
Polysaccharides from Basidiocarps of Cultivating Mushroom Pleurotus ostreatus: Isolation and Structural Characterization


 , ,
, , 

Abstract
1. Introduction
2. Results and Discussion
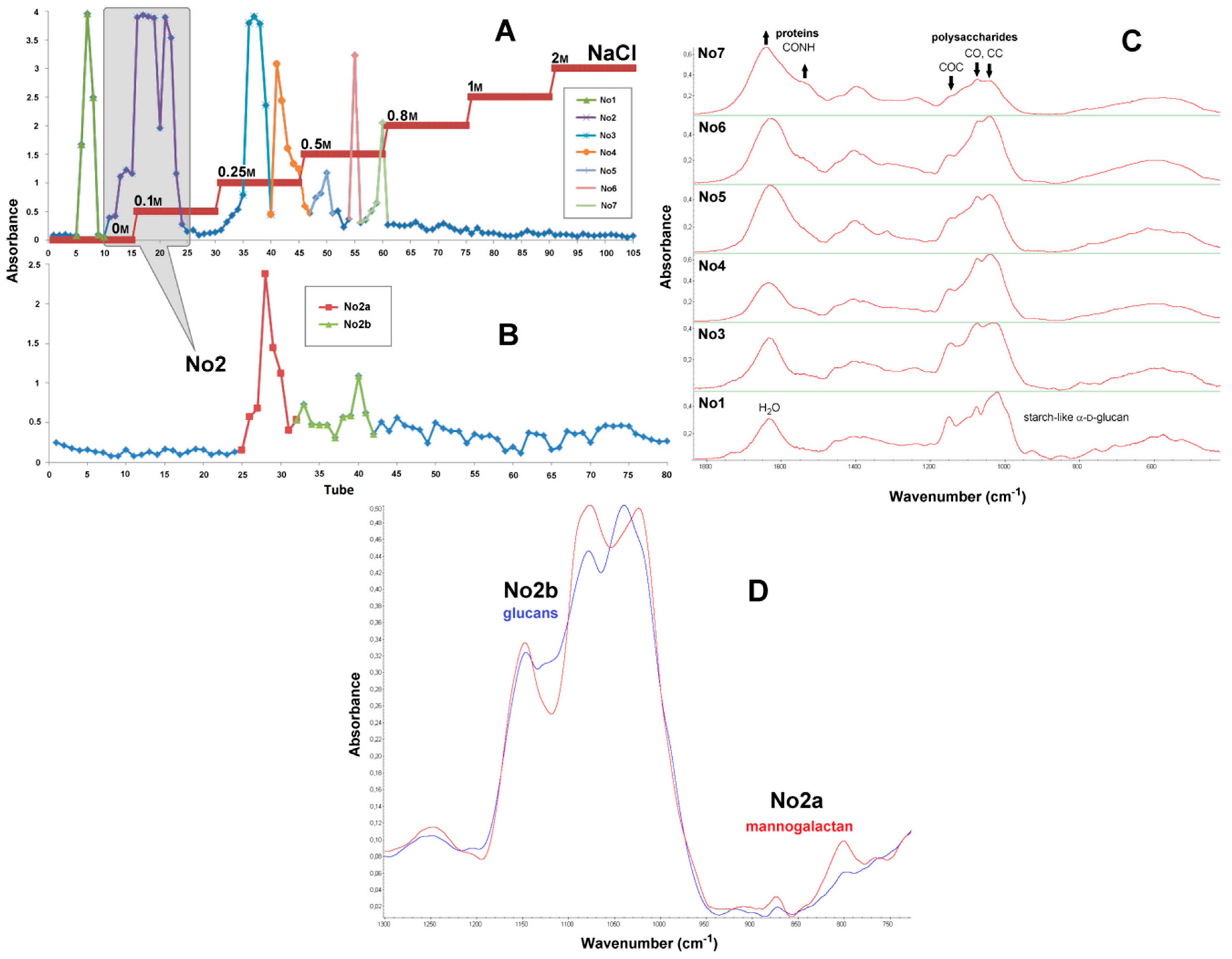
2.1. Yields of Isolations
2.2. Organic Elemental Composition
2.3. Glucan Assay
2.4. Monosaccharide Composition
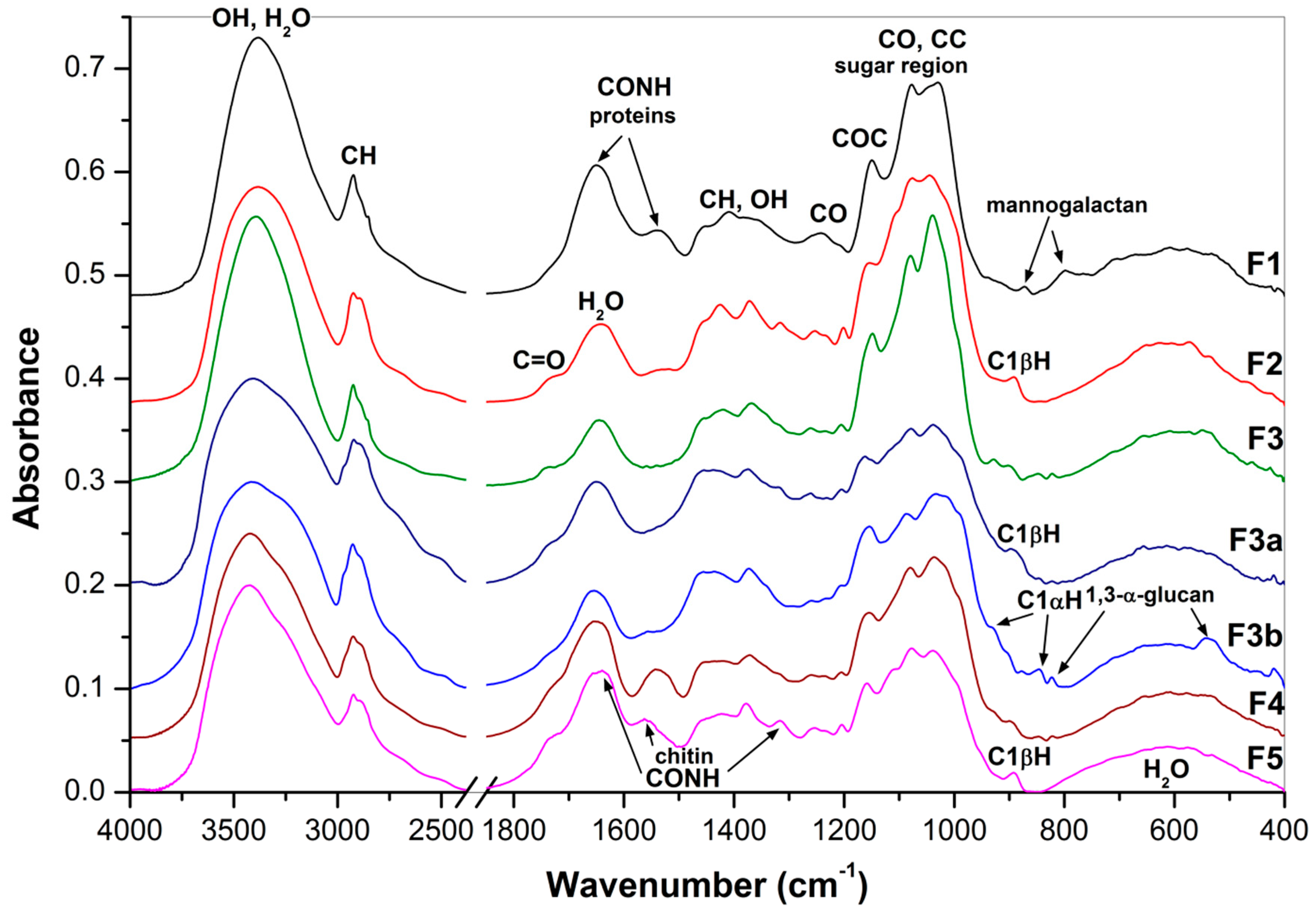
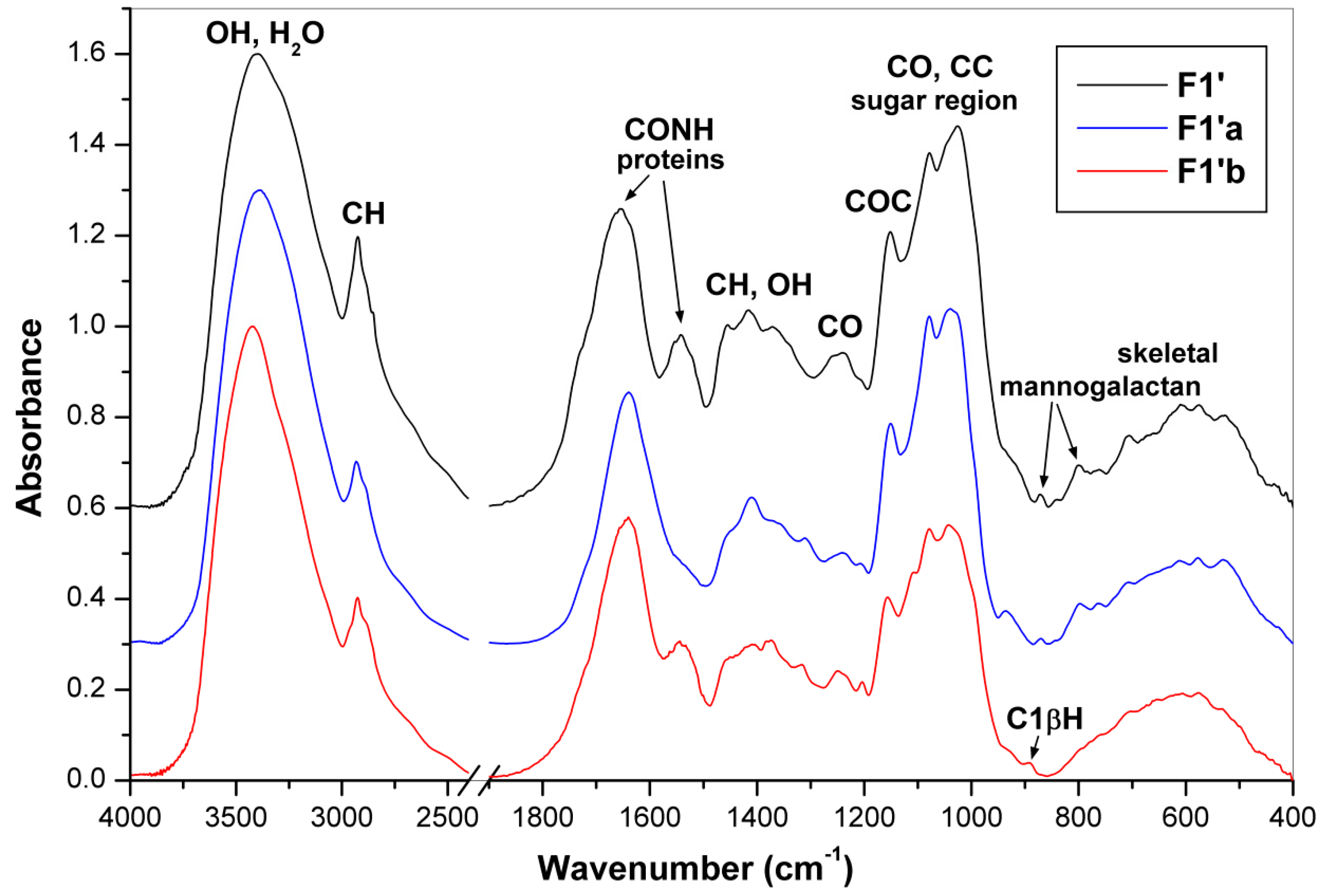
2.5. FTIR Spectra
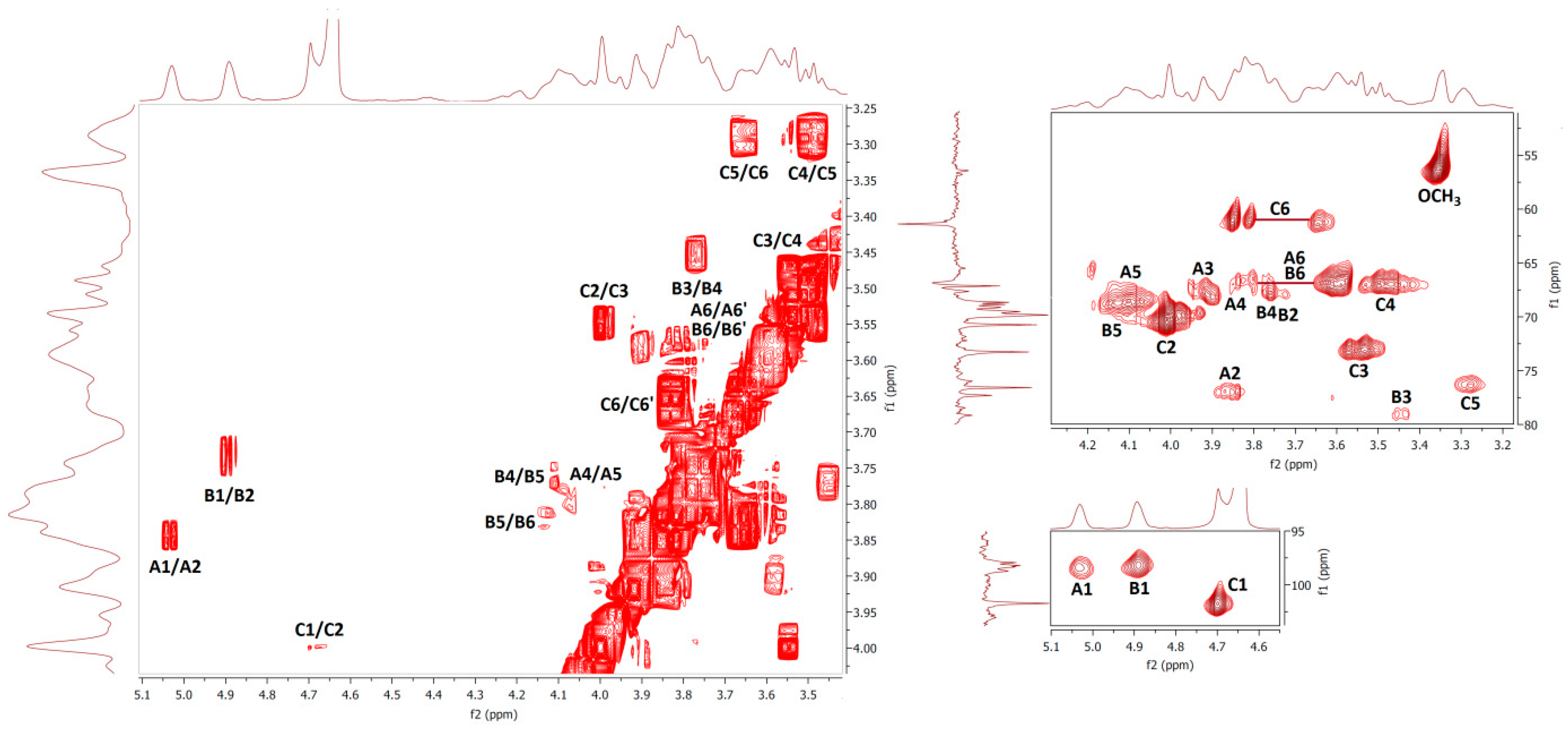
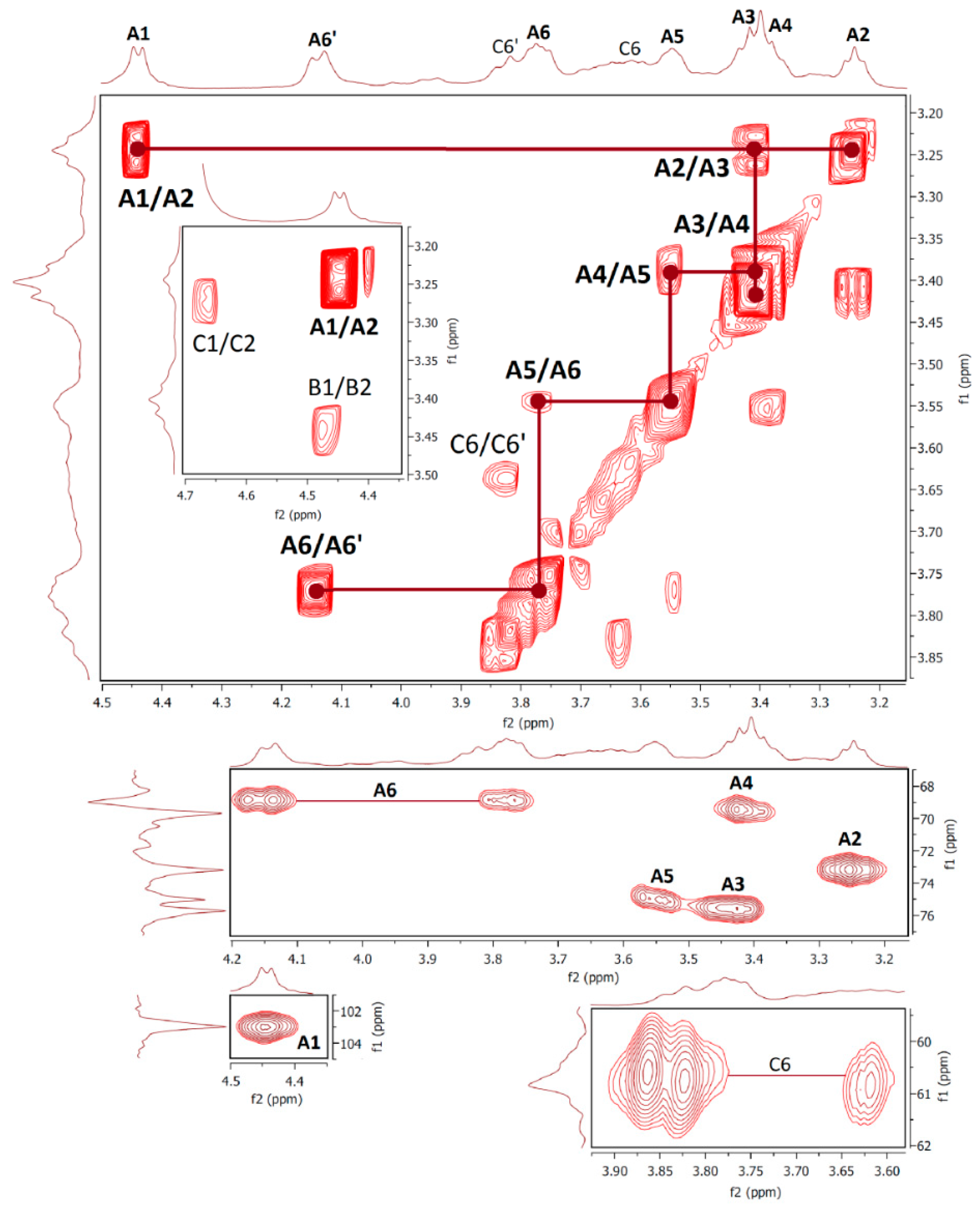
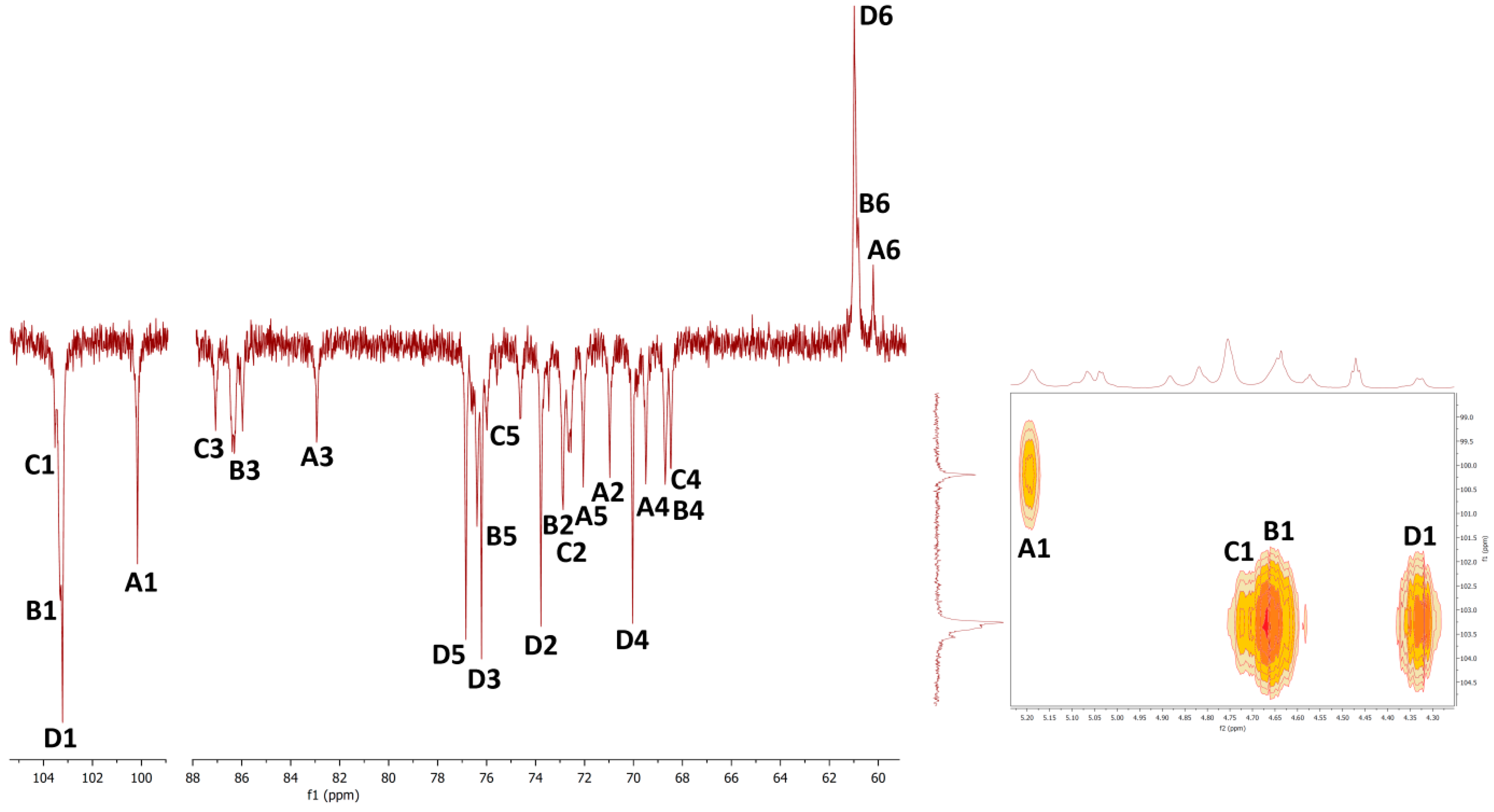
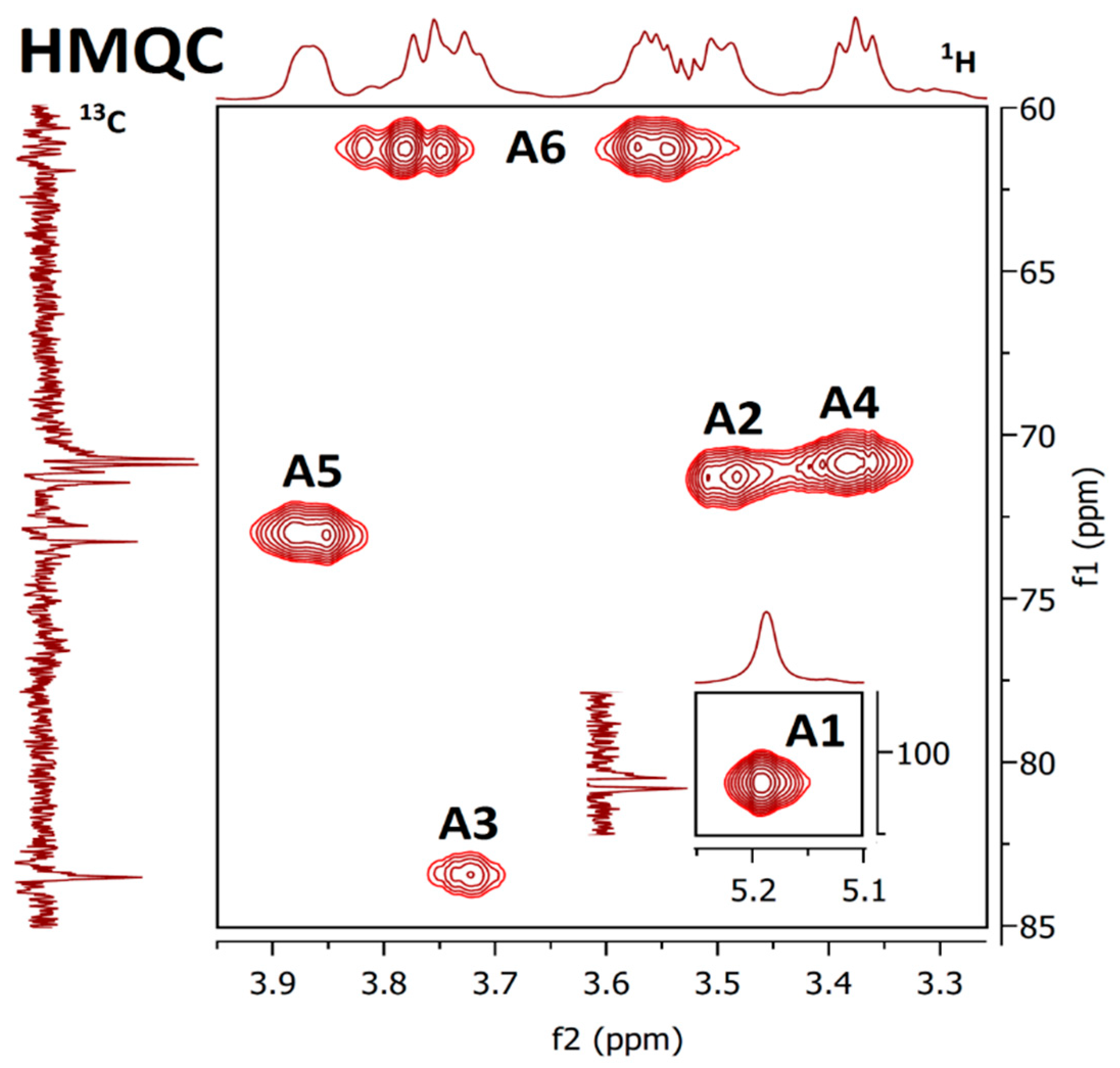
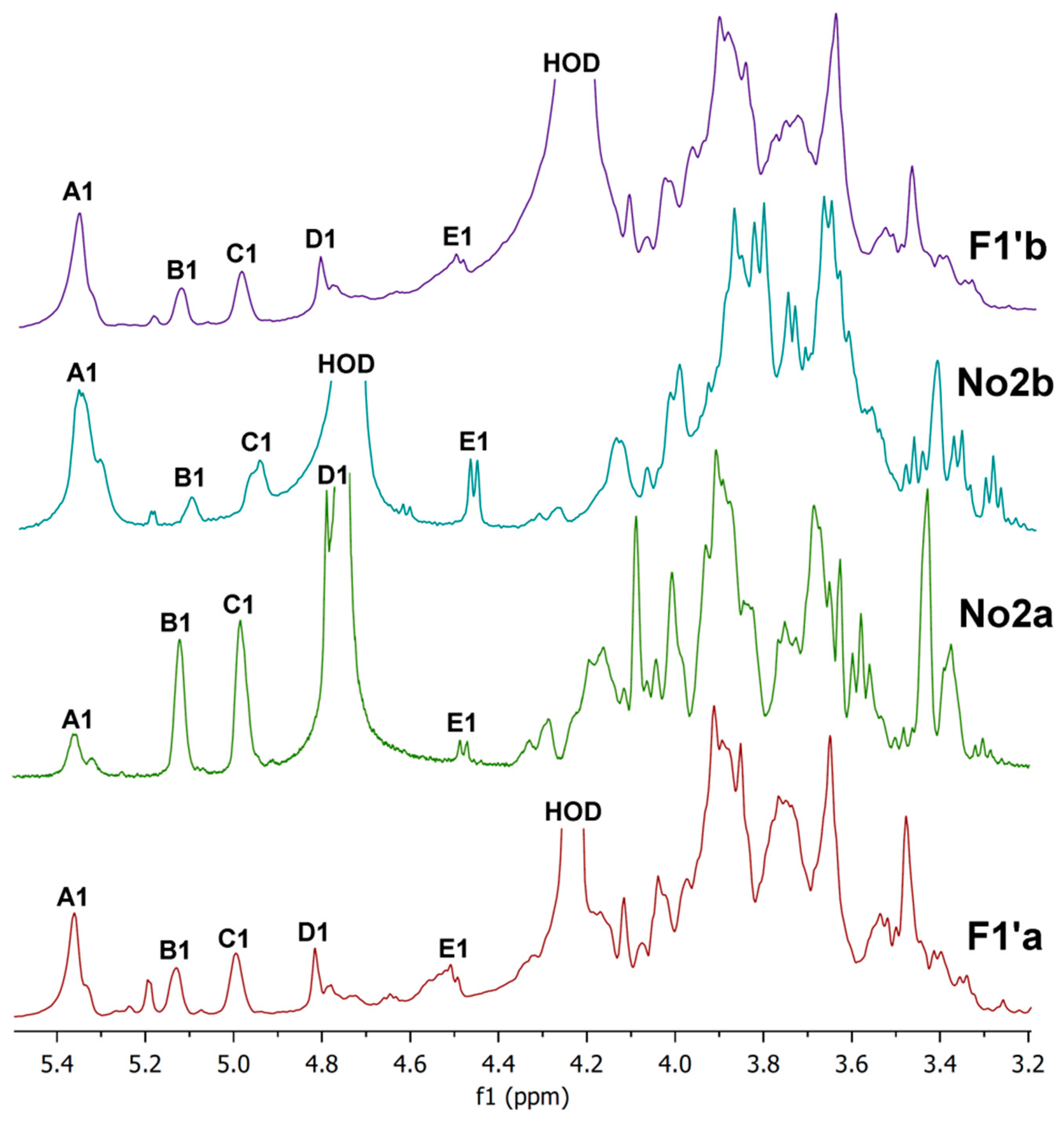
2.6. NMR Spectra
2.7. Preparative Chromatography
3. Materials and Methods
3.1. Materials
3.2. Isolation and Purification Procedures
3.3. Glucan Assay
3.4. Organic Elemental Analysis
3.5. Monosaccharide Composition Analysis
3.6. Spectroscopic Methods
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [PubMed]
- Karácsonyi, Š.; Kuniak, Ľ. Polysaccharides of Pleurotus ostreatus: Isolation and structure of pleuran, an alkali-insoluble β-D-glucan. Carbohydr. Polym. 1994, 24, 107–111. [Google Scholar] [CrossRef]
- Bobek, P.; Ozdín, Ĺ.; Kuniak, Ĺ. Effect of oyster mushroom and isolated β-glucan on lipid peroxidation and on the activities of antioxidative enzymes in rats fed the cholesterol diet. J. Nutr. Biochem. 1997, 8, 469–471. [Google Scholar] [CrossRef]
- Jesenak, M.; Majtan, J.; Rennerova, Z.; Kyselovic, J.; Banovcin, P.; Hrubisko, M. Immunomodulatory effect of pleuran (β-glucan from Pleurotus ostreatus) in children with recurrent respiratory tract infections. Int. Immunopharmacol. 2013, 15, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J. Pleuran (β-glucan from Pleurotus ostreatus): An effective nutritional supplement against upper respiratory tract infections? Med. Sport Sci. 2012, 59, 57–61. [Google Scholar] [PubMed]
- Bergendiova, K.; Tibenska, E.; Majtan, J. Pleuran (β-glucan from Pleurotus ostreatus) supplementation, cellular immune response and respiratory tract infections in athletes. Eur. J. Appl. Physiol. 2011, 111, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonský, I.; Spěváček, J.; Erban, V.; Kováříková, E.; Čopíková, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Akyuz, M.; Kirbag, S. Antimicrobial activity of Pleurotus eryngii var. ferulae grown on various agro-wastes. Eur. Asian J. BioSci. 2009, 3, 58–63. [Google Scholar]
