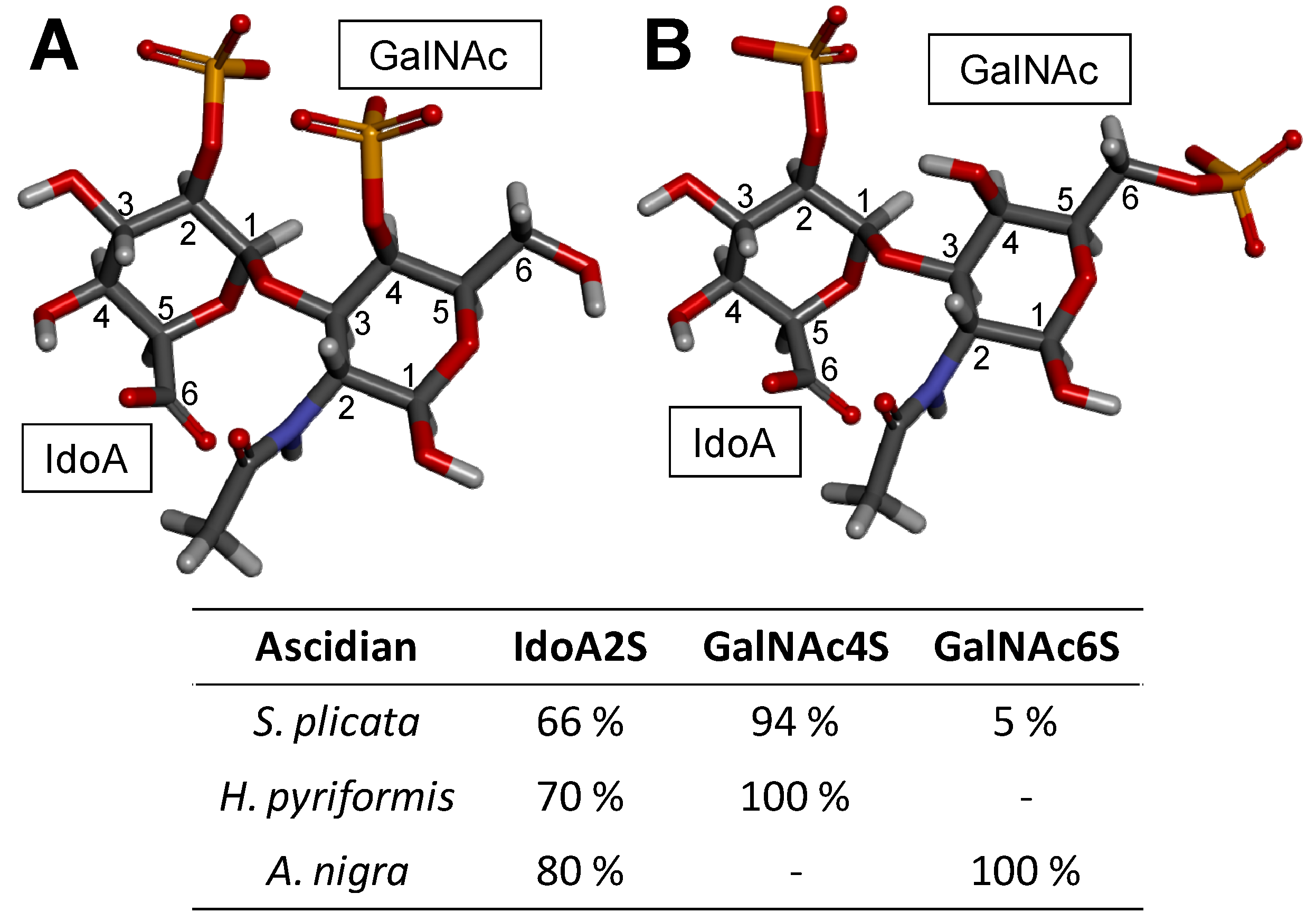{kind=link}
