1. Introduction
Ischemic stroke is a clinically common cerebrovascular disease accounting for 70 to 80% of all cerebrovascular patients; approximately 15 million people worldwide suffer from stroke each year, resulting in approximately 5 million deaths and making it a primary cause of disability and death worldwide [
1,
2]. Thus far, the primary concern of clinical treatment for ischemic stroke is to restore blood and oxygen supply to the ischemic brain tissue as soon as possible. However, re-opening the occluded cerebral vessels often produces pathological damage in the ischemic tissue and causes cerebral ischemia and reperfusion injury to be aggravated further or potentially rendered irreversible [
2,
3]. To explore better treatment options for ischemic stroke and reperfusion injury, researchers have carried out extensive studies on its pathogenesis; proposed a set of theories [
4,
5] mainly including energy metabolism disorders, oxidative stress, glutamate toxicity, Ca
2+ overload, excessive NO synthesis, and apoptosis; and developing a series of new drugs [
5], such as recombinant tissue plasminogen activator (r-TPA), aspirin and heparin. At present, significant progress has been made to alleviate the tremendous public health problems relating to stroke, such as the preventive efforts to reduce the morbidity and mortality associated with strokes, in addition to the establishment of special intensive care units to improve the functional consequence of stroke patients. However, the management of acute ischemic stroke has not made significant strides since the introduction of recombinant tissue plasminogen activator (r-TPA) two decades ago [
3], and r-TPA is the only thrombolytic agent approved by the US Food and Drug Administration for stroke treatment [
6]. Since its therapeutic time window (TTW) is narrow, r-TPA treatment for stroke is suitable for only a limited percentage of stroke patients (<10%) [
7].
After cerebral ischemia, particularly following the cerebral ischemia and reperfusion process, various damage factors exist, including oxygen free radicals, oxidative stress, glutamate toxicity, Ca
2+ overload, and excessive NO synthesis. Furthermore, apoptosis is triggered through various pathways, such as hydroxyl radicals and superoxide anions. In the pathogenesis and treatment of acute ischemic stroke, an acute ischemic cascade reaction can greatly damage brain cells due to energy metabolism disorders, oxidative stress or inhibition of the body’s oxidative defence, easily leading to neuronal apoptosis, necrosis or loss of neurons in the hippocampus and cortex of the brain [
8,
9,
10,
11,
12,
13]. Therefore, research and development of neuroprotective drugs to reduce apoptosis and necrosis of brain tissue for the treatment of acute ischemic stroke is clinically imperative [
14].
Apoptosis, also called programmed cell death, is involved in the pathogenesis and pathological mechanism of ischemic stroke. Apoptosis is widely involved in the activation of cell development and physiological cell turnover but can also contribute to pathological damage by environmental stimulation, leading to disorders such as stroke [
12,
14,
15,
16,
17]. It has been shown that arsenic-induced cytotoxicity and oxidative stress damage could lead to apoptosis in a variety of cell types, including neuronal cells, myoblasts, and osteoblasts, which are closely related to activation of PI3K-Akt/mTOR, mitogen-activated protein kinase (MAPK) and endoplasmic reticulum stress regulatory signaling pathways [
3,
4,
8,
9,
10]. While the mechanisms of neuronal apoptosis caused by cerebral ischemia are not yet completely elucidated, much evidence has shown that neuronal apoptosis is closely associated with cell cycle regulation and oxidative stress damage. Furthermore, the MAPK signaling pathway, which includes ERK1/2, JNK1/2 and p-38, may inhibit the activation and hyperproliferation of glial cells by regulating the cell cycle and reducing harmful secreted factors that damage neurons [
8,
9,
10,
13]; on the other hand, this pathway reduces glial proliferation and swelling caused by local microcirculatory disorders, which creates a favorable environment for survival, not neuronal apoptosis [
18]. The MAPK signaling pathway has been implicated in the regulation of cytokine expression and cell apoptosis after stroke, and this pathway might represent a novel therapeutic target [
19,
20]. Thus, studies of MAPK activation in ischemic brain tissue may provide a foundation for the discovery of novel therapeutic agents for stroke patients.
In China, the use of traditional Chinese herbal medicines and combination preparations to treat cerebrovascular diseases can be traced back to the Han Dynasty. Even thousands of years ago, the medical formulary recorded many valuable Chinese medicines and classic prescriptions, such as melon, salvia, mulberry, panax notoginseng, and ganoderma lucidum, for the treatment of brain I/R-related diseases [
21]. Recently, several new compound traditional Chinese medicine preparations have been formulated for the treatment of cerebrovascular diseases in China [
15,
16,
21]. Radix Scrophulariae, called Xuan Shen in Chinese, is a traditional Chinese medicine derived from the dried roots of
Scrophularia ningpoensis Hemsl. Radix Scrophulariae has been used for thousands of years in China owing to its excellent traditional therapeutic effects and various pharmacological activities, such as anti-myocardial ischemia, anti-atherosclerosis, antimyocardial hypertrophy, anticerebral ischemia, antiplatelet aggregation, anti-inflammatory, liver protection, immune regulation, antibacteria, neuron protection, hypnosis, and antihyperuricaemia [
22,
23]. Modern pharmacological studies and clinical practice have demonstrated that Radix Scrophulariae possesses anti-angiogenesis, anti-inflammatory, and antimicrobial activities, as well as the ability to promote ventricular remodeling [
21,
22,
24]. Some studies have demonstrated that Radix Scrophulariae extract has anti-apoptotic and anti-inflammatory effects, potentially operating by affecting the mitogen-activated protein kinases (MAPKs) signaling pathway and inhibition of the NF-κB pathway [
24]. Although some pharmacological studies have been reported, the effects and mechanisms of Radix Scrophulariae activity against cerebral ischemia has not been clearly elaborated, and the chemical basis of this activity remains unclear.
