Antifungal Activities of 4″,6″-Disubstituted Amphiphilic Kanamycins
by
 , ,
, ,
Madher N. Alfindee
1,† ,
,
Yagya P. Subedi
1,†,
Michelle M. Grilley
2,
Jon Y. Takemoto
2 and
Cheng-Wei T. Chang
1,* 1
Department of Chemistry and Biochemistry, Utah State University, 0300 Old Main Hill, Logan, UT 84322-0300, USA
2
Department of Biology, Utah State University, 5305 Old Main Hill, Logan, UT 84322-5305, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Molecules 2019, 24(10), 1882; https://doi.org/10.3390/molecules24101882
Submission received: 16 April 2019
/
Revised: 10 May 2019
/
Accepted: 14 May 2019
/
Published: 16 May 2019
(This article belongs to the Special Issue Recent Development on the New Applications of Aminoglycosides)
Abstract
:Amphiphilic kanamycins derived from the classic antibiotic kanamycin have attracted interest due to their novel bioactivities beyond inhibition of bacteria. In this study, the recently described 4″,6″-diaryl amphiphilic kanamycins reported as inhibitors of connexin were examined for their antifungal activities. Nearly all 4″,6″-diaryl amphiphilic kanamycins tested had antifungal activities comparable to those of 4″,6″-dialkyl amphiphilic kanamycins, reported previously against several fungal strains. The minimal growth inhibitory concentrations (MICs) correlated with the degree of amphiphilicity (cLogD) of the di-substituted amphiphilic kanamycins. Using the fluorogenic dyes, SYTOXTM Green and propidium iodide, the most active compounds at the corresponding MICs or at 2×MICs caused biphasic dye fluorescence increases over time with intact cells. Further lowering the concentrations to half MICs caused first-order dye fluorescence increases. Interestingly, 4×MIC or 8×MIC levels resulted in fluorescence suppression that did not correlate with the MIC and plasma membrane permeabilization. The results show that 4″,6″-diaryl amphiphilic kanamycins are antifungal and that amphiphilicity parameter cLogD is useful for the design of the most membrane-active versions. A cautionary limitation of fluorescence suppression was revealed when using fluorogenic dyes to measure cell-permeation mechanisms with these antifungals at high concentrations. Finally, 4″,6″-diaryl amphiphilic kanamycins elevate the production of cellular reactive oxygen species as other reported amphiphilic kanamycins.
1. Introduction
Although commonly unappreciated, fungal diseases cause tremendous economic loss and health impacts globally. Annually, more than a billion people acquire fungal infections. The death rate from fungal diseases is equal to that of tuberculosis and malaria combined, with approximately 10% of those deaths due to cryptococcal meningitis [1,2]. Immunosuppressed individuals, such as those infected with HIV or undergoing treatment for cancer or organ transplants, are especially susceptible to invasive fungal infections. Current treatment options for fungal infections include polyene-based compounds (e.g., amphotericin B) [3,4,5], cytosine-based compounds (e.g., flucytosine) [5,6], and azole-based compounds (e.g., itraconazole and fluconazole) [5,7,8]. However, as exemplified in the recent outbreak of Candida auris [9], drug resistance is a growing problem. Further research to develop new and effective treatments for fungal diseases is urgent.
Kanamycin belongs to a class of antibacterial compounds known as aminoglycosides that are active against both Gram-negative (G−) and Gram-positive (G+) bacteria, albeit its clinical use is limited due to the emergence of bacterial resistance (Figure 1) [10]. To overcome the problem of bacterial resistance, extensive research has been devoted to structural modifications of aminoglycosides that lead to the discovery of amphiphilic aminoglycosides [11,12]. In contrast to antibacterial kanamycin that is inactive against fungi, some of the amphiphilic kanamycins (AKs) were found to be active against a wide range of fungal strains [13,14] and concomitantly not active against bacteria. Two of the antifungal AKs produced good specific antifungal activity: FG08, which contains an octyl group (C8) chain attached at the 4″ position of kanamycin analog via an ether linkage [15] and K20, which has an octanesulfonyl group at the 6″ position (Figure 1) [16].
AKs are known to show their antimicrobial activity by increasing the membrane permeability of microorganisms [17,18,19,20,21]. Fluorogenic dyes, such as SYTOXTM green and propidium iodide (PI), are commonly employed for the study of membrane permeabilization (Figure 2). SYTOXTM green is non-fluorescent and cannot penetrate the plasma membrane of intact organisms. However, in the presence of agents that compromise membrane integrity, SYTOXTM green enters the cytoplasm, binds to nucleic acids, and emits fluorescence. Propidium iodide (PI) has similar properties and is widely used for evaluating membrane permeabilization of substances in fungi and bacteria. We recently reported the synthesis of 4″,6″-diaryl AKs (compounds 1–8) as connexin inhibitors [22,23] (Figure 3). Since these are structurally similar to the antifungal 4″,6″-dialkyl AKs (compounds 9–13), we decided to investigate their antifungal activities and to conduct mode of action studies of both groups of compounds using fluorogenic dyes.
2. Results and Discussion
2.1. Fungal Growth Inhibition by 4″,6″-Disubstituted AKs
Thirteen disubstituted AKs were examined for growth inhibition capabilities against a panel of fungi that included yeasts and the filamentous fungus Fusarium graminearum (Table 1). The 4″,6″- diaryl compounds 6, 7, and 8 were strongly inhibitory toward F. graminearum (minimal growth inhibitory concentrations, MICs, of 2–16 µg/mL) and compounds 1–5 had low (MICs, >32 µg/mL) to moderate (MICs, 16–32 µg/mL) inhibitory activities. Likewise, except with compounds 1 and 2, Cryptococcus neoformans H99 and C. neoformans VR-54 were highly susceptible to the diaryl compounds (MICs, 2–16 µg/mL), as was Rhodotorula pilimanae (ATCC 26423) (MICs, 2–32 µg/mL). In contrast, except with compounds 4 and 6, Candida albicans 64124 (azole-resistant) and C. albicans MYA2876 (azole sensitive) were not susceptible to the diaryl compounds (MICs, >32). The 4″,6″-dialkyl compounds, except 13, displayed strong inhibitory activities against the C. neoformans strains (MICs, 2–16 µg/mL), and moderate activities against the Candida strains 64124 and MYA2876 (MICs, 8–128 µg/mL). Compound 13, with a long linear alkyl chain (C16), had no antifungal activity (MICs, equal to or >256). Overall, except for compounds 1, 2 and 13, the 4″,6″-disubstituted AKs (10 of 13) were strongly inhibitory to F. graminearum, C. neoformans and R. pilimanae (ATCC 26423), but less so or moderately with the C. albicans strains.
