Silicon Dioxide Nanoparticles Enhance Endotoxin-Induced Lung Injury in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Physicochemical Characterization of SiONPs
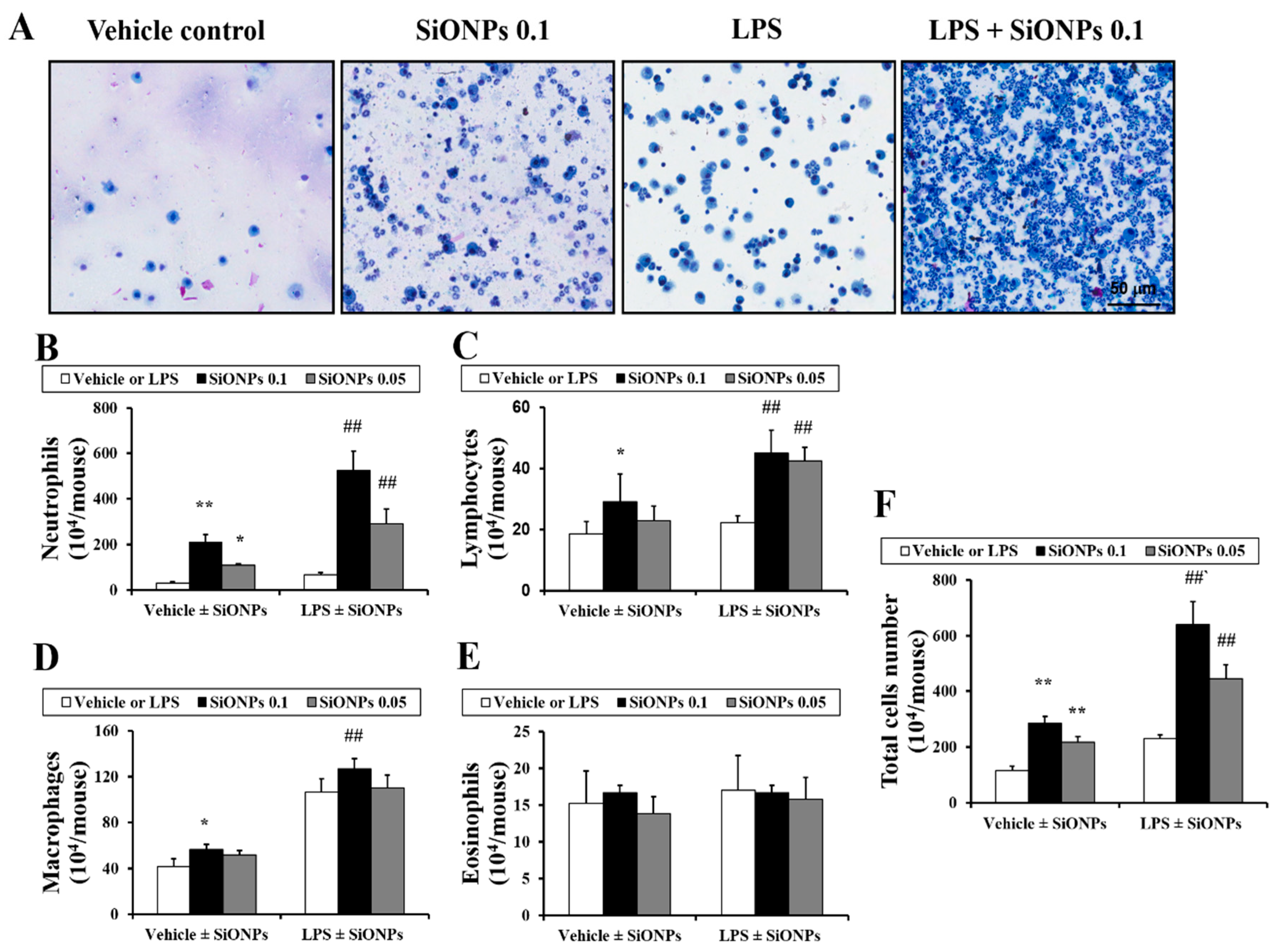
2.2. Effects of SiONPs on Inflammatory Cell Counts in the bronchoalveolar lavage fluid (BALF) from Vehicle- and LPS-Treated Mice
2.3. Effects of SiONPs on Inflammatory Cytokines in the BALF from the Vehicle- and LPS Treated-Mice
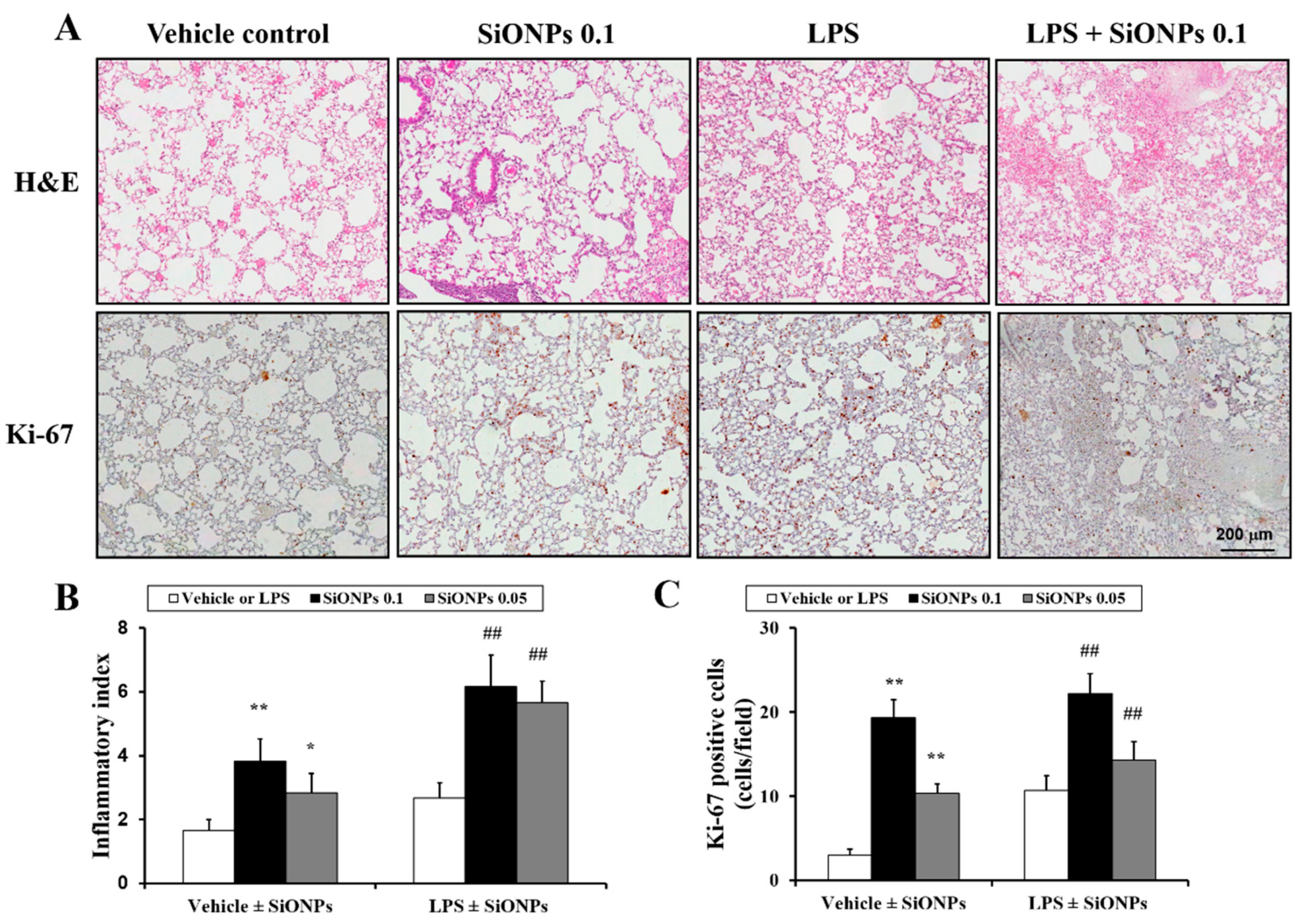
2.4. Effects of SiONPs on Histological Alteration in the Vehicle- and LPS-Treated Mice
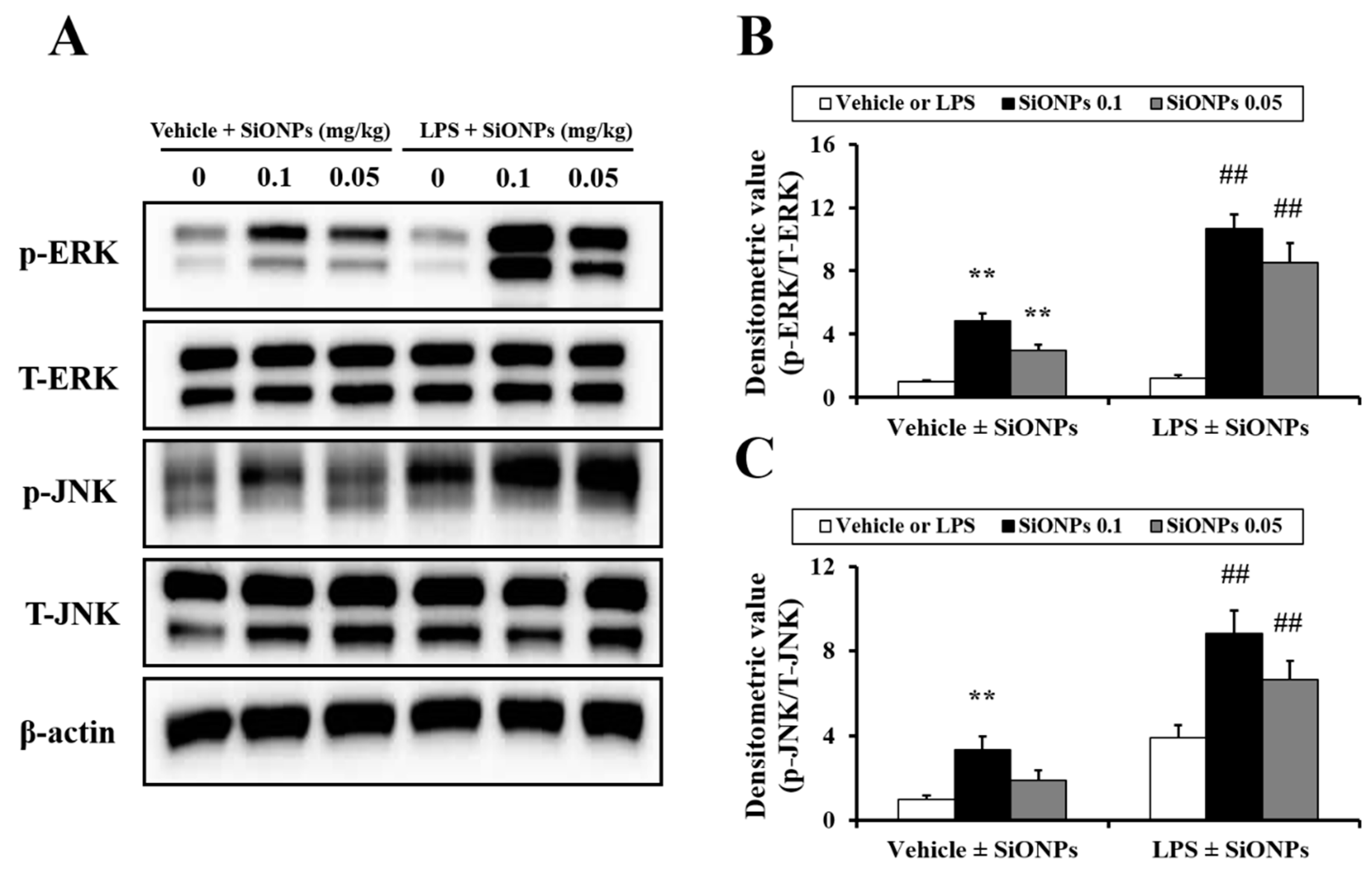
2.5. Effects of SiO2 on MAPK Expression in LPS-Treated Mice
3. Discussion
4. Materials and Methods
4.1. Nanoparticles
4.2. Experimental Procedure for the Animal Experiments
4.3. Measurement of Proinflammatory Cytokines in the BALF
4.4. Histology
4.5. Measurement of Mitogen-Activated Protein Kinase (MAPK) Protein Expression in Lung Tissue
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Ko, J.W.; Park, J.W.; Shin, N.R.; Kim, J.H.; Cho, Y.K.; Shin, D.H.; Kim, J.C.; Lee, I.C.; Oh, S.R.; Ahn, K.S.; et al. Copper oxide nanoparticle induces inflammatory response and mucus production via MAPK signaling in human bronchial epithelial cells. Environ. Toxicol. Pharmacol. 2016, 43, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Na, H.K.; Kim, Y.K.; Ryoo, S.R.; Cho, H.S.; Lee, K.E.; Jeon, H.; Ryoo, R.; Min, D.H.; Kim, M.H. Facile synthesis of monodispersed mesoporous silica nanoparticles with ultralarge pores and their application in gene delivery. ASC Nano 2011, 5, 3568–3576. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Hua, X.; Wu, Y.; Pan, X.; Liu, S. Polymer-functionalized silica nanosphere labels for ultrasensitive detection of tumor necrosis factor-alpha. Anal. Chem. 2011, 83, 6800–6809. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S.M.; Hunter, A.C.; Murray, J.C. Nanomedicine: Current status and future prospects. FASEB J. 2005, 19, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, J.; Ahamed, M.; Akhtar, M.J.; Alrokayan, S.A.; Siddiqui, M.A.; Musarrat, J.; Al-Khedhairy, A.A. Apoptosis induction by silica nanoparticles mediated through reactive oxygen species in human liver cell line HepG2. Toxicol. Appl. Pharmacol. 2012, 259, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Yu, Y.; Li, Y.; Yu, Y.; Li, Y.; Zhou, X.; Huang, P.; Sun, Z. Toxic effect of silica nanoparticles on endothelial cells through DNA damage response via Chk1-dependent G2/M checkpoint. PLoS ONE 2013, 8, e62087. [Google Scholar] [CrossRef] [PubMed]
- Hassankhani, R.; Esmaeillou, M.; Tehrani, A.A.; Nasirzadeh, K.; Khadir, F.; Maadi, H. In vivo toxicity of orally administrated silicon dioxide nanoparticles in healthy adult mice. Eviron. Sci. Pollut. Res. Int. 2015, 22, 1127–1132. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Muralikrishnan, S.; Ng, C.T.; Yung, L.Y.; Bay, B.H. Nanoparticle-induced pulmonary toxicity. Exp. Biol. Med. 2010, 235, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Naota, M.; Shiotsu, S.; Shimada, A.; Kohara, Y.; Morita, T.; Inoue, K.; Takano, H. Pathological study of chronic pulmonary toxicity induced by intratracheally instilled Asian sand dust (kosa). Toxicol. Pathol. 2013, 41, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Kashima, S.; Yorifuji, T.; Suzuki, E. Asian dust and daily emergency ambulance calls among elderly people in Japan: An analysis of its double role as a direct cause and as an effect modifier. J. Occup. Environ. Med. 2014, 56, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J. What are people dying of on high air pollution days? Environ. Res. 1994, 64, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.G.; Jung, J.H.; Kim, S.T. Asian sand dust enhances allergen-induced th2 allergic inflammatory changes and mucin production in BALB/c mouse lungs. Allergy Asthma Immunol. Res. 2012, 4, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Nafisi, S.; Schäfer-Korting, M.; Maibach, H.I. Perspectives on percutaneous penetration: Silica nanoparticles. Nanotoxicology 2015, 9, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Shimada, A.; Kohara, Y.; Naota, M.; Kobayashi, Y.; Morita, T.; Inoue, K.; Takano, H. Pathological study of chronic pulmonary toxicity induced by intratracheally instilled Asian sand dust (Kosa): Possible association of fibrosis with the development of granulomatous lesions. Folia Histochem. Cytobiol. 2015, 53, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Song, H.H.; Shin, I.S.; Woo, S.Y.; Lee, S.U.; Sung, M.H.; Ryu, H.W.; Kim, D.Y.; Ahn, K.S.; Lee, H.K.; Lee, D.; et al. Piscroside, C, a novel iridoid glycoside isolated from Pseudolysimachion rotundum var. subinegrum suppresses airway inflammation induced by cigarette smoke. J. Ethnopharmacol. 2015, 170, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Ko, J.W.; Shin, N.R.; Shin, D.H.; Cho, Y.K.; Seo, C.S.; Kim, J.C.; Kim, J.S.; Shin, I.S. 4-Hydroxycinnamic acid protects mice from cigarette smoke-induced pulmonary inflammation via MAPK pathways. Food. Chem. Toxicol. 2017, 110, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Sun, J.; Zhong, G.; Shi, L.; Zhang, D. Biodistribution and toxicity of intravenously administered silica nanoparticles in mice. Arch. Toxicol. 2010, 84, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Park, Y.H.; Park, H.J.; Lee, K.; Um, K.; Park, J.W.; Lee, J.H. Toxic and adjuvant effects of silica nanoparticles on ovalbumin-induced allergic airway inflammation in mice. Respir. Res. 2016, 17, 60. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Lee, I.C.; Shin, N.R.; Jeon, C.M.; Kwon, O.K.; Ko, J.W.; Kim, J.C.; Oh, S.R.; Shin, I.S.; Ahn, K.S. Copper oxide nanoparticles aggravate airway inflammation and mucus production in asthmatic mice via MAPK signaling. Nanotoxicology 2016, 10, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Grommes, J.; Mörgelin, M.; Soehnlein, O. Pioglitazone attenuates endotoxin-induced acute lung injury by reducing neutrophil recruitment. Eur. Respir. J. 2012, 40, 416–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.C.; Jivan, A.; Shao, C.; Duan, L.; Goad, D.; Zaganjor, E.; Osborne, J.; McGlynn, K.; Stippec, S.; Earnest, S.; et al. The roles of MAPKs in disease. Cell Res. 2008, 18, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, I.S.; Park, J.W.; Shin, N.R.; Jeon, C.M.; Kwon, O.K.; Lee, M.Y.; Kim, H.S.; Kim, J.C.; Oh, S.R.; Ahn, K.S. Melatonin inhibits MUC5AC production via suppression of MAPK signaling in human airway epithelial cells. J. Pineal. Res. 2014, 56, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Yang, M.; Jing, L.; Wang, J.; Yu, Y.; Li, Y.; Duan, J.; Zhou, X.; Li, Y.; Sun, Z. Amorphous silica nanoparticles trigger vascular endothelial cell injury through apoptosis and autophagy via reactive oxygen species-mediated MAPK/Bcl-2 and PI3K/Akt/mTOR signaling. Int. J. Nanomed. 2016, 11, 5257–5276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, J.; Liu, K.; Niu, W.; Chen, M.; Wang, M.; Xue, Y.; Gao, C.; Ma, P.X.; Lei, B. Gold and gold-silver alloy nanoparticles enhance the myogenic differentiation of myoblasts through p38 MAPK signaling pathway and promote in vivo skeletal muscle regeneration. Biomaterials 2018, 175, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Parnsamut, C.; Brimson, S. Effects of silver nanoparticles and gold nanoparticles on IL-2, IL-6, and TNF-α production via MAPK pathway in leukemic cell lines. Genet. Mol. Res. 2015, 14, 3650–3668. [Google Scholar] [CrossRef] [PubMed]