Based on the above, we hypothesized that Radix Scrophulariae extract may provide neuroprotective effects on cerebral ischemia and reperfusion injury by inhibiting oxidative stress damage and inhibiting apoptosis by regulating the MAPK pathway. Therefore, we investigated the effects of Radix Scrophulariae aqueous extract (RSAE) on PC12 cells subjected to an oxygen-glucose deprivation and reperfusion (OGD/R) model and MCAO/R-operated mice in cerebral ischemic stroke. Recently, chemists have identified various chemical components within this extract, such as iridoids, phenylpropanoids, anthraquinones, phenols, phenylethanoid glycosides, sterols, flavonoids, fatty acids, and sugars; as showed in
Figure 1, a total of 41 compounds were identified from RSAE based on the accurate mass measurement of precursor and product ions via MS/MS-Q-TOF/MS as previously described [
22,
23].
3. Discussion
It has been reported that in ischemic stroke, energy metabolism disorders and mitochondrial dysfunction may lead to the formation of large amounts of free radicals, mediate oxidative damage to DNA, inhibit antioxidant enzyme activity, upregulate Bax levels, downregulate Bcl-2 levels in astrocytes, and finally, suppress the activation of Caspase-3, thus inducing neuronal apoptosis [
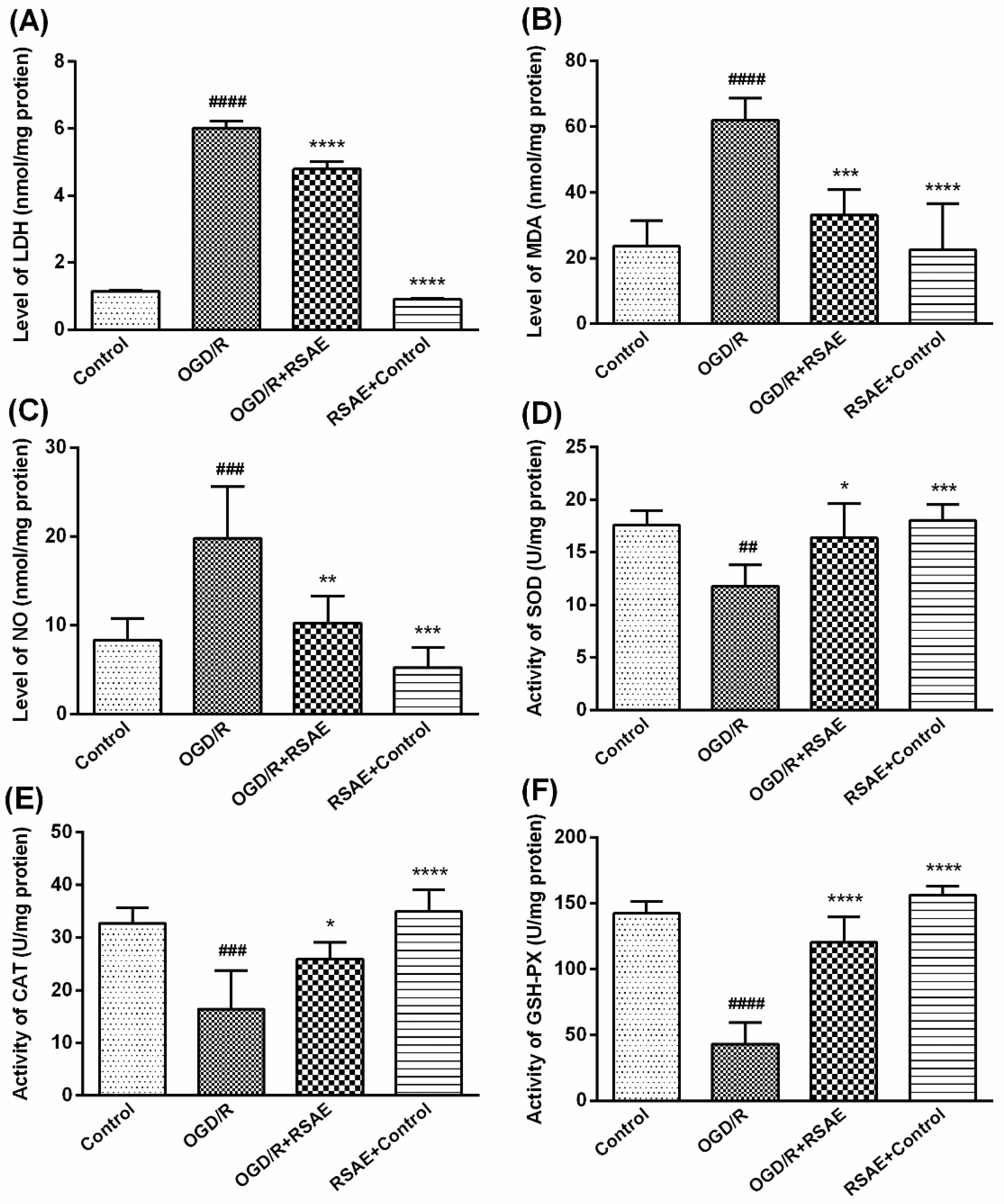
28]. Our research found that RSAE could increase the antioxidant capacity to mediate oxidative stress in OGD/R-induced PC12 cells and MCAO/R model mice: SOD, CAT and GSH-Px activity was remarkably improved (
Figure 3 and
Figure 5), indicating that ROS may be reduced as a result of inhibition of MCAO/R-induced ROS production, which leads to mitochondrial damage and induces mitochondrial apoptotic pathways. RSAE can reduce ROS accumulation caused by mitochondrial damage and energy metabolism disorders and inhibit neuronal cell injuries and apoptosis induced by oxidative stress (
Figure 2 and
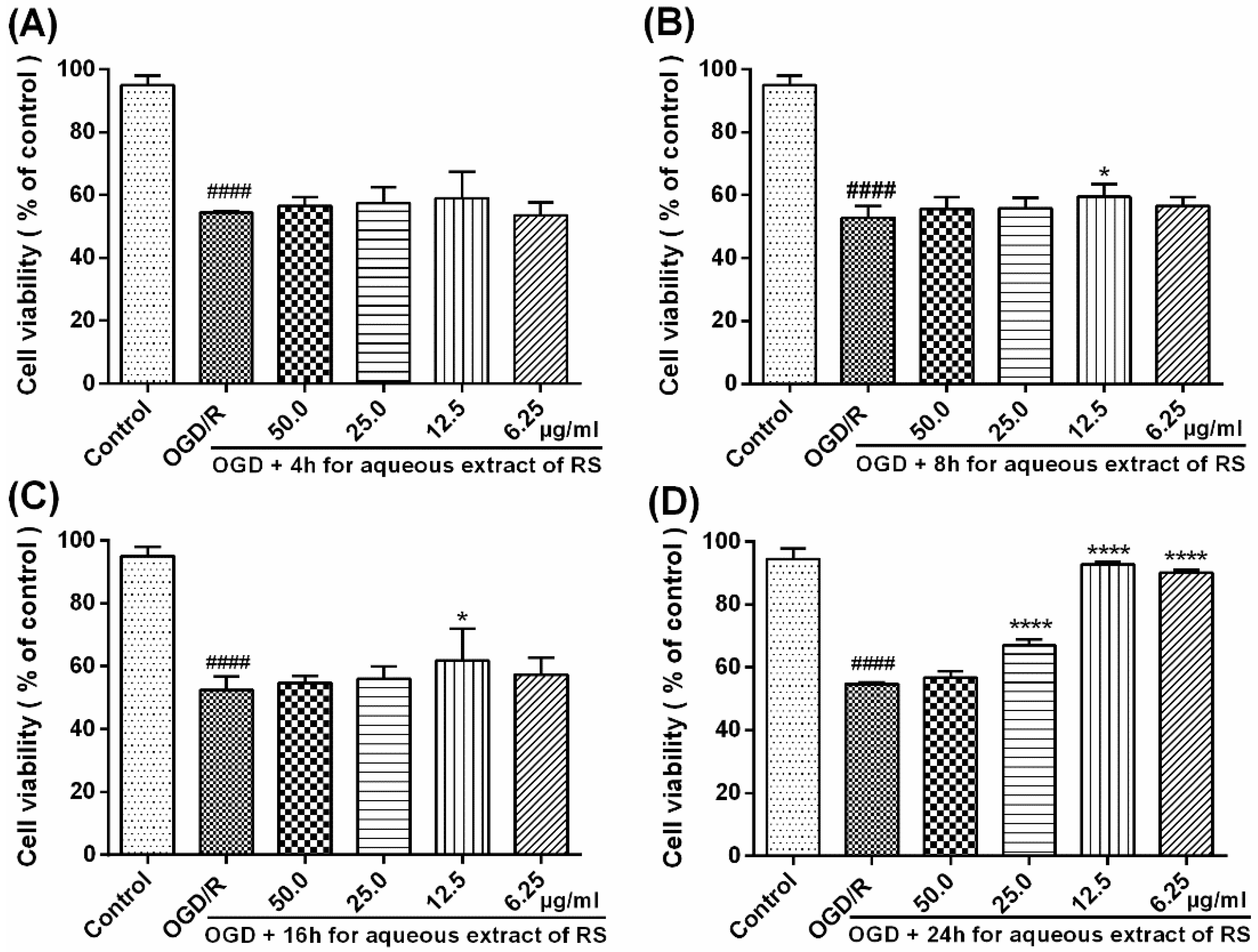
Figure 3). Therefore, our results showed that pretreatment with RSAE (12.5 μg/mL) for 8 h or 16 h significantly improved cell viability (
Figure 2) and inhibited OGD/R-induced damage (
Figure 3), indicating that RSAE may inhibit activated oxidative stress, reduce ROS production and decrease the mitochondrial membrane potential (MMP) (
Figure 4) to exert neuroprotective effects.
