
Xylopia aethiopica Seeds from Two Countries in West Africa Exhibit Differences in Their Proteomes, Mineral Content and Bioactive Phytochemical Composition

Abstract
:1. Introduction
2. Results and Discussion
2.1. External Characteristics of X. Aethiopica Seeds and Fruits
2.2. Proteomics
2.2.1. Total Number of Proteins and Significantly Changed Common Proteins
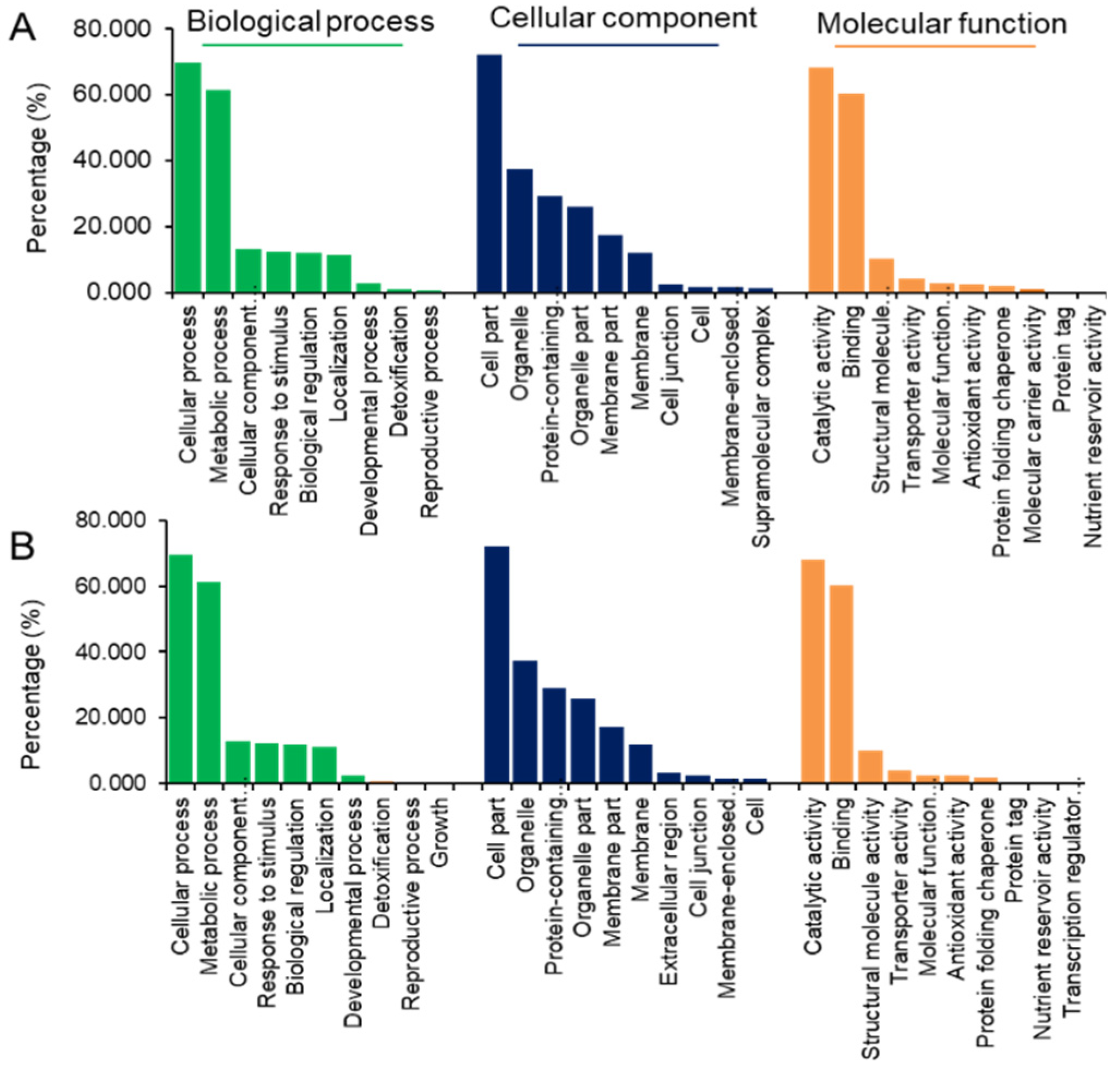
2.2.2. Functional Categorization
2.2.3. Pathway Enrichment Analysis
2.3. Elemental Analysis
2.4. Phytochemical Analysis
2.4.1. Total Phenolic Content (TPC)
2.4.2. Total Flavonoids Content (TFC)
2.4.3. Antioxidant Activities
3. Materials and Methods
3.1. Plant Material and Chemicals
3.2. Proteomics
3.2.1. Extraction of Protein
3.2.2. Protein Quantification and Digestion
3.2.3. Proteomic Data Processing
3.3. Quantitative Elemental Analysis
3.3.1. Sample Preparation (Microwave Acid-Assisted Digestion)
3.3.2. Inductively Coupled Plasma-Optical Emission Spectrometric (ICP-OES) Analysis
3.4. Phytochemical Analysis
3.4.1. Sample Preparation
3.4.2. Total Phenolic Content Determination (TPC)
3.4.3. Total Flavonoid Content Determination (TFC)
3.4.4. Antioxidant Activities
ABTS
Cupric Ion Reducing Capacity (CUPRIC)
DPPH Radical Scavenging Activity
Ferric Reducing Antioxidant Power Capacity (FRAP)
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fetse, J.P.; Kofie, W.; Adosraku, R.K. Ethnopharmacological importance of Xylopia aethiopica (Dunal) A. Rich (Annonaceae)—A Review. Br. J. Pharm. Res. 2016, 11, 1–21. [Google Scholar] [CrossRef]
- Boyom, F.F.; Ngouana, V.; Zollo, P.H.; Menut, C.; Bessiere, J.M.; Gut, J.; Rosenthal, P.J. Composition and anti-plasmodial activities of essential oils from some Cameroonian medicinal plants. Phytochemistry 2003, 64, 1269–1275. [Google Scholar] [CrossRef]
- Boampong, J.N.; Ameyaw, E.O.; Aboagye, B.; Asare, K.; Kyei, S.; Donfack, J.H.; Woode, E. The Curative and Prophylactic Effects of Xylopic Acid on Plasmodium berghei Infection in Mice. J. Parasitol. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Woode, E.; Ameyaw, E.O.; Boakye-Gyasi, E.; Abotsi, W.K.M. Analgesic effects of an ethanol extract of the fruits of Xylopia aethiopica (Dunal) A. Rich (Annonaceae) and the major constituent, xylopic acid in murine models. J. Pharm. Bioallied Sci. 2012, 4, 291–301. [Google Scholar] [CrossRef]
- Obiri, D.D.; Osafo, N. Aqueous ethanol extract of the fruit of Xylopia aethiopica (Annonaceae) exhibits anti-anaphylactic and anti-inflammatory actions in mice. J. Ethnopharmacol. 2013, 148, 940–945. [Google Scholar] [CrossRef]
