
Antimicrobial and Phytotoxic Activity of Origanum heracleoticum and O. majorana Essential Oils Growing in Cilento (Southern Italy)


 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
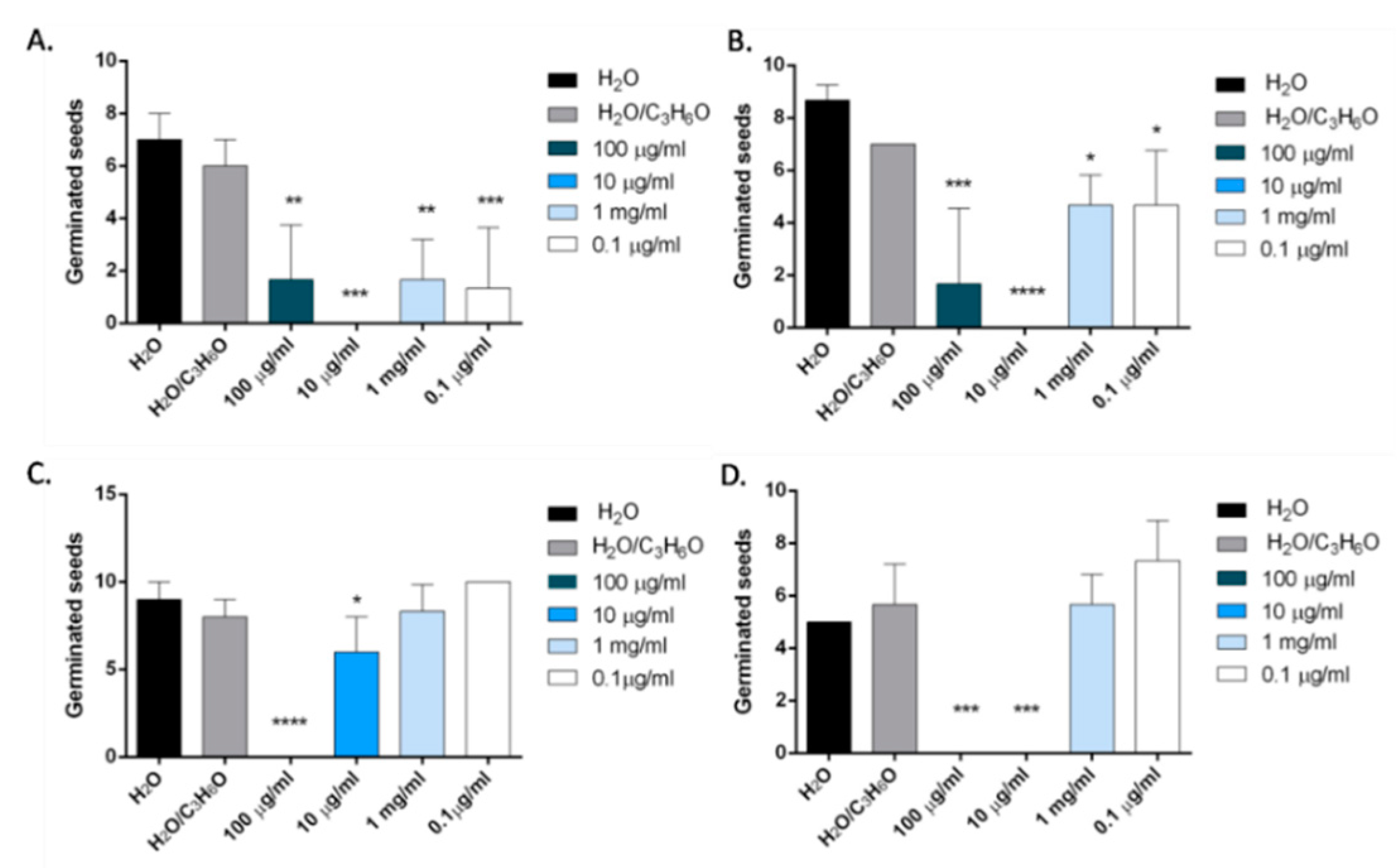
2. Results and Discussion
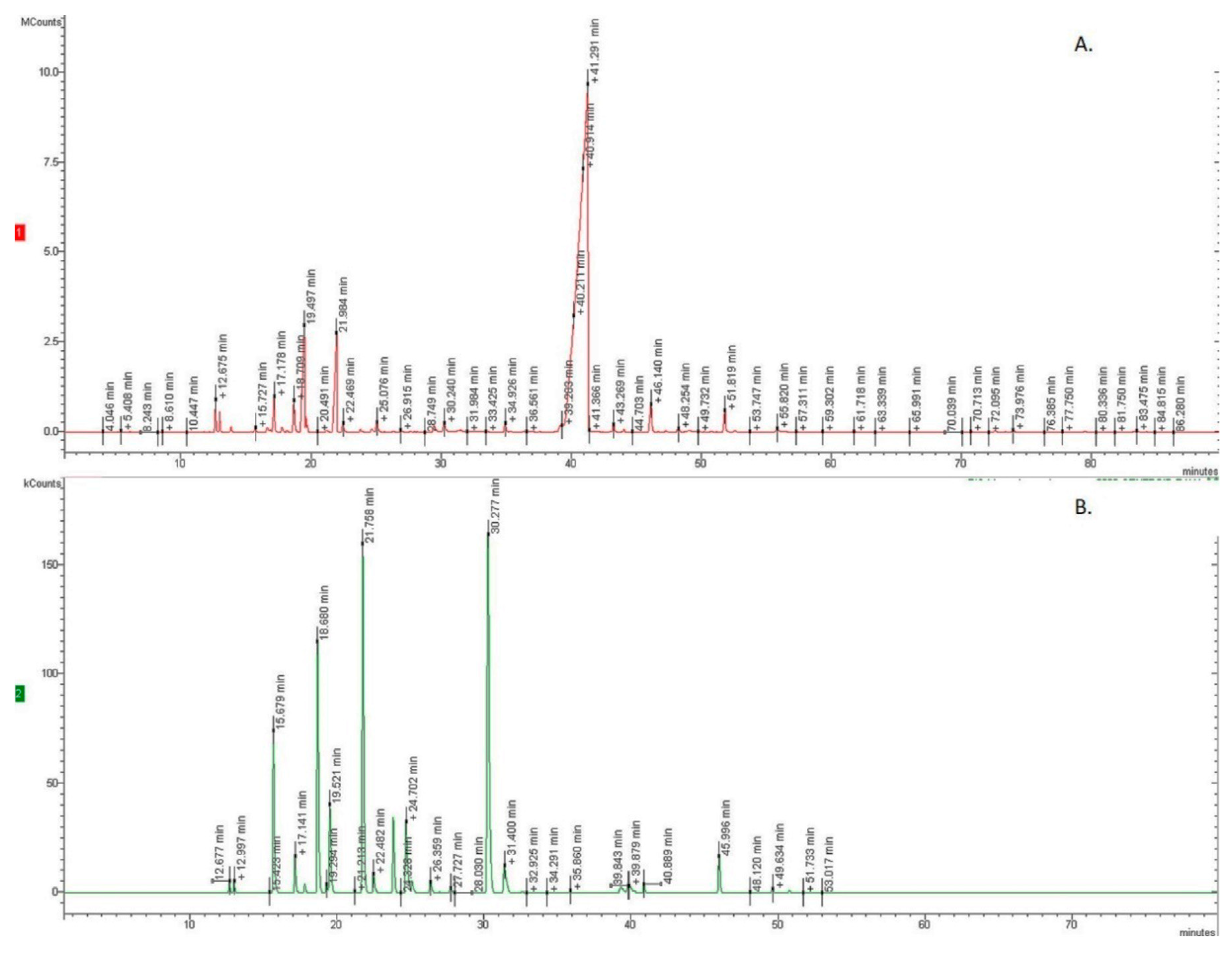
2.1. GC-MS Analysis
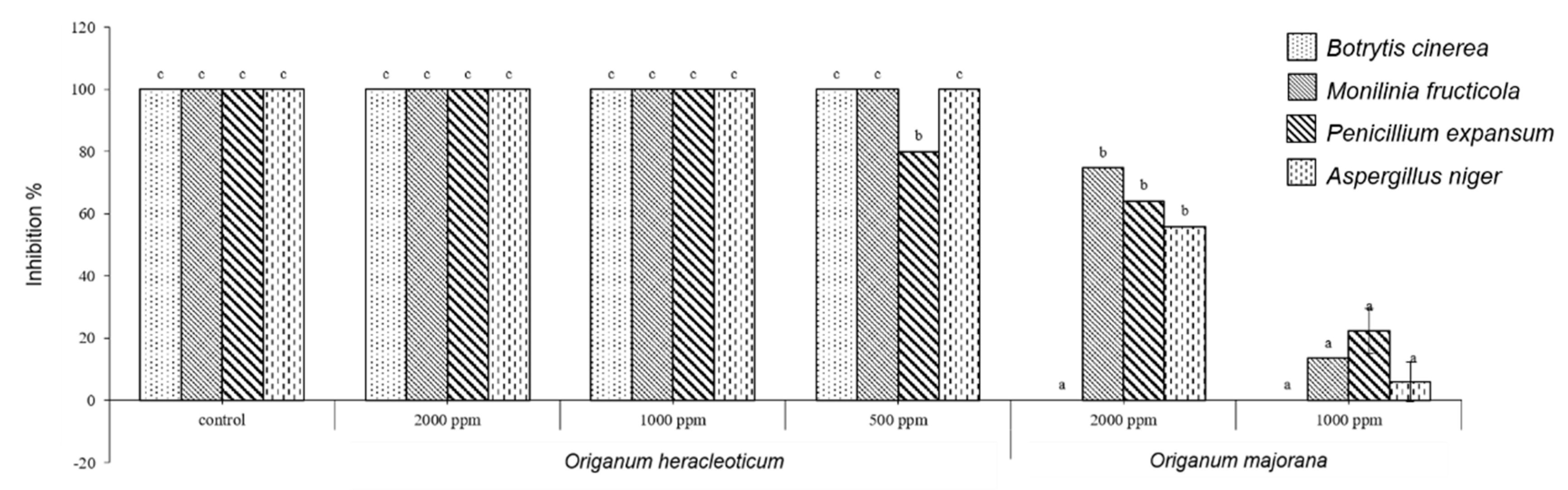
2.2. Antifungal In Vitro Test
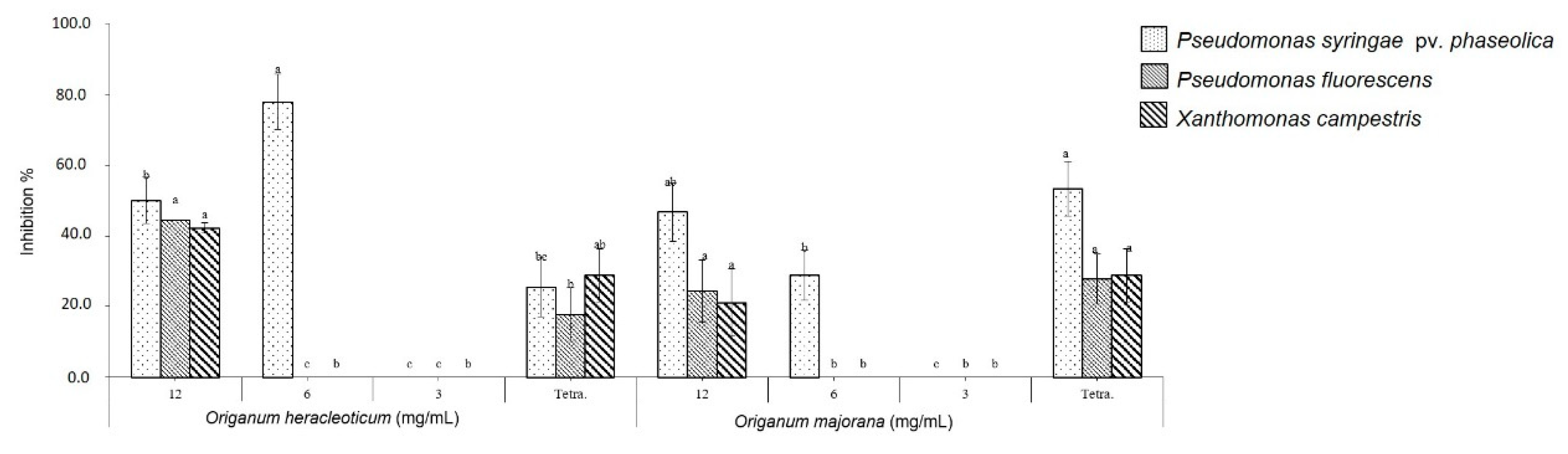
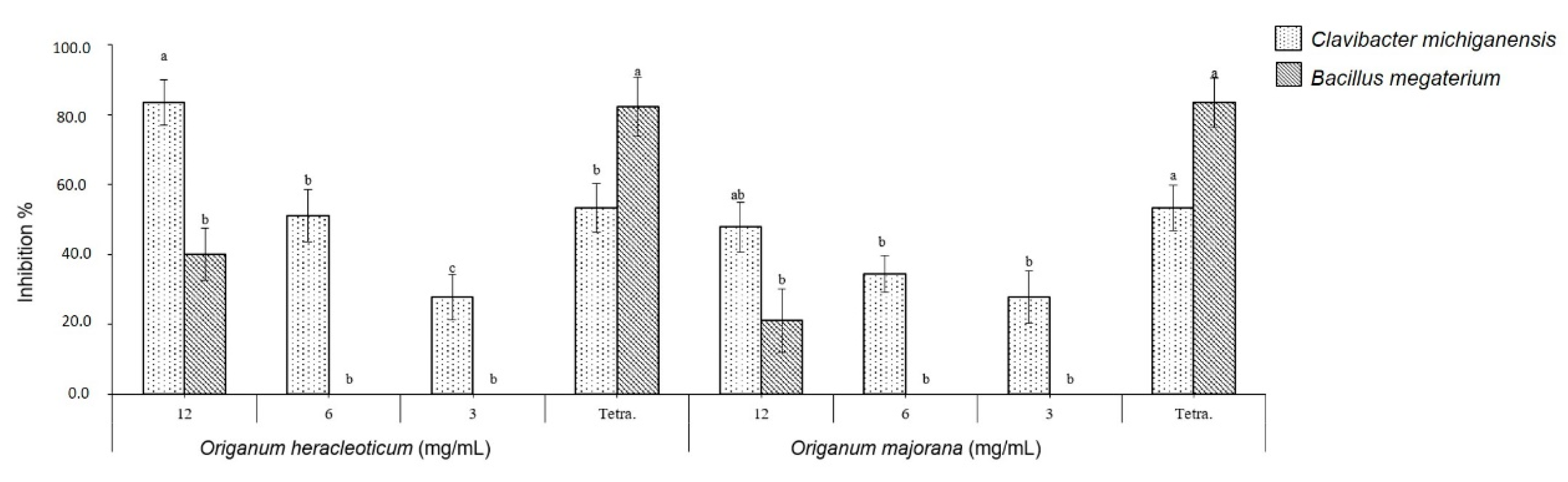
2.3. Antibacterial In Vitro Test
2.4. Fungal Spore Germination Assay
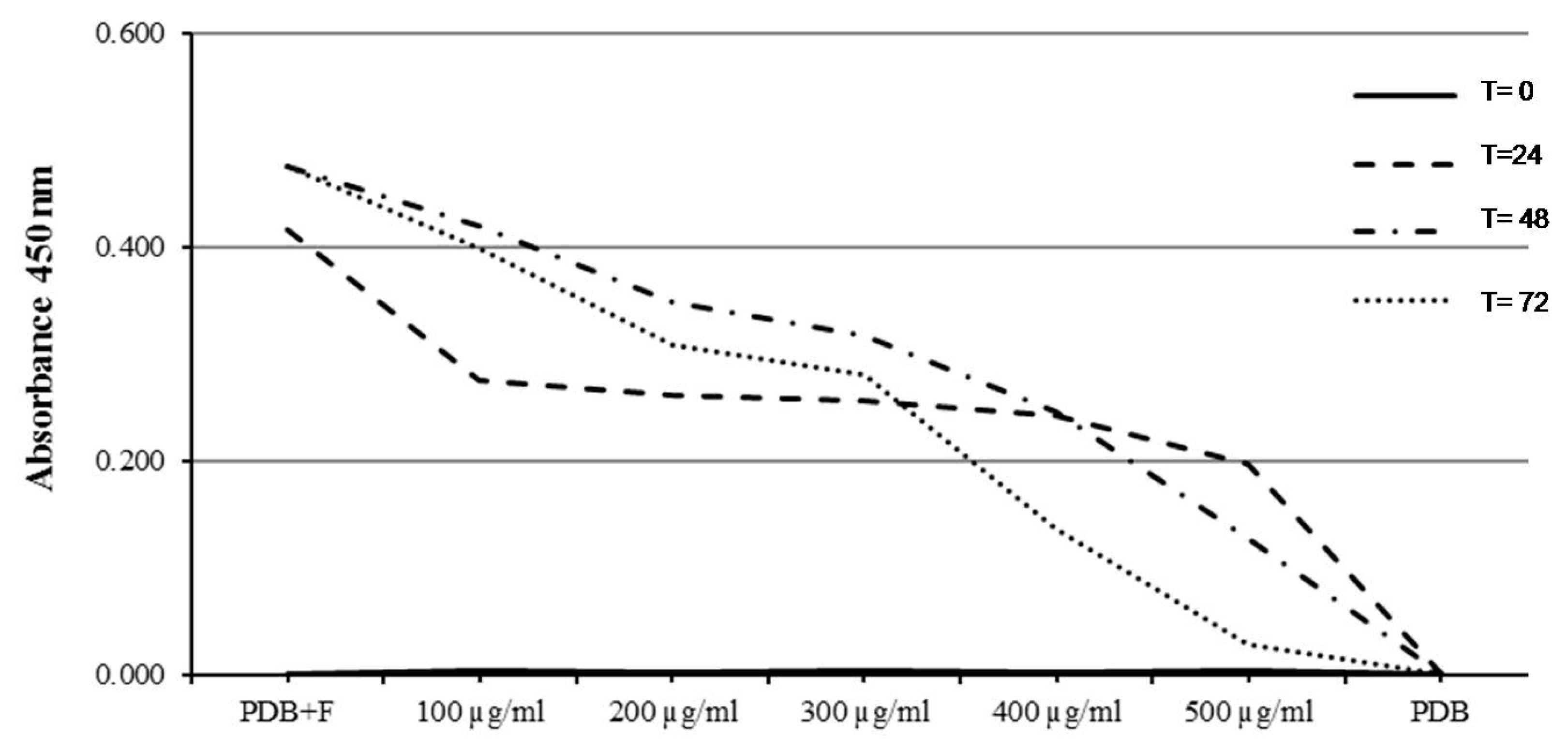
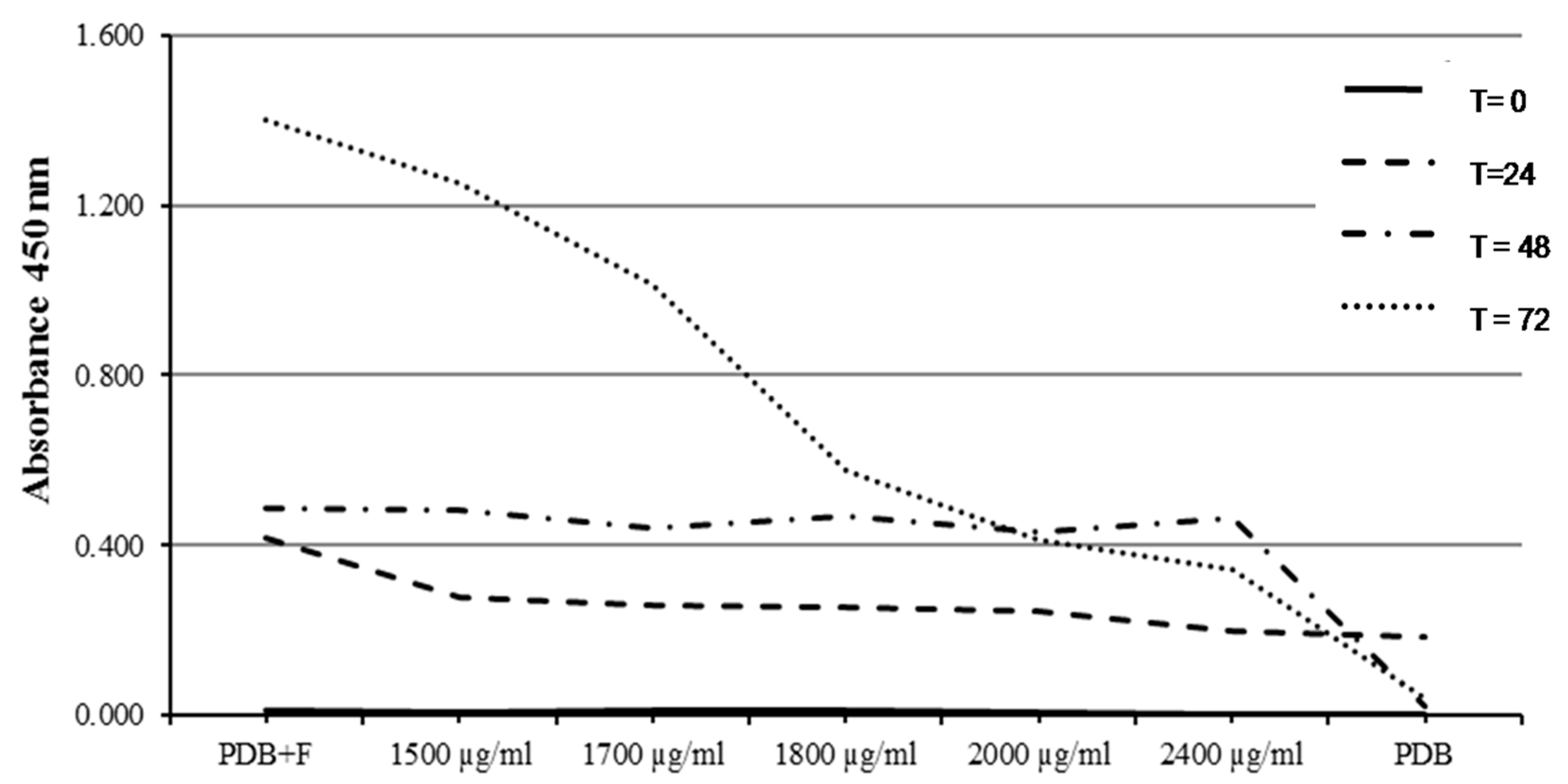
2.5. Determination of Minimum Inhibitory Concentration (MIC) (96-Well Microplate Method)
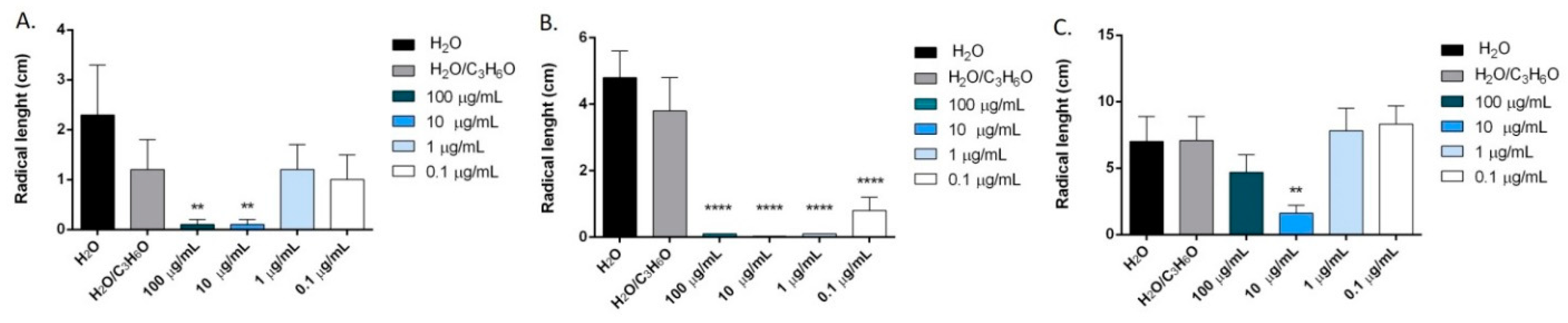
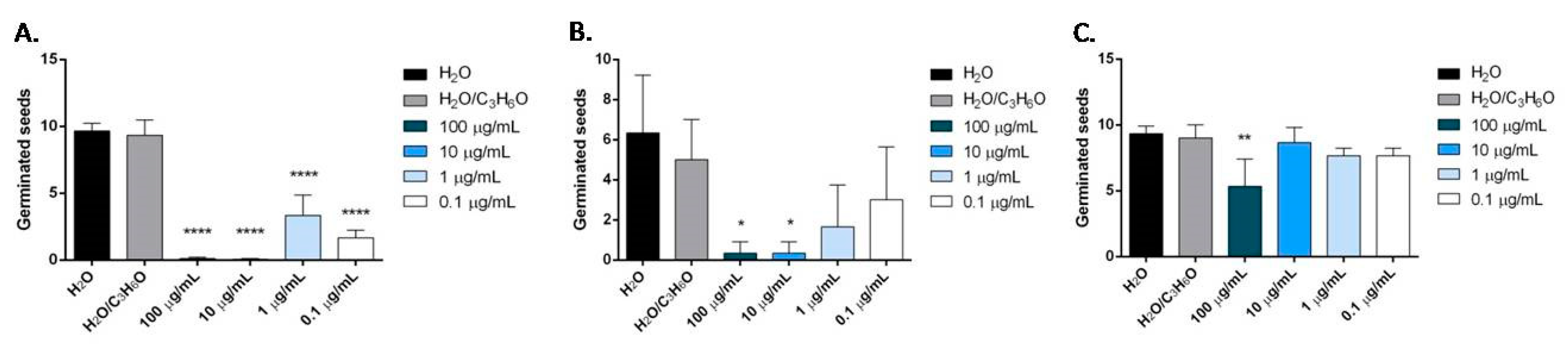
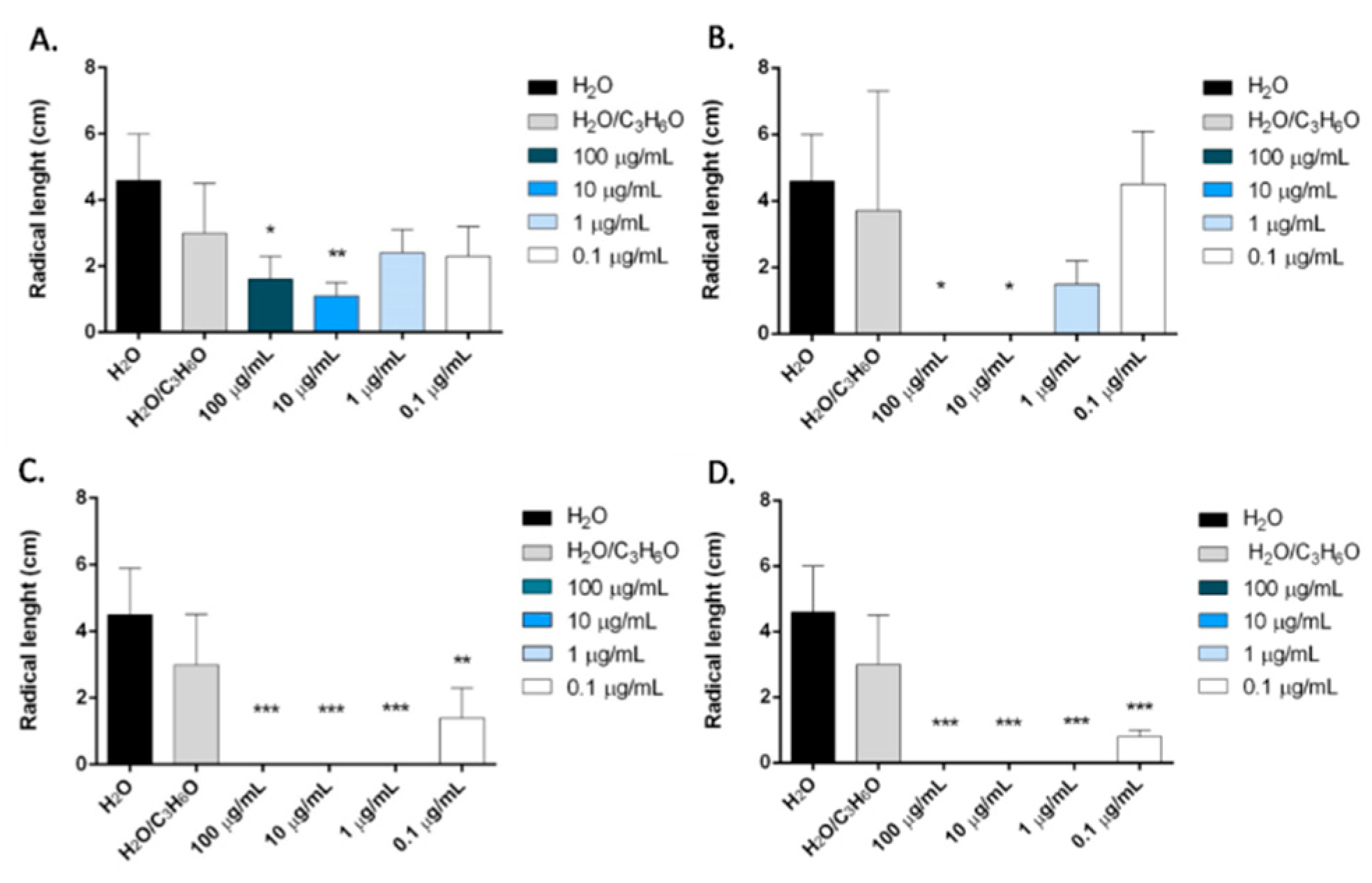
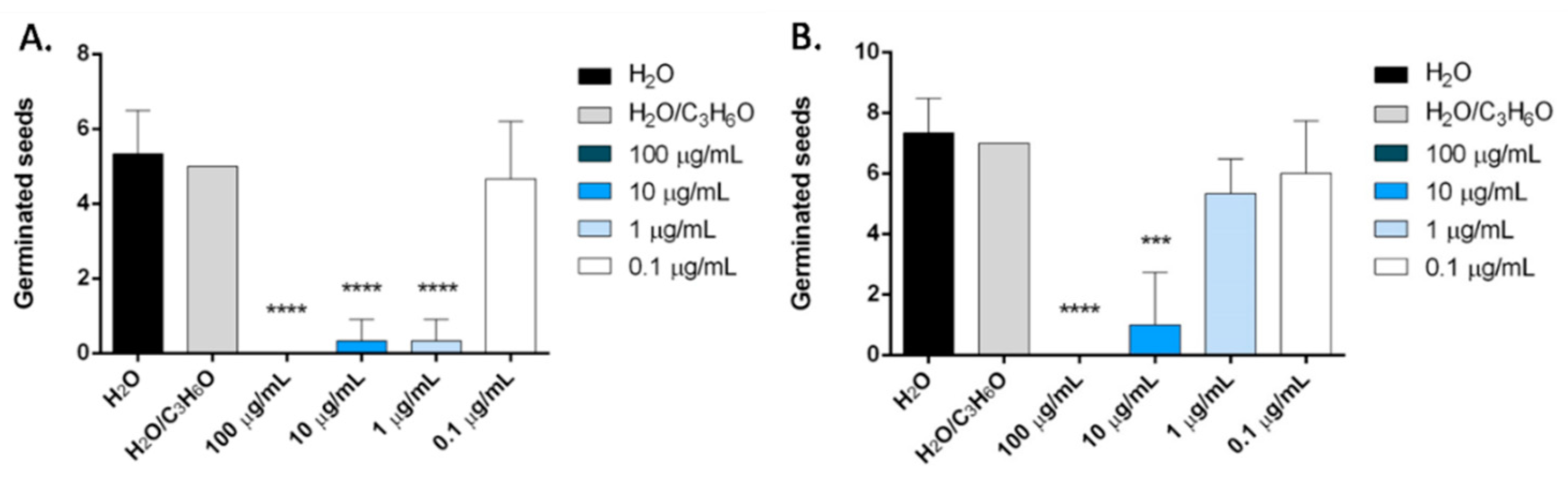
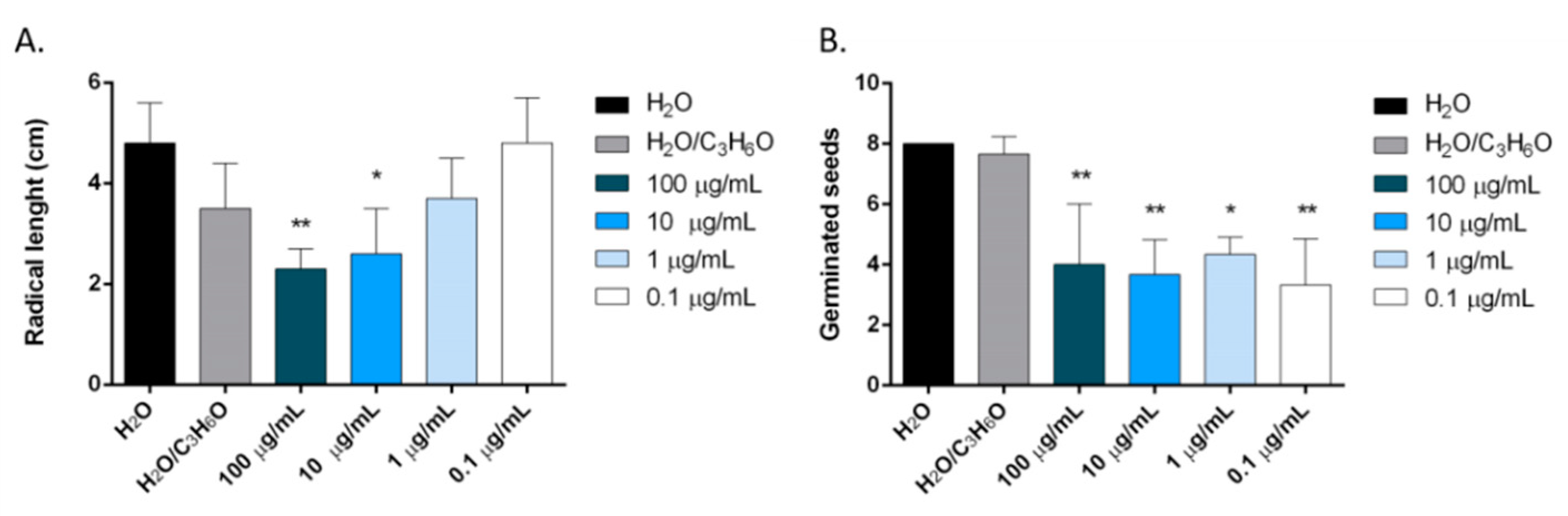
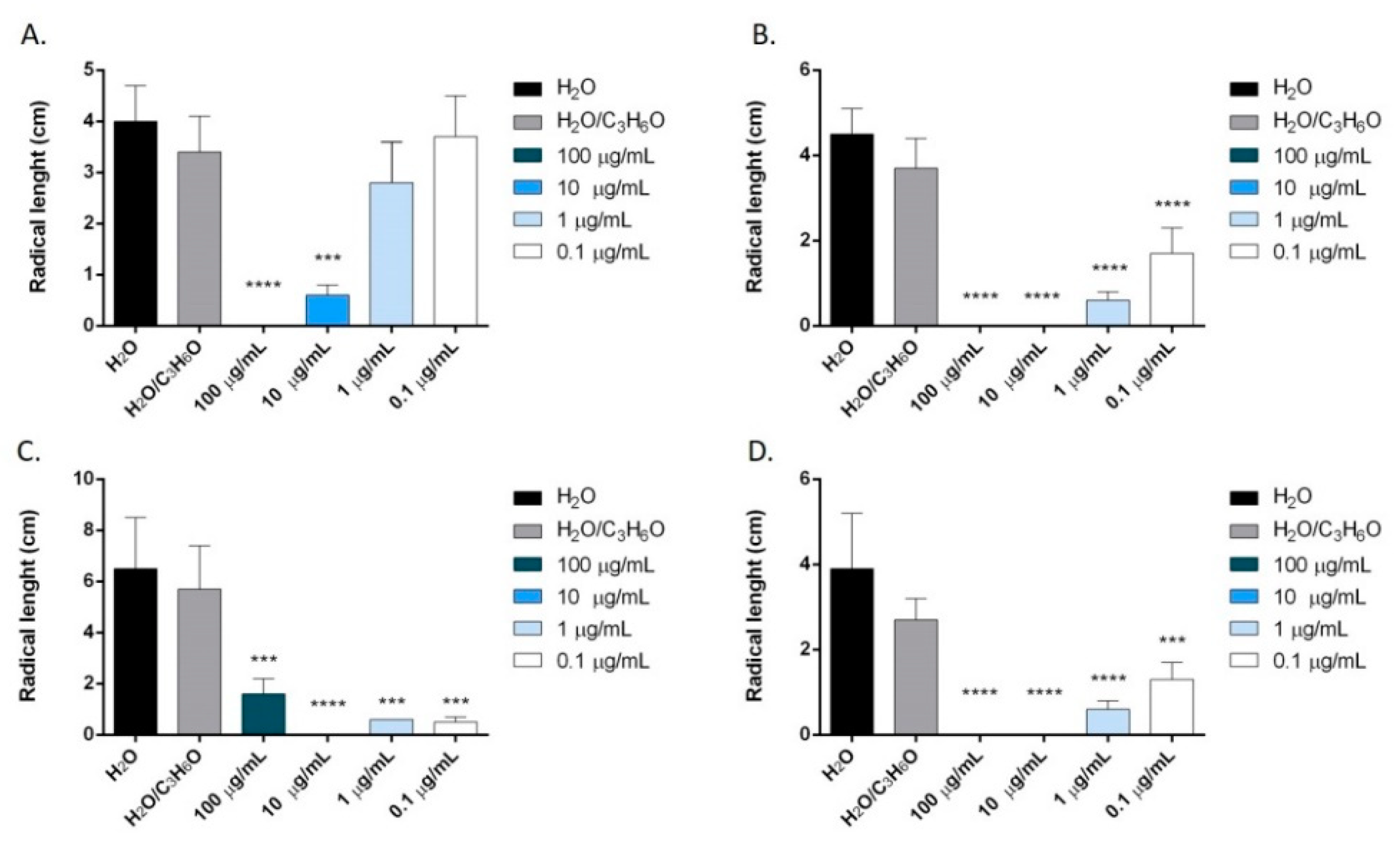
2.6. Phytotoxic Activity
3. Materials and Methods
3.1. Plant Materials
3.2. Isolation of the Volatile Oil
3.3. Gas Chromatography-Flame Ionization Detector (GC-FID) Analysis
3.4. GC/Mass Spectroscopy (MS) Analysis and Identification of Constituents
3.5. Antifungal Activity
3.5.1. Fungal Isolates
3.5.2. In Vitro Fungicidal Activity
3.6. Antibacterial Activity Assay
3.6.1. Bacterial Isolates
3.6.2. Antibacterial Activity Test
3.7. Fungal Spore Germination Assay
3.8. Determination of MIC (96-Well Microplate Method)
3.9. Phytotoxic Activity
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lopez-Reyes, J.G.; Spadaro, D.; Gullino, M.L.; Garibaldi, A. Efficacy of plant essential oils on postharvest control of rot caused by fungi on four cultivars of apples in vivo. Flavour Fragr. J. 2010, 25, 171–177. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Rolim De Almeida, L.F.R.; Frei, F.; Mancini, E.; De Martino, L.; De Feo, V. Phytotoxic Activities of Mediterranean Essential Oils. Molecules 2010, 15, 4309–4323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramezani, S.; Saharkhiz, M.J.; Ramezani, F.; Fotokian, M.H. Use of essential oils as bioherbicides. J. Essent. Oil Bear. Plants 2008, 11, 319–327. [Google Scholar] [CrossRef]