- Dey, B.; Bhunia, S.K.; Maity, K.K.; Patra, S.; Mandal, S.; Maiti, S.; Maiti, T.K.; Sikdar, S.R.; Islam, S.S. Glucans of Pleurotus florida blue variant: Isolation, purification, characterization and immunological studies. Int. J. Biol. Macromol. 2012, 50, 591–597. [Google Scholar] [CrossRef]
- Santos-Neves, J.C.; Pereira, M.I.; Carbonero, E.R.; Gracher, A.H.P.; Alquini, G.; Gorin, P.A.; Sasssaki, G.L.; Iacomini, M. A novel branched αβ-glucan isolated from the basidiocarps of the edible mushroom Pleurotus florida. Carbohydr. Polym. 2008, 73, 309–314. [Google Scholar] [CrossRef]
- Carbonero, E.R.; Gracher, A.H.P.; Smiderle, F.R.; Rosado, F.R.; Sassaki, G.L.; Gorin, P.A.; Iacomini, M. A β-glucan from the fruit bodies of edible mushrooms Pleurotus eryngii and Pleurotus ostreatoroseus. Carbohydr. Polym. 2006, 66, 252–257. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Olsen, L.M.; Carbonero, E.R.; Baggio, C.H.; Freitas, C.S.; Marcon, R.; Santos, A.R.S.; Gorin, P.A.; Iacomini, M. Anti-inflammatory and analgesic properties in a rodent model of a (1→3),(1→6)-linked β-glucan isolated from Pleurotus pulmonarius. Eur. J. Pharmacol. 2008, 597, 86–91. [Google Scholar] [CrossRef]
- Lavi, I.; Levinson, D.; Peri, I.; Tekoah, Y.; Hadar, Y.; Schwartz, B. Chemical characterization, antiproliferative and antiadhesive properties of polysaccharides extracted from Pleurotus pulmonarius mycelium and fruiting bodies. Appl. Microbiol. Biotechnol. 2010, 85, 1977–1990. [Google Scholar] [CrossRef]
- Wong, S.M.; Wong, K.K.; Chiu, L.C.M.; Cheung, P.C.K. Non-starch polysaccharides from different developmental stages of Pleurotus tuber-regium inhibited the growth of human acute promyelocytic leukemia HL-60 cells by cell-cycle arrest and/or apoptotic induction. Carbohydr. Polym. 2007, 68, 206–217. [Google Scholar] [CrossRef]
- Carbonero, E.R.; Ruthes, A.C.; Freitas, C.S.; Utrilla, P.; Gálvez, J.; da Silva, E.V.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Chemical and biological properties of a highly branched β-glucan from edible mushroom Pleurotus sajor-caju. Carbohydr. Polym. 2012, 90, 814–819. [Google Scholar] [CrossRef]