- Mohammed, A.; Koorbanally, N.A.; Islam, M.S. Anti-diabetic effect of Xylopia aethiopica (Dunal) A. Rich. (Annonaceae) fruit acetone fraction in a type 2 diabetes model of rats. J. Ethnopharmacol. 2016, 180, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Asekun, O.; Adeniyi, B. Antimicrobial and cytotoxic activities of the fruit essential oil of Xylopia aethiopica from Nigeria. Fitoterapia 2004, 75, 368–370. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, T.; Mensah, M.; Komlaga, G.; Gbedema, S.; Skaltsa, H. Antimicrobial Activity Of Essential Oils of Xylopia aethiopica. Afr. J. Tradit. Complement. Altern. Med. 2008, 5, 391. [Google Scholar] [CrossRef] [PubMed]
- Reguera, M.; Conesa, C.M.; Gil-Gómez, A.; Haros, C.M.; Pérez-Casas, M.Á.; Briones-Labarca, V.; Bolaños, L.; Bonilla, I.; Alvarez, R.; Pinto, K.; et al. The impact of different agroecological conditions on the nutritional composition of quinoa seeds. Peer J. 2018, 6, e4442. [Google Scholar] [CrossRef] [PubMed]
- Alolga, R.N.; Chavez, M.A.S.C.; Muyaba, M. Untargeted UPLC-Q/TOF-MS-based metabolomics and inductively coupled plasma optical emission spectroscopic analysis reveal differences in the quality of ginger from two provinces in Zambia. J. Pharm. Pharmacol. 2018, 70, 1262–1271. [Google Scholar] [CrossRef]
- Yin, X.; Wang, S.-L.; Alolga, R.N.; Mais, E.; Li, P.; Yang, P.; Komatsu, S.; Qi, L.-W. Label-free proteomic analysis to characterize ginger from China and Ghana. Food Chem. 2018, 249, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Vitamvas, P.; Urban, M.O.; Prášil, I.T.; Renaut, J. Plant Abiotic Stress Proteomics: The Major Factors Determining Alterations in Cellular Proteome. Front. Plant Sci. 2018, 9, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; He, D.; Teng, H.; Chen, L.; Song, H.; Huang, Q. Physiological and proteomic analyses of coix seed aging during storage. Food Chem. 2018, 260, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Livramento, K.G.D.; Borém, F.M.; José, A.C.; Santos, A.V.; Livramento, D.E.D.; Alves, J.D.; Paiva, L.V. Proteomic analysis of coffee grains exposed to different drying process. Food Chem. 2017, 221, 1874–1882. [Google Scholar] [CrossRef]
- Bao, F.; Huang, X.; Zhu, C.; Zhang, X.; Li, X.; Yang, S. Arabidopsis HSP90 protein modulates RPP4-mediated temperature-dependent cell death and defense responses. New Phytol. 2014, 202, 1320–1334. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Monaghan, J.; Zhong, X.; Lin, L.; Sun, T.; Dong, O.X.; Li, X. HSP90s are required for NLR immune receptor accumulation in Arabidopsis. Plant J. 2014, 79, 427–439. [Google Scholar] [CrossRef]