2.2. Analysis of Correlation Between MIC and cLogD
Structurally diverse AKs have been synthesized for the purpose of elucidating structure-activity relationships (SAR) [7,20]. These AKs carry variations of hydrophobic moieties or linkages of the hydrophobic moieties to the kanamycin core. Several factors that may contribute to the differences of antimicrobial activity, such as the chain length of the hydrophobic moiety and the linkage, were deduced primarily on the revealed MICs of these AKs. Despite these labor-intensive efforts, contradictory SARs were noted and these factors seemed to escalate the complexity in understanding the nature of the antimicrobial selectivity of AKs. Thus, we explored the use of a water/1-octanol distribution coefficient (cLogD) as a simplified means to evaluate the amphiphilicity of the disubstituted AK compounds and possible correlation with antifungal activity. The cLogD was calculated using Marvin Sketch (version 18.19) keeping a 0.1 molar concentration of Na+, K+, and Cl− ions (Table 1). When plotting cLogD (x-axis) vs. MICs (y-axis) for all tested fungi (z-axis), a clearer SAR was observed (Figure 4). Compounds with cLogD values between –9.5 and –6.4 had the lowest MIC values against F. graminearum, C. neoformans (two strains) and R. pilimanae. cLogD values above (e.g., compound 13) or below (e.g., compounds 1 and 2) this range corresponded to the highest MIC values. Such a trend was not observed with C. albicans (two strains) because of fluctuating MIC values. This analysis suggests that it is possible to use cLogD as a guideline for designing antifungal AK of the same class, which can drastically reduce the synthetic burden.
2.3. Plasma Membrane Permeabilization by 4″,6″-Disubstituted Kanamycins
Plasma membrane permeabilization studies were conducted by light microscopy using the dyes SYTOXTM green with C. neoformans H99 treated with diaryl compound 7 or the dialkyl compounds (11 and 13) at 1×MIC (Figure 5). Triton X-100 (1%), a non-fungal targeting agent, but known to cause membrane permeabilization of mammalian cells, was used for comparison. As expected, most fungal cells treated with compounds 7 and 11 showed fluorescence emitted from SYTOXTM green. In contrast, almost no cells emitted fluorescence when treated with compound 13 (256 μg/mL). A few fungal cells emitted fluorescence when treated with Triton X-100, suggesting that this agent at 1% concentration causes a small degree of membrane permeabilization.
We previously reported that fungi treated with active AKs with hydrophobic groups attached at the 6′ position displayed a fast increase of dye fluorescence observed in a time-dependent fashion [24]. To determine if the disubstituted AKs behave similarly, time-dependent kinetic membrane permeabilization experiments were performed using C. neoformans H99 fungi at 1×MIC of the AKs. The relative fluorescence unit (RFU) was monitored every 3 min for 4 h.
The most growth inhibitory 4″,6″-disubstituted AKs (compounds 5, 6, 7, and 8) caused the highest levels of SYTOXTM green fluorescence. For most of the tested compounds, the kinetics of the fluorescence signals revealed biphasic fluorescence increases over time: A fast membrane permeabilization within the first 15 min that quickly levels to various RFUs (Figure 6). The leveled units, in general, follow similar orders in all experiments using SYTOXTM green or PI. The less active AKs (1, 2 and 3) showed profiles similar to Triton X-100. Despite the overall similarities of the biphasic fluorescence profiles, differences were observed between the profiles. First, different total RFU levels (at 4 h) were achieved with different AKs and with the two dyes despite using the same fungal cell densities, fluorogenic dye concentrations, and compounds at 1×MIC. Second, in three experiments (Figure 6B–D), several AKs caused total RFU levels lower than controls with no AK. Third, certain compounds displayed different RFU kinetic profiles using SYTOXTM green vs. PI. For example, the RFU kinetic profiles of compounds 2 and 4 appeared flat or linear using PI (Figure 6B) but biphasic using SYTOXTM green (Figure 6A). For compounds 5 and 6, different RFU profiles were obtained from the experiments using SYTOXTM green vs. PI. For compound 12, the RFU profiles were linear using SYTOXTM green but biphasic using PI (Figure 6C,D). Because the only variable parameters in these experiments were the individual AKs and their MICs, it was speculated that the fluorescence properties of the dyes reflected in the RFU and kinetic profiles were directly affected by individual AKs and their concentrations. To explore this hypothesis, we carried out further experiments using 2-fold adjusted concentrations of the MICs of the AKs.
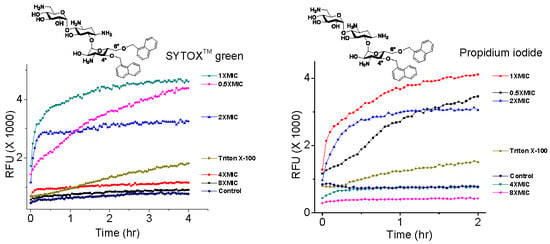
Fluorescence kinetics and levels with selected 4″,6″-disubstituted AKs at various concentrations were measured with C. neoformans H99 using SYTOXTM green and PI (Figure 7). It was observed that the AK concentration significantly influenced the degree of fluorescence independently of the MIC and membrane permeabilization capabilities. For example, compound 7 showed linear fluorescence kinetics at 0.5×MIC, biphasic kinetics at 1×MIC, biphasic kinetics but leveled at lower RFUs at 2×MIC, and suppression at 4×MIC and 8×MIC (Figure 7A,B). The same trend was observed when using PI. For compound 10, similar kinetic profiles were obtained at 1×MIC but not when 2×MIC was employed (Figure 7C,D). The kinetic RFU profiles of compound 11 behaved like those of compound 7 (Figure 7E,F). These data support our speculation that the fluorescence of dyes can be affected by the concentration of the tested AK compounds.
Images of fungi treated with compound 11 provide further evidence for the direct effect of the AK concentration on fluorescence (Figure 8). Cells treated with compound 11 and PI for 2 h emit fluorescence at the 0.5× and 1×MIC×MICs but little or no fluorescence at 8×MIC. A similar result was obtained with SYTOXTM green.