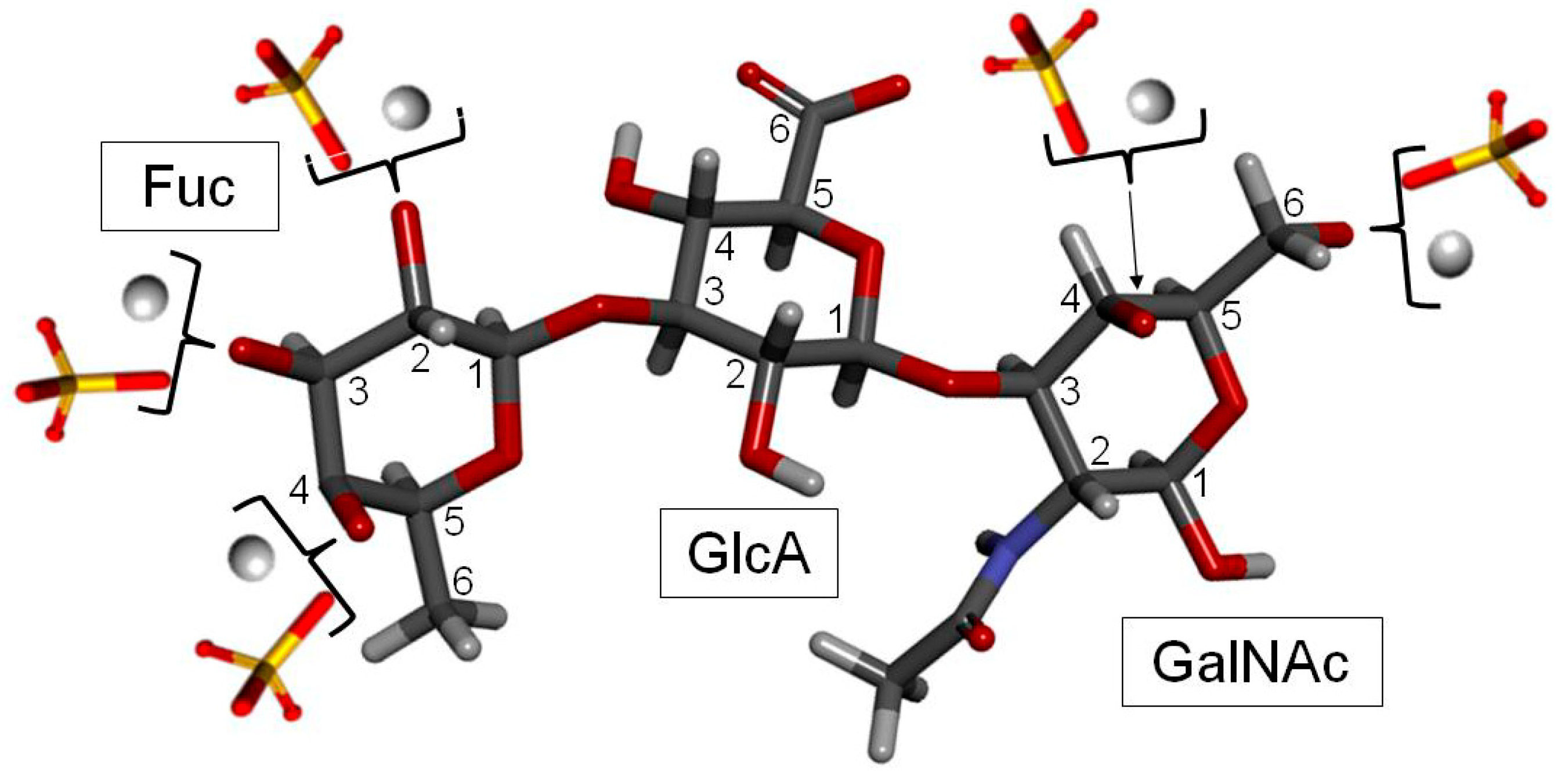{kind=link}
Table 1.