- Koul, O.; Walia, S.; Dhaliwal, G.S. Essential oils as green pesticides: Potential and constraints. Biopestic. Int. 2008, 4, 63–84. [Google Scholar]
- Mancini, E.; Camele, I.; Elshafie, H.S.; De Martino, L.; Pellegrino, C.; Grulova, D.; De Feo, V. Chemical Composition and Biological Activity of the Essential Oil of Origanum vulgare ssp. hirtum from Different Areas in the Southern Apennines. Chem. Biodivers. 2014, 11, 639–651. [Google Scholar] [CrossRef]
- Jerković, I.; Mastelić, J.; Miloš, M. The impact of both the season of collection and drying on the volatile constituents of Origanum vulgare L. ssp. hirtum grown wild in Croatia. Int. J. Food Sci. Technol. 2001, 36, 649–654. [Google Scholar] [CrossRef]
- Tripathy, B.; Satyanarayana, S.; Abedulla Khan, K.; Raja, K. An Updated Review on Traditional Uses, Taxonomy, Phytochemistry, Pharmacology and Toxicology of Origanum majorana. Int. J. Pharma Res. Health Sci. 2017, 5, 1717–1723. [Google Scholar]
- Dzamic, A.; Sokovic, M.; Ristic, M.S.; Grujic-Jovanovic, S.; Vukojevic, J.; Marin, P.D. Chemical composition and antifungal activity of Origanum heracleoticum essential oil. Chem. Nat. Compd. 2008, 44, 659–660. [Google Scholar] [CrossRef]
- Kokkini, S.; Karousou, R.; Hanlidou, E.; Lanaras, T. Essential oil composition of Greek (Origanum vulgare ssp. hirtum) and Turkish (O. onites) Origanum: A tool for their distinction. J. Essent. Oil Res. 2004, 16, 334–338. [Google Scholar] [CrossRef]
- Mith, H.; Clinquart, A.; Zhiri, A.; Daube, G.; Delcenserie, V. The impact of Origanum (Origanum heracleoticum) essential oil and carvacrol on virulence gene transcription by Escherichia coli O157:H7. FEMS Microbiol. Lett. 2015, 362, 1–7. [Google Scholar] [CrossRef]