As is known, activation of apoptosis can be induced by growth factor deficiency, toxins, and glucose-oxygen deficiency. It has also been found that apoptosis can be controlled by a variety of regulators and regulatory proteins that possess inhibitory (anti-apoptosis) effects on programmed cell death or block (pro-apoptosis) inhibitors. To prevent target cells from activating apoptosis too soon, each cell encodes their own anti-apoptosis genes as a defensive strategy [
29]. Bcl-2 and Bax are two major members of the Bcl-2 family, a family of proteins that primarily regulates cell apoptosis and necrosis.
Bax is a cytosolic sensor of cell damage and stimulation that functions by forming pores across the mitochondrial outer membrane, leading to a decrease in the MMP along with an outflow of Cyt C and other apoptosis-inducing factors. Bax also activates Caspase-3 and destroys the anti-apoptotic function of the Bcl-2 protein under normal conditions [
30]. The anti-apoptotic protein Bcl-2 is an important intracellular component that inhibits the overexpression of Bax by blocking Cyt C release from mitochondria and Caspase-3 aggregation and that links the mitochondrial outer membrane to stabilize membrane permeability and protect mitochondrial integrity, which, in turn, inhibits apoptosis [
30]. Both Bax and Bcl-2 play an important regulatory role in the mitochondrial pathway; therefore, the ratio of Bax/Bcl-2 is considered an important indicator of apoptosis.
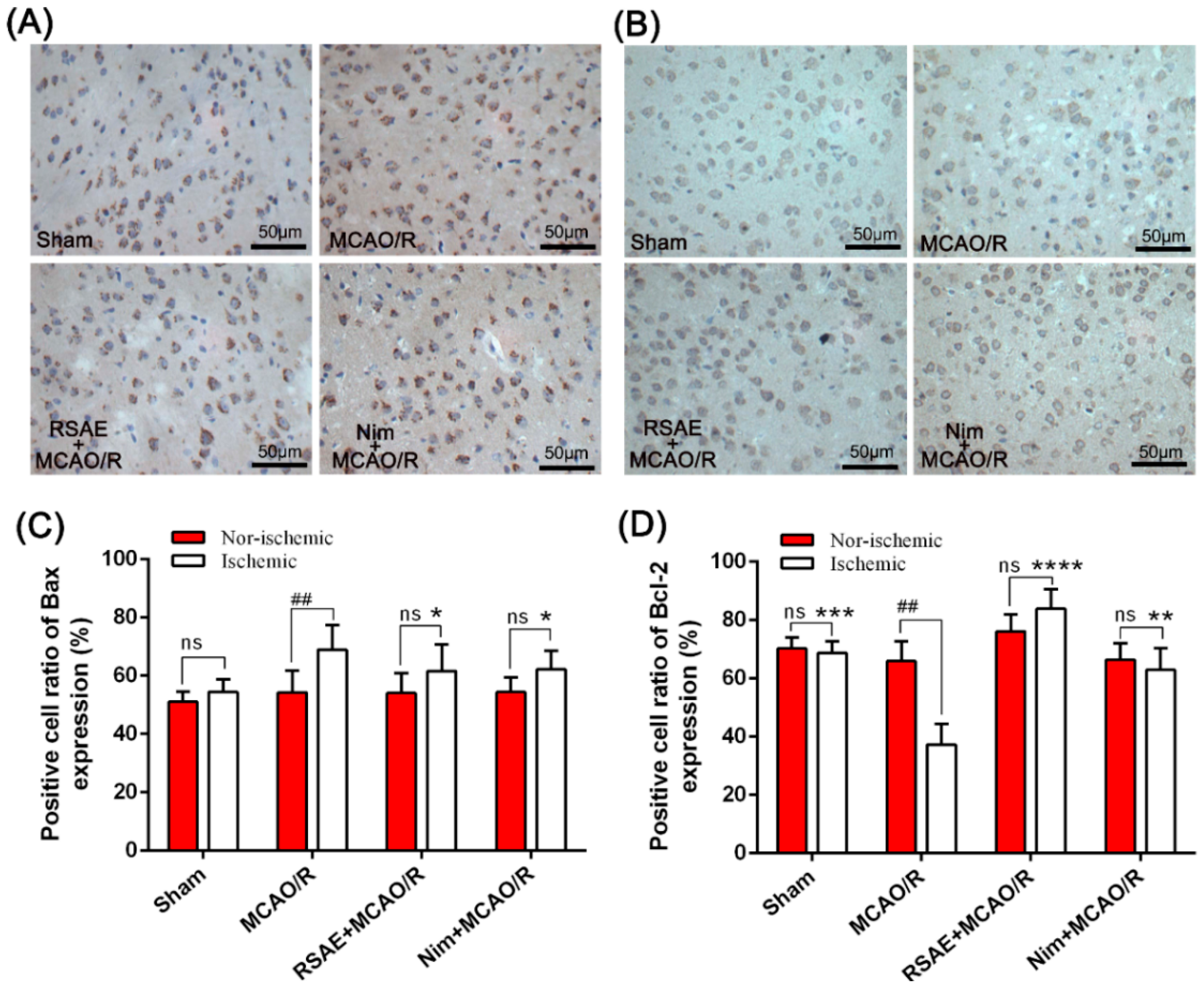
In this study, we used immunohistochemistry to detect changes in the expression of Bax and Bcl-2. It was suggested that the anti-apoptotic protection effects of RSAE treatment on ischemic stroke resulted from upregulating Bcl-2 protein expression and downregulating Bax protein expression (
Figure 4 and
Figure 6).