- Pramanik, M.; Chakraborty, I.; Mondal, S.; Islam, S.S. Structural analysis of a water-soluble glucan (Fr. I) of an edible mushroom, Pleurotus sajor-caju. Carbohydr. Res. 2007, 342, 2670–2675. [Google Scholar] [CrossRef]
- Silveira, M.L.; Smiderle, F.R.; Moraes, C.P.; Borato, D.G.; Baggio, C.H.; Ruthes, A.C.; Wisbeck, E.; Sassaki, G.L.; Cipriani, T.R.; Furlan, S.A.; et al. Structural characterization and anti-inflammatory activity of a linear β-d-glucan isolated from Pleurotus sajor-caju. Carbohydr. Polym. 2014, 113, 588–596. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Olsen, L.M.; Carbonero, E.R.; Marcon, R.; Baggio, C.H.; Freitas, C.S.; Santos, A.R.S.; Torri, G.; Gorin, P.A.J.; Iacomini, M. A 3-O-methylated mannogalactan from Pleurotus pulmonarius: Structure and antinociceptive effect. Phytochemistry 2008, 69, 2731–2736. [Google Scholar] [CrossRef]
- Zhang, A.Q.; Xu, M.; Fu, L.; Sun, P.L. Structural elucidation of a novel mannogalactan isolated from the fruiting bodies of Pleurotus geesteranus. Carbohydr. Polym. 2013, 92, 236–240. [Google Scholar] [CrossRef]
- Palacios, I.; García-Lafuente, A.; Guillamón, E.; Villares, A. Novel isolation of water-soluble polysaccharides from the fruiting bodies of Pleurotus ostreatus mushrooms. Carbohydr. Res. 2012, 358, 72–77. [Google Scholar] [CrossRef]
- Corrêa, R.C.G.; Brugnari, T.; Bracht, A.; Peralta, R.M.; Ferreira, I.C. Biotechnological, nutritional and therapeutic uses of Pleurotus spp. (Oyster mushroom) related with its chemical composition: A review on the past decade findings. Trends Food Sci. Technol. 2016, 50, 103–117. [Google Scholar] [CrossRef]
- Patel, Y.; Naraian, R.; Singh, V.K. Medicinal properties of Pleurotus species (oyster mushroom): A review. World J. Fungal Plant Biol. 2012, 3, 1–12. [Google Scholar]
- Snarr, B.; Qureshi, S.; Sheppard, D. Immune recognition of fungal polysaccharides. J. Fungi 2017, 3, 47. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. β-Glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef]