- Abbassy, M.A.; Abdelgaleil, S.A.; Rabie, R.Y. Insecticidal and synergistic effects of Origanum majorana essential oil and some of its major constituents. Entomol. Exp. Appl. 2009, 131, 225–232. [Google Scholar] [CrossRef]
- Brosche, T.; Vostrowsky, O.; Gemeinhardt, F.; Asmus, U.; Knobloch, K. On the Essential Oil Components from Majorana hortensis Moench. Z. Naturforsch. C 1981, 36, 23–29. [Google Scholar] [CrossRef]
- Kadoglidou, K.; Lagopodi, A.; Karamanoli, K.; Vokou, D.; Bardas, G.A.; Menexes, G.; Constantinidou, H.I.A. Inhibitory and stimulatory effects of essential oils and individual monoterpenoids on growth and sporulation of four soil-borne fungal isolates of Aspergillus terreus, Fusarium oxysporum, Penicillium expansum, and Verticillium dahliae. Eur. J. Plant Pathol. 2011, 130, 297–309. [Google Scholar] [CrossRef]
- Daferera, D.J.; Ziogas, B.N.; Polissiou, M.G. The effectiveness of plant essential oils on the growth of Botrytis cinerea, Fusarium sp. and Clavibacter michiganensis subsp. michiganensis. Crop Prot. 2003, 22, 39–44. [Google Scholar] [CrossRef]
- Stavropoulou, A.; Loulakakis, K.; Magan, N.; Tzortzakis, N. Origanum dictamnus oil vapour suppresses the development of grey mould in eggplant fruit in vitro. BioMed Res. Int. 2014, 2014, 562679. [Google Scholar] [CrossRef] [PubMed]
- Felsociova, S.; Kacaniova, M.; Horská, E.; Vukovic, N.; Hleba, L.; Petrová, J.; Hajduová, Z. Antifungal activity of essential oils against selected terverticillate penicillia. Ann. Agric. Environ. Med. 2015, 22, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Deleanu, M.; Popa, E.E.; Popa, M.E. Chemical Composition and Active Properties Evaluation of Wild Oregano (Origanum Vulgare) and Ginger (Zingiber Officinale-Roscoe) Essential Oils. Rev. Chim. 2018, 69, 1927–1933. [Google Scholar]
- Bayramoglu, E.E.; Gulumser, G.; Karaboz, I. Ecological and innovative fungicide for the leather industry: Essential oil of Origanum minutiflorum. JALCA 2006, 101, 96–104. [Google Scholar]