Thus, we speculated that since RSAE exerts an anti-apoptotic effect, it may inhibit the activation of Caspase-3 by regulating the expression of Bcl-2 and Bax. Concurrently, RSAE pretreatment increased antioxidant activity and stabilized the mitochondrial membrane potential, indicating that it may exert a protective effect by regulating oxidative stress and activation of apoptosis [
8,
9,
10,
13,
18,
27,
28].
Throughout the course of ischemic stroke, cerebral I/R following a cerebral infarction can cause neuronal apoptosis, reaching the summit 1 to 5 days after ischemia, and can last for approximately 4 weeks [
4,
14,
31,
32]. One study revealed that apoptosis was a dynamic progressive process that follows cerebral ischemia-reperfusion. During the early stage at 30 min, edema appeared in the anterior synovial striatum ischemic area surrounded by the appearance of a minor number of apoptotic cells, and with the prolongation of reperfusion time (6–72 h), the number of apoptotic cells increased. These cells were mainly located in the penumbra of the brain slices. The CA1 area of the hippocampus is considered to be sensitive to ischemic injury. After a certain time, ischemia-induced damage cannot be suppressed or corrected; consequently, the penumbra in the ischemic brain region may inevitably deteriorate and become part of a permanent infarct [
32]. In the central area of cerebral ischemia, the cells were mainly necrotic, with cell death primarily caused by apoptosis around the ischemic area [
8,
9,
10]. Similarly, our research showed that the number of positive cells in the ischemic (MCAO/R) group was significantly higher than that in the sham-operated group and that the number of neuron-positive cells was significantly decreased (
Figure 7). While the number of apoptosis-positive cells in the RSAE-treated group was significantly lower than that in the MCAO/R group, the number of positive cells increased. Furthermore, the infarct size of the intervention group was significantly smaller than that of the ischemic group, which indicated that RSAE seemed to exert neuroprotective effects on cerebral ischemia and reperfusion injury by inhibiting apoptosis.
Based on the present reports, the precise mechanism of neuronal death resulting from ischemic injury was not completely elucidated; this process is dependent on the severity of the injury and quantity of apoptotic cells, in addition to the mechanism of occurrence, location, extent, and duration of ischemia. Moreover, a timely and effective method of neuronal apoptosis inhibition during cerebral ischemia has not yet been fully resolved, but a large number of studies have shown that neuronal apoptosis is related to cell cycle regulation [
13,
18,
27,
28].
In our research, western blot analysis showed that MCAO/R significantly increased the phosphorylation levels of ERK1/2 and p38 MAPK proteins in the ischemic penumbra compared to those of controls. However, these increased phosphorylation levels of ERK1/2 and p38 induced by MCAO/R were strikingly abrogated by pretreatment with RSAE, which indicated that RSAE may modulate the ERK1/2 and p-38 MAPK pathway by mediating the phosphorylation levels of MAPK pathway components in MCAO/R mice. In addition, RSAE had no effect on p-JNK1/2, JNK1/2 and p38 expression (
Figure 8), indicating that RSAE may regulate cytokine expression and suppress cell apoptosis after stroke at least partially by modulating the MAPK pathway. However, it has not yet been determined whether RSAE binds directly to membrane receptors or regulates MAPKs through a secondary signal. Furthermore, the identification of specific active ingredients in RSAE has not yet been performed, necessitating further in-depth studies.
4. Methods
4.1. RSAE Preparation
Radix Scrophulariae was purchased from Xinyi Town, Chun’an Town, Zhejiang Province in China; the aqueous extract of Radix Scrophulariae (RSAE) was prepared by the Chinese Herbal Medicine Germplasm Resources and Evaluation Laboratory of Zhejiang University of Traditional Chinese Medicine, Hangzhou, in China. According to experimental requirements, RSAE was prepared and diluted to different concentrations.
4.2. Cell Cultures and OGD/R Models
PC12 (pheochromocytoma) cells were obtained from the Institute of Basic Medical Sciences at the Chinese Academy of Medical Sciences [
33] and were cultured at 37 °C in Dulbecco’s modified Eagle’s medium (Gibco, Grand Island, NY, USA) with 10% foetal bovine serum (Gibco, Grand Island, NY, USA), 100 U/mL penicillin and 100 mg/mL streptomycin in a normal incubator, which was used as complete medium (CM) containing 4.5 g/L D-glucose. PC12 cells at a density of 1 × 10
5 cells/mL were seeded in 96-well plates in a total volume of 100 μL per well, and the cells were allowed to adhere and grow for 24 h.