- Khatri, N.; Mudgil, Y. Hypothesis: NDL proteins function in stress responses by regulating microtubule organization. Front. Plant Sci. 2015, 6, 6115. [Google Scholar] [CrossRef] [PubMed]
- Yazdanpanah, F.; Maurino, V.G.; Mettler-Altmann, T.; Buijs, G.; Bailly, M.; Jashni, M.K.; Willems, L.; Sergeeva, L.I.; Rajjou, L.; Hilhorst, H.W.M.; et al. NADP-MALIC ENZYME 1 affects germination after seed storage in Arabidopsis thaliana. Plant Cell Physiol. 2018, 60, 318–328. [Google Scholar] [CrossRef]
- Arias, C.L.; Pavlovic, T.; Torcolese, G.; Badia, M.B.; Gismondi, M.; Maurino, V.G.; Andreo, C.S.; Drincovich, M.F.; Wheeler, M.C.G.; Saigo, M. NADP-Dependent Malic Enzyme 1 Participates in the Abscisic Acid Response in Arabidopsis thaliana. Front. Plant Cell. 2018, 9, 1637. [Google Scholar] [CrossRef] [Green Version]
- Larson, E.R.; Van Zelm, E.; Roux, C.; Marion-Poll, A.; Blatt, M.R. Clathrin Heavy Chain Subunits Coordinate Endo- and Exocytic Traffic and Affect Stomatal Movement1[CC-BY]. Plant Physiol. 2017, 175, 708–720. [Google Scholar]
- Wu, G.; Liu, S.; Zhao, Y.; Wang, W.; Kong, Z.; Tang, D. ENHANCED DISEASE RESISTANCE4 associates with CLATHRIN HEAVY CHAIN 2 and modulates plant immunity by regulating relocation of EDR1 in Arabidopsis. Plant Cell 2015, 27, 857–873. [Google Scholar] [CrossRef]
- Komatsu, S.; Ahsan, N. Soybean proteomics and its application to functional analysis. J. Proteom. 2009, 72, 325–336. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2018, 221, 32–49. [Google Scholar] [CrossRef]
- Kanwal, P.; Gupta, S.; Arora, S.; Kumar, A. Identification of genes involved in carbon metabolism from Eleusine coracana (L.) for understanding their light-mediated entrainment and regulation. Plant Cell Rep. 2014, 33, 1403–1411. [Google Scholar] [CrossRef]
- Zhang, D.-Y.; Wan, Y.; Hao, J.-Y.; Hu, R.-Z.; Chen, C.; Yao, X.-H.; Zhao, W.-G.; Liu, Z.-Y.; Li, L. Evaluation of the alkaloid, polyphenols, and antioxidant contents of various mulberry cultivars from different planting areas in eastern China. Ind. Crop. Prod. 2018, 122, 298–307. [Google Scholar] [CrossRef]
- Papoulias, E.; Siomos, A.S.; Koukounaras, A.; Gerasopoulos, D.; Kazakis, E. Effects of Genetic, Pre- and Post-Harvest Factors on Phenolic Content and Antioxidant Capacity of White Asparagus Spears. Int. J. Mol. Sci. 2009, 10, 5370–5380. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wu, X.; Huang, L. Correlation between Antioxidant Activities and Phenolic Contents of Radix Angelicae sinensis (Danggui). Molecules 2009, 14, 5349–5361. [Google Scholar] [CrossRef]