- Soylu, E.M.; Kurt, S.; Soylu, S. In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. Int. J. Food Microbiol. 2010, 143, 183–189. [Google Scholar] [CrossRef]
- Veldhuizen, E.J.; Tjeerdsma-van Bokhoven, J.L.; Zweijtzer, C.; Burt, S.A.; Haagsman, H.P. Structural requirements for the antimicrobial activity of carvacrol. J. Agric. Food Chem. 2006, 54, 1874–1879. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Armentano, M.; Carmosino, M.; Bufo, S.; De Feo, V.; Camele, I. Cytotoxic activity of Origanum vulgare L. on hepatocellular carcinoma cell line HepG2 and evaluation of its biological activity. Molecules 2017, 22, 1435. [Google Scholar] [CrossRef] [PubMed]
- Sarac, N.; Ugur, A. Antimicrobial activities of the essential oils of Origanum onites L., Origanum vulgare L. subspecies hirtum (Link) Ietswaart, Satureja thymbra L., and Thymus cilicicus Boiss. & Bal. growing wild in Turkey. J. Med. Food 2008, 11, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Özcan, M.M.; Sadıç, O.; Özkan, G. Inhibitory effects of spice essential oils on the growth of Bacillus species. J. Med. Food 2006, 9, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Chudasama, K.S.; Thaker, V.S. Screening of potential antimicrobial compounds against Xanthomonas campestris from 100 essential oils of aromatic plants used in India: An ecofriendly approach. Arch. Phytopathol. Plant Protect. 2012, 45, 783–795. [Google Scholar] [CrossRef]
- Araniti, F.; Landi, M.; Lupini, A.; Sunseri, F.; Guidi, L.; Abenavoli, M.R. Origanum vulgare essential oils inhibit glutamate and aspartate metabolism altering the photorespiratory pathway in Arabidopsis thaliana seedlings. J. Plant Physiol. 2018, 231, 297–309. [Google Scholar] [CrossRef]
- Kordali, S.; Cakir, A.; Ozer, H.; Cakmakci, R.; Kesdek, M.; Mete, E. Antifungal, phytotoxic and insecticidal properties of essential oil isolated from Turkish Origanum acutidens and its three components, carvacrol, thymol and p-cymene. Bioresour. Technol. 2008, 99, 8788–8795. [Google Scholar] [CrossRef]