- Bueter, C.L.; Specht, C.A.; Levitz, S.M. Innate sensing of chitin and chitosan. PLoS Pathog. 2013, 9, e1003080. [Google Scholar] [CrossRef]
- Rappleye, C.A.; Eissenberg, L.G.; Goldman, W.E. Histoplasma capsulatum α-(1, 3)-glucan blocks innate immune recognition by the β-glucan receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 1366–1370. [Google Scholar] [CrossRef]
- Gomba, G.K.; Synytsya, A.; Švecová, P.; Coimbra, M.A.; Čopíková, J. Distinction of fungal polysaccharides by N/C ratio and mid infrared spectroscopy. Int. J. Biol. Macromol. 2015, 80, 271–281. [Google Scholar] [CrossRef]
- Synytsya, A.; Míčková, K.; Jablonsky, I.; Sluková, M.; Copikova, J. Mushrooms of genus Pleurotus as a source of dietary fibres and glucans for food supplements. Czech J. Food Sci. 2008, 26, 441–446. [Google Scholar] [CrossRef]
- Rout, D.; Mondal, S.; Chakraborty, I.; Islam, S.S. The structure of a polysaccharide from Fraction-II of an edible mushroom, Pleurotus florida. Carbohydr. Res. 2006, 341, 995–1002. [Google Scholar] [CrossRef]
- Zhang, A.Q.; Zhang, Y.; Yang, J.H.; Sun, P.L. Structural elucidation of a novel heteropolysaccharide from the fruiting bodies of Pleurotus eryngii. Carbohydr. Polym. 2013, 92, 2239–2244. [Google Scholar] [CrossRef]
- Rosado, F.R.; Carbonero, E.R.; Claudino, R.F.; Tischer, C.A.; Kemmelmeier, C.; Iacomini, M. The presence of partially 3-O-methylated mannogalactan from the fruit bodies of edible basidiomycetes Pleurotus ostreatus ‘florida’Berk. and Pleurotus ostreatoroseus Sing. FEMS Microbiol. Lett. 2003, 221, 119–124. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, J. Purification, structure and immunobiological activity of a water-soluble polysaccharide from the fruiting body of Pleurotus ostreatus. Bioresour. Technol. 2009, 100, 983–986. [Google Scholar] [CrossRef]
- Maity, K.K.; Patra, S.; Dey, B.; Bhunia, S.K.; Mandal, S.; Das, D.; Majumdar, D.K.; Maiti, S.; Maiti, T.K.; Islam, S.S. A heteropolysaccharide from aqueous extract of an edible mushroom, Pleurotus ostreatus cultivar: Structural and biological studies. Carbohydr. Res. 2011, 346, 366–372. [Google Scholar] [CrossRef]