Common and novel CS disaccharide units described by sulfation patterns and by common marine sources.
Table 1.
Common and novel CS disaccharide units described by sulfation patterns and by common marine sources.
| Type | Structure | Source | Reference |
|---|---|---|---|
| CS-A | GlcA(β1−3)GalNAc(4OSO3−) | Whale | [58] |
| CS-C | GlcA(β1−3)GalNAc(6OSO3−) | Shark | [59] |
| CS-D | GlcA(2OSO3−)(β1−3)GalNAc(6OSO3−) | Fish, Shark, Squid | [47,52] |
| CS-E | GlcA(β1−3)GalNAc(4,6diOSO3−) | Squid | [46,60] |
| CS-K | GlcA(3OSO3−)(β1−3)GalNAc(4OSO3−) | King Crab and Octopus | [46,61,62] |
| CS-L | GlcA(3OSO3−)(β1−3)GalNAc(6OSO3−) | Squid | [46] |
| CS-M | GlcA(3OSO3−)(β1−3)GalNAc(4,6diOSO3−) | Squid | [46] |
Table 2.
Common and novel DS (CS-B) disaccharide units described by sulfation patterns and by common sources.
Table 2.
Common and novel DS (CS-B) disaccharide units described by sulfation patterns and by common sources.
| Type | Structure | Source | Reference |
|---|---|---|---|
| DS (4S) | IdoA(α1−3)GalNAc(4OSO3−) | Bovine and porcine mucosa | [108] |
| DS (2S4S) | IdoA(2OSO3−)(α1−3)GalNAc(4OSO3−) | Tunic of ascidian | [104,109] |
| DS (6S) | IdoA(α1−3)GalNAc(6OSO3−) | Bovine liver, bovine spleen, rabbit liver and hog spleen | [58] |
| DS (2S6S) | IdoA(2OSO3−)(α1−3)GalNAc(6OSO3−) | Tunic of ascidian | [105] |
| DS (2S4,6S) (CS-H) | IdoA(2OSO3−)(α1−3)GalNAc(4,6diOSO3−) | Hagfish skin and Hagfish notochord | [72,110,111,112] |
Table 3.
Sulfation patterns (proportions of the branching sulfated fucose units) of FucCS from 27 sea cucumber species analyzed so far.
Table 3.
Sulfation patterns (proportions of the branching sulfated fucose units) of FucCS from 27 sea cucumber species analyzed so far.
| Species | Fuc0S | Fuc3S | Fuc4S | Fuc2S4S | Fuc3S4S | References |
|---|---|---|---|---|---|---|
| Ludwigothurea griseaa | 0 | − | ~49 | ~20 | ~17 | [19,165] |
| Pearsonothuria graeffei | − | − | 81.6 | 18.4 | − | [166] |
| Holothuriava gabunda | 25.6 | − | 50.2 | 15.8 | 8.4 | [166] |
| Stichopus tremulus | − | − | 24.8 | 22.4 | 52.8 | [166] |
| Isostichopus badionotus | − | − | 4.1 | 95.9 | − | [166] |
| Thelenota ananas | 0 | ~25 | ~22 | ~53 | 0 | [167,168] |
| Stichopus japonicusb | 18 | 17 c | 0 | 16 | 23 | [169] |
| Holothuria edulisd | − | − | Nd | 18 | Nd | [170] |
| Apostichopus japonicusd | − | − | Nd | 45 | Nd | [170] |
| Holothuria nobilise | − | Nd | Nd | − | Nd | [170] |
| Acaudina molpadioideaf | − | − | − | − | − | [171] |
| Athyonidium chilensisf | − | − | − | − | − | [172] |
| Patalus mollis | 0 | 0 | 26 | 34 | 40 | [173] |
| Massinium magnumg | _ | _ | _ | _ | Nd | [174] |
| Apostichopus mauritania | _ | _ | _ | 16.7 | 66.7 | [175] |
| Holothuria mexicana | 6.24 | 5.58 | 37.16 | _ | 51 | [176] |
| Cucumaria djakonovi | _ | _ | ~25 | ~50 | ~25 | [177] |
| Eupentaca fraudatrixh | 33.3 | _ | _ | _ | 66.7 | [178] |
| Stichopus horrensi | _ | _ | _ | Nd | _ | [179] |
| Stichopus chlorontusi | _ | _ | _ | Nd | _ | [179] |
| Holothuria scabraj | _ | _ | 27.1 | 72.9 | _ | [180] |
| Holothuria tubulosak | _ | _ | 14.3 | 42.85 | 42.85 | [181] |