- Ben Farhat, M.; Landoulsi, A.; Chaouch-Hamada, R.; Sotomayor, J.A.; Jordán, M.J. Characterization and quantification of phenolic compounds and antioxidant properties of Salvia species growing in different habitats. Ind. Crop. Prod. 2013, 49, 904–914. [Google Scholar] [CrossRef]
- Djouahri, A.; Boualem, S.; Boudarene, L.; Baaliouamer, A. Geographic’s variation impact on chemical composition, antioxidant and anti-inflammatory activities of essential oils from wood and leaves of Tetraclinis articulata (Vahl) Masters. Ind. Crop. Prod. 2015, 63, 138–146. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Q.; Zhou, W.; Li, P.; Alolga, R.N.; Qi, L.-W.; Yin, X. A comparative proteomic characterization and nutritional assessment of naturally- and artificially-cultivated Cordyceps sinensis. J. Proteom. 2018, 181, 24–35. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially Modified Protein Abundance Index (emPAI) for Estimation of Absolute Protein Amount in Proteomics by the Number of Sequenced Peptides per Protein. Mol. Cell. Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [Green Version]
- Jelled, A.; Fernandes, Â.; Barros, L.; Chahdoura, H.; Achour, L.; Ferreira, I.C.; Ben Cheikh, H. Chemical and antioxidant parameters of dried forms of ginger rhizomes. Ind. Crop. Prod. 2015, 77, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Hong, Y.; Han, Y.; Wang, Y.; Xia, L. Chemical characterization and antioxidant activities comparison in fresh, dried, stir-frying and carbonized ginger. J. Chromatogr. B 2016, 1011, 223–232. [Google Scholar] [CrossRef]
Sample Availability: Samples of the X. aethiopica fruits used can be provided upon request from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Description | PSM (Nigeria) | PSM (Ghana) | Ratio (Nigeria/Ghana) | p-Value | ||
|---|---|---|---|---|---|---|---|
| N1 | N2 | G1 | G2 | ||||
| I1MQ89 | Uncharacterized protein | 51 | 38 | 1 | 1 | 44.500 | 0.022 |
| I1K4W1 | Uncharacterized protein | 25 | 32 | 1 | 1 | 28.500 | 0.016 |
| I1MPI1 | Uncharacterized protein | 31 | 22 | 1 | 1 | 26.500 | 0.030 |
| I1MT10 | Uncharacterized protein | 25 | 23 | 1 | 1 | 24.000 | 0.002 |
| D6C500 | HSP90-2 | 26 | 19 | 1 | 1 | 22.500 | 0.025 |
| I1JT28 | Tubulin beta chain | 15 | 11 | 1 | 1 | 13.000 | 0.027 |
| I1KZK0 | Uncharacterized protein | 17 | 14 | 1 | 2 | 10.333 | 0.013 |
| I1K5Y5 | Malic enzyme | 8 | 10 | 1 | 1 | 9.000 | 0.015 |
| I1LMS9 | Clathrin heavy chain | 8 | 10 | 1 | 1 | 9.000 | 0.015 |
| I1JY29 | Uncharacterized protein | 9 | 7 | 1 | 1 | 8.000 | 0.020 |
| I1JWK3 | Uncharacterized protein | 9 | 6 | 1 | 1 | 7.500 | 0.049 |
| I1M3L8 | Uncharacterized protein | 16 | 14 | 3 | 1 | 7.500 | 0.012 |
| I1JPX8 | Fructose-bisphosphate aldolase | 8 | 7 | 1 | 1 | 7.500 | 0.006 |