These results indicate that there is an optimal ratio of AK vs. fluorogenic dye when measuring plasma membrane permeabilization by AK compounds. By considering the MICs that gave rise to the maximum leveled RFU, the concentration fell in the region around 5 µM of AK and 0.01 µM of SYTOXTM green or 0.4 µM of PI. Judging from the structures of the AKs, SYTOXTM green and PI (Figure 2), it is likely that ionic or hydrogen bonding contributed to the AK inhibiting effect toward these fluorogenic dyes. Therefore, caution needs to be taken when evaluating the cellular effect of AKs or other classes of compounds using fluorogenic probes. Finally, since high RFUs were observed with the use of AK levels at <1×MIC, possibilities are opened for using such low AK concentrations to detect fungi without significant fungicidal effect.
2.4. Effect of 4″,6″-Disubstituted Kanamycins on the Production of Reactive Oxygen Species
It has been reported that a wide range of antimicrobials of different classes, albeit having various modes of action, will have a common effect of increased oxidative stress by promoting the production of reactive oxygen species (ROS), which can lead to cell death. We have also demonstrated that 6′-substituted AKs exert this common effect. Thus, several selective 4″,6″-disubstituted kanamycins—compounds 4, 7 and 11—were subjected to the investigation of induced ROS production. The experiments of ROS production were conducted using 1,1′-(hexane-1,6-diyl)bis(3-decyl-4,9-dioxo-4,9-dihydro-1H-naphtho [2,3-d][1,2,3]triazol-3-ium triflate (14) (Figure 9) [25,26], known to produce ROS, as the positive control. Compound 4 showed almost no ROS generation as compared to control, while compound 11 showed moderate ROS generation, and compound 7 showed a much higher level of ROS generation (Figure 9).
Compound 7 contains a naphthalene motif that can better stabilize radicals. Hence, it explains why compound 7 promoted ROS production to the level of the positive control, compound 14. Compound 11, with the linear alkyl chain, exerted a moderate level of ROS production similar to the 6′-alkylated AKs, as we have noted previously. Fluoro-substituted molecules, such as in the case of compound 4, are noted to inhibit the formation of radicals [27]. Therefore, it is not surprising that compound 4 displayed almost no elevation of ROS production, as compared to the control (blank). However, compound 4 showed similar antifungal activities as compounds 7 and 11. The ROS measurement was conducted following 3 h of incubation of fungi with the AKs, a much shorter time compared to the determination of the MIC, which often requires 48 h of incubation. Thus, compound 4 may exert fast membrane permeabilization, as observed with the fast RFU increases. However, the presence of fluoro-substituents deters the formation of ROS, resulting in the lower ROS generation but similar antifungal activity. Hence, it is likely that the actual mode of the antifungal mode of action is a combination of two main factors: (1) The rate of membrane permeabilization and (2) the rate of ROS production.
3. Materials and Methods
Procedure for the antifungal activity. The fungal MIC test was carried out similar to previously reported protocols [28]. In brief, fungi were grown in RPMI 1640 medium for 48 h at 37 °C. Cells, or spores in the case of F. graminearum, were counted and diluted to 4 × 104 cells/mL in the growth medium. Fungi were added to the compounds dissolved in the growth medium, maintaining final concentrations of the compounds from 256 to 0.125 µg/mL. Fungi, treated with compounds, were incubated at 37 °C for 48 h to see the inhibitory effect of the compounds. The fungal MIC testing was done in triplicate trials.
Procedure for kinetic cell permeabilization using 1×MIC of compounds. The fungi cell membrane permeabilization study was performed in a 96-well cell culture plate. C. neoformans H99 was grown in PDB medium at 37 °C for 48 h. From the growth, 4 mL of fungi were taken and washed with water twice and resuspended in 1 mL of water. Fungi cells were counted, and cells were diluted to 4 × 106 cells/mL as the final confluence. Then, compounds and dye were added to the fungi, maintaining the final concentration of compounds to 1×MIC, 0.01 µM for SYTOXTM green and 0.4 µg/mL for PI. The control cells were treated with dye only. The fluorescence intensity was measured in every 3 min for a period of four hours at 28 °C with an excitation/emission wavelength of 480Ex/525Em nm for SYTOXTM green and 537Ex/617Em nm for PI using the Cytation 5 imaging reader. The images of compounds 7, 11, 13, and Triton X-100 were taken after 60 min of incubation using the green channel and bright field filter set in the Cytation 5 imaging reader and Olympus IX 71.
Procedure for kinetic cell permeabilization using multiple MIC of the compounds. This assay was performed similarly to the fungi cell membrane permeabilization study using 0.125×MIC to 8×MIC of compounds 2, 7, 10, and 11. The fluorescence intensity was measured every 3 min for a period of two or four hours. The images of the representative compound 11 were taken after the 60 min of incubation using the green channel for SYTOXTM green and after 120 min using the red channel for PI along with the bright field filter set.
Procedure for reactive oxygen species (ROS) generation study. The effect of the amphiphilic kanamycin to generate ROS species in the fungal cells was studied in the C. neoformans H99 strain, using the previously reported ROS-generating amphiphilic antifungal compound 14, as the positive control [25,26]. The fungal cells were grown in PDB medium at 28 °C for 48 h with gentle shaking. The cells were washed with water twice before using for the assay by centrifugation at 10,000 rpm for 2 min. A volume of 0.5 mL of cells in water (final cell confluence, 4 × 107 cells/mL) was incubated with 1×MIC of the compounds for 3 h at 37 °C. Then, the cells were washed twice with 0.5 mL water and finally resuspended in 0.5 mL water. The cells were then incubated for another 30 min with 2′,7′-dichlorodihydrofluorescein diacetate (DCF-DA) dye (25 mM final concentration) at 37 °C. After incubation, the cells were washed twice with the same volume of water and resuspended in water. The fluorescence of the cells was measured in the Cytation 5 imaging reader with an excitation and emission wavelength of 485 nm and 525 nm, respectively. The experiment was performed at least in triplicate.
4. Conclusions
We have examined the antifungal activities of 4″,6″-diaryl kanamycins, as well as the previously described 4″,6″-dialkyl kanamycins. These two classes of AKs show similar antifungal profiles against a panel of fungi despite having different hydrophobic groups. The amphiphilicity parameter cLogD can be useful for the structural design when no obvious SAR can be deduced. Using the fluorogenic dye SYTOXTM green—4″,6″-diaryl kanamycins were shown to exert their antifungal activities via the increase of plasma membrane permeability. It was shown, however, that the fluorescent properties of the commonly used fluorogenic dyes SYTOXTM green and PI are drastically affected by the concentrations of the AKs. Therefore, to acquire meaningful results in such approaches, prudent practice is needed to establish the optimal ratio of AKs to fluorogenic dyes for the experiments. Finally, from a combination of studying kinetic membrane permeabilization, ROS production, cLogD and fungal growth MICs, we conclude that the mode of antifungal action of the 4″,6″-disubstituted AKs is a combination of plasma membrane permeabilization and oxidative stress.