According to the oxygen-glucose deprivation and reperfusion (OGD/R) model established in our lab [
33,
34], we made proper adjustments, replaced growth medium with glucose-free DMEM (Gibco, U.S.), and placed the plates into an anaerobic incubator under 95% N
2 and 5% CO
2 at 37 °C for 120 min. Then, the glucose-free DMEM was rapidly replaced with CM so that cells were fed for a 24 h reperfusion period with complete medium in a normal incubator. Control cell cultures were incubated under normal conditions and were not deprived of oxygen or glucose. The cells were pretreated with different concentrations of RSAE, namely, 6.25 μg/mL, 12.5 μg/mL, 25.0 μg/mL and 50.0 μg/mL, and were then exposed to OGD/R injury.
4.3. Cell Viability Analysis
Cell viability of PC12 cells was assessed according to the MTT assay. After OGD/R, the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (Amresco, Atlanta, GA, USA) assay was used to detect cell viability as previously described [
33,
34,
35]. Briefly, MTT was dissolved in DMEM and added to each well for incubation at a final concentration of 0.5 mg/ml at 37 °C for 4 h. Then, to dissolve the formazan crystals, the medium was replaced with 150 μL of DMSO per well for 10 min. An automatic microplate reader (Spectrauor, TECAN, Sunrise, Austria) was used to record the optical density (OD) of all groups at 570 nm. Cell viability was expressed as a percentage of the control value. Each experiment was repeated in quintuplicate using three independent cultures.
4.4. Assessment of Cell Injury or Death
Cell viability analysis showed pretreatment with 12.5 μg/mL RSAE could improve the viability of OGD/R-induced PC12 cells. To confirm the reliability of the MTT results, we used diagnostic kits to measure the amount of LDH leakage release as an indicator of cell injury or death. The four sets of experiments were carried out as follows: (1) control PC12 cells, (2) OGD/R-induced PC12 cells, (3) PC12 cells pretreated with 12.5 μg/mL RSAE for 24 h and then induced by OGD/R, and (4) control PC12 cells pretreated with 12.5 μg/mL RSAE.
Based on the manufacturer’s instructions, the amount of LDH leakage release was determined using an assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Culture media were harvested after OGD/R injury to determine LDH levels. LDH leakage was expressed as a percentage of total LDH activity (LDH in the medium + LDH in the cells).
4.5. Determination of the Levels of Oxidative Stress
Following treatment, PC12 cells induced by OGD/R were lysed using 0.2% Triton X-100 for 30 min to release intracellular superoxide malondialdehyde (MDA), dismutase (SOD), glutathione peroxidase (GSH-Px) and catalase (CAT). The levels of oxidative stress were determined by quantifying MDA, SOD, GSH-Px and CAT enzyme activity using an automatic microplate reader (Spectrauor, TECAN, Sunrise, Austria) according to the kit instructions [
31]. Additionally, a BCA Protein Assay Kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) was used to detect protein concentrations according to the manufacturer’s instructions. The following four sets of experiments were performed: (1) control PC12 cells, (2) OGD/R-induced PC12 cells, (3) cells pretreated with 12.5 μg/mL RSAE for 24 h and then induced by OGD/R, and (4) control cells pretreated with 12.5 μg/mL RSAE.
4.6. Measurement of the Mitochondrial Membrane Potential
The change in the mitochondrial membrane potential (MMP) was detected by JC-1 staining [
36]. PC12 cells (cell density, 1 × 10
5 cells/mL) were cultured in 24-well plates. Posttreatment, cells were harvested and incubated with JC-1 (2 mM final concentration) at 37 °C in the dark for 30 min. Then, the cells were immediately observed using fluorescence microscopy (Leica, Germany Q9). The results were analysed using the ImageJ software (National Institutes of Health, Bethesda, MD, USA) [
33]. The following four sets of experiments were performed: (1) control PC12 cells, (2) OGD/R induced-PC12 cells, (3) cells pretreated with 12.5 μg/mL RSAE for 24 h and then induced by OGD/R, and (4) control cells pretreated with 12.5 μg/mL RSAE.
4.7. Mouse MCAO/R Model and Drug Administration
Male Kunming (KM) mice (SPF, weighed 18–22 g) used in this study were purchased from the Animal Experimental Center of Zhejiang University of Traditional Chinese Medicine. Mice were housed at an ambient temperature of 20 ± 1 °C with a 12-h light/dark cycle and free access to a standard laboratory chow diet and water. All human subjects gave informed consent for inclusion before participation in the study. The study was conducted in accordance with the Declaration of Helsinki. The protocol was approved by the Laboratory Animal Ethics Committee of the Institute of Medicinal Plant Development, Peking Union Medical College, and conformed to the Guide for the Care and Use of Laboratory Animals (Permit Number: SYXK 2017-0020).
For focal brain ischemia, the mouse middle cerebral artery occlusion and reperfusion model (MCAO/R) was used as described previously [
37,
38,
39,
40]. In brief, animals were anaesthetized with 50 mg·kg
−1 Zoletil 50 via intramuscular injection (Virbac S.A, Carros, France) and maintained at a half-dose. A silicone-coated 8–0 monofilament was introduced into the left internal carotid artery and advanced to occlude the middle cerebral artery for 2 h. After 2 h MCAO, the animals were briefly re-anaesthetized, and the filament was withdrawn for reperfusion studies.