- Pramanik, M.; Mondal, S.; Chakraborty, I.; Rout, D.; Islam, S.S. Structural investigation of a polysaccharide (Fr. II) isolated from the aqueous extract of an edible mushroom Pleurotus sajor-caju. Carbohydr. Res. 2005, 340, 629–636. [Google Scholar] [CrossRef]
- Chen, J.N.; Wang, Y.T.; Wu, J.S.B. A glycoprotein extracted from golden oyster mushroom Pleurotus citrinopileatus exhibiting growth inhibitory effect against U937 leukemia cells. J. Agric. Food Chem. 2009, 57, 6706–6711. [Google Scholar] [CrossRef]
- Chen, J.N.; de Mejia, E.G.; Wu, J.S.B. Inhibitory effect of a glycoprotein isolated from golden oyster mushroom (Pleurotus citrinopileatus) on the lipopolysaccharide-induced inflammatory reaction in RAW 264.7 macrophage. J. Agric. Food Chem. 2011, 59, 7092–7097. [Google Scholar] [CrossRef]
- Kong, J.; Yu, S. Fourier transform infrared spectroscopic analysis of protein secondary structures. Acta Biochim. Biophys. Sinica 2007, 39, 549–559. [Google Scholar] [CrossRef]
- Figueiro, S.D.; Góes, J.C.; Moreira, R.A.; Sombra, A.S.B. On the physico-chemical and dielectric properties of glutaraldehyde crosslinked galactomannan–collagen films. Carbohydr. Polym. 2004, 56, 313–320. [Google Scholar] [CrossRef]
- Cerqueira, M.A.; Souza, B.W.; Simões, J.; Teixeira, J.A.; Domingues, M.R.M.; Coimbra, M.A.; Vicente, A.A. Structural and thermal characterization of galactomannans from non-conventional sources. Carbohydr. Polym. 2011, 83, 179–185. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Prieto, A.; Martínez, A.T. Structural characterization of extracellular polysaccharides produced by fungi from the genus Pleurotus. Carbohydr. Res. 1996, 281, 143–154. [Google Scholar] [CrossRef]
- Šandula, J.; Kogan, G.; Kačuráková, M.; Machová, E. Microbial (1→3)-β-d-glucans, their preparation, physico-chemical characterization and immunomodulatory activity. Carbohydr. Res. 1999, 38, 247–253. [Google Scholar] [CrossRef]
- Wang, T.; Deng, L.; Li, S.; Tan, T. Structural characterization of a water insoluble α-(1→3)-d-glucan isolated from the Penicillium chrysogenum. Carbohydr. Polym. 2007, 67, 133–137. [Google Scholar] [CrossRef]
- Das, D.; Mondal, S.; Roy, S.K.; Maiti, D.; Bhunia, B.; Maiti, T.K.; Sikdar, S.R.; Islam, S.S. A (1→6)-β-glucan from a somatic hybrid of Pleurotus florida and Volvariella volvacea: Isolation, characterization, and study of immunoenhancing properties. Carbohydr. Res. 2010, 345, 974–978. [Google Scholar] [CrossRef]