Supplementary Materials
Graphs of the kinetic membrane permeabilization of C. neoformans with 1×MIC of compounds with the standard deviation and graphs of the kinetic membrane permeabilization of C. neoformans with multiple MICs of compounds with standard deviation are available online.
Author Contributions
M.N.A. and Y.P.S. performed the experiments in equal contribution. C.-W.T.C. designed the experiments and instructed the data analysis. J.Y.T. provides general guidance for fungal strains to be tested and instruction on the details of fungal membrane compositions. M.M.G. provides specific technical instruction for growing and assaying fungi.
Funding
CTC was funded by USU RC grant.
Acknowledgments
We thank the support from USU RC grant. We are also grateful to ChemAxon for providing free academic license.
Conflicts of Interest
No conflicts of interest declared.
References
- Brown, G.D.; Denning, D.W.; Levitz, S.M. Tackling human fungal infections. Science 2012, 336, 647. [Google Scholar] [CrossRef]
- Armstrong-James, D.; Meintjes, G.; Brown, G.D. A neglected epidemic: Fungal infections in HIV/AIDS. Trends Microbiol. 2014, 22, 120–127. [Google Scholar] [CrossRef]
- Dutcher, J.D. The discovery and development of amphotericin b. Chest 1968, 54, 296–298. [Google Scholar] [CrossRef]
- Donovick, R.; Gold, W.; Pagano, J.F.; Stout, H.A. Amphotericins a and b, antifungal antibiotics produced by a streptomycete. I. In vitro studies. Antibiot. Annu. 1955, 3, 579–586. [Google Scholar]
- Lyman, C.A.; Walsh, T.J. Systemically administered antifungal agents. Drugs 1992, 44, 9–35. [Google Scholar] [CrossRef]
- Montgomery, J.A.; Hewson, K. Synthesis of potential anticancer agents. X. 2-fluoroadenosine1. J. Am. Chem. Soc. 1957, 79, 4559. [Google Scholar] [CrossRef]
- Shrestha, S.K.; Fosso, M.Y.; Green, K.D.; Garneau-Tsodikova, S. Amphiphilic tobramycin analogues as antibacterial and antifungal agents. Antimicrob Agents Chemother. 2015, 59, 4861–4869. [Google Scholar] [CrossRef] [PubMed]
- Woolley, D.W. Some biological effects produced by benzimidazole and their reversal by purines. J. Biol. Chem. 1944, 152, 225–232. [Google Scholar]
- Tracking Candida auris | Candida auris | Fungal Diseases | CDC. Available online: https://www.cdc.gov/fungal/candida-auris/tracking-c-auris.html (accessed on 13 April 2019).
- Chandrika, N.T.; Garneau-Tsodikova, S. Comprehensive review of chemical strategies for the preparation of new aminoglycosides and their biological activities. Chem. Soc. Rev. 2018, 47, 1189–1249. [Google Scholar] [CrossRef]
- Zhang, J.; Chiang, F.-I.; Wu, L.; Czyryca, P.G.; Li, D.; Chang, C.-W.T. Surprising alteration of antibacterial activity of 5′′-modified neomycin against resistant bacteria. J. Med. Chem. 2008, 51, 7563–7573. [Google Scholar] [CrossRef]
- Bera, S.; Zhanel, G.G.; Schweizer, F. Antibacterial activities of aminoglycoside antibiotics-derived cationic amphiphiles. Polyol-modified neomycin b-, kanamycin a-, amikacin-, and neamine-based amphiphiles with potent broad spectrum antibacterial activity. J. Med. Chem. 2010, 53, 3626–3631. [Google Scholar] [CrossRef]
- Chang, C.-W.T.; Takemoto, J.Y. Antifungal amphiphilic aminoglycosides. MedChemComm 2014, 5, 1048–1057. [Google Scholar] [CrossRef]
- Subedi, Y.P.; AlFindee, M.N.; Takemoto, J.Y.; Chang, C.-W.T. Antifungal amphiphilic kanamycins: New life for an old drug. MedChemComm 2018, 9, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-W.T.; Fosso, M.; Kawasaki, Y.; Shrestha, S.; Bensaci, M.F.; Wang, J.; Evans, C.K.; Takemoto, J.Y. Antibacterial to antifungal conversion of neamine aminoglycosides through alkyl modification. Strategy for reviving old drugs into agrofungicides. J. Antibiot. 2010, 63, 667–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.-W.T.; Takemoto, J. Aminoglycosides: Synthesis and Use as Antifungals. U.S. Patent US 13/316,720; U.S. Patent US8865665 B2, 21 October 2014. [Google Scholar]