- Vasilakoglou, I.; Dhima, K.; Paschalidis, K.; Ritzoulis, C. Herbicidal potential on Lolium rigidum of nineteen major essential oil components and their synergy. J. Essent. Oil Res. 2013, 25, 1–10. [Google Scholar] [CrossRef]
- De Assis Alves, T.; Pinheiro, P.F.; Praça-Fontes, M.M.; Andrade-Vieira, L.F.; Corrêa, K.B.; de Assis Alves, T.; Soares, T.C.B. Toxicity of thymol, carvacrol and their respective phenoxyacetic acids in Lactuca sativa and Sorghum bicolor. Ind. Crops Prod. 2018, 114, 59–67. [Google Scholar] [CrossRef]
- Pinheiro, P.F.; Costa, A.V.; Alves, T.D.A.; Galter, I.N.; Pinheiro, C.A.; Pereira, A.F.; Fontes, M.M.P. Phytotoxicity and cytotoxicity of essential oil from leaves of Plectranthus amboinicus, carvacrol, and thymol in plant bioassays. J. Agric. Food Chem. 2015, 63, 8981–8990. [Google Scholar] [CrossRef]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole: Bologna, Italy, 2017; Available online: https://img.ibs.it/pdf/9788850652426.pdf (accessed on 20 June 2019).
- Council of Europe. European Pharmacopeia, 5th ed.; Council of Europe: Strasbourg Cedex, France, 2014; Volume I, pp. 217–218. [Google Scholar]
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicone and Carbowax 20M phases. J. Chromatogr. 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 2007. [Google Scholar]
- Goodner, K.L. Practical retention index models of OV-101, DB-1, DB-5, and DB-Wax for flavor and fragrance compounds. LWT Food Sci. Technol. 2008, 41, 951–958. [Google Scholar] [CrossRef]
- McLafferty, F.W. The Wiley Registry of Mass Spectral Data, with NIST Spectral Data CD Rom, 7th ed.; John Wiley & Sons: New York, NY, USA, 1998. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSIBLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Ghanney, N.; Mang, S.M.; Ferchichi, A.; Camele, I. An in vitro attempt for controlling severe phyto and human pathogens using essential oils from Mediterranean plants of genus Schinus. J. Med. Food 2016, 19, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Zygadlo, J.A.; Guzman, C.A.; Grosso, N.R. Antifungal properties of the leaf oils of Tagetes minuta L. and Tagetes filifolia Lag. J. Essent. Oil Res. 1994, 6, 617–621. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Sakr, S.; Mang, S.M.; De Feo, V.; Camele, I. Antimicrobial activity and chemical composition of three essential oils extracted from Mediterranean aromatic plants. J. Med. Food 2016, 19, 1096–1103. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Racioppi, R.; Bufo, S.A.; Camele, I. In vitro study of biological activity of four strains of Burkholderia gladioli pv. agaricicola and identification of their bioactive metabolites using GC-MS. Saudi J. Biol. Sci. 2017, 24, 295–301. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Mancini, E.; De Martino, L.; Pellegrino, C.; Camele, I.; De Feo, V. Antifungal activity of some constituents of Origanum vulgare L. essential oil against postharvest disease of peach fruit. J. Med. Food 2015, 18, 929–934. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055. [Google Scholar] [CrossRef]
Sample Availability: Samples of the O. heracleoticum and O. majorana essential oils are available from the authors. |














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds | % | KI a | KI b | Identification c |
|---|---|---|---|---|---|
| 1 | α-Thujene | 0.7 | 856 | 930 | 1,2,3 |
| 2 | α-Pinene | 0.4 | 860 | 939 | 1,2,3 |
| 3 | Camphene | 0.1 | 871 | 954 | 1,2,3 |
| 4 | Sabinene | 0.4 | 896 | 975 | 1,2,3 |
| 5 | β-Pinene | 1.0 | 914 | 979 | 1,2,3 |
| 6 | α-Phellandrene | 0.2 | 922 | 1002 | 1,2,3 |
| 7 | δ-3-Carene | 0.1 | 927 | 1011 | 1,2 |
| 8 | α-Terpinene | 1.1 | 934 | 1017 | 1,2,3 |
| 9 | ρ–Cimene | 5.3 | 944 | 1024 | 1,2 |
| 10 | Limonene | 0.3 | 946 | 1029 | 1,2,3 |
| 11 | 1,8 Cineol | 0.1 | 947 | 1031 | 1,2,3 |
| 12 | (E)-β-Ocimene | 0.1 | 966 | 1050 | 1,2,3 |
| 13 | γ-Terpinene | 4.9 | 977 | 1059 | 1,2,3 |