| A0A0R0GRY6 | Uncharacterized protein | 11 | 10 | 2 | 1 | 7.000 | 0.006 |
| A0A0R0GJT8 | Uncharacterized protein | 6 | 8 | 1 | 1 | 7.000 | 0.027 |
| I1NDI4 | Uncharacterized protein | 8 | 6 | 1 | 1 | 7.000 | 0.027 |
| A0A0R0FJR0 | 40S ribosomal protein SA | 8 | 6 | 1 | 1 | 7.000 | 0.027 |
| I1NAI7 | Uncharacterized protein | 39 | 29 | 9 | 1 | 6.800 | 0.045 |
| I1KEN4 | Uncharacterized protein | 16 | 14 | 1 | 4 | 6.000 | 0.020 |
| I1MJU7 | Uncharacterized protein | 5 | 6 | 1 | 1 | 5.500 | 0.012 |
| I1KU21 | Uncharacterized protein | 10 | 12 | 3 | 1 | 5.500 | 0.024 |
| I1M5G8 | Malic-enzyme | 6 | 5 | 1 | 1 | 5.500 | 0.012 |
| I1L0G9 | Tubulin beta chain | 20 | 15 | 3 | 4 | 5.000 | 0.032 |
| A0A0R4J4C3 | Elongation factor 1-alpha | 17 | 20 | 7 | 1 | 4.625 | 0.050 |
| C6T1V2 | Uncharacterized protein | 5 | 4 | 1 | 1 | 4.500 | 0.020 |
| I1MQS6 | EF1Bgamma class glutathione S-transferase | 4 | 5 | 1 | 1 | 4.500 | 0.020 |
| I1KPH6 | 6-phosphogluconate dehydrogenase | 5 | 4 | 1 | 1 | 4.500 | 0.020 |
| A0A0R0F2E8 | Uncharacterized protein | 7 | 6 | 1 | 2 | 4.333 | 0.019 |
| I1KH24 | Phospholipase D | 5 | 6 | 1 | 2 | 3.667 | 0.030 |
| K7KYB1 | Aconitate hydratase | 6 | 5 | 2 | 1 | 3.667 | 0.030 |
| I1N0W4 | Xyloglucan endotransglucosylase/hydrolase | 3 | 4 | 1 | 1 | 3.500 | 0.038 |
| I1JDR2 | Uncharacterized protein | 4 | 3 | 1 | 1 | 3.500 | 0.038 |
| A0A0R0FB78 | Uncharacterized protein | 4 | 3 | 1 | 1 | 3.500 | 0.038 |
| I1MC13 | Uncharacterized protein | 4 | 3 | 1 | 1 | 3.500 | 0.038 |
| I1KZJ0 | Uncharacterized protein | 3 | 4 | 1 | 1 | 3.500 | 0.038 |
| I1K672 | Uncharacterized protein | 4 | 3 | 1 | 1 | 3.500 | 0.038 |
| I1JCQ7 | Succinate dehydrogenase [ubiquinone] | 3 | 4 | 1 | 1 | 3.500 | 0.038 |
| I1JXI9 | Proteasome subunit beta | 4 | 3 | 1 | 1 | 3.500 | 0.038 |
| I1JJM3 | Fructose-bisphosphate aldolase | 7 | 7 | 1 | 3 | 3.500 | 0.038 |
| I1L314 | Heat shock protein 90-1 | 36 | 27 | 8 | 11 | 3.316 | 0.043 |
| I1LNM2 | NADH dehydrogenase subunit 9 | 4 | 4 | 1 | 2 | 2.667 | 0.038 |
| A0A0R0HEQ3 | Histone H2B | 4 | 4 | 2 | 1 | 2.667 | 0.038 |
| C6TGU2 | Proteasome subunit alpha type | 6 | 7 | 2 | 3 | 2.600 | 0.030 |
| O48548 | Aspartate aminotransferase | 5 | 4 | 2 | 2 | 2.250 | 0.038 |
| I1MC31 | Uncharacterized protein | 19 | 18 | 10 | 10 | 1.850 | 0.003 |
| A0A0R0HEM1 | Uncharacterized protein | 1 | 1 | 12 | 14 | 0.077 | 0.007 |
| I1L3K7 | Uncharacterized protein | 1 | 1 | 13 | 16 | 0.069 | 0.012 |
| Element | Ghana | Nigeria | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| G1 | G2 | G3 | Average | N1 | N2 | N3 | Average | ||
| Zn | 0.052 ± 0.001 | 0.049 ± 0.016 | 0.037 ± 0.07 | 0.046 | 0.045 ± 0.10 | 0.032 ± 0.12 | 0.040 ± 0.09 | 0.039 | 0.304 |