- Fosso, M.; AlFindee, M.N.; Zhang, Q.; Nziko, V.d.P.N.; Kawasaki, Y.; Shrestha, S.K.; Bearss, J.; Gregory, R.; Takemoto, J.Y.; Chang, C.-W.T. Structure–activity relationships for antibacterial to antifungal conversion of kanamycin to amphiphilic analogues. J. Org. Chem. 2015, 80, 4398–4411. [Google Scholar] [CrossRef] [PubMed]
- Fosso, M.Y.; Shrestha, S.K.; Green, K.D.; Garneau-Tsodikova, S. Synthesis and bioactivities of kanamycin b-derived cationic amphiphiles. J. Med. Chem. 2015, 58, 9124–9132. [Google Scholar] [CrossRef]
- Jaber, Q.Z.; Benhamou, R.I.; Herzog, I.M.; Ben Baruch, B.; Fridman, M. Cationic amphiphiles induce macromolecule denaturation and organelle decomposition in pathogenic yeast. Angew. Chem. Int. Ed. 2018, 57, 16391–16395. [Google Scholar] [CrossRef] [PubMed]
- Steinbuch, K.B.; Benhamou, R.I.; Levin, L.; Stein, R.; Fridman, M. Increased degree of unsaturation in the lipid of antifungal cationic amphiphiles facilitates selective fungal cell disruption. ACS Infect. Dis. 2018, 4, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Salta, J.; Benhamou, R.I.; Herzog, I.M.; Fridman, M. Tuning the effects of bacterial membrane permeability through photo-isomerization of antimicrobial cationic amphiphiles. Chem. Eur. J. 2017, 23, 12724–12728. [Google Scholar] [CrossRef] [PubMed]
- AlFindee, M.N.; Subedi, Y.P.; Fiori, M.C.; Krishnan, S.; Kjellgren, A.; Altenberg, G.A.; Chang, C.-W.T. Inhibition of connexin hemichannels by new amphiphilic aminoglycosides without antibiotic activity. ACS Med. Chem. Lett. 2018, 9, 697–701. [Google Scholar] [CrossRef]
- Kjellgren, A.; Fiori, M.C.; AlFindee, M.N.; Subedi, Y.P.; Krishnan, S.; Chang, C.-W.T.; Altenberg, G.A. Inhibition of connexion hemichannels by new aminoglycosides without antibiotic activity. Biophys. J. 2019, 116, 250a. [Google Scholar] [CrossRef]
- Subedi, Y.P.; Roberts, P.; Grilley, M.; Takemoto, J.Y.; Chang, C.-W.T. Development of fungal selective amphiphilic kanamycin: Cost-effective synthesis and use of fluorescent analogs for mode of action investigation. ACS Infect. Dis. 2019, 5, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, J.P.; Baker, C.; Kawasaki, Y.; Subedi, Y.P.; de Paul, N.N.V.; Takemoto, J.Y.; Chang, C.-W.T. Synthesis and bioactivity investigation of quinone-based dimeric cationic triazolium amphiphiles selective against resistant fungal and bacterial pathogens. Eur. J. Med. Chem. 2017, 126, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, J.P.; Subedi, Y.P.; Chen, L.; Chang, C.-W.T. A mode of action study of cationic anthraquinone analogs: A new class of highly potent anticancer agents. MedChemComm 2015, 6, 2012–2022. [Google Scholar] [CrossRef]
- Shrestha, A.; Jin Oh, H.; Kim, M.J.; Pun, N.T.; Magar, T.B.T.; Bist, G.; Choi, H.; Park, P.-H.; Lee, E.-S. Design, synthesis, and structure-activity relationship study of halogen containing 2-benzylidene-1-indanone derivatives for inhibition of lps-stimulated ros production in raw 264.7 macrophages. Eur. J. Med. Chem. 2017, 133, 121–138. [Google Scholar] [CrossRef] [PubMed]
- AlFindee, M.N.; Zhang, Q.; Subedi, Y.P.; Shrestha, J.P.; Kawasaki, Y.; Grilley, M.; Takemoto, J.Y.; Chang, C.-W.T. One-step synthesis of carbohydrate esters as antibacterial and antifungal agents. Bioorgan. Med. Chem. 2018, 26, 765–774. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds 9, 11, K20 and FG08 are available from the authors. |
Figure 1.
Structure of aminoglycosides and selected kanamycins (AKs).
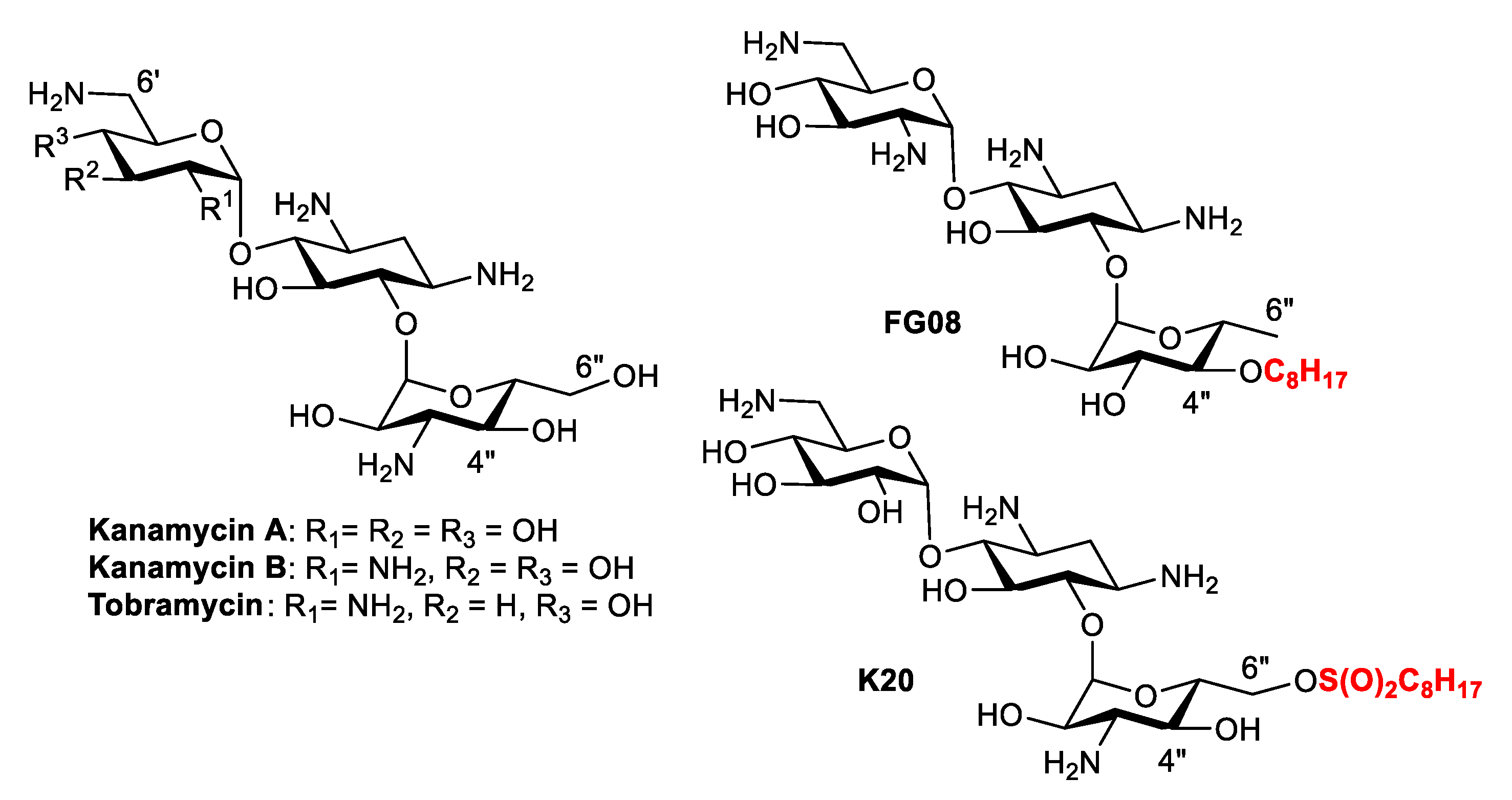
Figure 2.