| 14 | cis-Sabinene hydrate | 0.3 | 983 | 1070 | 1,2 |
| 15 | Terpinolene | 0.1 | 1000 | 1088 | 1,2 |
| 16 | Linalool | 0.4 | 1017 | 1096 | 1,2,3 |
| 17 | trans–Sabinene hydrate | t | 1033 | 1098 | 1,2 |
| 18 | allo-Ocimene | 0.1 | 1040 | 1132 | 1,2 |
| 19 | Borneol | 0.4 | 1073 | 1169 | 1,2,3 |
| 20 | Terpinen-4-ol | 0.5 | 1083 | 1177 | 1,2,3 |
| 21 | α-Terpineol | 0.1 | 1097 | 1188 | 1,2,3 |
| 22 | cis–Dihydrocarvone | 0.1 | 1110 | 1192 | 1,2 |
| 23 | trans-Dihydrocarvone | t | 1106 | 1200 | 1,2 |
| 24 | Thymol methyl ether | 0.3 | 1146 | 1235 | 1,2 |
| 25 | Thymoquinone | t | 1151 | 1252 | 1,2 |
| 26 | Thymol | 0.1 | 1204 | 1290 | 1,2,3 |
| 27 | Carvacrol | 77.8 | 1218 | 1299 | 1,2,3 |
| 28 | Eugenol | 0.2 | 1261 | 1359 | 1,2,3 |
| 29 | (E)-Caryophyllene | 1.3 | 1303 | 1419 | 1,2 |
| 30 | Aromadendrene | t | 1320 | 1441 | 1,2,3 |
| 31 | α-Humulene | 0.2 | 1335 | 1454 | 1,2,3 |
| 32 | Aceto-vanillone | 0.1 | 1346 | 1482 | 1,2 |
| 33 | Germacrene A | 0.8 | 1389 | 1509 | 1,2 |
| 34 | Caryophyllene oxide | 0.2 | 1452 | 1583 | 1,2,3 |
| 35 | α Bisabolol | t | 1150 | 1685 | 1,2,3 |
| Total | 97.8 | ||||
| Monoterpene hydrocarbons | 15.1 | ||||
| Oxygenated monoterpenes | 80.1 | ||||
| Sesquiterpene hydrocarbons | 2.3 | ||||
| Oxygenated sesquiterpenes | 0.2 |
| No. | Compounds | % | K a | K b | Identification c |
|---|---|---|---|---|---|
| 1 | α-Thujene | 0.4 | 856 | 930 | 1,2,3 |
| 2 | α-Pinene | 7.9 | 895 | 939 | 1,2,3 |
| 3 | α– Fenchene | 2.2 | 914 | 951 | 1,2 |
| 4 | Camphene | 13.4 | 934 | 954 | 1,2,3 |
| 5 | Verbetene | 0.4 | 942 | 967 | 1,2 |
| 6 | trans-Isolimonene | 5.4 | 945 | 984 | 1,2 |
| 7 | δ-2-Carene | 20.1 | 974 | 1002 | 1,2 |
| 8 | α-Terpinene | 4.2 | 1000 | 1017 | 1,2,3 |
| 9 | cis- Sabinene hydrate | 5.4 | 1009 | 1070 | 1,2 |
| 10 | Linalool | 1.0 | 1017 | 1096 | 1,2,3 |
| 11 | cis-p-Menth-2-en-1-ol | 0.8 | 1033 | 1121 | 1,2 |
| 12 | allo-Ocimene | t | 1040 | 1132 | 1,2,3 |
| 13 | trans-p-Menth-2-en-1-ol | 0.4 | 1051 | 1177 | 1,2 |