- Cárdenas, G.; Cabrera, G.; Taboada, E.; Miranda, S.P. Chitin characterization by SEM, FTIR, XRD, and 13C cross polarization/mass angle spinning NMR. J. Appl. Polymer Sci. 2004, 93, 1876–1885. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, M.; Dong, J.; Guo, J.; Song, Y.; Cheung, P.C.K. Chemical structure and chain conformation of the water-insoluble glucan isolated from Pleurotus tuber-regium. Biopolym. Orig. Res. Biomol. 2001, 59, 457–464. [Google Scholar] [CrossRef]
- Fan, D.; Ma, W.; Wang, L.; Huang, J.; Zhao, J.; Zhang, H.; Chen, W. Determination of structural changes in microwaved rice starch using Fourier transform infrared and Raman spectroscopy. Starch-Stärke 2012, 64, 598–606. [Google Scholar] [CrossRef]
- Nilsson, G.S.; Gorton, L.; Bergquist, K.E.; Nilsson, U. Determination of the degree of branching in normal and amylopectin type potato starch with 1H-NMR spectroscopy improved resolution and two-dimensional spectroscopy. Starch-Stärke 1996, 48, 352–357. [Google Scholar] [CrossRef]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.I.; Lee, Y.C. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef]
- Schiavone, M.; Vax, A.; Formosa, C.; Martin-Yken, H.; Dague, E.; François, J.M. A combined chemical and enzymatic method to determine quantitatively the polysaccharide components in the cell wall of yeasts. FEMS Yeast Res. 2014, 14, 933–947. [Google Scholar] [CrossRef]
- Ivshin, V.P.; Artamonova, S.D.; Ivshina, T.N.; Sharnina, F.F. Methods for isolation of chitin-glucan complexes from higher fungi native biomass. Polymer Sci. Ser. B 2007, 49, 305–310. [Google Scholar] [CrossRef]
- Passos, C.P.; Coimbra, M.A. Microwave superheated water extraction of polysaccharides from spent coffee grounds. Carbohydr. Polym. 2013, 94, 626–633. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds F1, F2, F3a, F3b and F5 are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Description | Yield (% w/w) | Main Component(s) |
|---|---|---|---|
| F1 | cold water extract | 6.14 | mannogalactan + proteins |
| F2 | hot water extract | 3.40 | (1→6)-β-d-glucan |