| P | 1.197 ± 0.03 | 1.190 ± 0.06 | 0.713 ± 0.01 | 1.034 | 1.286 ± 0.21 | 0.595 ± 0.18 | 0.920 ± 0.13 | 0.934 | 0.717 |
| Fe | 1.721 ± 0.20 | 1.248 ± 0.08 | 0.771 ± 0.11 | 1.246 | 0.785 ± 0.11 | 0.452 ± 0.23 | 0.847 ± 0.31 | 0.695 | 0.140 |
| Mg | 10.505 ± 0.05 | 11.142 ± 0.01 | 4.835 ± 0.1 | 8.827 | 13.025 ± 0.19 | 5.318 ± 0.24 | 8.039 ± 0.14 | 8.794 | 0.992 |
| Ca | 3.270 ± 0.16 | 3.201 ± 0.20 | 1.839 ± 0.09 | 2.77 | 3.023 ± 0.35 | 1.772 ± 0.24 | 2.740 ± 0.28 | 2.512 | 0.689 |
| Na | 0.024 ± 0.03 | 0.222 ± 0.01 | 0.251 ± 0.07 | 0.165 | 0.326 ± 0.76 | 0.479 ± 0.81 | 0.499 ± 0.98 | 0.435 | 0.040 * |
| K | 8.149 ± 0.09 | 6.760 ± 0.06 | 4.049 ± 0.02 | 6.319 | 4.878 ± 0.78 | 2.168 ± 0.89 | 3.772 ± 0.68 | 3.606 | 0.132 |
| Samples | Total Phenolic and Flavonoid Content | Antioxidant Activities | ||||
|---|---|---|---|---|---|---|
| TPC (mg GAE/g) | TFC (mg CE/g) | ABTS (mg/g db) | CUPRIC (mg/g db) | DPPH (mg/g db) | FRAP (mg/g db) | |
| G1 | 58.94 ± 8.54 a | 67.73 ± 3.39 ab | 18.15 ± 5.06 e | 32.44 ± 16.70 c | 89.20 ± 0.40 c | 66.84 ± 4.46 c |
| G2 | 67.34 ± 7.47 a | 77.39 ± 0.77 ab | 20.33 ± 3.95 e | 26.72 ± 3.36 c | 82.39 ± 0.33 d | 62.11 ± 3.77 cd |
| G3 | 49.61 ± 0.49 a | 65.46 ± 2.62 b | 36.70 ± 6.81 d | 36.96 ± 10.37 c | 83.73 ± 0.48 d | 55.57 ±10.08 d |
| N1 | 84.85 ± 5.32 ab | 90.03 ± 4.21 a | 46.40 ± 3.63 a | 60.35 ± 2.67 ab | 107.07 ± 0.81 a | 83.50 ± 4.09 b |
| N2 | 81.92 ± 7.96 bc | 95.59 ± 2.89 a | 39.53 ± 1.37 c | 79.89 ± 5.93 a | 99.53 ± 1.06 b | 87.27 ± 0.42 ab |
| N3 | 79.65 ± 5.08 c | 92.5 ± 3.9 a | 42.89 ± 4.04 b | 80.20 ± 6.27 a | 104.105 ± 0.82 a | 89.70 ± 3.09 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, X.; Chávez León, M.A.S.C.; Osae, R.; Linus, L.O.; Qi, L.-W.; Alolga, R.N. Xylopia aethiopica Seeds from Two Countries in West Africa Exhibit Differences in Their Proteomes, Mineral Content and Bioactive Phytochemical Composition. Molecules 2019, 24, 1979. https://doi.org/10.3390/molecules24101979
Yin X, Chávez León MASC, Osae R, Linus LO, Qi L-W, Alolga RN. Xylopia aethiopica Seeds from Two Countries in West Africa Exhibit Differences in Their Proteomes, Mineral Content and Bioactive Phytochemical Composition. Molecules. 2019; 24(10):1979. https://doi.org/10.3390/molecules24101979
Chicago/Turabian StyleYin, Xiaojian, María A.S.C. Chávez León, Richard Osae, Loveth O. Linus, Lian-Wen Qi, and Raphael N. Alolga. 2019. "Xylopia aethiopica Seeds from Two Countries in West Africa Exhibit Differences in Their Proteomes, Mineral Content and Bioactive Phytochemical Composition" Molecules 24, no. 10: 1979. https://doi.org/10.3390/molecules24101979