Structures of SYTOXTM green and propidium iodide (PI).
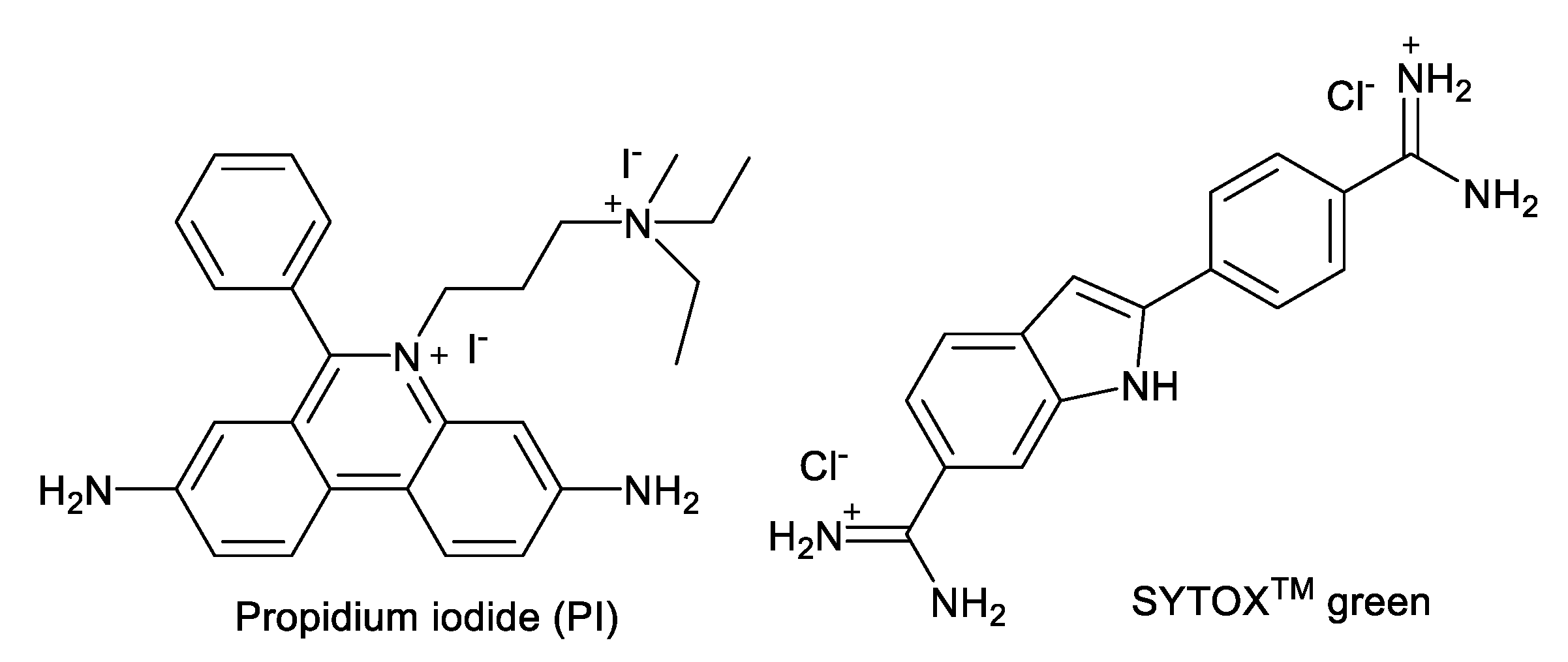
Figure 3.
Structure of selected AKs.

Figure 4.
Relationship of the water/1-octanol distribution coefficient (cLogD) vs. minimal growth inhibitory concentration (MIC).
Figure 4.
Relationship of the water/1-octanol distribution coefficient (cLogD) vs. minimal growth inhibitory concentration (MIC).
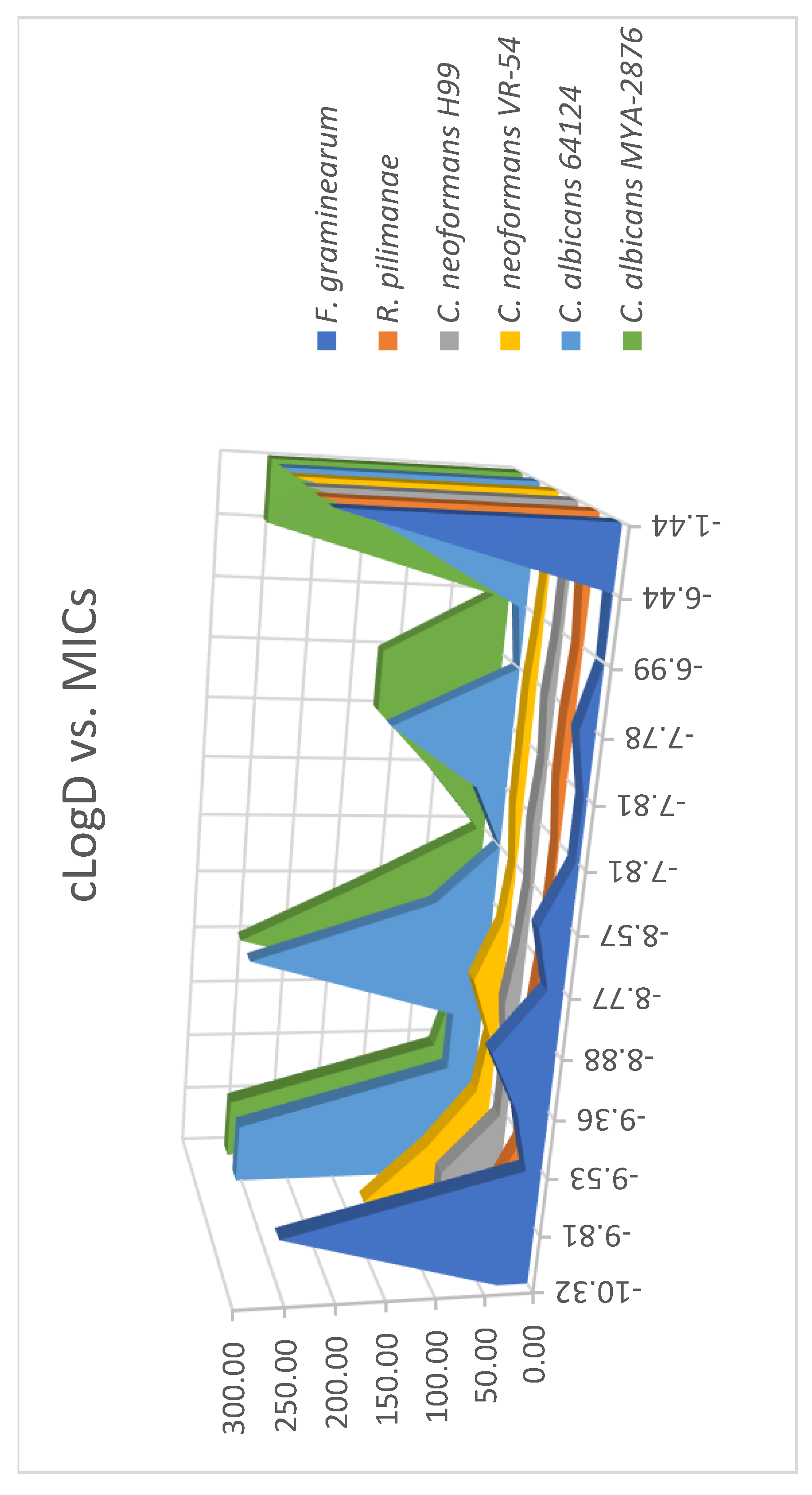
Figure 5.