All animals were divided into 4 groups (n = 10 per group) according to the random number table, namely, the MCAO/R operation model group; the sham operation group; the group treated with 2.4 g·kg−1 Radix Scrophulariae aqueous extract (RSAE), a dose equivalent to the original drug; and the positive drug group, with a dose of 12 mg·kg−1 nimodipine. Continuous gastric administration lasted for 7 days, once per day. The model and sham operation groups were given an equal volume of physiological saline for injection.
4.8. Quantification of Infarct Volume and Brain Water Content
After the MCAO/R operation, mice were anaesthetized with 50 mg·kg
−1 Zoletil 50 via intramuscular injection (Virbac S.A, Carros, France), and brains were removed, sectioned coronally at a thickness of 2 mm, incubated in a phosphate-buffered solution (pH = 7.3) with 2% (
w/
v) 2,3,5-triphenyltetrazolium chloride (TTC), and stained for 15 min at 37 °C, followed by overnight immersion [
41]. The normal nonischemic tissue was stained red, while the infarct tissue area remained unstained (white). The infarct areas on each slice were quantified by using ImageJ software. To compensate for the effect of brain edema, the corrected infarct volume was calculated as described previously (ten male mice per group): Corrected infarct area = left hemisphere area − (right hemisphere area-infarct area) [
42].
The mice were anaesthetized, and mouse brain tissue was acquired and processed as needed at 24 h after MCAO. A 3-mm section of the ischemic hemisphere brain was cut from the anterior pole to detect the water content in the brain tissue. The wet-dry method was applied to determine brain water content (BWC) in another subgroup (n = 10 per group). An electronic scale was used to weigh the ischemic and nonischemic hemispheres (wet weight). After the ischemic brain hemisphere was dried overnight at 105 °C in a desiccating oven, it was weighed again (dry weight), and the total brain water content was calculated according to BWC % = [(wet weight − dry weight)/wet weight] × 100% [
43,
44].
4.9. Assessment of the Levels of Superoxide Dismutase Activity and Malondialdehyde and Nitric Oxide Contents
At 24 h post-surgery, animals were sacrificed and decapitated, and the right hemisphere of the brain was retrieved. The remaining brain tissue was washed with prechilled saline before blood was removed, and the tissue was then dried on filter paper. After being weighed, tissues were prepared as 10% brain tissue homogenate in a homogenizer and centrifuged at 3000 rpm·min
−1 for 10 min. The supernatant was collected and stored at −20 °C. The Bradford method was used to determine protein concentrations in the collected supernatants. SOD activity was determined by xanthine oxidase, the MDA content was determined by thiobarbituric acid, and the nitric oxide (NO) content was determined by nitrate reductase. All the determinations were carried out using assay kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) [
31]. Ten male mice were used for each group.
4.10. Haematoxylin-Eosin and Nissl Staining
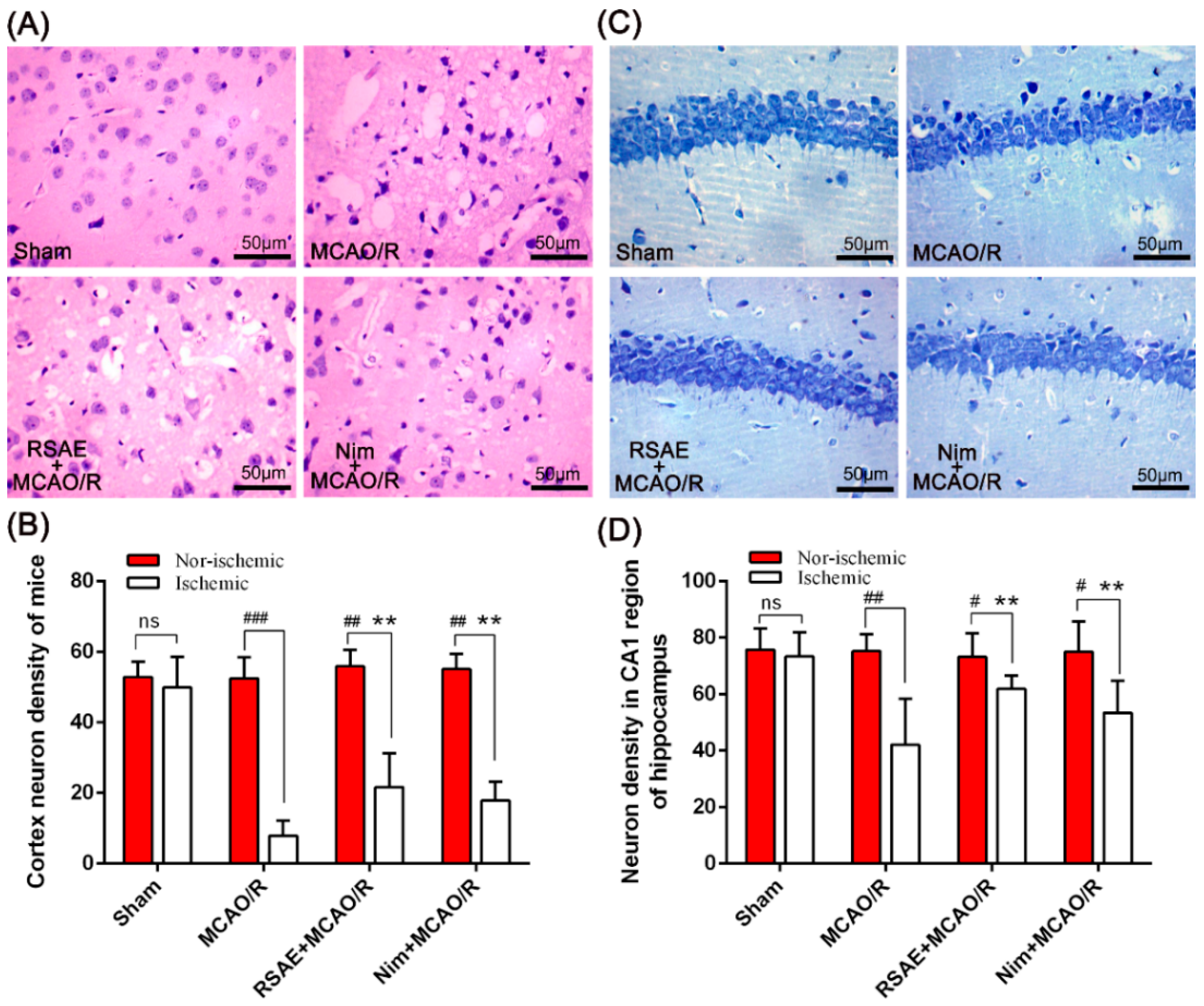
Following the MCAO/R operation, animals were anaesthetized with Zoletil 50 and immediately sacrificed. Then, the animals were perfused through the heart with 4°C phosphate-buffered saline (PBS), followed by 4% paraformaldehyde. Finally, the intact brain was carefully removed and placed in brain buffer containing 4% formaldehyde fixative for haematoxylin and eosin staining (H&E) staining, toluidine blue staining and other immunohistochemical staining.