| F1′ | hot water extract | 10.32 | mannogalactan + glucans + proteins |
| F1′a | cold-water-soluble part | 5.83 | mannogalactan + glucans + proteins |
| F1′b | cold-water-insoluble part | 4.27 | mannogalactan + glucans + proteins |
| F3 | 1st alkaline extract | 15.63 | (1→3)-α-d-glucan + (1→3)(1→6)-β-d-glucan |
| F3a | DMSO-soluble part | 1.80 | (1→3)(1→6)-β-d-glucan |
| F3b | DMSO-insoluble part | 11.24 | (1→3)-α-d-glucan |
| F4 | 2nd alkaline extract | 1.54 | (1→3)(1→6)-β-d-glucan + proteins |
| F5 | insoluble part | 50.86 | (1→3)(1→6)-β-d-glucan + chitin |
| Fraction | Content (% w/w) | |||
|---|---|---|---|---|
| N | C | H | S | |
| F1 | 3.81 | 41.11 | 6.68 | 0.36 |
| F1′ | 2.58 | 37.53 | 6.12 | 0.28 |
| F2 | 0.51 | 39.72 | 6.10 | 0.09 |
| F3a | 0.39 | 40.75 | 6.78 | 0.75 |
| F3b | 0.19 | 41.83 | 6.87 | 0.23 |
| F4 | 2.01 | 41.93 | 6.82 | 0.15 |
| F5 | 2.22 | 40.47 | 6.74 | 0.10 |
| Content (% m/m of Dry Matter) | Fraction | |||
|---|---|---|---|---|
| F1 | F2 | F3 | F5 | |
| total glucans | 6.9 | 60.27 | 55.28 | 45.51 |
| α-glucans | 0.80 | 0.37 | 0.07 * | 0.01 |
| β-glucans | 6.14 | 59.90 | 55.21† | 45.49 |
| Fraction | Molar Ratio (%) | ||||||
|---|---|---|---|---|---|---|---|
| Glc | Fuc | Rha | Gal | Xyl | Man | Ara | |
| F1 | 10.6 | 2.0 | 3.5 | 45.6 | 4.6 | 28.3 | 5.4 |
| F1‘ | 48.4 | 1.9 | 2.7 | 17.8 | 4.4 | 18.8 | 6.0 |
| F2 | 97.1 | 0.1 | 0.2 | 0.9 | 0.3 | 1.2 | 0.2 |
| F3 | 96.0 | 0.3 | 0.2 | 0.6 | 0.1 | 0.9 | 1.9 |
| F5 | 94.9 | 0.6 | 1.2 | 0.6 | 1.1 | 0.8 | 0.8 |
| Fraction F1 | ||||||||
| Unit | H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | OCH3 | Assignment |
| A | 5.04 | 3.84 | 3.91 | 3.80 | 4.09 | 3.81; 3.59 | 1,2,6-β-Galp | |
| 98.12 | 77.02 | 68.57 | 66.84 | 68.91 | 66.76 | |||
| B | 4.89 | 3.73 | 3.44 | 3.77 | 4.12 | 3.81; 3.59 | 3.35 | 1,6-β-Galp3Me |
| 98.27 | 68.41 | 78.87 | 66.90 | 68.91 | 66.74 | 56.55 | ||
| C | 4.72 | 4.01 | 3.56 | 3.48 | 3.29 | 3.82; 3.63 | t-α-Manp | |
| 101.76 | 70.53 | 73.06 | 66.89 | 76.28 | 61.20 | |||
| Fraction F2 | ||||||||
| Unit | H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | Assignment | |
| A | 4.44 | 3.24 | 3.41 | 3.38 | 3.55 | 3.77 | 1,6-β-Glcp | |
| 103.07 | 73.17 | 75.50 | 69.42 | 75.00 | 68.96 | |||