Phase contrast (left panels ) and fluorescent (right panels) images of C. neoformans H99. (A) and (B): Images of cells treated with SYTOXTM green alone; (C) and (D): Images of cells treated with SYTOXTM green and 1% Triton X-100; (E) and (F): Images of cells treated with SYTOXTM green and compound 7 (1×MIC); (G) and (H): Images of cells treated with SYTOXTM green and compound 11 (1xMIC); (I) and (J): Images of cells treated with SYTOXTM green and compound 13 (256 μg/mL).
Figure 5.
Phase contrast (left panels ) and fluorescent (right panels) images of C. neoformans H99. (A) and (B): Images of cells treated with SYTOXTM green alone; (C) and (D): Images of cells treated with SYTOXTM green and 1% Triton X-100; (E) and (F): Images of cells treated with SYTOXTM green and compound 7 (1×MIC); (G) and (H): Images of cells treated with SYTOXTM green and compound 11 (1xMIC); (I) and (J): Images of cells treated with SYTOXTM green and compound 13 (256 μg/mL).

Figure 6.
Kinetic membrane permeabilization of C. neoformans H99. (A) Cells treated with 4″,6″-diaryl AKs using SYTOXTM green; (B) cells treated with 4″,6″-diaryl AKs using PI; (C) cells treated with 4″,6″-dialkyl AKs using SYTOXTM green; (D) cells treated with 4″,6″-dialkyl AKs using PI. The figure with standard deviation is available in Supplementary Materials.
Figure 6.
Kinetic membrane permeabilization of C. neoformans H99. (A) Cells treated with 4″,6″-diaryl AKs using SYTOXTM green; (B) cells treated with 4″,6″-diaryl AKs using PI; (C) cells treated with 4″,6″-dialkyl AKs using SYTOXTM green; (D) cells treated with 4″,6″-dialkyl AKs using PI. The figure with standard deviation is available in Supplementary Materials.
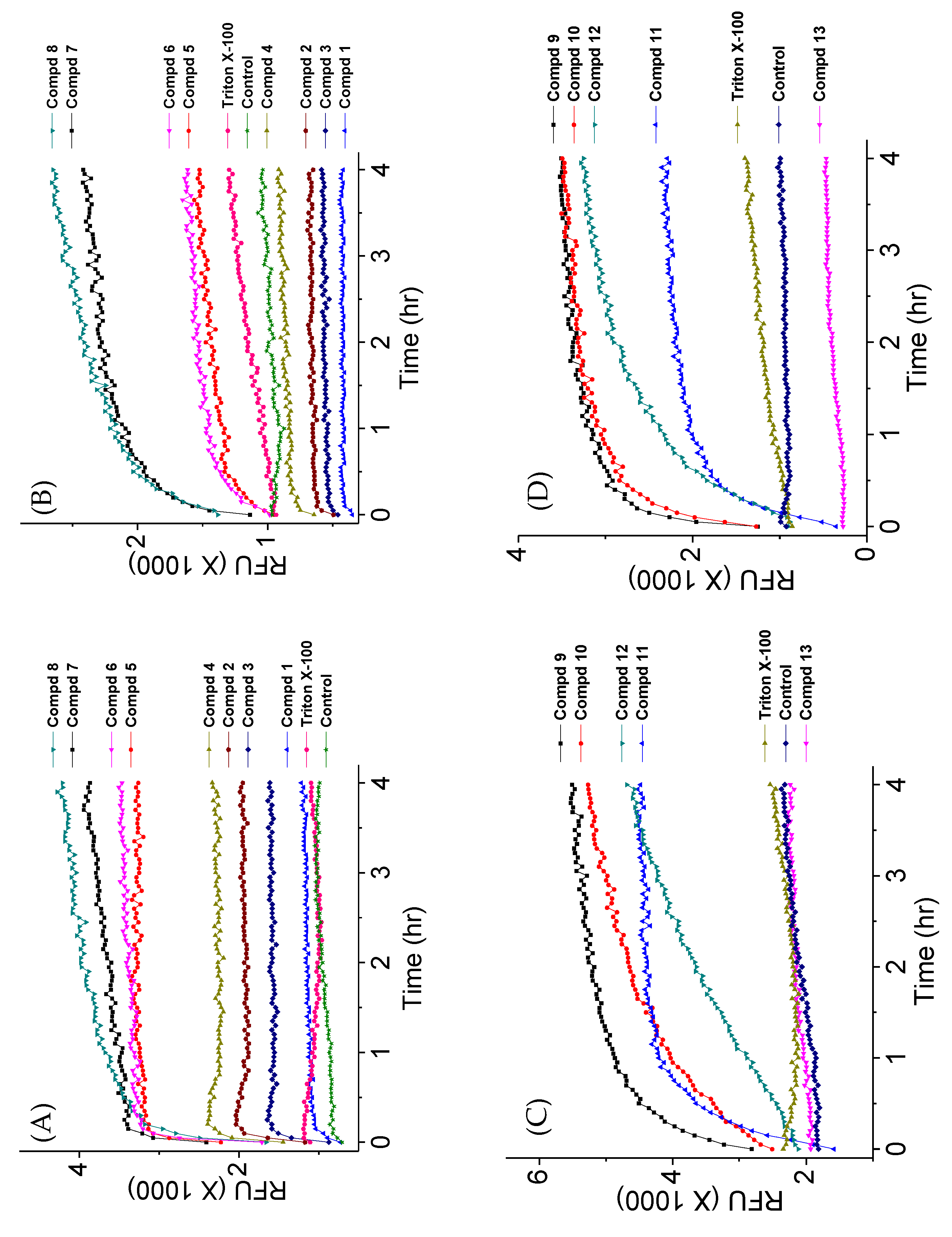
Figure 7.