Haematoxylin-eosin (H&E) staining was used to revealed morphological features of injured neurons in the cerebral cortex. The samples were embedded in paraffin and cut in 5-μm slices; 5-μm-thick serial coronal sections were generated and mounted on slides. The sections were stained using haematoxylin and H&E according to the described standard protocol [
39,
45]. Images of stained slides were acquired using a light microscope (Leica, Wetzlar, Germany). Nissl staining was applied to observe morphologic changes in hippocampus cells within the ischemic penumbra after MCAO/R operation, mainly containing CA1 in the hippocampus [
44]. After 24 h following MCAO/R operation, the brains were processed as above. Samples were embedded in paraffin and cut in 5-μm slices; 5-μm-thick serial coronal sections were generated and mounted on slides. After the sliced sections were washed in cold water, the paraffin sections were stained with 1% toluidine blue for 20 min, rinsed with PBS, dehydrated with graded alcohol, made transparent with xylene, and fixed with neutral glue. An optical microscope was used to observe each section.
Normal neurons showed typical cell size, prominent protrusions, uniform cytoplasm, a clear nuclear membrane, obvious nucleoli, and no chromatin condensation. Three high-magnitude (10 × 40) fields in both the cortex and the CA1 of the hippocampus were randomly selected for image analysis; the number of pyramidal cells was also counted. The mean value was taken as the number of pyramidal cells in the sample (n = 10 per group).
4.11. Immunohistochemical Evaluation
After MCAO/R operation, the animals were anaesthetized with Zoletil 50 and immediately sacrificed. The tissue samples obtained from mice were histologically processed and embedded in paraffin for immunohistochemical staining. Five micro slides were removed from these blocks and mounted on poly-L-lysine coated slides; these slides were immunohistochemically stained with primary antibodies recognizing Bcl-2 and Bax. The remaining procedure was performed as described previously with little modification [
12,
16]. Finally, the slides were fixed with a water-based mounting medium and evaluated under an optical microscope. Three sections of ischemic brain tissue with 10 × 40 magnification (400×) were randomly selected for image analysis via ImageJ software.
4.12. Western Blot Analysis
Twenty-four hours after MCAO, ischemic brain sections that included both the ischemic area and corresponding regions from the contralateral hemisphere were collected, and protein extracts were prepared as described previously [
33,
36]. PVDF membranes (Millipore, Bedford, MA, USA) were blocked for 2 h in 5% nonfat milk in Tris-buffered saline (TBS)/Tween 20 and probed with the following antibodies: anti-Bax (1:200), anti-Bcl-2 (1:200), anti-p-p38 (1:500), anti-p-38 (1:500), anti-p-ERK (1:200), anti-ERK (1:200), anti-cleaved caspase-3 (1:500), anti-p-JNK (1:200), anti-JNK (1:200), and anti-β-actin (1:1000). Protein expression was detected by an enhanced chemiluminescence method and imaged by using ChemiDoc XRS (Bio-Rad, Hercules, CA, USA). To reduce variations in protein expression quantification, three independent experiments were performed.
4.13. Statistical Analysis
Data are presented as the mean values ± standard error of the mean. All analyses were performed using GraphPad Prism 7.0 statistical software (GraphPad Software, Inc., La Jolla, San Diego, CA, USA). One-way ANOVA was used to determine the differences between groups, and then post-hoc LSD testing was applied. Comparisons between two groups were performed by using unpaired Student’s t-test. The p values < 0.05 were considered significant. At least three independent experiments were performed for each experiment.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