| B | 4.46 | 3.47 | 3.68 | 3.48 | 3.58 | 3.77 | 1,3,6-β-Glcp | |
| 72.86 | 84.55 | 68.85 | 74.87 | 68.96 | ||||
| C | 4.66 | 3.27 | 3.45 | 3.32 | 3.37 | 3.64 | t-β-Glcp | |
| 73.17 | 75.70 | 69.66 | 60.84 | |||||
| Fraction F3a | ||||||||
| Unit | H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | Assignment | |
| B | 4.74 | 3.60 | 3.82 | 3.55 | 3.45 | 3.98; 3.73 | 1,3-β-Glcp | |
| 103.25 | 73.12 | 86.61 | 68.95 | 76.50 | 61.10 | |||
| C | 4.75 | 3.61 | 3.86 | 3.59 | 3.68 | 4.23; 3.85 | 1,3,6-β-Glcp | |
| 103.35 | 72.89 | 86.25 | 68.95 | 76.22 | 68.66 | |||
| D | 4.53 | 3.31 | 3.46 | 3.38 | 3.41 | 3.98; 3.73 | t-β-Glcp | |
| 103.57 | 73.93 | 76.45 | 70.30 | 77.12 | 61.23 | |||
| Fractions F3b | ||||||||
| Unit | H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | Assignment | |
| A | 5.19 | 3.50 | 3.72 | 3.38 | 3.87 | 3.78; 3.56 | 1,3-α-Glcp | |
| 100.92 | 71.25 | 83.45 | 70.83 | 73.07 | 61.22 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baeva, E.; Bleha, R.; Lavrova, E.; Sushytskyi, L.; Čopíková, J.; Jablonsky, I.; Klouček, P.; Synytsya, A. Polysaccharides from Basidiocarps of Cultivating Mushroom Pleurotus ostreatus: Isolation and Structural Characterization. Molecules 2019, 24, 2740. https://doi.org/10.3390/molecules24152740
Baeva E, Bleha R, Lavrova E, Sushytskyi L, Čopíková J, Jablonsky I, Klouček P, Synytsya A. Polysaccharides from Basidiocarps of Cultivating Mushroom Pleurotus ostreatus: Isolation and Structural Characterization. Molecules. 2019; 24(15):2740. https://doi.org/10.3390/molecules24152740
Chicago/Turabian StyleBaeva, Ekaterina, Roman Bleha, Ekaterina Lavrova, Leonid Sushytskyi, Jana Čopíková, Ivan Jablonsky, Pavel Klouček, and Andriy Synytsya. 2019. "Polysaccharides from Basidiocarps of Cultivating Mushroom Pleurotus ostreatus: Isolation and Structural Characterization" Molecules 24, no. 15: 2740. https://doi.org/10.3390/molecules24152740
APA StyleBaeva, E., Bleha, R., Lavrova, E., Sushytskyi, L., Čopíková, J., Jablonsky, I., Klouček, P., & Synytsya, A. (2019). Polysaccharides from Basidiocarps of Cultivating Mushroom Pleurotus ostreatus: Isolation and Structural Characterization. Molecules, 24(15), 2740. https://doi.org/10.3390/molecules24152740