- Christen, V.; Fent, K. Silica nanoparticles induce endoplasmic reticulum stress response and activate mitogen activated kinase (MAPK) signalling. Toxicol. Rep. 2016, 3, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Ryu, H.W.; Ko, J.W.; Park, J.W.; Kwon, O.K.; Oh, S.R.; Kim, J.C.; Shin, I.S.; Ahn, K.S. A standardized bark extract of Pinus pinaster Aiton (Pycnogenol®) attenuated chronic obstructive pulmonary disease via Erk-sp1 signaling pathway. J. Ethnopharmacol. 2016, 194, 412–420. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds including SiONPs are available from the authors. |



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, J.-W.; Lee, H.-J.; Shin, N.-R.; Seo, Y.-S.; Kim, S.-H.; Shin, I.-S.; Kim, J.-S. Silicon Dioxide Nanoparticles Enhance Endotoxin-Induced Lung Injury in Mice. Molecules 2018, 23, 2247. https://doi.org/10.3390/molecules23092247
Ko J-W, Lee H-J, Shin N-R, Seo Y-S, Kim S-H, Shin I-S, Kim J-S. Silicon Dioxide Nanoparticles Enhance Endotoxin-Induced Lung Injury in Mice. Molecules. 2018; 23(9):2247. https://doi.org/10.3390/molecules23092247
Chicago/Turabian StyleKo, Je-Won, Hae-Jun Lee, Na-Rae Shin, Yun-Soo Seo, Sung-Ho Kim, In-Sik Shin, and Joong-Sun Kim. 2018. "Silicon Dioxide Nanoparticles Enhance Endotoxin-Induced Lung Injury in Mice" Molecules 23, no. 9: 2247. https://doi.org/10.3390/molecules23092247
APA StyleKo, J.-W., Lee, H.-J., Shin, N.-R., Seo, Y.-S., Kim, S.-H., Shin, I.-S., & Kim, J.-S. (2018). Silicon Dioxide Nanoparticles Enhance Endotoxin-Induced Lung Injury in Mice. Molecules, 23(9), 2247. https://doi.org/10.3390/molecules23092247