| Holothuria stellatik | _ | _ | 20 | 40 | 40 | [181] |
| Holothuria forskai | _ | _ | 15 | 39 | 46 | [182] |
| Cucumaria frondosal | _ | _ | _ | 25 | 62.5 | [183] |
| Cucumaria japonica | _ | _ | _ | 20 | 80 | [184] |
| Actinopyga mauritiana | _ | _ | _ | 20 | 80 | [175] |
a The CS backbone of FucCS from L. grisea has been extensively characterized. It is composed of GalNAc units with the following substitution percentages: 12% 4,6-di-sulfated, 53% 6-mono-sulfated, 4% 4-mono-sulfated, and 31% non-sulfated [165]. Some 3-O-sulfation of GlcA has also been identified [163]. b The CS backbone of this FucCS was mostly characterized as CS-E [155], which is predominantly composed of 4,6-O-di-sulfated GalNAc units. c Not determined. d Although the mono-4S and di-3S4S fucosyl units have been assigned in the FucCS of H. edulis and A. japonicus in [170], the amounts of these units were not provided therein. e The FucCS from H. nobilis was studied by NMR but the anomeric signals belonging to the fucose residues were rather too weak and broad to allow integration and further quantitation of their proportions. However, mono-3S, mono-4S, and di-3S4Sfucosyl units were clearly observed [170]. f Structures studied by Fourier transform–infrared spectroscopy. Just a few structural features were raised. The sulfation patterns of FucCS from these two holothurian species are still an unknown. g The CS backbone was characterized and found to be mainly composed of CS-E and CS-A at a ratio of 9:1, respectively [174]. h The CS backbone of FucCS from E. fraudatrix was characterized to be composed of a 1:1 ratio of CS-E to CS-A [178]. i Both FucCS from S. horrens and S. chlorontus were characterized with a backbone mostly composed of CS-E [179]. j FucCS from H. scabra was characterized with a backbone comprised mostly of CS-C with some CS-E and CS-A units [180]. k The CS backbone of FucCS from H. tubulosa was found to be composed of units of CS-E and CS-A at a ratio of 4:1 while the FucCS backbone from H. stellati is also composed of CS-E and CS-A in a ratio of 4.5:1 [181]. l The FucCS characterized from C. frondosa was found to have an unusual sulfated alpha-L-Fucp linked to the O-6 of the GalNac residue, resulting in 12.2% of the sulfation pattern in addition to the sulfated fucose residues indicated in the table above [183]. Table modified with permission from [6].
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pomin, V.H.; Vignovich, W.P.; Gonzales, A.V.; Vasconcelos, A.A.; Mulloy, B. Galactosaminoglycans: Medical Applications and Drawbacks. Molecules 2019, 24, 2803. https://doi.org/10.3390/molecules24152803
AMA Style
Pomin VH, Vignovich WP, Gonzales AV, Vasconcelos AA, Mulloy B. Galactosaminoglycans: Medical Applications and Drawbacks. Molecules. 2019; 24(15):2803. https://doi.org/10.3390/molecules24152803
Chicago/Turabian StylePomin, Vitor H., William P. Vignovich, Alysia V. Gonzales, Ariana A. Vasconcelos, and Barbara Mulloy. 2019. "Galactosaminoglycans: Medical Applications and Drawbacks" Molecules 24, no. 15: 2803. https://doi.org/10.3390/molecules24152803