Kinetic membrane permeabilization of C. neoformans H99 using varied concentrations of AKs. (A) Cells treated with compound 7 using SYTOXTM green; (B) cells treated with compound 7 using PI; (C cells treated with compound 10 using SYTOXTM green; (D): cells treated with compound 10 using PI; (E cells treated with compound 11 using SYTOXTM green; (F): cells treated with compound 11 using PI. The figure with standard deviation is available in Supplementary Materials.
Figure 7.
Kinetic membrane permeabilization of C. neoformans H99 using varied concentrations of AKs. (A) Cells treated with compound 7 using SYTOXTM green; (B) cells treated with compound 7 using PI; (C cells treated with compound 10 using SYTOXTM green; (D): cells treated with compound 10 using PI; (E cells treated with compound 11 using SYTOXTM green; (F): cells treated with compound 11 using PI. The figure with standard deviation is available in Supplementary Materials.

Figure 8.
Images of C. neoformans treated with varied concentrations of compound 11. (A1,A2) Cells treated with only SYTOXTM green; (B1,B2) cells treated with Triton X-100 and SYTOXTM green; (C1,C2) cells treated with 0.5×MIC of 11 and SYTOXTM green; (D1,D2) cells treated with 1×MIC of 11 and SYTOXTM green; (E1,E2) cells treated with 2×MIC of 11 and SYTOXTM green; (F1,F2) cells treated with 4×MIC of 11 and SYTOXTM green; (G1,G2) cells treated with 8×MIC of 11 and SYTOXTM green; (H1,H2) cells treated with only PI; (I1,I2) cells treated with Triton X-100 and PI; (J1,J2) cells treated with 0.5×MIC of 11 and PI; (K1,K2) cells treated with 1×MIC of 11 and PI; (L1,L2) cells treated with 2×MIC of 11 and PI; (M1,M2) cells treated with 4×MIC of 11 and PI; (N1,N2) cells treated with 8×MIC of 11 and PI.
Figure 8.
Images of C. neoformans treated with varied concentrations of compound 11. (A1,A2) Cells treated with only SYTOXTM green; (B1,B2) cells treated with Triton X-100 and SYTOXTM green; (C1,C2) cells treated with 0.5×MIC of 11 and SYTOXTM green; (D1,D2) cells treated with 1×MIC of 11 and SYTOXTM green; (E1,E2) cells treated with 2×MIC of 11 and SYTOXTM green; (F1,F2) cells treated with 4×MIC of 11 and SYTOXTM green; (G1,G2) cells treated with 8×MIC of 11 and SYTOXTM green; (H1,H2) cells treated with only PI; (I1,I2) cells treated with Triton X-100 and PI; (J1,J2) cells treated with 0.5×MIC of 11 and PI; (K1,K2) cells treated with 1×MIC of 11 and PI; (L1,L2) cells treated with 2×MIC of 11 and PI; (M1,M2) cells treated with 4×MIC of 11 and PI; (N1,N2) cells treated with 8×MIC of 11 and PI.

Figure 9.
Reactive oxygen species (ROS) study for 4″,6″-disubstituted AKs and compound 14 structure.
Figure 9.
Reactive oxygen species (ROS) study for 4″,6″-disubstituted AKs and compound 14 structure.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Minimal growth inhibitory concentrations (MICs) of AKs against fungal strains.
| Compound | cLogD | Strains | |||||
|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | ||
| 1 | −10.32 | 32–64 | 256 | 256 | 32 | 64 | 128 |
| 2 | −9.81 | 256 | 256 | >256 | 32 | 64 | 64 |
| 3 | −8.88 | 64–128 | >256 | 256 | 8 | 16 | 32 |
| 4 | −9.53 | 16–32 | 32–64 | 32–64 | 4 | 8 | 16 |
| 5 | −8.77 | 16–32 | 64 | 128 | 4–8 | 8 | 8 |
| 6 | −7.81 | 4–16 | 32–64 | 64 | 2–4 | 8 | 8 |
| 7 | −7.81 | 4−8 | 128 | 128 | 8–16 | 4 | 8 |
| 8 | −6.44 | 2–8 | 128–256 | 256 | 8–16 | 4 | 2–4 |
| 9 | −9.36 | 32 | 32 | 16–32 | 8–16 | 8 | 8 |
| 10 | −8.57 | 32 | ND | 8–16 | 2 | 4 | 2 |
| 11 | −7.78 | 16 | ND | 128 | 8–16 | 8 | 8–16 |
| 12 | −6.99 | ND | 8 | 16–32 | 4 | 4 | 4 |
| 13 | −1.44 | >256 | >256 | >256 | >256 | 256 | >256 |
| K20 | −11.62 | 32 | 16 | 32 | 8–16 | 8 | 16 |
MIC unit: µg/mL; ND: Not determined; strains: (A) Fusarium graminearum B4-5A, (B) Candida albicans 64124, (C) Candida albicans MYA-2876, (D) Rhodotorula pilimanae (ATCC 26423), (E) Cryptococcus neoformans H99, (F) Cryptococcus neoformans VR-54.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alfindee, M.N.; Subedi, Y.P.; Grilley, M.M.; Takemoto, J.Y.; Chang, C.-W.T. Antifungal Activities of 4″,6″-Disubstituted Amphiphilic Kanamycins. Molecules 2019, 24, 1882. https://doi.org/10.3390/molecules24101882
AMA Style
Alfindee MN, Subedi YP, Grilley MM, Takemoto JY, Chang C-WT. Antifungal Activities of 4″,6″-Disubstituted Amphiphilic Kanamycins. Molecules. 2019; 24(10):1882. https://doi.org/10.3390/molecules24101882
Chicago/Turabian StyleAlfindee, Madher N., Yagya P. Subedi, Michelle M. Grilley, Jon Y. Takemoto, and Cheng-Wei T. Chang. 2019. "Antifungal Activities of 4″,6″-Disubstituted Amphiphilic Kanamycins" Molecules 24, no. 10: 1882. https://doi.org/10.3390/molecules24101882