| 14 | Terpinen-4-ol | 29.6 | 1083 | 1177 | 1,2 |
| 15 | α-Terpineol | 2.8 | 1087 | 1188 | 1,2 |
| 16 | Thymol | 0.3 | 1204 | 1290 | 1,2,3 |
| 17 | Carvacrol | 0.6 | 1218 | 1299 | 1,2,3 |
| 18 | δ-Elemene | 0.5 | 1227 | 1338 | 1,2 |
| 19 | (E)-Caryophyllene | 2.2 | 1003 | 1419 | 1,2,3 |
| 20 | cis-Muurola-3,5-diene | 0.2 | 1353 | 1450 | 1,2 |
| Total | 98.0 | ||||
| Monoterpene hydrocarbons | 54.0 | ||||
| Oxygenated monoterpenes | 40.9 | ||||
| Sesquiterpene hydrocarbons | 2.9 |
| EOs | * Spore/mL | ** Spore Germination % | *** Inhibition Rate % | |
|---|---|---|---|---|
| Control | 80.0 × 104 | 100.0 | 0.0 | |
| O. heracleoticum EO | 15 µg/µL | 16.7 × 104 | 20.9 a | 79.1 ab |
| 22 µg/µL | 92.5 × 104 | 11.6 b | 88.4 a | |
| 45 µg/µL | 45.0 × 104 | 5.6 b | 94.4 a | |
| O. majorana EO | 15 µg/µL | 16.8 × 104 | 23.5 a | 76.5 ab |
| 22 µg/µL | 10.2 × 104 | 12.8 ab | 87.2 a | |
| 45 µg/µL | 5.5 × 104 | 6.9 ab | 93.1 a | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Della Pepa, T.; Elshafie, H.S.; Capasso, R.; De Feo, V.; Camele, I.; Nazzaro, F.; Scognamiglio, M.R.; Caputo, L. Antimicrobial and Phytotoxic Activity of Origanum heracleoticum and O. majorana Essential Oils Growing in Cilento (Southern Italy). Molecules 2019, 24, 2576. https://doi.org/10.3390/molecules24142576
Della Pepa T, Elshafie HS, Capasso R, De Feo V, Camele I, Nazzaro F, Scognamiglio MR, Caputo L. Antimicrobial and Phytotoxic Activity of Origanum heracleoticum and O. majorana Essential Oils Growing in Cilento (Southern Italy). Molecules. 2019; 24(14):2576. https://doi.org/10.3390/molecules24142576
Chicago/Turabian StyleDella Pepa, Teresa, Hazem S. Elshafie, Raffaele Capasso, Vincenzo De Feo, Ippolito Camele, Filomena Nazzaro, Maria Rosa Scognamiglio, and Lucia Caputo. 2019. "Antimicrobial and Phytotoxic Activity of Origanum heracleoticum and O. majorana Essential Oils Growing in Cilento (Southern Italy)" Molecules 24, no. 14: 2576. https://doi.org/10.3390/molecules24142576